
The Novel RXR Agonist MSU-42011 Differentially Regulates Gene Expression in Mammary Tumors of MMTV-Neu Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
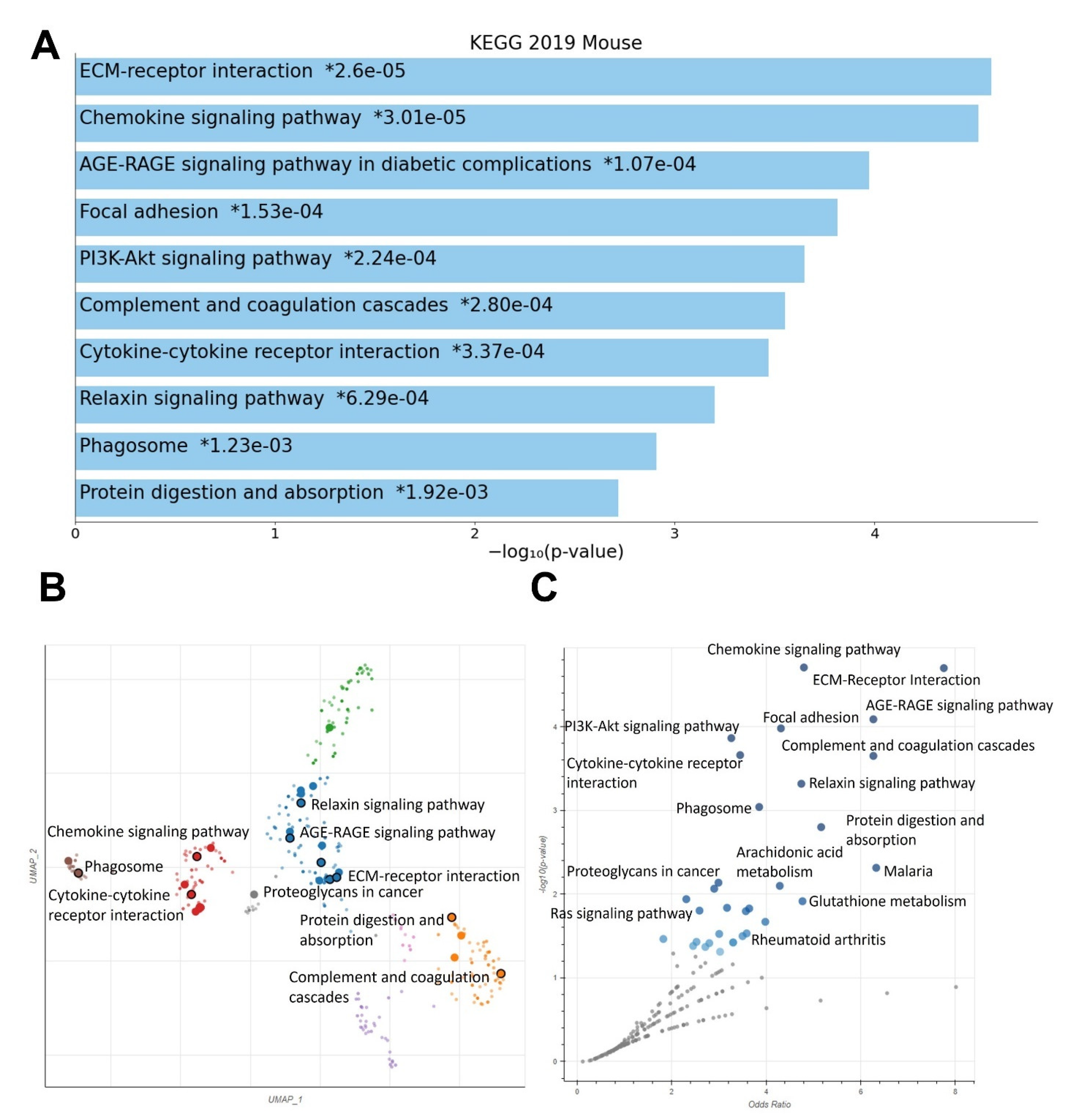
2.1. RXR Agonists Regulate Pathways Relevant in Breast Cancer
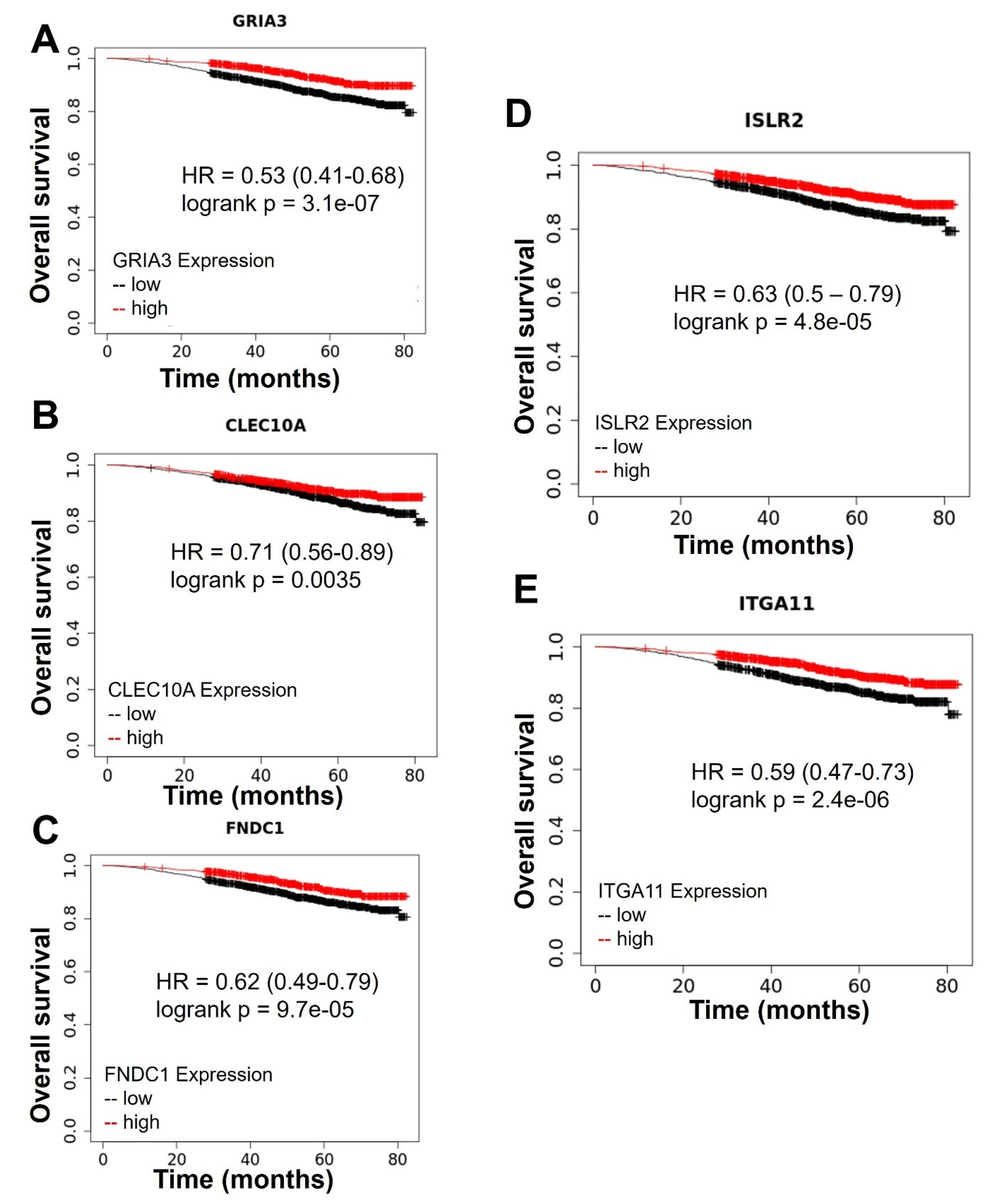
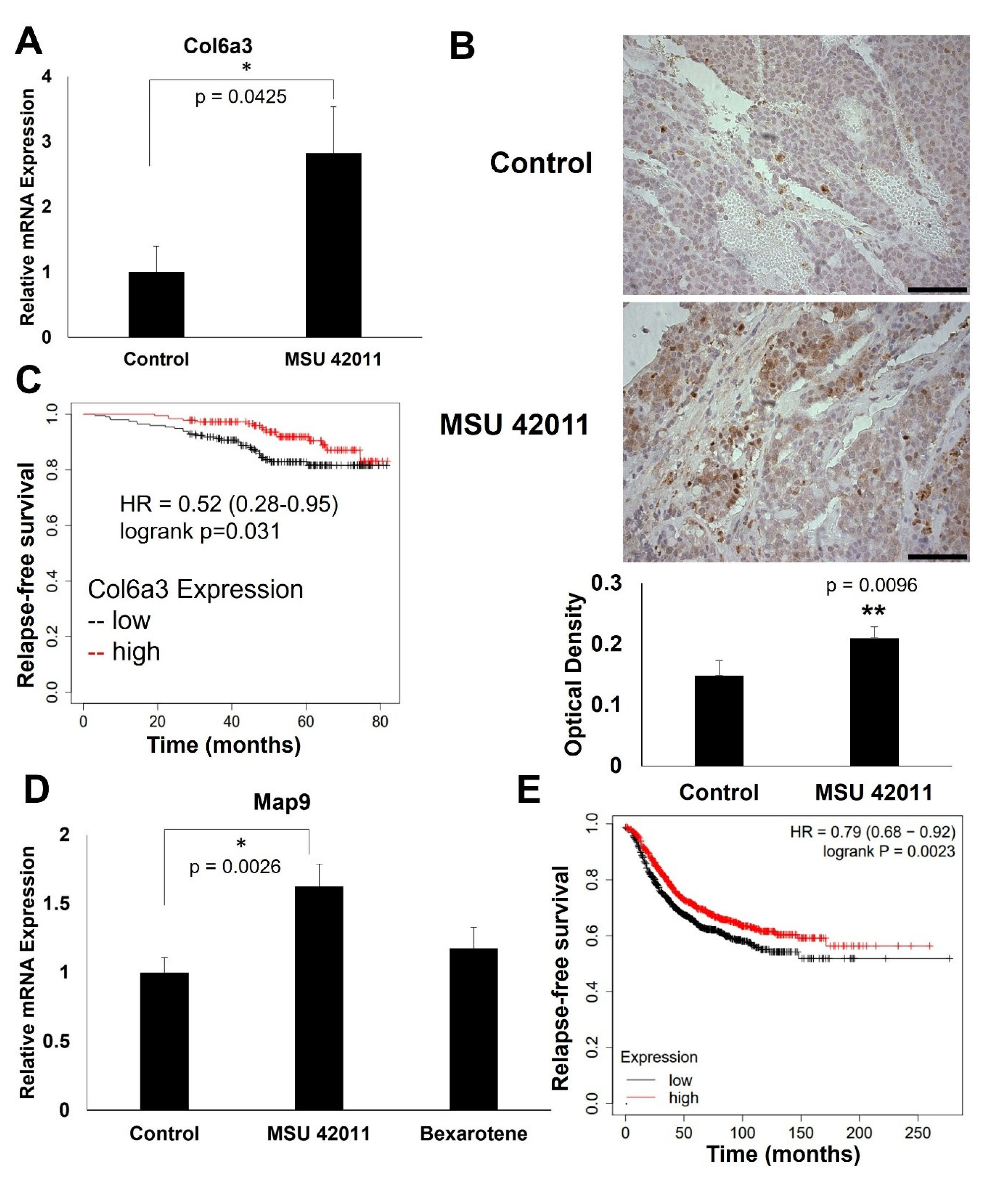
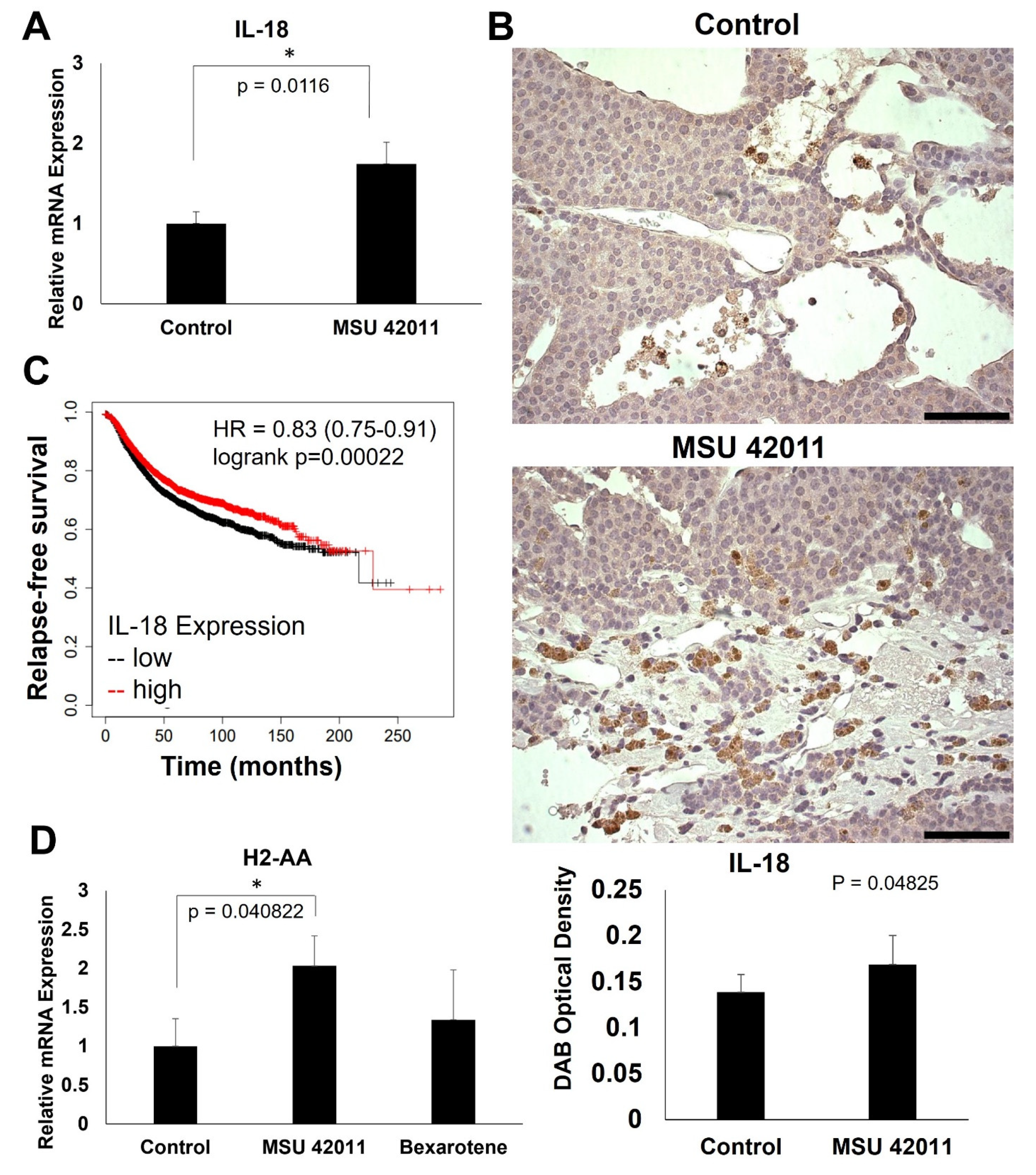
2.2. Top Genes Differentially Expressed in Tumors Treated with MSU-42011 and Bexarotene Correlate with Patient Survival
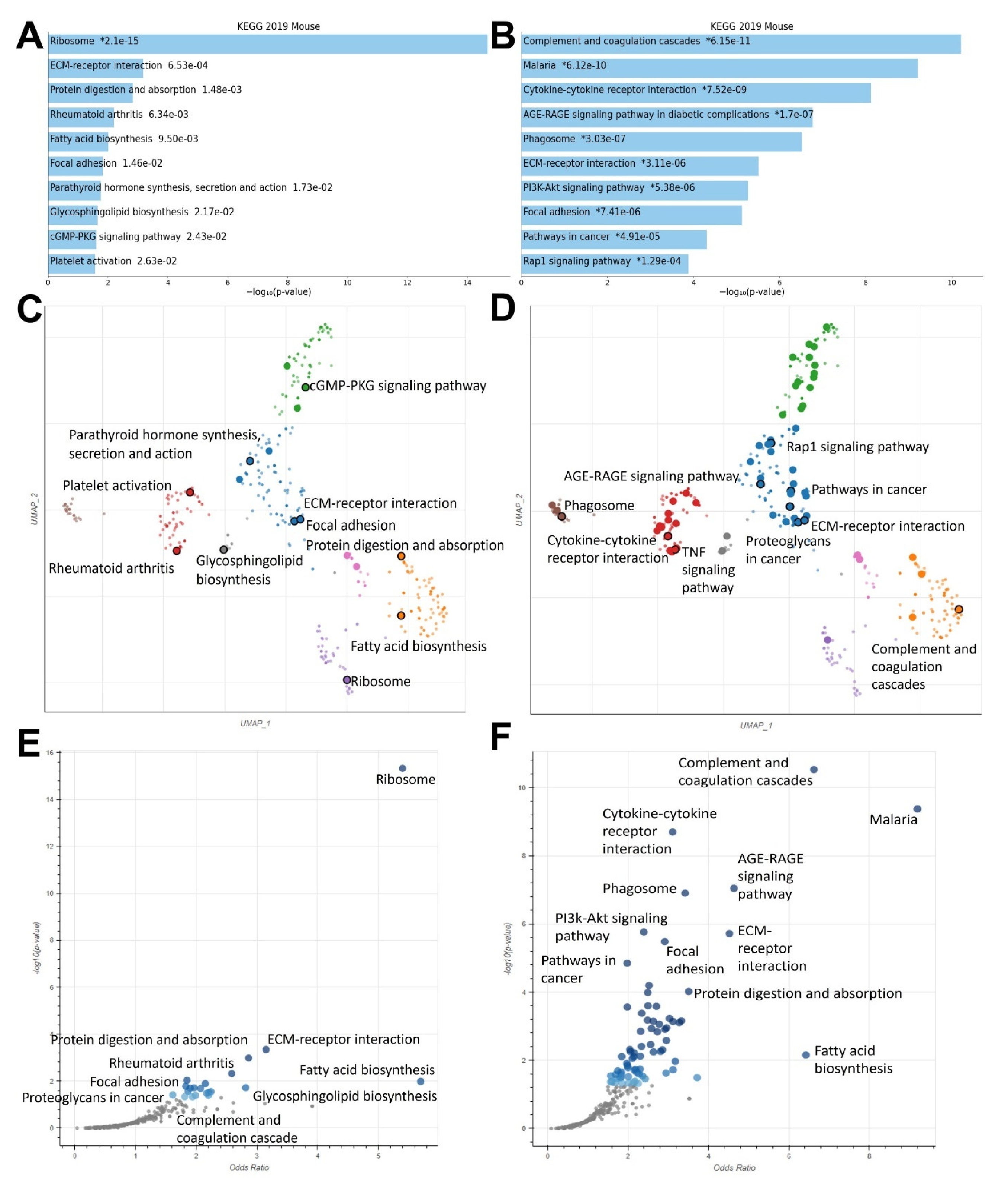
2.3. RXR Agonists Regulate Cancer-Relevant Biological Pathways in MMTV-Neu Tumors
2.4. MSU-42011 and Bexarotene Induce Unique Gene Expression Profiles with Some Unifying Characteristics in Treated Tumors of a HER2+ Murine Model
2.5. MSU-42011 Increases Col6a3 and Map9 Expression in Mouse Mammary Tumors
2.6. MSU-42011 Increases IL-18 and H2-AA Expression in Mouse Mammary Tumors
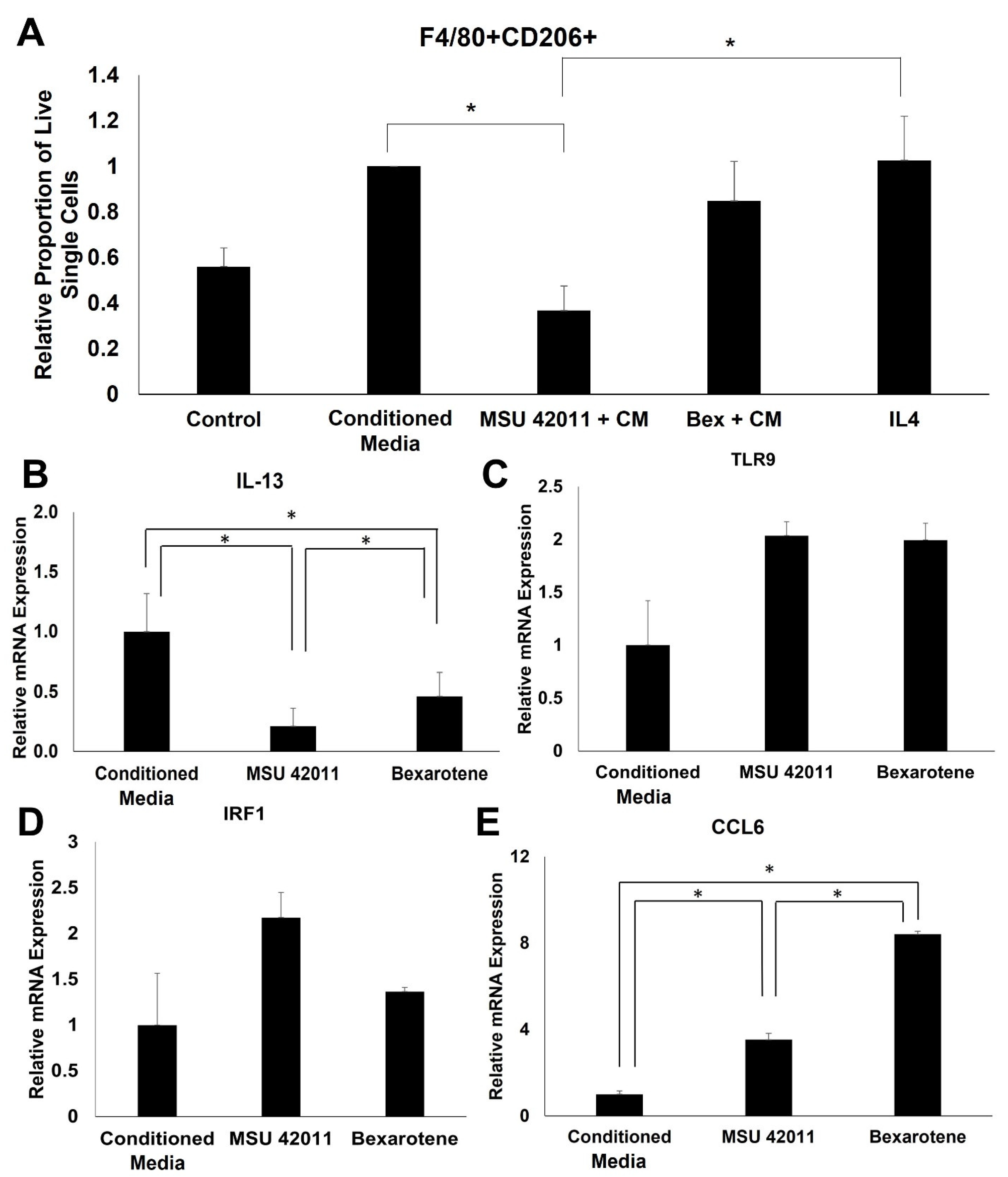
2.7. MSU-42011 Polarizes Bone Marrow-Derived Macrophages (BMDMs) towards an Anti-Tumor Phenotype
3. Discussion
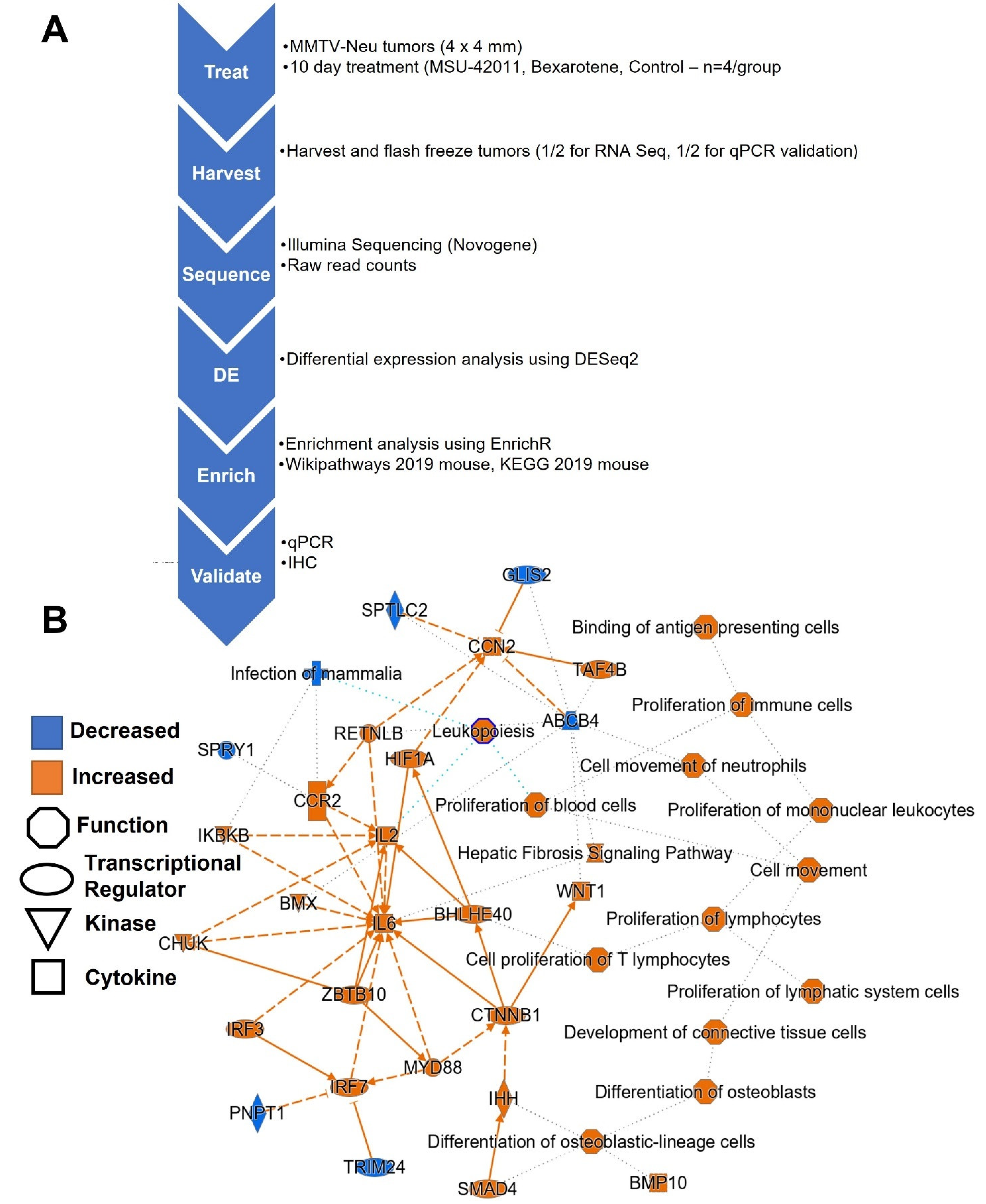
4. Materials and Methods
4.1. Drugs
4.2. In Vitro Experiments
4.3. Flow Cytometry
4.4. In Vivo Experiments
4.5. RNA Sequencing
4.6. qPCR
4.7. Immunohistochemistry
4.8. KmPlot Generation
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenfeld, M.G.; Lunyak, V.V.; Glass, C.K. Sensors and signals: A coactivator/corepressor/epigenetic code for integrating signal-dependent programs of transcriptional response. Genes Dev. 2006, 20, 1405–1428. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, B.W.; Malovannaya, A.; Qin, J. Minireview: Nuclear receptor and coregulator proteomics—2012 and beyond. Mol. Endocrin. 2012, 26, 1646–1650. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.M.; Mangelsdorf, D.J. Nuclear Receptors, RXR, and the Big Bang. Cell 2014, 157, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, A.S.; Reich, L.A.; Moerland, J.A.; Zhang, D.; Liby, K.T. Potential therapeutic uses of rexinoids. Adv. Pharmacol. 2021, 91, 141–183. [Google Scholar] [PubMed]
- Hurst, R.E. Bexarotene ligand pharmaceuticals. Curr. Opin. Investig. Drugs 2000, 1, 514–523. [Google Scholar] [PubMed]
- Esteva, F.J.; Glaspy, J.; Baidas, S.; Laufman, L.; Hutchins, L.; Dickler, M.; Tripathy, D.; Cohen, R.; DeMichele, A.; Yocum, R.C.; et al. Multicenter phase II study of oral bexarotene for patients with metastatic breast cancer. J. Clin. Oncol. 2003, 21, 999–1006. [Google Scholar] [CrossRef]
- Khuri, F.R.; Rigas, J.R.; Figlin, R.A.; Gralla, R.J.; Shin, D.M.; Munden, R.; Fox, N.; Huyghe, M.R.; Kean, Y.; Reich, S.D.; et al. Multi-Institutional Phase I/II Trial of Oral Bexarotene in Combination with Cisplatin and Vinorelbine in Previously Untreated Patients with Advanced Non–Small-Cell Lung Cancer. J. Clin. Oncol. 2001, 19, 2626–2637. [Google Scholar] [CrossRef]
- Zhang, Q.; Pan, J.; Zhang, J.; Liu, P.; Chen, R.; Chen, D.; Lubet, R.; Wang, Y.; You, M. Aerosolized Bexarotene Inhibits Lung Tumorigenesis without Increasing Plasma Triglyceride and Cholesterol Levels in Mice. Can. Prev. Res. 2011, 4, 270. [Google Scholar] [CrossRef] [Green Version]
- Moerland, J.A.; Zhang, D.; Reich, L.A.; Carapellucci, S.; Lockwood, B.; Leal, A.S.; Krieger-Burke, T.; Aleiwi, B.; Ellsworth, E.; Liby, K.T. The novel rexinoid MSU-42011 is effective for the treatment of preclinical Kras-driven lung cancer. Sci. Rep. 2020, 10, 22244. [Google Scholar] [CrossRef]
- Reich, L.A.; Moerland, J.A.; Leal, A.S.; Zhang, D.; Carapellucci, S.; Lockwood, B.; Jurutka, P.W.; Marshall, P.A.; Wagner, C.E.; Liby, K.T. The rexinoid V-125 reduces tumor growth in preclinical models of breast and lung cancer. Sci. Rep. 2022, 12, 293. [Google Scholar] [CrossRef]
- Leal, A.S.; Moerland, J.A.; Zhang, D.; Carapellucci, S.; Lockwood, B.; Krieger-Burke, T.; Aleiwi, B.; Ellsworth, E.; Liby, K.T. The RXR Agonist MSU42011 Is Effective for the Treatment of Preclinical HER2+ Breast Cancer and Kras-Driven Lung Cancer. Cancers 2021, 13, 5004. [Google Scholar] [CrossRef] [PubMed]
- Andrechek, E.R.; Laing, M.A.; Girgis-Gabardo, A.A.; Siegel, P.M.; Cardiff, R.D.; Muller, W.J. Gene Expression Profiling of Neu-induced Mammary Tumors from Transgenic Mice Reveals Genetic and Morphological Similarities to ErbB2-expressing Human Breast Cancers1. Can. Res. 2003, 63, 4920–4926. [Google Scholar]
- Rennhack, J.P.; To, B.; Swiatnicki, M.; Dulak, C.; Ogrodzinski, M.P.; Zhang, Y.; Li, C.; Bylett, E.; Ross, C.; Szczepanek, K.; et al. Integrated analyses of murine breast cancer models reveal critical parallels with human disease. Nat. Comm. 2019, 10, 3261. [Google Scholar] [CrossRef] [Green Version]
- Guy, C.T.; Webster, M.A.; Schaller, M.; Parsons, T.J.; Cardiff, R.D.; Muller, W.J. Expression of the neu protooncogene in the mammary epithelium of transgenic mice induces metastatic disease. Proc. Natl. Acad. Sci. USA 1992, 89, 10578–10582. [Google Scholar] [CrossRef] [Green Version]
- Shulman, A.I.; Larson, C.; Mangelsdorf, D.J.; Ranganathan, R. Structural determinants of allosteric ligand activation in RXR heterodimers. Cell 2004, 116, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Shiota, G.; Kanki, K. Retinoids and their target genes in liver functions and diseases. J. Gastroenterol. Hepatol. 2013, 28, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, R.; Pedersen, T.A.; Hagenbeek, D.; Moulos, P.; Siersbaek, R.; Megens, E.; Denissov, S.; Børgesen, M.; Francoijs, K.J.; Mandrup, S.; et al. Genome-wide profiling of PPARgamma:RXR and RNA polymerase II occupancy reveals temporal activation of distinct metabolic pathways and changes in RXR dimer composition during adipogenesis. Genes Dev. 2008, 22, 2953–2967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, N.; Yu, R.T.; Subramaniam, N.; Sherman, M.H.; Wilson, C.; Rao, R.; Leblanc, M.; Coulter, S.; He, M.; Scott, C.; et al. A vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic response. Cell 2013, 153, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Delfosse, V.; Huet, T.; Harrus, D.; Granell, M.; Bourguet, M.; Gardia-Parège, C.; Chiavarina, B.; Grimaldi, M.; Le Mével, S.; Blanc, P. Mechanistic insights into the synergistic activation of the RXR–PXR heterodimer by endocrine disruptor mixtures. Proc. Natl. Acad. Sci. USA 2021, 118, e2020551118. [Google Scholar] [CrossRef]
- Ihunnah, C.A.; Jiang, M.; Xie, W. Nuclear receptor PXR, transcriptional circuits and metabolic relevance. Biochim. Biophys. Acta 2011, 1812, 956–963. [Google Scholar] [CrossRef] [Green Version]
- Abba, M.C.; Hu, Y.; Levy, C.C.; Gaddis, S.; Kittrell, F.S.; Zhang, Y.; Hill, J.; Bissonnette, R.P.; Medina, D.; Brown, P.H.; et al. Transcriptomic signature of Bexarotene on mammary gland from three transgenic mouse mammary cancer models. BMC Med. Genom. 2008, 1, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Győrffy, B. Survival analysis across the entire transcriptome identifies biomarkers with the highest prognostic power in breast cancer. Comput. Struct. Biotech. J. 2021, 19, 4101–4109. [Google Scholar] [CrossRef] [PubMed]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, R.; Cai, Q.; Gao, X.; Tong, F.; Dong, J.; Hu, Y.; Wu, G.; Dong, X. MicroRNA-330-3p promotes brain metastasis and epithelial-mesenchymal transition via GRIA3 in non-small cell lung cancer. Aging 2019, 11, 6734–6761. [Google Scholar] [CrossRef]
- Heger, L.; Balk, S.; Lühr, J.J.; Heidkamp, G.F.; Lehmann, C.H.K.; Hatscher, L.; Purbojo, A.; Hartmann, A.; Garcia-Martin, F.; Nishimura, S.-I.; et al. CLEC10A Is a Specific Marker for Human CD1c+ Dendritic Cells and Enhances Their Toll-like Receptor 7/8-Induced Cytokine Secretion. Front. Immunol. 2018, 9, 744. [Google Scholar] [CrossRef]
- Jiang, T.; Gao, W.; Lin, S.; Chen, H.; Du, B.; Liu, Q.; Lin, X.; Chen, Q. FNDC1 Promotes the Invasiveness of Gastric Cancer via Wnt/β-Catenin Signaling Pathway and Correlates With Peritoneal Metastasis and Prognosis. Front. Oncol. 2020, 10, 590492. [Google Scholar] [CrossRef]
- Abudureyimu, S.; Asai, N.; Enomoto, A.; Weng, L.; Kobayashi, H.; Wang, X.; Chen, C.; Mii, S.; Takahashi, M. Essential Role of Linx/Islr2 in the Development of the Forebrain Anterior Commissure. Sci. Rep. 2018, 8, 7292. [Google Scholar] [CrossRef] [Green Version]
- Navab, R.; Strumpf, D.; To, C.; Pasko, E.; Kim, K.S.; Park, C.J.; Hai, J.; Liu, J.; Jonkman, J.; Barczyk, M.; et al. Integrin α11β1 regulates cancer stromal stiffness and promotes tumorigenicity and metastasis in non-small cell lung cancer. Oncogene 2016, 35, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.; Kim, I.K.; Kim, H.; Choi, M.J.; Lee, J.; Jo, Y.S. Liver X Receptor β Related to Tumor Progression and Ribosome Gene Expression in Papillary Thyroid Cancer. Endocrinol. Metab. 2020, 35, 656–668. [Google Scholar] [CrossRef]
- Lu, X.; Gao, C.; Liu, C.; Zhuang, J.; Su, P.; Li, H.; Wang, X.; Sun, C. Identification of the key pathways and genes involved in HER2-positive breast cancer with brain metastasis. Pathol. Res. Pract. 2019, 215, 152475. [Google Scholar] [CrossRef] [PubMed]
- Schenck, J.; Fitzgerald, J. Analysis of COL6A3, COL6A5 and COL6A6 gene expression in breast and prostate tumors. Adv. Clin. Transl. Res. 2018, 2, 1–10. [Google Scholar]
- Rouquier, S.; Pillaire, M.-J.; Cazaux, C.; Giorgi, D. Expression of the Microtubule-Associated Protein MAP9/ASAP and Its Partners AURKA and PLK1 in Colorectal and Breast Cancers. Dis. Markers 2014, 2014, 798170. [Google Scholar] [CrossRef] [Green Version]
- Leal, A.S.; Zydeck, K.; Carapellucci, S.; Reich, L.A.; Zhang, D.; Moerland, J.A.; Sporn, M.B.; Liby, K.T. Retinoid X receptor agonist LG100268 modulates the immune microenvironment in preclinical breast cancer models. Npj Breast Cancer 2019, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Royce, D.B.; Risingsong, R.; Williams, C.R.; Sporn, M.B.; Liby, K.T. The Rexinoids LG100268 and LG101506 Inhibit Inflammation and Suppress Lung Carcinogenesis in A/J Mice. Cancer Prev. Res. 2016, 9, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguiar, M.A.N.; Wanderley, C.W.S.; Nobre, L.M.S.; Alencar, M.R.M.; Saldanha, M.d.P.S.; Souza, A.M.; Wong, D.V.T.; Barros, P.G.; Almeida, P.R.C.; Lima-Júnior, R.C.P.; et al. Interleukin-18 (IL-18) is equally expressed in inflammatory breast cancer and noninflammatory locally advanced breast cancer: A possible association with chemotherapy response. Asia Pac. J. Clin. Oncol. 2018, 14, e138–e144. [Google Scholar] [CrossRef] [Green Version]
- Senju, H.; Kumagai, A.; Nakamura, Y.; Yamaguchi, H.; Nakatomi, K.; Fukami, S.; Shiraishi, K.; Harada, Y.; Nakamura, M.; Okamura, H.; et al. Effect of IL-18 on the Expansion and Phenotype of Human Natural Killer Cells: Application to Cancer Immunotherapy. Int. J. Biol. Sci. 2018, 14, 331–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, T.B.; Meza-Perez, S.; Londoño, A.; Katre, A.; Peabody, J.E.; Smith, H.J.; Forero, A.; Norian, L.A.; Straughn, J.M., Jr.; Buchsbaum, D.J.; et al. Epigenetic modifiers upregulate MHC II and impede ovarian cancer tumor growth. Oncotarget 2017, 8, 44159–44170. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, M.L.; Cook, R.S.; Johnson, D.B.; Balko, J.M. Biological Consequences of MHC-II Expression by Tumor Cells in Cancer. Clin. Can. Res. 2019, 25, 2392–2402. [Google Scholar] [CrossRef] [PubMed]
- Magner, W.J.; Kazim, A.L.; Stewart, C.; Romano, M.A.; Catalano, G.; Grande, C.; Keiser, N.; Santaniello, F.; Tomasi, T.B. Activation of MHC class I, II, and CD40 gene expression by histone deacetylase inhibitors. J. Immunol. 2000, 165, 7017–7024. [Google Scholar] [CrossRef] [Green Version]
- Cycon, K.A.; Mulvaney, K.; Rimsza, L.M.; Persky, D.; Murphy, S.P. Histone deacetylase inhibitors activate CIITA and MHC class II antigen expression in diffuse large B-cell lymphoma. Immunology 2013, 140, 259–272. [Google Scholar] [CrossRef]
- Forero, A.; Li, Y.; Chen, D.; Grizzle, W.E.; Updike, K.L.; Merz, N.D.; Downs-Kelly, E.; Burwell, T.C.; Vaklavas, C.; Buchsbaum, D.J.; et al. Expression of the MHC Class II Pathway in Triple-Negative Breast Cancer Tumor Cells Is Associated with a Good Prognosis and Infiltrating Lymphocytes. Can. Immunol. Res. 2016, 4, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Li, L.; Ye, H.; Tao, H.; He, H. Role of COL6A3 in colorectal cancer. Oncol. Rep. 2018, 39, 2527–2536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, L.; Yang, H.; Li, C.; Fang, C. Identification of potential genes related to breast cancer brain metastasis in breast cancer patients. Biosci. Rep. 2021, 41, BSR20211615. [Google Scholar] [CrossRef]
- Al Amri, W.S.; Baxter, D.E.; Hanby, A.M.; Stead, L.F.; Verghese, E.T.; Thorne, J.L.; Hughes, T.A. Identification of candidate mediators of chemoresponse in breast cancer through therapy-driven selection of somatic variants. Breast Can. Res. Treat. 2020, 183, 607–616. [Google Scholar] [CrossRef]
- Wang, J.; Pan, W. The Biological Role of the Collagen Alpha-3 (VI) Chain and Its Cleaved C5 Domain Fragment Endotrophin in Cancer. Onco Targets Ther. 2020, 13, 5779–5793. [Google Scholar] [CrossRef]
- Guglielmi, V.; Cardellini, M.; Cinti, F.; Corgosinho, F.; Cardolini, I.; D’Adamo, M.; Zingaretti, M.C.; Bellia, A.; Lauro, D.; Gentileschi, P.; et al. Omental adipose tissue fibrosis and insulin resistance in severe obesity. Nutr. Diabetes 2015, 5, e175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehbid, M.; Aleyasin, S.; Vaziri, H. Evaluation of DNA Methylation of MAP9 Gene in Breast Cancer as Epigenetic Biomarker. J. Mol. Biomark. Diagn. S 2016, 8, 2. [Google Scholar]
- Basbous, J.; Knani, D.; Bonneaud, N.; Giorgi, D.; Brondello, J.-M.; Rouquier, S. Induction of ASAP (MAP9) contributes to p53 stabilization in response to DNA damage. Cell Cycle 2012, 11, 2380–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Yang, Y.; Li, Z.; Zhao, D.; Zhang, W.; Jiang, Y.; Xue, D. Modular bioinformatics analysis demonstrates that a Toll-like receptor signaling pathway is involved in the regulation of macrophage polarization. Mol. Med. Rep. 2018, 18, 4313–4320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Leal, A.S.; Carapellucci, S.; Shahani, P.H.; Bhogal, J.S.; Ibrahim, S.; Raban, S.; Jurutka, P.W.; Marshall, P.A.; Sporn, M.B.; et al. Testing Novel Pyrimidinyl Rexinoids: A New Paradigm for Evaluating Rexinoids for Cancer Prevention. Cancer Prev. Res. 2019, 12, 211–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Rennhack, J.; Andrechek, E.R.; Rockwell, C.E.; Liby, K.T. Identification of an Unfavorable Immune Signature in Advanced Lung Tumors from Nrf2-Deficient Mice. Antioxid. Redox Signal. 2018, 29, 1535–1552. [Google Scholar] [CrossRef] [PubMed]
- Crowe, A.R.; Yue, W. Semi-quantitative Determination of Protein Expression using Immunohistochemistry Staining and Analysis: An Integrated Protocol. Bio Protoc. 2019, 9, e3465. [Google Scholar] [CrossRef] [PubMed]
- Gheban, B.-A.; Colosi, H.A.; Gheban-Roșca, I.-A.; Georgiu, C.; Gheban, D.; Crişan, D.; Crişan, M. Techniques for digital histological morphometry of the pineal gland. Acta Histochem. 2022, 124, 151897. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; European Molecular Biology Laboratory: Heidelberg, Germany, 2012; Volume 10. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reich, L.A.; Leal, A.S.; Ellsworth, E.; Liby, K.T. The Novel RXR Agonist MSU-42011 Differentially Regulates Gene Expression in Mammary Tumors of MMTV-Neu Mice. Int. J. Mol. Sci. 2023, 24, 4298. https://doi.org/10.3390/ijms24054298
Reich LA, Leal AS, Ellsworth E, Liby KT. The Novel RXR Agonist MSU-42011 Differentially Regulates Gene Expression in Mammary Tumors of MMTV-Neu Mice. International Journal of Molecular Sciences. 2023; 24(5):4298. https://doi.org/10.3390/ijms24054298
Chicago/Turabian StyleReich, Lyndsey A., Ana S. Leal, Edmund Ellsworth, and Karen T. Liby. 2023. "The Novel RXR Agonist MSU-42011 Differentially Regulates Gene Expression in Mammary Tumors of MMTV-Neu Mice" International Journal of Molecular Sciences 24, no. 5: 4298. https://doi.org/10.3390/ijms24054298