
Increased Density of Endogenous Adenosine A2A Receptors in Atrial Fibrillation: From Cellular and Porcine Models to Human Patients

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
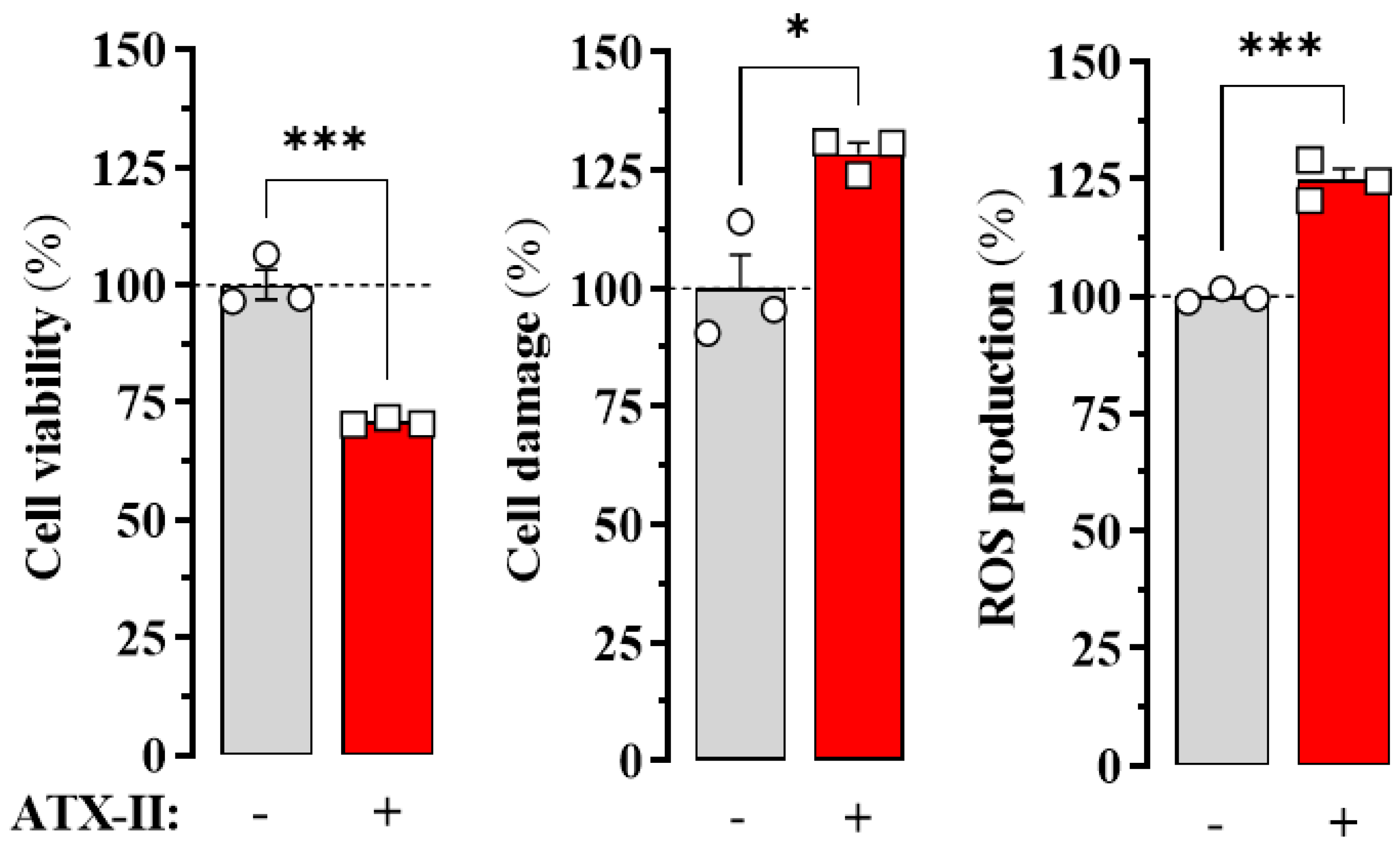
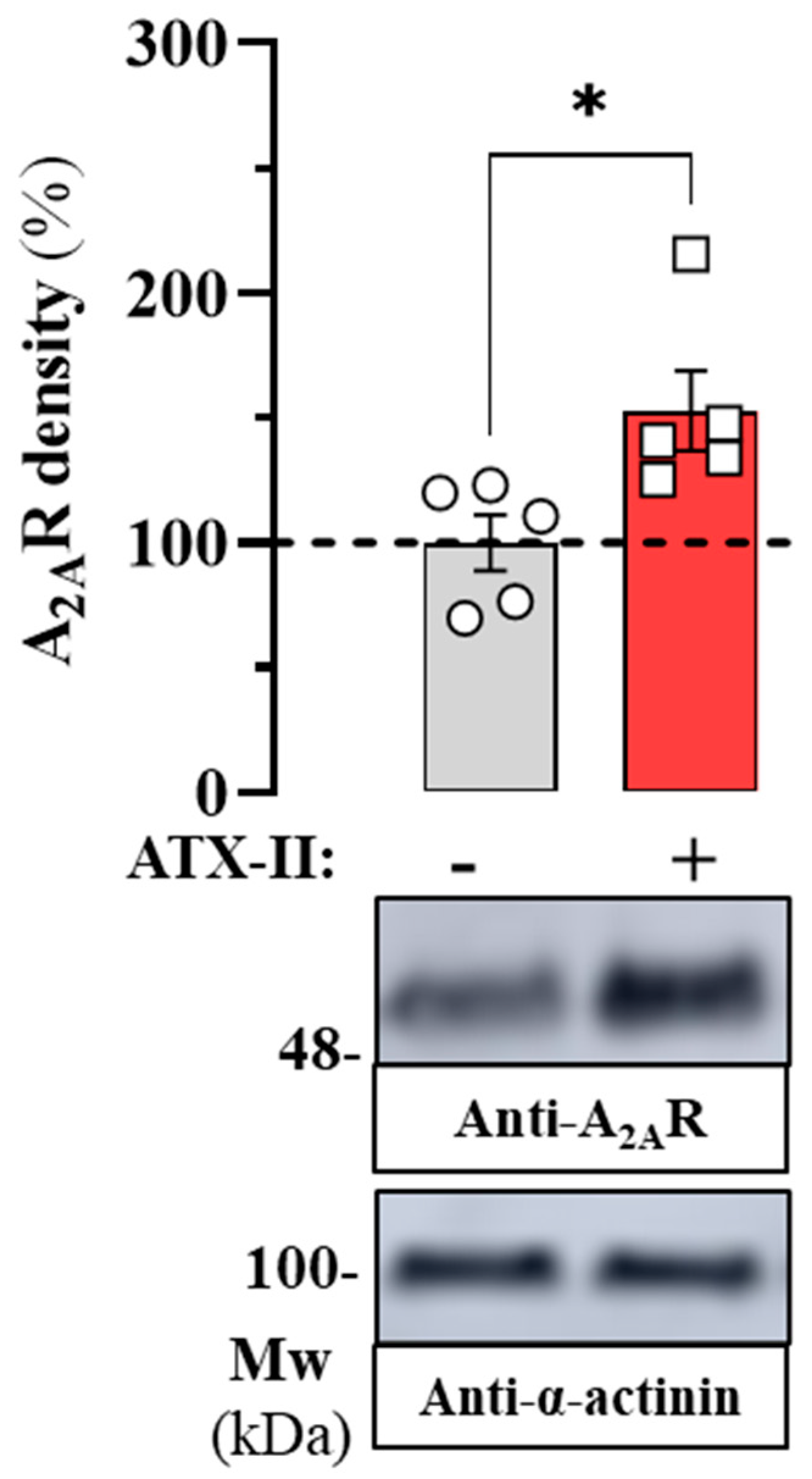
2.1. Endogenous A2AR Density in the HL-1 Cell Line
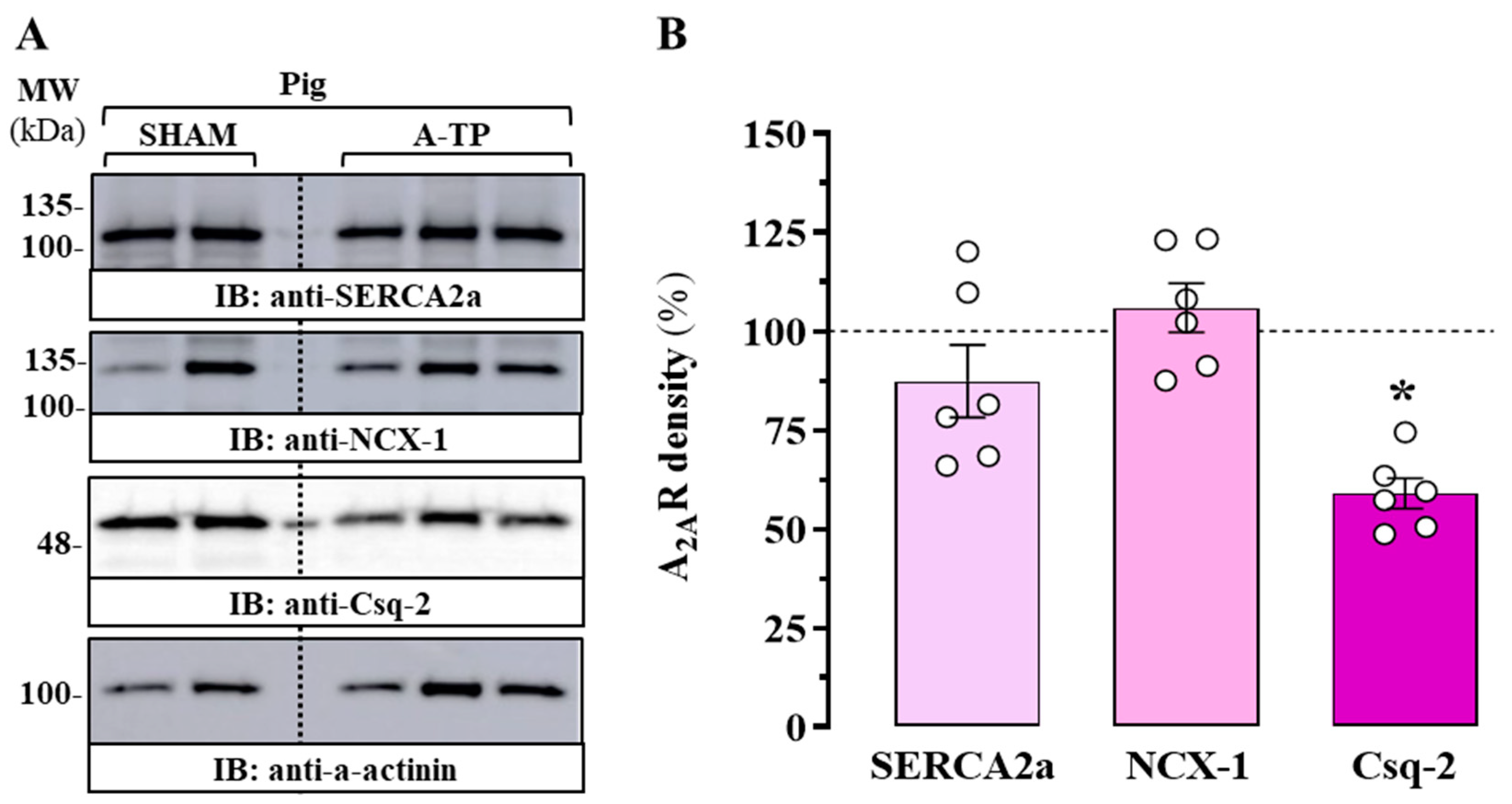
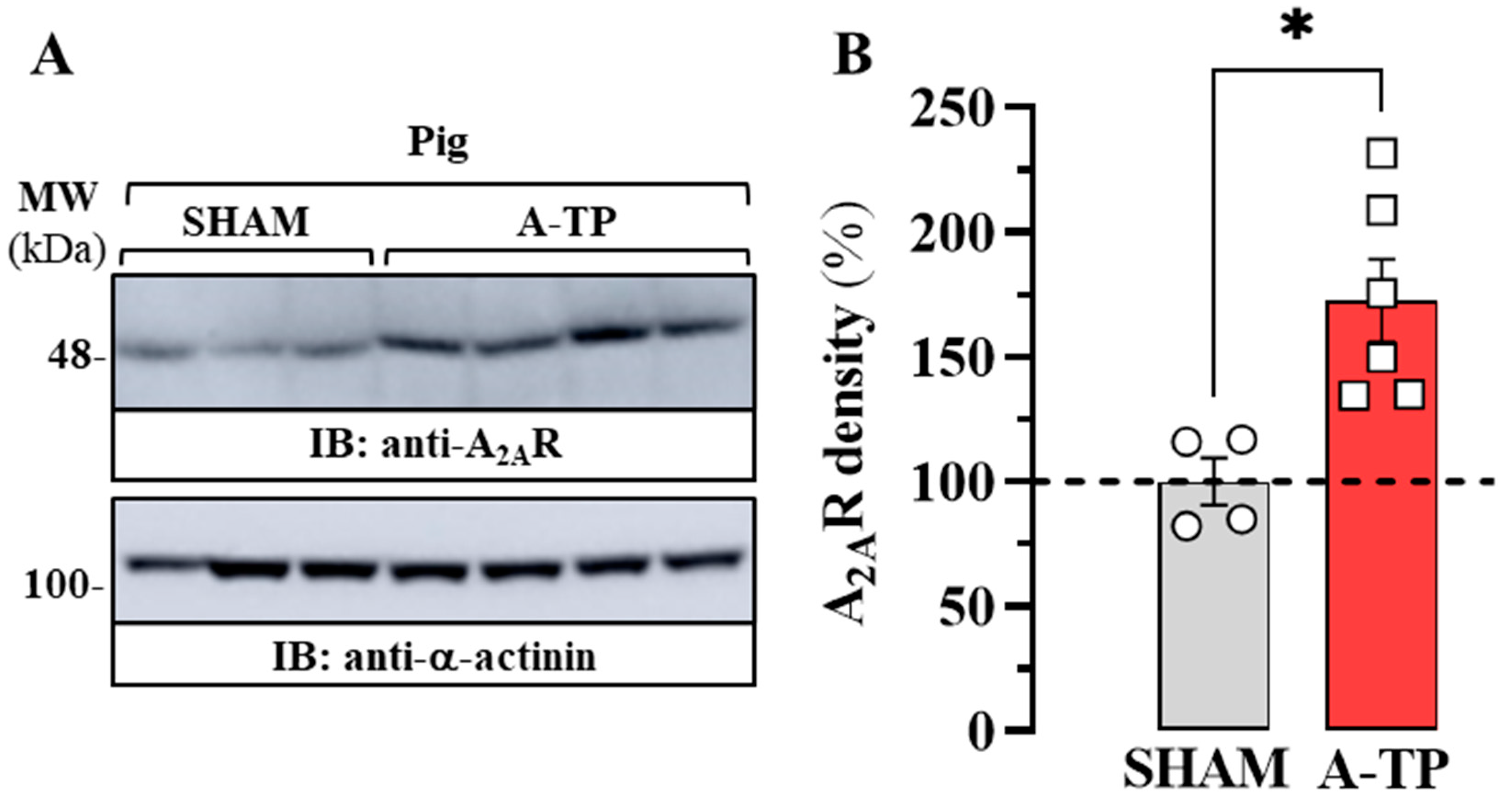
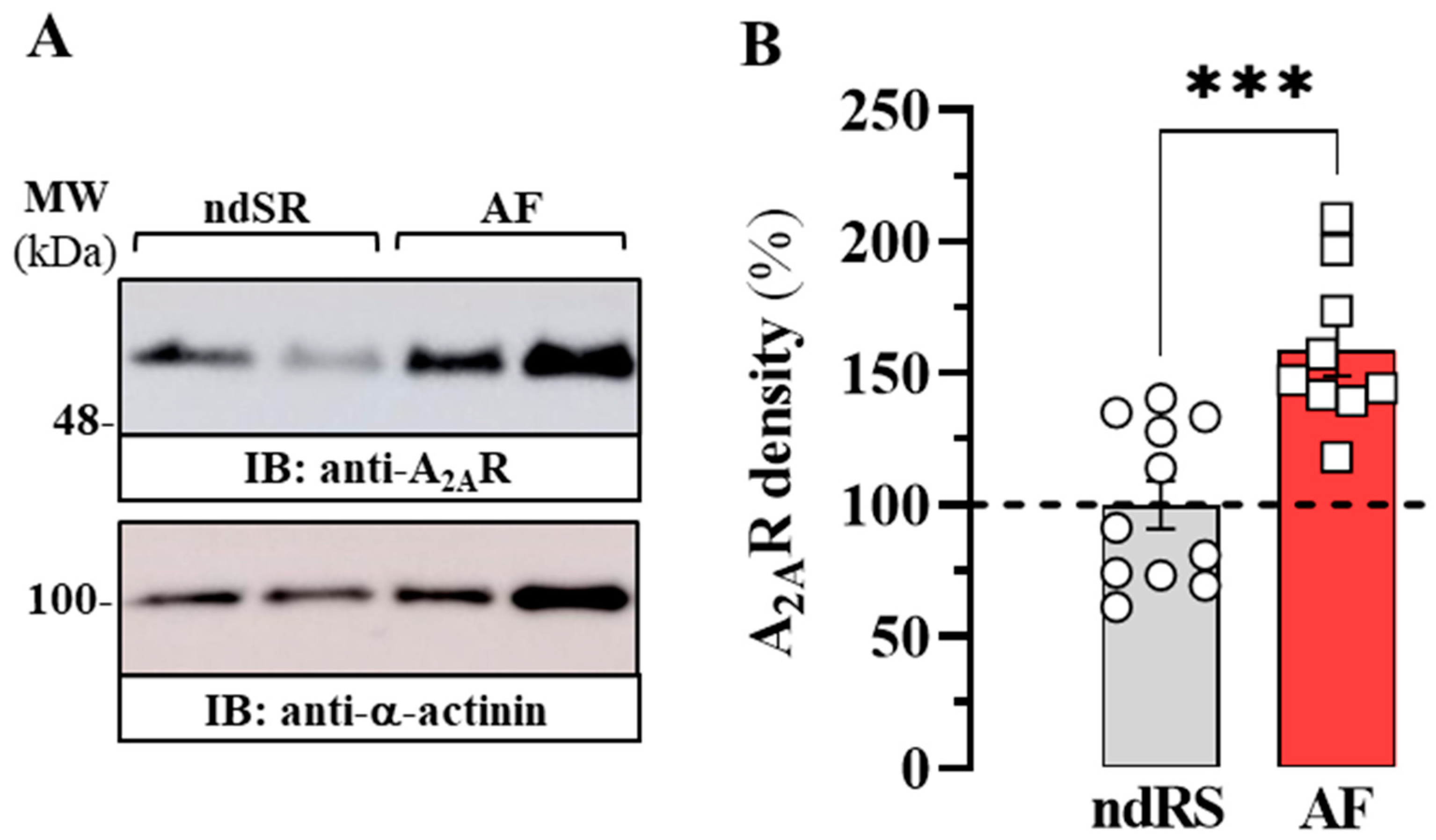
2.2. Expression of A2AR in the Atrium of a Porcine Model of AF
2.3. Expression of A2AR in the Atrium of Patients with AF
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Viability Assays
4.3. Porcine Model of AF
4.4. Human Samples and Subject Demographics
4.5. Preparation of Membrane Extracts
4.6. Gel Electrophoresis and Immunoblotting
4.7. Data and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vest, J.A.; Wehrens, X.H.T.; Reiken, S.R.; Lehnart, S.E.; Dobrev, D.; Chandra, P.; Danilo, P.; Ravens, U.; Rosen, M.R.; Marks, A.R. Defective Cardiac Ryanodine Receptor Regulation during Atrial Fibrillation. Circulation 2005, 111, 2025–2032. [Google Scholar] [CrossRef]
- Kabell, G.; Buchanan, L.V.; Gibson, J.K.; Belardinelli, L. Effects of Adenosine on Atrial Refractoriness and Arrhythmias. Cardiovasc. Res. 1994, 28, 1385–1389. [Google Scholar] [CrossRef]
- Workman, A.J. Cardiac Adrenergic Control and Atrial Fibrillation. Naunyn Schmiedebergs Arch. Pharmacol. 2010, 381, 235. [Google Scholar] [CrossRef]
- Hove-Madsen, L.; Prat-Vidal, C.; Llach, A.; Ciruela, F.; Casadó, V.; Lluis, C.; Bayes-Genis, A.; Cinca, J.; Franco, R. Adenosine A<inf>2A</Inf>receptors Are Expressed in Human Atrial Myocytes and Modulate Spontaneous Sarcoplasmic Reticulum Calcium Release. Cardiovasc. Res. 2006, 72, 292–302. [Google Scholar] [CrossRef]
- Soattin, L.; Lubberding, A.F.; Bentzen, B.H.; Christ, T.; Jespersen, T. Inhibition of Adenosine Pathway Alters Atrial Electrophysiology and Prevents Atrial Fibrillation. Front. Physiol. 2020, 11, 493. [Google Scholar] [CrossRef]
- Godoy-Marín, H.; Duroux, R.; Jacobson, K.A.; Soler, C.; Colino-Lage, H.; Jiménez-Sábado, V.; Montiel, J.; Hove-Madsen, L.; Ciruela, F. Adenosine A2a Receptors Are Upregulated in Peripheral Blood Mononuclear Cells from Atrial Fibrillation Patients. Int. J. Mol. Sci 2021, 22, 3467. [Google Scholar] [CrossRef]
- Llach, A.; Molina, C.E.; Prat-Vidal, C.; Fernandes, J.; Casadó, V.; Ciruela, F.; Lluís, C.; Franco, R.; Cinca, J.; Hove-Madsen, L. Abnormal Calcium Handling in Atrial Fibrillation Is Linked to Up-Regulation of Adenosine A2A Receptors. Eur. Heart J. 2011, 32, 721–729. [Google Scholar] [CrossRef]
- Molina, C.E.; Llach, A.; Herraiz-Martínez, A.; Tarifa, C.; Barriga, M.; Wiegerinck, R.F.; Fernandes, J.; Cabello, N.; Vallmitjana, A.; Benitéz, R.; et al. Prevention of Adenosine A2A Receptor Activation Diminishes Beat-to-Beat Alternation in Human Atrial Myocytes. Basic Res. Cardiol. 2016, 111, 5. [Google Scholar] [CrossRef]
- Dobrev, D.; Friedrich, A.; Voigt, N.; Jost, N.; Wettwer, E.; Christ, T.; Knaut, M.; Ravens, U. The G Protein-Gated Potassium Current IK,ACh Is Constitutively Active in Patients with Chronic Atrial Fibrillation. Circulation 2005, 112, 3697–3706. [Google Scholar] [CrossRef]
- Takemoto, Y.; Slough, D.P.; Meinke, G.; Katnik, C.; Graziano, Z.A.; Chidipi, B.; Reiser, M.; Alhadidy, M.M.; Ramirez, R.; Salvador-Montañés, O.; et al. Structural Basis for the Antiarrhythmic Blockade of a Potassium Channel with a Small Molecule. FASEB J. 2018, 32, 1778–1793. [Google Scholar] [CrossRef]
- Noujaim, S.F.; Pandit, S.V.; Berenfeld, O.; Vikstrom, K.; Cerrone, M.; Mironov, S.; Zugermayr, M.; Lopatin, A.N.; Jalife, J. Up-Regulation of the Inward Rectifier K+ Current (IK1) in the Mouse Heart Accelerates and Stabilizes Rotors. J. Physiol. 2007, 578, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Chaudary, N.; Shuralyova, I.; Liron, T.; Sweeney, G.; Coe, I.R. Transport Characteristics of HL-1 Cells: A New Model for the Study of Adenosine Physiology in Cardiomyocytes. Biochem. Cell Biol. 2002, 80, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.Y.; Cheng, C.C.; Chen, Y.C.; Chen, S.A.; Chen, Y.J. ATX-II-Induced Pulmonary Vein Arrhythmogenesis Related to Atrial Fibrillation and Long QT Syndrome. Eur. J. Clin. Investig. 2012, 42, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Cai, L.; Zhang, J.; Qi, X.; Lu, C. Aconitine-Induced Cardiac Arrhythmia in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Exp. Ther. Med. 2018, 16, 3497–3503. [Google Scholar] [CrossRef] [PubMed]
- Citerni, C.; Kirchhoff, J.; Olsen, L.H.; Sattler, S.M.; Gentilini, F.; Forni, M.; Zannoni, A.; Grunnet, M.; Edvardsson, N.; Bentzen, B.H.; et al. Characterization of Atrial and Ventricular Structural Remodeling in a Porcine Model of Atrial Fibrillation Induced by Atrial Tachypacing. Front. Vet. Sci 2020, 7, 179. [Google Scholar] [CrossRef] [PubMed]
- Greiser, M.; Lederer, W.J.; Schotten, U. Alterations of Atrial Ca(2+) Handling as Cause and Consequence of Atrial Fibrillation. Cardiovasc. Res. 2011, 89, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Herraiz-Martínez, A.; Tarifa, C.; Jiménez-Sábado, V.; Llach, A.; Godoy-Marín, H.; Colino-Lage, H.; Nolla-Colomer, C.; Casabella-Ramon, S.; Izquierdo-Castro, P.; Benítez, I.; et al. Influence of Sex on Intracellular Calcium Homoeostasis in Patients with Atrial Fibrillation. Cardiovasc. Res. 2022, 118, 1033–1045. [Google Scholar] [CrossRef]
- van Gorp, P.R.R.; Trines, S.A.; Pijnappels, D.A.; de Vries, A.A.F. Multicellular In Vitro Models of Cardiac Arrhythmias: Focus on Atrial Fibrillation. Front. Cardiovasc. Med. 2020, 7, 43. [Google Scholar] [CrossRef]
- Antzelevitch, C.; Nesterenko, V.; Shryock, J.C.; Rajamani, S.; Song, Y.; Belardinelli, L. The Role of Late Ina in Development of Cardiac Arrhythmias. Handb. Exp. Pharmacol. 2014, 221, 137–168. [Google Scholar] [CrossRef]
- Schüttler, D.; Bapat, A.; Kaäb, S.; Lee, K.; Tomsits, P.; Clauss, S.; Hucker, W.J. Animal Models of Atrial Fibrillation. Circ. Res. 2020, 127, 91–110. [Google Scholar] [CrossRef]
- Ai, X. SR Calcium Handling Dysfunction, Stress-Response Signaling Pathways, and Atrial Fibrillation. Front. Physiol. 2015, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Dinanian, S.; Boixel, C.; Juin, C.; Hulot, J.S.; Coulombe, A.; Rücker-Martin, C.; Bonnet, N.; le Grand, B.; Slama, M.; Mercadier, J.J.; et al. Downregulation of the Calcium Current in Human Right Atrial Myocytes from Patients in Sinus Rhythm but with a High Risk of Atrial Fibrillation. Eur. Heart J. 2008, 29, 1190–1197. [Google Scholar] [CrossRef]
- Motulsky, H.J.; Brown, R.E. Detecting Outliers When Fitting Data with Nonlinear Regression—A New Method Based on Robust Nonlinear Regression and the False Discovery Rate. BMC Bioinform. 2006, 7, 123. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ndSR | AF | |
|---|---|---|
| Number of patients | 11 | 9 |
| Weight (mean ± SD) (Kg) | 79.75 ± 8.62 | 72.01 ± 12.01 |
| Height (mean ± SD) (cm) | 168.8 ± 8.7 | 162.92 ± 8.06 |
| Age (mean ± SD) | 64.7 ± 9.9 | 70.9 ± 10.5 |
| Sex (male/female) | 9/2 | 6/3 |
| Body Surface (mean ± SD) (m2) | 1.90 ± 0.12 | 9 |
| LA diameter (mean ± SD) (mm) | 41.1 ± 5.9 | 72.01 ± 12.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godoy-Marín, H.; Jiménez-Sábado, V.; Tarifa, C.; Ginel, A.; Santos, J.L.D.; Bentzen, B.H.; Hove-Madsen, L.; Ciruela, F. Increased Density of Endogenous Adenosine A2A Receptors in Atrial Fibrillation: From Cellular and Porcine Models to Human Patients. Int. J. Mol. Sci. 2023, 24, 3668. https://doi.org/10.3390/ijms24043668
Godoy-Marín H, Jiménez-Sábado V, Tarifa C, Ginel A, Santos JLD, Bentzen BH, Hove-Madsen L, Ciruela F. Increased Density of Endogenous Adenosine A2A Receptors in Atrial Fibrillation: From Cellular and Porcine Models to Human Patients. International Journal of Molecular Sciences. 2023; 24(4):3668. https://doi.org/10.3390/ijms24043668
Chicago/Turabian StyleGodoy-Marín, Héctor, Verónica Jiménez-Sábado, Carmen Tarifa, Antonino Ginel, Joana Larupa Dos Santos, Bo Hjorth Bentzen, Leif Hove-Madsen, and Francisco Ciruela. 2023. "Increased Density of Endogenous Adenosine A2A Receptors in Atrial Fibrillation: From Cellular and Porcine Models to Human Patients" International Journal of Molecular Sciences 24, no. 4: 3668. https://doi.org/10.3390/ijms24043668