
Ca2+- and Voltage-Activated K+ (BK) Channels in the Nervous System: One Gene, a Myriad of Physiological Functions

 , and
, and
Abstract
:1. Introduction
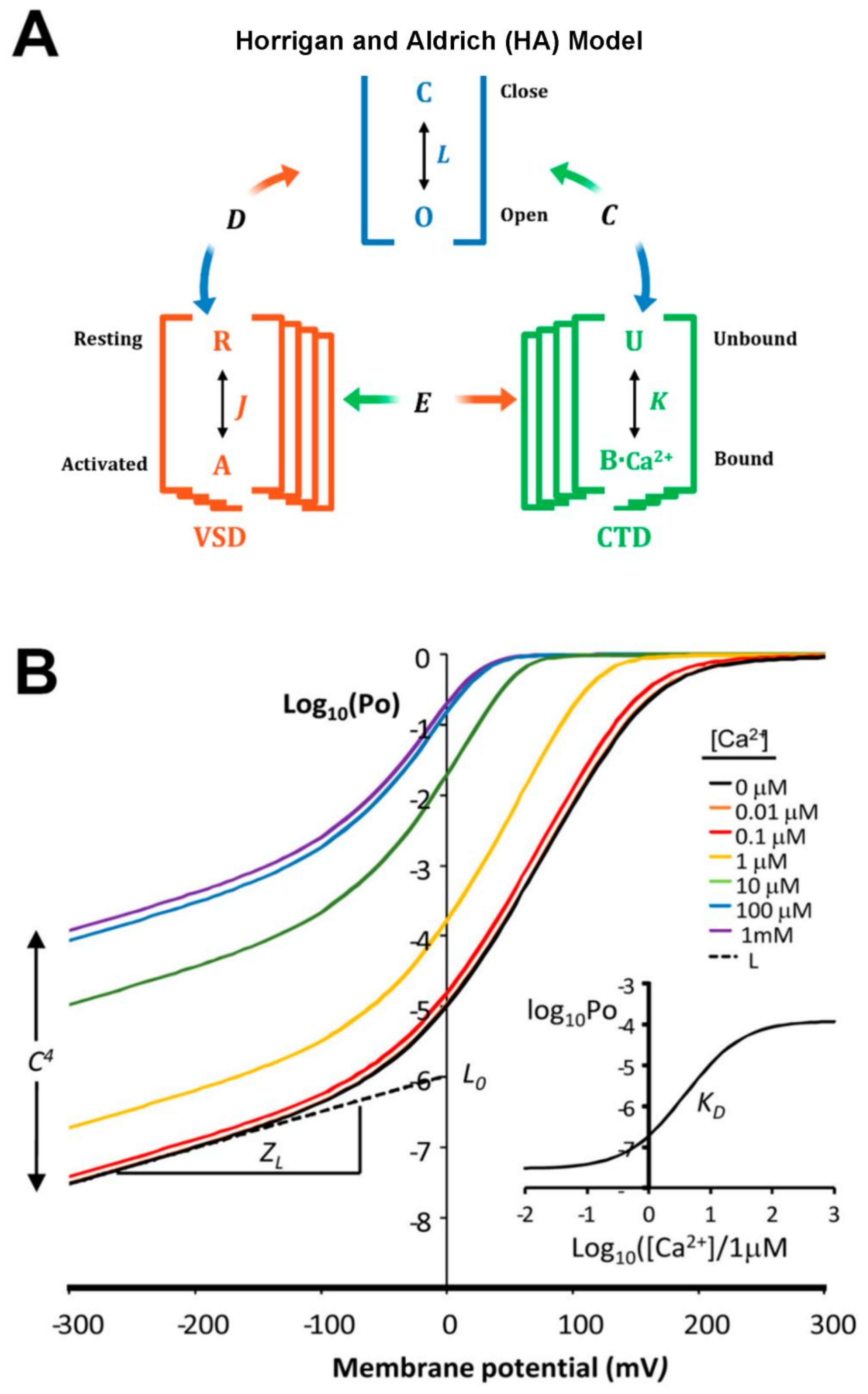
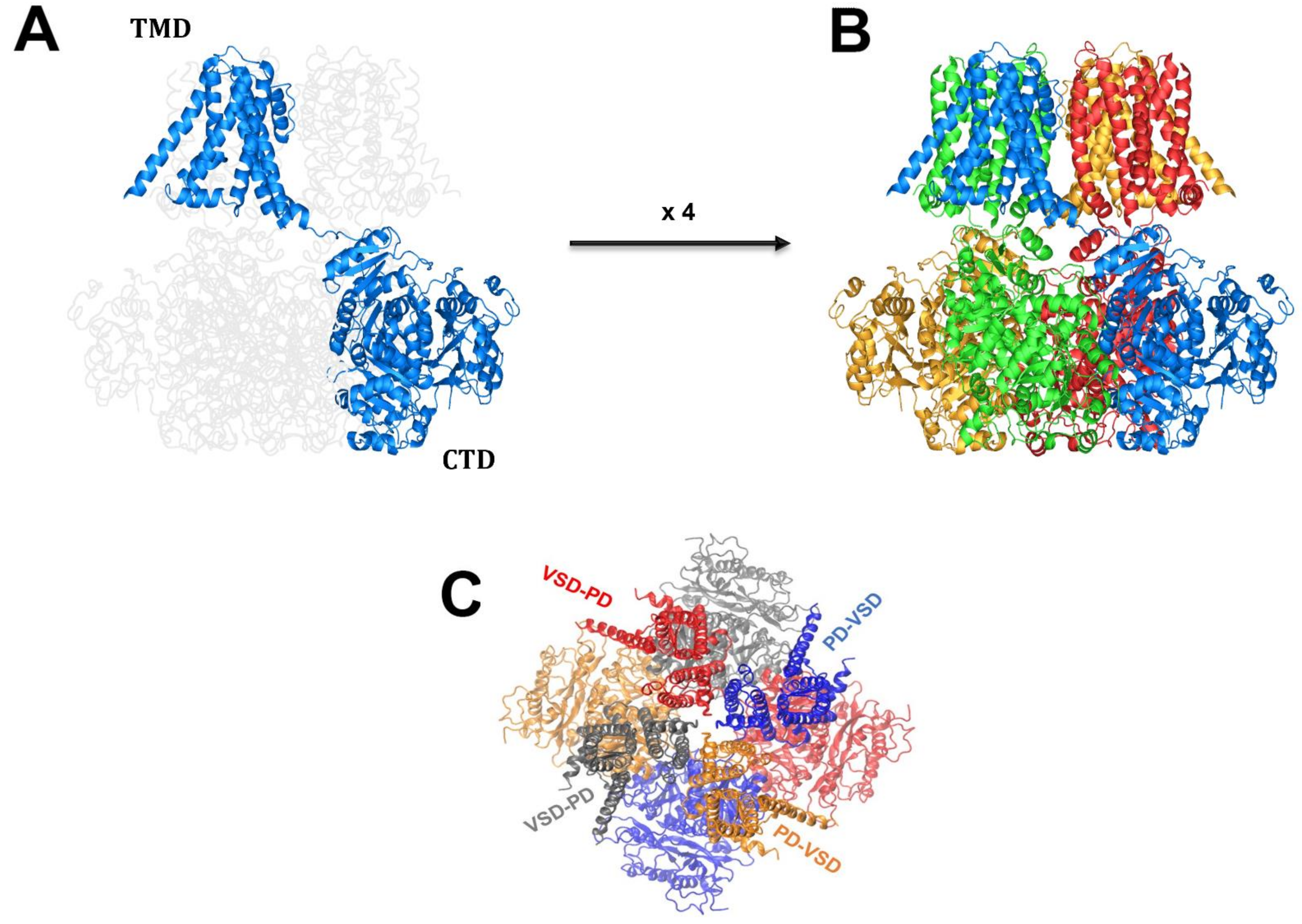
2. BK Biophysical and Structural Characteristics
3. BK Auxiliary Subunits
4. BK Channel Coupling with Ca2+-Permeable Channels
5. BK Post-Translational Modifications
6. BK Channels Differentially Contribute to Control of Brain Function
7. BK Channel Dysfunction in Neurological Disorders
8. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Pyott, S.; Duncan, R. BK Channels in the Vertebrate Inner Ear. Int. Rev. Neurobiol. 2016, 128, 369–399. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy-Natarajan, G.; Koide, M. BK Channels in the Vascular System. Int. Rev. Neurobiol. 2016, 128, 401–438. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.R.M.; Plante, A.E.; Meredith, A.L. Ion Channels Controlling Circadian Rhythms in Suprachiasmatic Nucleus Excitability. Physiol. Rev. 2020, 100, 1415–1454. [Google Scholar] [CrossRef]
- Knaus, H.; Schwarzer, C.; Koch, R.; Eberhart, A.; Kaczorowski, G.; Glossmann, H.; Wunder, F.; Pongs, O.; Garcia, M.; Sperk, G. Distribution of high-conductance Ca(2+)-activated K+ channels in rat brain: Targeting to axons and nerve terminals. J. Neurosci. 1996, 16, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Sausbier, U.; Sausbier, M.; Sailer, C.A.; Arntz, C.; Knaus, H.-G.; Neuhuber, W.; Ruth, P. Ca2+-activated K+ channels of the BK-type in the mouse brain. Histochem. Cell Biol. 2006, 125, 725–741. [Google Scholar] [CrossRef]
- Wanner, S.G.; Koch, R.O.; Koschak, A.; Trieb, M.; Garcia, M.L.; Kaczorowski, G.J.; Knaus, H.-G. High-Conductance Calcium-Activated Potassium Channels in Rat Brain: Pharmacology, Distribution, and Subunit Composition. Biochemistry 1999, 38, 5392–5400. [Google Scholar] [CrossRef]
- González-Sanabria, N.; Echeverría, F.; Segura, I.; Alvarado-Sánchez, R.; Latorre, R. BK in Double-Membrane Organelles: A Biophysical, Pharmacological, and Functional Survey. Front. Physiol. 2021, 12, 761474. [Google Scholar] [CrossRef]
- Hayashi, Y.; Morinaga, S.; Zhang, J.; Satoh, Y.; Meredith, A.; Nakata, T.; Wu, Z.; Kohsaka, S.; Inoue, K.; Nakanishi, H. BK channels in microglia are required for morphine-induced hyperalgesia. Nat. Commun. 2016, 7, 11697. [Google Scholar] [CrossRef]
- Kyle, B.D.; Braun, A.P. The regulation of BK channel activity by pre- and post-translational modifications. Front. Physiol. 2014, 5, 316. [Google Scholar] [CrossRef]
- Latorre, R.; Castillo, K.; Carrasquel-Ursulaez, W.; Sepulveda, R.V.; Gonzalez-Nilo, F.; Gonzalez, C.; Alvarez, O. Molecular Determinants of BK Channel Functional Diversity and Functioning. Physiol. Rev. 2017, 97, 39–87. [Google Scholar] [CrossRef]
- Li, B.; Gao, T.-M. Functional Role of Mitochondrial and Nuclear BK Channels. Int. Rev. Neurobiol. 2016, 128, 163–191. [Google Scholar] [CrossRef]
- Seidel, K.N.; Derst, C.; Salzmann, M.; Höltje, M.; Priller, J.; Markgraf, R.; Heinemann, S.H.; Heilmann, H.; Skatchkov, S.N.; Eaton, M.J.; et al. Expression of the voltage- and Ca2+-dependent BK potassium channel subunits BKβ1 and BKβ4 in rodent astrocytes. Glia 2011, 59, 893–902. [Google Scholar] [CrossRef]
- Seifert, G.; Henneberger, C.; Steinhäuser, C. Diversity of astrocyte potassium channels: An update. Brain Res. Bull. 2018, 136, 26–36. [Google Scholar] [CrossRef]
- Contet, C.; Goulding, S.P.; Kuljis, D.A.; Barth, A.L. BK channel sin the CNS. Int. Rev. Neurobiol. 2016, 128, 281–342. [Google Scholar]
- Deng, P.-Y.; Rotman, Z.; Blundon, J.A.; Cho, Y.; Cui, J.; Cavalli, V.; Zakharenko, S.; Klyachko, V.A. FMRP Regulates Neurotransmitter Release and Synaptic Information Transmission by Modulating Action Potential Duration via BK Channels. Neuron 2013, 77, 696–711. [Google Scholar] [CrossRef]
- Griguoli, M.; Sgritta, M.; Cherubini, E. Presynaptic BK channels control transmitter release: Physiological relevance and potential therapeutic implications. J. Physiol. 2016, 594, 3489–3500. [Google Scholar] [CrossRef]
- Sausbier, M.; Hu, H.; Arntz, C.; Feil, S.; Kamm, S.; Adelsberger, H.; Sausbier, U.; Sailer, C.A.; Feil, R.; Hofmann, F.; et al. Cerebellar ataxia and Purkinje cell dysfunction caused by Ca2+-activated K+channel deficiency. Proc. Natl. Acad. Sci. USA 2004, 101, 9474–9478. [Google Scholar] [CrossRef]
- Bock, T.; Stuart, G.J. The Impact of BK Channels on Cellular Excitability Depends on their Subcellular Location. Front. Cell. Neurosci. 2016, 10, 206. [Google Scholar] [CrossRef]
- Storm, J.F. Understanding the Brain Through the Hippocampus the Hippocampal Region as a Model for Studying Brain Structure and Function. Prog. Brain Res. 1990, 83, 161–187. [Google Scholar]
- Bielefeldt, K.; Jackson, M.B. A calcium-activated potassium channel causes frequency-dependent action-potential failures in a mammalian nerve terminal. J. Neurophysiol. 1993, 70, 284–298. [Google Scholar] [CrossRef]
- Goldberg, J.A.; Wilson, C.J. Control of spontaneous firing patterns by the selective coupling of calcium currents to calcium-activated potassium currents in striatal cholinergic interneurons. J. Neurosci. 2005, 25, 10230–10238. [Google Scholar] [CrossRef]
- Grimes, W.N.; Li, W.; Chávez, A.E.; Diamond, J.S. BK channels modulate pre- and postsynaptic signaling at reciprocal synapses in retina. Nat. Neurosci. 2009, 12, 585–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Shao, L.-R.; Chavoshy, S.; Gu, N.; Trieb, M.; Behrens, R.; Laake, P.; Pongs, O.; Knaus, H.G.; Ottersen, O.P.; et al. Presynaptic Ca2+-Activated K+Channels in Glutamatergic Hippocampal Terminals and Their Role in Spike Repolarization and Regulation of Transmitter Release. J. Neurosci. 2001, 21, 9585–9597. [Google Scholar] [CrossRef] [PubMed]
- Raffaelli, G.; Saviane, C.; Mohajerani, M.H.; Pedarzani, P.; Cherubini, E. BK potassium channels control transmitter release at CA3-CA3 synapses in the rat hippocampus. J. Physiol. 2004, 557, 147–157. [Google Scholar] [CrossRef]
- Tazerart, S.; Blanchard, M.G.; Miranda-Rottmann, S.; Mitchell, D.E.; Pina, B.N.; Thomas, C.I.; Kamasawa, N.; Araya, R. Selective activation of BK channels in small-headed dendritic spines suppresses excitatory postsynaptic potentials. J. Physiol. 2022, 600, 2165–2187. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-Y.; Liu, S.-B.; Cui, G.-B.; Ma, L.; Feng, B.; Xing, J.-H.; Yang, Q.; Li, X.-Q.; Wu, Y.-M.; Xiong, L.-Z.; et al. Acute stress induces down-regulation of large-conductance Ca2+-activated potassium channels in the lateral amygdala. J. Physiol. 2012, 590, 875–886. [Google Scholar] [CrossRef]
- Zhang, J.; Guan, X.; Li, Q.; Meredith, A.L.; Pan, H.-L.; Yan, J. Glutamate-activated BK channel complexes formed with NMDA receptors. Proc. Natl. Acad. Sci. USA 2018, 115, E9006–E9014. [Google Scholar] [CrossRef]
- Cavaccini, A.; Gritti, M.; Giorgi, A.; Locarno, A.; Heck, N.; Migliarini, S.; Bertero, A.; Mereu, M.; Margiani, G.; Trusel, M.; et al. Serotonergic Signaling Controls Input-Specific Synaptic Plasticity at Striatal Circuits. Neuron 2018, 98, 801–816.e7. [Google Scholar] [CrossRef]
- Xu, J.W.; Slaughter, M.M. Large-Conductance Calcium-Activated Potassium Channels Facilitate Transmitter Release in Salamander Rod Synapse. J. Neurosci. 2005, 25, 7660–7668. [Google Scholar] [CrossRef]
- Eisenman, G.; Latorre, R.; Miller, C. Multi-ion conduction and selectivity in the high-conductance Ca++-activated K+ channel from skeletal muscle. Biophys. J. 1986, 50, 1025–1034. [Google Scholar] [CrossRef]
- Latorre, R.; Miller, C. Conduction and selectivity in potassium channels. J. Membr. Biol. 1983, 71, 11–30. [Google Scholar] [CrossRef]
- Yellen, G. Ionic permeation and blockade in Ca2+-activated K+ channels of bovine chromaffin cells. J. Gen. Physiol. 1984, 84, 157–186. [Google Scholar] [CrossRef]
- Adelman, J.P.; Shen, K.-Z.; Kavanaugh, M.P.; Warren, R.A.; Wu, Y.-N.; Lagrutta, A.; Bond, C.T.; North, R.A. Calcium-activated potassium channels expressed from cloned complementary DNAs. Neuron 1992, 9, 209–216. [Google Scholar] [CrossRef]
- Butler, A.; Tsunoda, S.; McCobb, D.P.; Wei, A.; Salkoff, L. mSlo, a complex mouse gene encoding “maxi” calcium-activated potassium channels. Science 1993, 261, 221–224. [Google Scholar] [CrossRef]
- Meera, P.; Wallner, M.; Song, M.; Toro, L. Large conductance voltage- and calcium-dependent K+ channel, a distinct member of voltage-dependent ion channels with seven N-terminal transmembrane segments (SO-S6), an extracellular N terminus, and an intracellular (S9-S10) C terminus. Proc. Natl. Acad. Sci. USA 1997, 94, 14066–14071. [Google Scholar] [CrossRef]
- Bao, L.; Rapin, A.M.; Holmstrand, E.C.; Cox, D.H. Elimination of the BKCa Channel’s High-Affinity Ca2+ Sensitivity. J. Gen. Physiol. 2002, 120, 173–189. [Google Scholar] [CrossRef]
- Salkoff, L.; Butler, A.; Ferreira, G.; Santi, C.; Wei, A. High-conductance potassium channels of the SLO family. Nat. Rev. Neurosci. 2006, 7, 921–931. [Google Scholar] [CrossRef]
- Tao, X.; Hite, R.K.; MacKinnon, R. Cryo-EM structure of the open high-conductance Ca2+-activated K+ channel. Nature 2016, 541, 46–51. [Google Scholar] [CrossRef]
- Xia, X.-M.; Zeng, X.; Lingle, C.J. Multiple regulatory sites in large-conductance calcium-activated potassium channels. Nature 2002, 418, 880–884. [Google Scholar] [CrossRef]
- McManus, O.B.; Blatz, A.L.; Magleby, K.L. Inverse relationship of the durations of adjacent open and shut intervals for Cl and K channels. Nature 1985, 317, 625–627. [Google Scholar] [CrossRef]
- Pallotta, B.S. Calcium-activated potassium channels in rat muscle inactivate from a short-duration open state. J. Physiol. 1985, 363, 501–516. [Google Scholar] [CrossRef] [PubMed]
- Rothberg, B.S.; Magleby, K.L. Voltage and Ca2+ Activation of Single Large-Conductance Ca2+-Activated K+ Channels Described by a Two-Tiered Allosteric Gating Mechanism. J. Gen. Physiol. 2000, 116, 75–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horrigan, F.T.; Aldrich, R.W. Coupling between Voltage Sensor Activation, Ca2+ Binding and Channel Opening in Large Conductance (BK) Potassium Channels. J. Gen. Physiol. 2002, 120, 267–305. [Google Scholar] [CrossRef] [PubMed]
- Carrasquel-Ursulaez, W.; Contreras, G.F.; Sepúlveda, R.V.; Aguayo, D.; González-Nilo, F.; González, C.; Latorre, R. Hydrophobic interaction between contiguous residues in the S6 transmembrane segment acts as a stimuli integration node in the BK channel. J. Gen. Physiol. 2014, 145, 61–74. [Google Scholar] [CrossRef]
- Lorenzo-Ceballos, Y.; Carrasquel-Ursulaez, W.; Castillo, K.; Alvarez, O.; Latorre, R. Calcium-driven regulation of voltagesensingdomains in BK channels. Elife 2019, 8, 1–24. [Google Scholar] [CrossRef]
- Hite, R.K.; Tao, X.; MacKinnon, R. Structural basis for gating the high-conductance Ca2+-activated K+ channel. Nature 2016, 541, 52–57. [Google Scholar] [CrossRef]
- Ma, Z.; Lou, X.J.; Horrigan, F.T. Role of Charged Residues in the S1–S4 Voltage Sensor of BK Channels. J. Gen. Physiol. 2006, 127, 309–328. [Google Scholar] [CrossRef]
- Carrasquel-Ursulaez, W.; Segura, I.; Díaz-Franulic, I.; Echeverría, F.; Lorenzo-Ceballos, Y.; Espinoza, N.; Rojas, M.; Garate, J.A.; Perozo, E.; Alvarez, O. Mechanism of voltage sensing in Ca2+- and voltage-activated K+ (BK) channels. Proc. Natl. Acad. Sci. USA 2022, 119, 1–11. [Google Scholar] [CrossRef]
- Zeng, X.-H.; Xia, X.-M.; Lingle, C.J. Divalent Cation Sensitivity of BK Channel Activation Supports the Existence of Three Distinct Binding Sites. J. Gen. Physiol. 2005, 125, 273–286. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, X.-H.; Lingle, C.J. Barium ions selectively activate BK channels via the Ca 2+ -bowl site. Proc. Natl. Acad. Sci. USA 2012, 109, 11413–11418. [Google Scholar] [CrossRef]
- Lee, C.-H.; MacKinnon, R. Structures of the Human HCN1 Hyperpolarization-Activated Channel. Cell 2017, 168, 111–120.e11. [Google Scholar] [CrossRef]
- Wang, W.; MacKinnon, R. Cryo-EM Structure of the Open Human Ether-à-go-go -Related K + Channel hERG. Cell 2017, 169, 422–430.e10. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.-Y.; Zeng, X.-H.; Lingle, C.J. Closed-channel block of BK potassium channels by bbTBA requires partial activation. J. Gen. Physiol. 2009, 134, 409–436. [Google Scholar] [CrossRef]
- Thompson, J.; Begenisich, T. Selectivity filter gating in large-conductance Ca2+-activated K+ channels. J. Gen. Physiol. 2012, 139, 235–244. [Google Scholar] [CrossRef]
- Wilkens, C.M.; Aldrich, R.W. State-independent Block of BK Channels by an Intracellular Quaternary Ammonium. J. Gen. Physiol. 2006, 128, 347–364. [Google Scholar] [CrossRef]
- Jia, Z.; Yazdani, M.; Zhang, G.; Cui, J.; Chen, J. Hydrophobic gating in BK channels. Nat. Commun. 2018, 9, 3408. [Google Scholar] [CrossRef]
- Torres, Y.P.; Granados, S.T.; Latorre, R. Pharmacological consequences of the coexpression of BK channel α and auxiliary β subunits. Front Physiol. 2014, 5, 383. [Google Scholar] [CrossRef]
- Orio, P.; Rojas, P.; Ferreira, G.; Latorre, R. New Disguises for an Old Channel: MaxiK Channel β-Subunits. Physiology 2002, 17, 156–161. [Google Scholar] [CrossRef]
- Zhou, Y.; Lingle, C.J. Paxilline inhibits BK channels by an almost exclusively closed-channel block mechanism. J. Gen. Physiol. 2014, 144, 415–440. [Google Scholar] [CrossRef]
- Meera, P.; Wallner, M.; Toro, L. A neuronal β subunit (KCNMB4) makes the large conductance, voltage- and Ca2+ -activated K + channel resistant to charybdotoxin and iberiotoxin. Proc. Natl. Acad. Sci. USA 2000, 97, 5562–5567. [Google Scholar] [CrossRef]
- Ha, T.S.; Heo, M.-S.; Park, C.-S. Functional Effects of Auxiliary β4-Subunit on Rat Large-Conductance Ca2+-Activated K+ Channel. Biophys. J. 2004, 86, 2871–2882. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Rothberg, B.S.; Brenner, R. Mechanism of β4 Subunit Modulation of BK Channels. J. Gen. Physiol. 2006, 127, 449–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffe, D.; Wang, B.; Brenner, R. Shaping of action potentials by type I and type II large-conductance Ca2+-activated K+ channels. Neuroscience 2011, 192, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jaffe, D.B.; Brenner, R. Current understanding of iberiotoxin-resistant BK channels in the nervous system. Front. Physiol. 2014, 5, 382. [Google Scholar] [CrossRef]
- Reinhart, P.H.; Chung, S.; Levitan, I.B. A family of calcium-dependent potassium channels from rat brain. Neuron 1989, 2, 1031–1041. [Google Scholar] [CrossRef]
- Behrens, R.; Nolting, A.; Reimann, F.; Schwarz, M.; Waldschütz, R.; Pongs, O. hKCNMB3 and hKCNMB4, cloning and characterization of two members of the large-conductance calcium-activated potassium channel β subunit family. FEBS Lett. 2000, 474, 99–106. [Google Scholar] [CrossRef]
- Lippiat, J.D.; Standen, N.B.; Harrow, I.D.; Phillips, S.C.; Davies, N.W. Properties of BKCa channels formed by bicistronic expression of hSloα and β1-4 subunits in HEK293 cells. J. Membr. Biol. 2003, 192, 141–148. [Google Scholar]
- Gan, G.; Yi, H.; Chen, M.; Sun, L.; Li, W.; Wu, Y.; Ding, J. Structural Basis for Toxin Resistance of β4-Associated Calcium-activated Potassium (BK) Channels. J. Biol. Chem. 2008, 283, 24177–24184. [Google Scholar] [CrossRef]
- Tao, X.; Mackinnon, R. Molecular structures of the human slo1 k+ channel in complex with b4. Elife 2019, 8, e51409. [Google Scholar] [CrossRef]
- Candia, S.; Garcia, M.; Latorre, R. Mode of action of iberiotoxin, a potent blocker of the large conductance Ca(2+)-activated K+ channel. Biophys. J. 1992, 63, 583–590. [Google Scholar] [CrossRef]
- Weiger, T.M.; Holmqvist, M.H.; Levitan, I.B.; Clark, F.T.; Sprague, S.; Huang, W.-J.; Ge, P.; Wang, C.; Lawson, D.; Jurman, M.E.; et al. A Novel Nervous System β Subunit that Downregulates Human Large Conductance Calcium-Dependent Potassium Channels. J. Neurosci. 2000, 20, 3563–3570. [Google Scholar] [CrossRef]
- Brenner, R.; Chen, Q.H.; Vilaythong, A.; Toney, G.M.; Noebels, J.; Aldrich, R.W. BK channel β4 subunit reduces dentate gyrus excitability and protects against temporal lobe seizures. Nat. Neurosci. 2005, 8, 1752–1759. [Google Scholar] [CrossRef]
- Zhao, H.; Xue, Q.; Li, C.; Wang, Q.; Han, S.; Zhou, Y.; Yang, T.; Xie, Y.; Fu, H.; Lu, C.; et al. Upregulation of Beta4 subunit of BKCa channels in the anterior cingulate cortex contributes to mechanical allodynia associated anxiety-like behaviors. Mol. Brain 2020, 13, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Piwonska, M.; Wilczek, E.; Szewczyk, A.; Wilczynski, G. Differential distribution of Ca2+-activated potassium channel β4 subunit in rat brain: Immunolocalization in neuronal mitochondria. Neuroscience 2008, 153, 446–460. [Google Scholar] [CrossRef]
- Benton, M.D.; Lewis, A.H.; Bant, J.; Raman, I.M. Iberiotoxin-sensitive and -insensitive BK currents in Purkinje neuron somata. J. Neurophysiol. 2013, 109, 2528–2541. [Google Scholar] [CrossRef]
- Jiang, Z.; Wallner, M.; Meera, P.; Toro, L. Human and Rodent MaxiK Channel β-Subunit Genes: Cloning and Characterization. Genomics 1999, 55, 57–67. [Google Scholar] [CrossRef]
- Salzmann, M.; Seidel, K.N.; Bernard, R.; Prüss, H.; Veh, R.W.; Derst, C. BKβ1 Subunits Contribute to BK Channel Diversity in Rat Hypothalamic Neurons. Cell. Mol. Neurobiol. 2010, 30, 967–976. [Google Scholar] [CrossRef]
- Tseng-Crank, J.; Godinot, N.; Johansen, T.E.; Ahring, P.K.; Strøbæk, D.; Mertz, R.; Foster, C.D.; Olesen, S.P.; Reinhart, P.H. Cloning, expression, and distribution of a Ca2+-activated K+ channel β-subunit from human brain. Proc. Natl. Acad. Sci. USA 1996, 93, 9200–9205. [Google Scholar] [CrossRef]
- Wynne, P.M.; Puig, S.I.; Martin, G.E.; Treistman, S.N. Compartmentalized β subunit distribution determines characteristics and ethanol sensitivity of somatic, dendritic, and terminal large-conductance calcium-activated potassium channels in the rat central nervous system. J. Pharmacol. Exp. Ther. 2009, 329, 978–986. [Google Scholar] [CrossRef]
- Deng, P.-Y.; Klyachko, V.A. Genetic upregulation of BK channel activity normalizes multiple synaptic and circuit defects in a mouse model of fragile X syndrome. J. Physiol. 2015, 594, 83–97. [Google Scholar] [CrossRef]
- Kshatri, A.; Cerrada, A.; Gimeno, R.; Bartolomé-Martín, D.; Rojas, P.; Giraldez, T. Differential regulation of BK channels by fragile X mental retardation protein. J. Gen. Physiol. 2020, 152, e201912502. [Google Scholar] [CrossRef] [PubMed]
- Pyott, S.; Glowatzki, E.; Trimmer, J.S.; Aldrich, R.W. Extrasynaptic Localization of Inactivating Calcium-Activated Potassium Channels in Mouse Inner Hair Cells. J. Neurosci. 2004, 24, 9469–9474. [Google Scholar] [CrossRef] [PubMed]
- Faber, E.S.L.; Sah, P. Ca2+ -activated K+ (BK) channel inactivation contributes to spike broadening during repetitive firing in the rat lateral amygdala. J Physiol. 2003, 552, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Hicks, G.A.; Marrion, N.V. Ca2+-dependent inactivation of large conductance Ca2+-activated K+ (BK) channels in rat hippocampal neurones produced by pore block from an associated particle. J. Physiol. 1998, 508, 721–734. [Google Scholar] [CrossRef]
- Li, W.; Gao, S.-B.; Lv, C.-X.; Wu, Y.; Guo, Z.-H.; Ding, J.-P.; Xu, T. Characterization of voltage-and Ca2+-activated K+ channels in rat dorsal root ganglion neurons. J. Cell. Physiol. 2007, 212, 348–357. [Google Scholar] [CrossRef]
- Whitt, J.P.; Montgomery, J.R.; Meredith, A.L. BK channel inactivation gates daytime excitability in the circadian clock. Nat. Commun. 2016, 7, 10837. [Google Scholar] [CrossRef]
- Meredith, A.L.; Wiler, S.W.; Miller, B.H.; Takahashi, J.S.; Fodor, A.A.; Ruby, N.F.; Aldrich, R.W. BK calcium-activated potassium channels regulate circadian behavioral rhythms and pacemaker output. Nat. Neurosci. 2006, 9, 1041–1049. [Google Scholar] [CrossRef]
- Pitts, G.R.; Ohta, H.; McMahon, D.G. Daily rhythmicity of large-conductance Ca2+-activated K+ currents in suprachiasmatic nucleus neurons. Brain Res. 2006, 1071, 54–62. [Google Scholar] [CrossRef]
- Berkefeld, H.; Fakler, B.; Schulte, U. Ca2+-activated K+ channels: From protein complexes to function. Physiol. Rev. 2010, 90, 1437–1459. [Google Scholar] [CrossRef]
- Shah, K.R.; Guan, X.; Yan, J. Structural and Functional Coupling of Calcium-Activated BK Channels and Calcium-Permeable Channels Within Nanodomain Signaling Complexes. Front. Physiol. 2022, 12, 2556. [Google Scholar] [CrossRef]
- Sweet, T.B.; Cox, D.H. Measurements of the BK Ca channel’s high-affinity Ca 2+ binding constants: Effects of membrane voltage. J. Gen. Physiol. 2008, 132, 491–505. [Google Scholar] [CrossRef]
- Fakler, B.; Adelman, J.P. Control of KCa Channels by Calcium Nano/Microdomains. Neuron 2008, 59, 873–881. [Google Scholar] [CrossRef]
- Naraghi, M.; Neher, E. Linearized Buffered Ca2+Diffusion in Microdomains and Its Implications for Calculation of [Ca2+] at the Mouth of a Calcium Channel. J. Neurosci. 1997, 17, 6961–6973. [Google Scholar] [CrossRef] [Green Version]
- Bellono, N.W.; Leitch, D.B.; Julius, D. Molecular basis of ancestral vertebrate electroreception. Nature 2017, 543, 391–396. [Google Scholar] [CrossRef]
- Berkefeld, H.; Fakler, B. Repolarizing Responses of BKCa-Cav Complexes Are Distinctly Shaped by Their Cav Subunits. J. Neurosci. 2008, 28, 8238–8245. [Google Scholar] [CrossRef]
- Berkefeld, H.; Sailer, C.A.; Bildl, W.; Rohde, V.; Thumfart, J.O.; Eble, S.; Klugbauer, N.; Reisinger, E.; Bischofberger, J.; Oliver, D.; et al. BKCa-Cav channel complexes mediate rapid and localized Ca 2+-activated K+ signaling. Science 2006, 314, 615–620. [Google Scholar] [CrossRef]
- Marrion, N.V.; Tavalin, S.J. Selective activation of Ca2+-activated K+ channels by co-localized Ca2+ channels in hippocampal neurons. Nature 1998, 395, 900–905. [Google Scholar] [CrossRef]
- Rehak, R.; Bartoletti, T.M.; Engbers, J.D.T.; Berecki, G.; Turner, R.W.; Zamponi, G.W. Low Voltage Activation of KCa1.1 Current by Cav3-KCa1.1 Complexes. PLoS ONE 2013, 8, e61844. [Google Scholar] [CrossRef]
- Müller, A.; Kukley, M.; Uebachs, M.; Beck, H.; Dietrich, D. Nanodomains of single Ca2+ channels contribute to action potential repolarization in cortical neurons. J. Neurosci. 2007, 27, 483–495. [Google Scholar] [CrossRef]
- Gola, M.; Crest, M. Colocalization of active KCa channels and Ca2+ channels within Ca2+ Domains in helix neurons. Neuron 1993, 10, 689–699. [Google Scholar] [CrossRef]
- Monreal, M.R.; Pérez, J.Q.; Escalera, M.P.; Lazalde, A.R.; Bonilla, M.E.P. Development of the L-type CaV/BK Complex Simulator (II): Estimation of the distance between the channels. South Florida J. Dev. 2021, 2, 1258–1265. [Google Scholar] [CrossRef]
- Edgerton, J.R.; Reinhart, P.H. Distinct contributions of small and large conductance Ca2+- activated K+ channels to rat Purkinje neuron function. J. Physiol. 2003, 548, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Gutzmann, J.J.; Lin, L.; Hoffman, D.A. Functional Coupling of Cav2.3 and BK Potassium Channels Regulates Action Potential Repolarization and Short-Term Plasticity in the Mouse Hippocampus. Front. Cell. Neurosci. 2019, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Vivas, O.; Moreno, C.M.; Santana, L.F.; Hille, B. Proximal clustering between BK and CaV1.3 channels promotes functional coupling and BK channel activation at low voltage. Elife 2017, 6, e28029. [Google Scholar] [CrossRef]
- Smith, M.R.; Nelson, A.B.; du Lac, S. Regulation of Firing Response Gain by Calcium-Dependent Mechanisms in Vestibular Nucleus Neurons. J. Neurophysiol. 2002, 87, 2031–2042. [Google Scholar] [CrossRef]
- Loane, D.J.; Hicks, G.A.; Perrino, B.A.; Marrion, N.V. Inhibition of BKCa channel activity by association with calcineurin in rat brain. Eur. J. Neurosci. 2006, 24, 433–441. [Google Scholar] [CrossRef]
- Sun, X.; Gu, X.Q.; Haddad, G.G. Calcium Influx via L- and N-Type Calcium Channels Activates a Transient Large-Conductance Ca2+-Activated K+Current in Mouse Neocortical Pyramidal Neurons. J. Neurosci. 2003, 23, 3639–3648. [Google Scholar] [CrossRef]
- Plante, A.E.; Whitt, J.P.; Meredith, A.L. BK channel activation by L-type Ca2+ channels CaV1.2 and CaV1.3 during the subthreshold phase of an action potential. J. Neurophysiol. 2021, 126, 427–439. [Google Scholar] [CrossRef]
- Davies, P.J.; Ireland, D.R.; McLachlan, E.M. Sources of Ca2+ for different Ca2+-activated K+ conductances in neurones of the rat superior cervical ganglion. J. Physiol. 1996, 495, 353–366. [Google Scholar] [CrossRef]
- Womack, M.D.; Chevez, C.; Khodakhah, K. Calcium-Activated Potassium Channels Are Selectively Coupled to P/Q-Type Calcium Channels in Cerebellar Purkinje Neurons. J. Neurosci. 2004, 24, 8818–8822. [Google Scholar] [CrossRef]
- Isaacson, J.S.; Murphy, G. Glutamate-Mediated Extrasynaptic Inhibition: Direct Coupling of NMDA Receptors to Ca2+-Activated K+ Channels. Neuron 2001, 31, 1027–1034. [Google Scholar] [CrossRef]
- Gómez, R.; Maglio, L.E.; Gonzalez-Hernandez, A.J.; Rivero-Pérez, B.; Bartolomé-Martín, D.; Giraldez, T. NMDA receptor–BK channel coupling regulates synaptic plasticity in the barrel cortex. Proc. Natl. Acad. Sci. USA 2021, 118, e2107026118. [Google Scholar] [CrossRef]
- Irie, T.; Trussell, L.O. Double-Nanodomain Coupling of Calcium Channels, Ryanodine Receptors, and BK Channels Controls the Generation of Burst Firing. Neuron 2017, 96, 856–870.e4. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Bugay, V.; Ling, L.; Chuang, H.H.; Jaffe, D.B.; Brenner, R. Knockout of the BK β4-subunit promotes a functional coupling of BK channels and ryanodine receptors that mediate a fAHP-induced increase in excitability. J. Neurophysiol. 2016, 116, 456–465. [Google Scholar] [CrossRef]
- Whitt, J.P.; McNally, B.A.; Meredith, A.L. Differential contribution of Ca2+ sources to day and night BK current activation in the circadian clock. J. Gen. Physiol. 2017, 150, 259–275. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, Y.; Hou, P.; Yan, Z.; Kong, W.; Liu, B.; Li, X.; Yao, J.; Zhang, Y.; Qin, F.; et al. TRPV1 Channels Are Functionally Coupled with BK(mSlo1) Channels in Rat Dorsal Root Ganglion (DRG) Neurons. PLoS ONE 2013, 8, e78203. [Google Scholar] [CrossRef]
- Lin, M.T.; Hessinger, D.A.; Pearce, W.J.; Longo, L.D. Modulation of BK channel calcium affinity by differential phosphorylation in developing ovine basilar artery myocytes. Am. J. Physiol. Circ. Physiol. 2006, 291, H732–H740. [Google Scholar] [CrossRef]
- Yan, J.; Olsen, J.V.; Park, K.-S.; Li, W.; Bildl, W.; Schulte, U.; Aldrich, R.W.; Fakler, B.; Trimmer, J.S. Profiling the Phospho-status of the BKCa Channel α Subunit in Rat Brain Reveals Unexpected Patterns and Complexity. Mol. Cell. Proteom. 2008, 7, 2188–2198. [Google Scholar] [CrossRef]
- Wang, Z.-W. Regulation of Synaptic Transmission by Presynaptic CaMKII and BK Channels. Mol. Neurobiol. 2008, 38, 153–166. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Y.; Wen, H.; Levitan, I.B. Simultaneous Binding of Two Protein Kinases to a Calcium-Dependent Potassium Channel. J. Neurosci. 1999, 19, RC4. [Google Scholar] [CrossRef]
- Lin, M.T.; Longo, L.D.; Pearce, W.J.; Hessinger, D.A. Ca2+-activated K+ channel-associated phosphatase and kinase activities during development. Am. J. Physiol.-Hear Circ. Physiol. 2005, 289, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Shelley, C.; Whitt, J.P.; Montgomery, J.R.; Meredith, A.L. Phosphorylation of a constitutive serine inhibits BK channel variants containing the alternate exon “SRKR”. J. Gen. Physiol. 2013, 142, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Weiger, T.M.; Wu, Y.; Levitan, I.B. Phosphorylation-dependent Functional Coupling of hSlo Calcium-dependent Potassium Channel and Its hβ4 Subunit. J. Biol. Chem. 2002, 277, 10014–10020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, A.B.; Gittis, A.; du Lac, S. Decreases in CaMKII Activity Trigger Persistent Potentiation of Intrinsic Excitability in Spontaneously Firing Vestibular Nucleus Neurons. Neuron 2005, 46, 623–631. [Google Scholar] [CrossRef]
- van Welie, I.; du Lac, S. Bidirectional control of BK channel open probability by CAMKII and PKC in medial vestibular nucleus neurons. J. Neurophysiol. 2011, 105, 1651–1659. [Google Scholar] [CrossRef]
- Widmer, H.A.; Rowe, I.C.M.; Shipston, M.J.; D’Antona, G.; Pellegrino, M.A.; Adami, R.; Rossi, R.; Carlizzi, C.N.; Canepari, M.; Saltin, B.; et al. Conditional protein phosphorylation regulates BK channel activity in rat cerebellar Purkinje neurons. J. Physiol. 2003, 552, 379–391. [Google Scholar] [CrossRef]
- Watanave, M.; Takahashi, N.; Hosoi, N.; Konno, A.; Yamamoto, H.; Yasui, H.; Kawachi, M.; Horii, T.; Matsuzaki, Y.; Hatada, I.; et al. Protein kinase Cγ in cerebellar Purkinje cells regulates Ca2+-activated large-conductance K+ channels and motor coordination. Proc. Natl. Acad. Sci. USA 2022, 119, 1–11. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, B.; Ge, Q.; Wang, Z.W. Presynaptic Ca2+/calmodulin-dependent protein kinase II modulates neurotransmitter release by activating BK channels at Caenorhabditis elegans neuromuscular junction. J. Neurosci. 2007, 27, 10404–10413. [Google Scholar] [CrossRef]
- Zaman, T.; De Oliveira, C.; Smoka, M.; Narla, C.; Poulter, M.O.; Schmid, S. BK Channels Mediate Synaptic Plasticity Underlying Habituation in Rats. J. Neurosci. 2017, 37, 4540–4551. [Google Scholar] [CrossRef]
- Sansom, S.C.; Stockand, J.D.; Hall, D.; Williams, B. Regulation of Large Calcium-activated Potassium Channels by Protein Phosphatase 2A. J. Biol. Chem. 1997, 272, 9902–9906. [Google Scholar] [CrossRef]
- Tian, L.; Knaus, H.-G.; Shipston, M. Glucocorticoid Regulation of Calcium-activated Potassium Channels Mediated by Serine/Threonine Protein Phosphatase. J. Biol. Chem. 1998, 273, 13531–13536. [Google Scholar] [CrossRef]
- Zhou, X.-B.; Ruth, P.; Schlossmann, J.; Hofmann, F.; Korth, M. Protein Phosphatase 2A Is Essential for the Activation of Ca2+-activated K+ Currents by cGMP-dependent Protein Kinase in Tracheal Smooth Muscle and Chinese Hamster Ovary Cells. J. Biol. Chem. 1996, 271, 19760–19767. [Google Scholar] [CrossRef]
- Petrik, D.; Wang, B.; Brenner, R. Modulation by the BK accessory β4 subunit of phosphorylation-dependent changes in excitability of dentate gyrus granule neurons. Eur. J. Neurosci. 2011, 34, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Latorre, J.A. Functional Upregulation of Ca2+ -Activated K+ Channels in the Development of Substantia Nigra Dopamine Neurons. PLoS ONE 2012, 7, e51610. [Google Scholar] [CrossRef]
- Kang, J.; Huguenard, J.R.; Prince, D.A. Development of BK channels in neocortical pyramidal neurons. J. Neurophysiol. 1996, 76, 188–198. [Google Scholar] [CrossRef]
- Kros, C.J.; Ruppersberg, J.P.; Rüsch, A. Expression of a potassium current in inner hair cells during development of hearing in mice. Nature 1998, 394, 281–284. [Google Scholar] [CrossRef]
- Hunsberger, M.S.; Mynlieff, M. BK potassium currents contribute differently to action potential waveform and firing rate as rat hippocampal neurons mature in the first postnatal week. J. Neurophysiol. 2020, 124, 703–714. [Google Scholar] [CrossRef]
- Niday, Z.; Bean, B.P. BK Channel Regulation of Afterpotentials and Burst Firing in Cerebellar Purkinje Neurons. J. Neurosci. 2021, 41, 2854–2869. [Google Scholar] [CrossRef]
- Kaufmann, W.; Kasugai, Y.; Ferraguti, F.; Storm, J. Two distinct pools of large-conductance calcium-activated potassium channels in the somatic plasma membrane of central principal neurons. Neuroscience 2010, 169, 974–986. [Google Scholar] [CrossRef]
- Womack, M.D.; Khodakhah, K. Characterization of large conductance Ca2+-activated K+ channels in cerebellar Purkinje neurons. Eur. J. Neurosci. 2002, 16, 1214–1222. [Google Scholar] [CrossRef]
- Sánchez-Aguilera, A.; Monedero, G.; Colino, A.; Vicente-Torres, M. Development of Action Potential Waveform in Hippocampal CA1 Pyramidal Neurons. Neuroscience 2020, 442, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Gu, N.; Vervaeke, K.; Storm, J.F. BK potassium channels facilitate high-frequency firing and cause early spike frequency adaptation in rat CA1 hippocampal pyramidal cells. J. Physiol. 2007, 580, 859–882. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.R.; Halvorsrud, R.; Borg-Graham, L.; Storm, J.F. The role of BK-type Ca2+-dependent K+ channels in spike broadening during repetitive firing in rat hippocampal pyramidal cells. J. Physiol. 1999, 521, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Benhassine, N.; Berger, T. Large-conductance calcium-dependent potassium channels prevent dendritic excitability in neocortical pyramidal neurons. Pflügers Arch.-Eur. J. Physiol. 2008, 457, 1133–1145. [Google Scholar] [CrossRef]
- Kurowski, P.; Grzelka, K.; Szulczyk, P. Ionic Mechanism Underlying Rebound Depolarization in Medial Prefrontal Cortex Pyramidal Neurons. Front. Cell. Neurosci. 2018, 12, 93. [Google Scholar] [CrossRef]
- Galarraga, E.; Vilchis, C.; Tkatch, T.; Salgado, H.; Tecuapetla, F.; Perez-Rosello, T.; Perez-Garci, E.; Hernandez-Echeagaray, E.; Surmeier, D.; Bargas, J. Somatostatinergic modulation of firing pattern and calcium-activated potassium currents in medium spiny neostriatal neurons. Neuroscience 2007, 146, 537–554. [Google Scholar] [CrossRef]
- Vilchis, C.; Bargas, J.; Ayala, G.; Galván, E.; Galarraga, E. Ca2+ channels that activate Ca2+-dependent K+ currents in neostriatal neurons. Neuroscience 1999, 95, 745–752. [Google Scholar] [CrossRef]
- Wang, H.; Ardiles, A.O.; Yang, S.; Tran, T.; Posada-Duque, R.; Valdivia, G.; Baek, M.; Chuang, Y.-A.; Palacios, A.G.; Gallagher, M.; et al. Metabotropic Glutamate Receptors Induce a Form of LTP Controlled by Translation and Arc Signaling in the Hippocampus. J. Neurosci. 2016, 36, 1723–1729. [Google Scholar] [CrossRef]
- Kohashi, T.; Carlson, B.A. A fast BK-type KCa current acts as a postsynaptic modulator of temporal selectivity for communication signals. Front. Cell. Neurosci. 2014, 8, 286. [Google Scholar] [CrossRef]
- Klyachko, V.A.; Ahern, G.P.; Jackson, M.B. cGMP-mediated facilitation in nerve terminals by enhancement of the spike afterhyperpolarization. Neuron 2001, 31, 1015–1025. [Google Scholar] [CrossRef]
- Zhang, Y.; Bonnan, A.; Bony, G.; Ferezou, I.; Pietropaolo, S.; Ginger, M.; Sans, N.; Rossier, J.; Oostra, B.; LeMasson, G.; et al. Dendritic channelopathies contribute to neocortical and sensory hyperexcitability in Fmr1-/y mice. Nat. Neurosci. 2014, 17, 1701–1709. [Google Scholar] [CrossRef]
- Roshchin, M.V.; Matlashov, M.E.; Ierusalimsky, V.N.; Balaban, P.M.; Belousov, V.V.; Kemenes, G.; Staras, K.; Nikitin, E.S. A BK channel–mediated feedback pathway links single-synapse activity with action potential sharpening in repetitive firing. Sci. Adv. 2018, 4, 1–9. [Google Scholar] [CrossRef]
- Książek, A.; Ładno, W.; Szulczyk, B.P.; Grzelka, K.; Szulczyk, P.J. Properties of BK-type Ca++-dependent K+ channel currents in medial prefrontal cortex pyramidal neurons in rats of different ages. Front. Cell. Neurosci. 2013, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Benhassine, N.; Berger, T. Homogeneous distribution of large-conductance calcium-dependent potassium channels on soma and apical dendrite of rat neocortical layer 5 pyramidal neurons. Eur. J. Neurosci. 2005, 21, 914–926. [Google Scholar] [CrossRef]
- Dong, P.; Zhang, Y.; Hunanyan, A.S.; Mikati, M.A.; Cui, J.; Yang, H. Neuronal mechanism of a BK channelopathy in absence epilepsy and dyskinesia. Proc. Natl. Acad. Sci. USA 2022, 119, e2200140119. [Google Scholar] [CrossRef]
- Grewe, B.F.; Bonnan, A.; Frick, A. Back-propagation of physiological action potential output in dendrites of slender-tufted L5A pyramidal neurons. Front. Cell. Neurosci. 2010, 4, 13. [Google Scholar] [CrossRef]
- Sailer, C.A.; Kaufmann, W.A.; Kogler, M.; Chen, L.; Sausbier, U.; Ottersen, O.P.; Ruth, P.; Shipston, M.J.; Knaus, H.-G. Immunolocalization of BK channels in hippocampal pyramidal neurons. Eur. J. Neurosci. 2006, 24, 442–454. [Google Scholar] [CrossRef]
- Golding, N.L.; Jung, H.-Y.; Mickus, T.; Spruston, N. Dendritic Calcium Spike Initiation and Repolarization Are Controlled by Distinct Potassium Channel Subtypes in CA1 Pyramidal Neurons. J. Neurosci. 1999, 19, 8789–8798. [Google Scholar] [CrossRef]
- Shanley, L.J.; Irving, A.J.; Rae, M.G.; Ashford, M.L.; Harvey, J. Leptin inhibits rat hippocampal neurons via activation of large conductance calcium-activated K+ channels. Nat. Neurosci. 2002, 5, 299–300. [Google Scholar] [CrossRef]
- Springer, S.J.; Burkett, B.J.; Schrader, L.A. Modulation of BK channels contributes to activity-dependent increase of excitability through MTORC1 activity in CA1 pyramidal cells of mouse hippocampus. Front. Cell. Neurosci. 2015, 8, 451. [Google Scholar] [CrossRef]
- Misonou, H.; Menegola, M.; Buchwalder, L.; Park, E.W.; Meredith, A.; Rhodes, K.J.; Aldrich, R.W.; Trimmer, J.S. Immunolocalization of the Ca2+-activated K+channel Slo1 in axons and nerve terminals of mammalian brain and cultured neurons. J. Comp. Neurol. 2006, 496, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Shruti, S.; Urban-Ciecko, J.; Fitzpatrick, J.A.; Brenner, R.; Bruchez, M.P.; Barth, A.L. The brain-specific beta4 subunit downregulates BK channel cell surface expression. PLoS ONE 2012, 7, e33429. [Google Scholar] [CrossRef]
- Jalini, S.; Ye, H.; Tonkikh, A.A.; Charlton, M.P.; Carlen, P.L. Raised Intracellular Calcium Contributes to Ischemia-Induced Depression of Evoked Synaptic Transmission. PLoS ONE 2016, 11, e0148110. [Google Scholar] [CrossRef] [PubMed]
- Alle, H.; Kubota, H.; Geiger, J.R.P. Sparse But Highly Efficient Kv3 Outpace BKCa Channels in Action Potential Repolarization at Hippocampal Mossy Fiber Boutons. J. Neurosci. 2011, 31, 8001–8012. [Google Scholar] [CrossRef]
- Wang, D.; Wang, X.; Liu, P.; Jing, S.; Du, H.; Zhang, L.; Jia, F.; Li, A. Serotonergic afferents from the dorsal raphe decrease the excitability of pyramidal neurons in the anterior piriform cortex. Proc. Natl. Acad. Sci. USA 2020, 117, 3239–3247. [Google Scholar] [CrossRef]
- Casale, A.E.; Foust, A.J.; Bal, T.; McCormick, D.A. Cortical Interneuron Subtypes Vary in Their Axonal Action Potential Properties. J. Neurosci. 2015, 35, 15555–15567. [Google Scholar] [CrossRef]
- Ji, J.J.; Chen, L.; Duan, X.; Song, X.; Su, W.; Zhang, P.; Li, L.; Bai, S.; Sun, Y.; Inagaki, N. BK Channels Reveal Novel Phosphate Sensitivity in SNr Neurons. PLoS ONE 2012, 7, e52148. [Google Scholar] [CrossRef]
- Authement, M.E.; Langlois, L.D.; Shepard, R.D.; Browne, C.A.; Lucki, I.; Kassis, H.; Nugent, F.S. A role for corticotropin-releasing factor signaling in the lateral habenula and its modulation by early-life stress. Sci. Signal. 2018, 11, eaan6480. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Han, X.; Liu, Y.; Chen, J.; Hua, L.; Ma, Q.; Huang, Y.Y.X.; Tang, Q.Y.; Zhang, Z. +MRNA expression of LRRC55 protein (leucine-rich repeat-containing protein 55) in the adult mouse brain. PLoS ONE 2018, 13, e0191749. [Google Scholar]
- Ster, J.; De Bock, F.; Guérineau, N.C.; Janossy, A.; Barrère-Lemaire, S.; Bos, J.L.; Bockaert, J.; Fagni, L. Exchange protein activated by cAMP (Epac) mediates cAMP activation of p38 MAPK and modulation of Ca2+-dependent K+ channels in cerebellar neurons. Proc. Natl. Acad. Sci. USA 2007, 104, 2519–2524. [Google Scholar] [CrossRef]
- Rancz, E.A.; Häusser, M. Dendritic calcium spikes are tunable triggers of cannabinoid release and short-term synaptic plasticity in cerebellar purkinje neurons. J. Neurosci. 2006, 26, 5428–5437. [Google Scholar] [CrossRef]
- Ouares, K.A.; Filipis, L.; Tzilivaki, A.; Poirazi, P.; Canepari, M. Two Distinct Sets of Ca2+ and K+ Channels Are Activated at Different Membrane Potentials by the Climbing Fiber Synaptic Potential in Purkinje Neuron Dendrites. J. Neurosci. 2019, 39, 1969–1981. [Google Scholar] [CrossRef]
- Hirono, M.; Ogawa, Y.; Misono, K.; Zollinger, D.R.; Trimmer, J.S.; Rasband, M.N.; Misonou, H. BK Channels Localize to the Paranodal Junction and Regulate Action Potentials in Myelinated Axons of Cerebellar Purkinje Cells. J. Neurosci. 2015, 35, 7082–7094. [Google Scholar] [CrossRef]
- Montgomery, J.R.; Meredith, A.L. Genetic activation of BK currents in vivo generates bidirectional effects on neuronal excitability. Proc. Natl. Acad. Sci. USA 2012, 109, 18997–19002. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Wang, Y.-X.; Petralia, R.S.; Brenowitz, S.D. Cholinergic Modulation of Large-Conductance Calcium-Activated Potassium Channels Regulates Synaptic Strength and Spine Calcium in Cartwheel Cells of the Dorsal Cochlear Nucleus. J. Neurosci. 2014, 34, 5261–5272. [Google Scholar] [CrossRef]
- Garaycochea, J.; Slaughter, M.M. GABAB receptors enhance excitatory responses in isolated rat retinal ganglion cells. J. Physiol. 2016, 594, 5543–5554. [Google Scholar] [CrossRef]
- Womack, M.; Hoang, C.; Khodakhah, K. Large conductance calcium-activated potassium channels affect both spontaneous firing and intracellular calcium concentration in cerebellar Purkinje neurons. Neuroscience 2009, 162, 989–1000. [Google Scholar] [CrossRef]
- Nelson, A.B.; Krispel, C.M.; Sekirnjak, C.; du Lac, S. Long-Lasting Increases in Intrinsic Excitability Triggered by Inhibition. Neuron 2003, 40, 609–620. [Google Scholar] [CrossRef]
- Kimm, T.; Khaliq, Z.M.; Bean, B.P. Differential Regulation of Action Potential Shape and Burst-Frequency Firing by BK and Kv2 Channels in Substantia Nigra Dopaminergic Neurons. J. Neurosci. 2015, 35, 16404–16417. [Google Scholar] [CrossRef]
- Gittis, A.; Moghadam, S.H.; Du Lac, S. Mechanisms of Sustained High Firing Rates in Two Classes of Vestibular Nucleus Neurons: Differential Contributions of Resurgent Na, Kv3, and BK Currents. J. Neurophysiol. 2010, 104, 1625–1634. [Google Scholar] [CrossRef]
- Petrucci, C.; Resta, V.; Fieni, F.; Bigiani, A.; Bagnoli, P. Modulation of potassium current and calcium influx by somatostatin in rod bipolar cells isolated from the rabbit retina via sst2 receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2001, 363, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Gittis, A.H.; Lac, S. Firing Properties of GABAergic Versus Non-GABAergic Vestibular Nucleus Neurons Conferred by a Differential Balance of Potassium Currents. J. Neurophysiol. 2007, 2003, 3986. [Google Scholar] [CrossRef] [PubMed]
- Kohlmeier, K.A.; Christensen, M.H.; Kristensen, M.P.; Kristiansen, U. Pharmacological evidence of functional inhibitory metabotrophic glutamate receptors on mouse arousal-related cholinergic laterodorsal tegmental neurons. Neuropharmacology 2013, 66, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Savtchouk, I.; Acharjee, S.; Liu, S.J. Inhibition of Ca2+-activated large-conductance K+ channel activity alters synaptic AMPA receptor phenotype in mouse cerebellar stellate cells. J. Neurophysiol. 2011, 106, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Bock, T.; Stuart, G.J. Impact of calcium-activated potassium channels on NMDA spikes in cortical layer 5 pyramidal neurons. J. Neurophysiol. 2016, 115, 1740–1748. [Google Scholar] [CrossRef]
- Golding, N.L.; Staff, N.P.; Spruston, N. Dendritic spikes as a mechanism for cooperative long-term potentiation. Nature 2002, 418, 326–331. [Google Scholar] [CrossRef]
- Poolos, N.P.; Johnston, D. Calcium-Activated Potassium Conductances Contribute to Action Potential Repolarization at the Soma But Not the Dendrites of Hippocampal CA1 Pyramidal Neurons. J. Neurosci. 1999, 19, 5205–5212. [Google Scholar] [CrossRef]
- Egaña-Huguet, J.; Saumell-Esnaola, M.; Achicallende, S.; Soria-Gomez, E.; Río, I.B.-D.; del Caño, G.G.; Barrondo, S.; Sallés, J.; Gerrikagoitia, I.; Puente, N.; et al. Lack of the Transient Receptor Potential Vanilloid 1 Shifts Cannabinoid-Dependent Excitatory Synaptic Plasticity in the Dentate Gyrus of the Mouse Brain Hippocampus. Front. Neuroanat. 2021, 15, 701573. [Google Scholar] [CrossRef]
- Sclip, A.; Acuna, C.; Luo, F.; Südhof, T.C. RIM-binding proteins recruit BK-channels to presynaptic release sites adjacent to voltage-gated Ca2+ -channels. EMBO J. 2018, 37, 1–14. [Google Scholar] [CrossRef]
- Brenes, O.; Vandael, D.; Carbone, E.; Montarolo, P.; Ghirardi, M. Knock-down of synapsin alters cell excitability and action potential waveform by potentiating BK and voltage-gated Ca2+ currents in Helix serotonergic neurons. Neuroscience 2015, 311, 430–443. [Google Scholar] [CrossRef]
- Oh, K.H.; Abraham, L.S.; Gegg, C.; Silvestri, C.; Huang, Y.-C.; Alkema, M.J.; Furst, J.; Raicu, D.; Kim, H. Presynaptic BK channel localization is dependent on the hierarchical organization of alpha-catulin and dystrobrevin and fine-tuned by CaV2 calcium channels. BMC Neurosci. 2015, 16, 1–13. [Google Scholar] [CrossRef]
- Chen, B.; Liu, P.; Zhan, H.; Wang, Z.-W. Dystrobrevin Controls Neurotransmitter Release and Muscle Ca2+ Transients by Localizing BK Channels in Caenorhabditis elegans. J. Neurosci. 2011, 31, 17338–17347. [Google Scholar] [CrossRef]
- Abrahamsson, T.; Chou, C.Y.C.; Li, S.Y.; Mancino, A.; Costa, R.P.; Brock, J.A.; Nuro, E.; Buchanan, K.A.; Elgar, D.; Blackman, A.V.; et al. Differential Regulation of Evoked and Spontaneous Release by Presynaptic NMDA Receptors. Neuron 2017, 96, 839–855.e5. [Google Scholar] [CrossRef]
- Banerjee, A.; Larsen, R.; Philpot, B.D.; Paulsen, O. Roles of Presynaptic NMDA Receptors in Neurotransmission and Plasticity. Trends Neurosci. 2015, 39, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Lituma, P.J.; Kwon, H.-B.; Alviña, K.; Luján, R.; Castillo, P.E. Presynaptic NMDA receptors facilitate short-term plasticity and BDNF release at hippocampal mossy fiber synapses. Elife 2021, 10, e66612. [Google Scholar] [CrossRef]
- Martire, M.; Barrese, V.; D’Amico, M.; Iannotti, F.A.; Pizzarelli, R.; Samengo, I.; Viggiano, D.; Ruth, P.; Cherubini, E.; Taglialatela, M. Pre-synaptic BK channels selectively control glutamate versus GABA release from cortical and hippocampal nerve terminals. J. Neurochem. 2010, 115, 411–422. [Google Scholar] [CrossRef]
- Samengo, I.; Curro’, D.; Barrese, V.; Taglialatela, M.; Martire, M. Large Conductance Calcium-Activated Potassium Channels: Their Expression and Modulation of Glutamate Release from Nerve Terminals Isolated from Rat Trigeminal Caudal Nucleus and Cerebral Cortex. Neurochem. Res. 2014, 39, 901–910. [Google Scholar] [CrossRef]
- Shoudai, K.; Nonaka, K.; Maeda, M.; Wang, Z.-M.; Jeong, H.-J.; Higashi, H.; Murayama, N.; Akaike, N. Effects of various K+ channel blockers on spontaneous glycine release at rat spinal neurons. Brain Res. 2007, 1157, 11–22. [Google Scholar] [CrossRef]
- De San Martín, J.Z.; Pyott, S.; Ballestero, J.; Katz, E. Ca2+ and Ca2+-activated K+ channels that support and modulate transmitter release at the olivocochlear efferent-inner hair cell synapse. J. Neurosci. 2010, 30, 12157–12167. [Google Scholar] [CrossRef]
- Choi, T.-Y.; Lee, S.-H.; Kim, Y.-J.; Bae, J.R.; Lee, K.M.; Jo, Y.; Kim, S.-J.; Lee, A.-R.; Choi, S.; Choi, L.-M.; et al. Cereblon Maintains Synaptic and Cognitive Function by Regulating BK Channel. J. Neurosci. 2018, 38, 3571–3583. [Google Scholar] [CrossRef]
- Price, D.L.; Ludwig, J.W.; Mi, H.; Schwarz, T.L.; Ellisman, M.H. Distribution of rSlo Ca2+-activated K+ channels in rat astrocyte perivascular endfeet. Brain Res. 2002, 956, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Girouard, H.; Bonev, A.D.; Hannah, R.M.; Meredith, A.; Aldrich, R.W.; Nelson, M.T. Astrocytic endfoot Ca 2+ and BK channels determine both arteriolar dilation and constriction. Proc. Natl. Acad. Sci. USA 2010, 107, 3811–3816. [Google Scholar] [CrossRef] [PubMed]
- Kyle, B.; Mishra, R.C.; Braun, A.P. The augmentation of BK channel activity by nitric oxide signaling in rat cerebral arteries involves co-localized regulatory elements. J. Cereb. Blood Flow Metab. 2017, 37, 3759–3773. [Google Scholar] [CrossRef] [PubMed]
- Filosa, J.A.; Bonev, A.D.; Straub, S.V.; Meredith, A.; Wilkerson, M.K.; Aldrich, R.W.; Nelson, M. Local potassium signaling couples neuronal activity to vasodilation in the brain. Nat. Neurosci. 2006, 9, 1397–1403. [Google Scholar] [CrossRef]
- Gebremedhin, D.; Yamaura, K.; Zhang, C.; Bylund, J.; Koehler, R.C.; Harder, D.R. Metabotropic Glutamate Receptor Activation Enhances the Activities of Two Types of Ca2+-Activated K+Channels in Rat Hippocampal Astrocytes. J. Neurosci. 2003, 23, 1678–1687. [Google Scholar] [CrossRef]
- Brandalise, F.; Ratto, D.; Leone, R.; Olivero, F.; Roda, E.; Locatelli, C.A.; Grazia Bottone, M.; Rossi, P. Deeper and Deeper on the Role of BK and Kir4.1 Channels in Glioblastoma Invasiveness: A Novel Summative Mechanism? Front Neurosci. 2020, 14, 1–6. [Google Scholar] [CrossRef]
- Singh, H.; Li, M.; Hall, L.; Chen, S.; Sukur, S.; Lu, R.; Caputo, A.; Meredith, A.; Stefani, E.; Toro, L. MaxiK channel interactome reveals its interaction with GABA transporter 3 and heat shock protein 60 in the mammalian brain. Neuroscience 2016, 317, 76–107. [Google Scholar] [CrossRef]
- Hayashi, Y.; Nakanishi, H. BK channel in microglia as a potent therapeutic molecular target for neuropathic pain. J. Oral Biosci. 2015, 57, 131–134. [Google Scholar] [CrossRef]
- Huang, S.; Chen, T.; Suo, Q.; Shi, R.; Khan, H.; Ma, Y.; Tang, Y.; Yang, G.-Y.; Zhang, Z. BK Channel-Mediated Microglial Phagocytosis Alleviates Neurological Deficit After Ischemic Stroke. Front. Cell. Neurosci. 2021, 15, 683769. [Google Scholar] [CrossRef]
- Hayashi, Y.; Kawaji, K.; Sun, L.; Zhang, X.; Koyano, K.; Yokoyama, T.; Kohsaka, S.; Inoue, K.; Nakanishi, H. Microglial Ca2+-Activated K+ Channels Are Possible Molecular Targets for the Analgesic Effects of S-Ketamine on Neuropathic Pain. J. Neurosci. 2011, 31, 17370–17382. [Google Scholar] [CrossRef]
- Bordey, A.; Spencer, D.D. Chemokine modulation of high-conductance Ca2+-sensitive K+ currents in microglia from human hippocampi. J. FENS 2003, 18, 2893–2898. [Google Scholar] [CrossRef]
- Schilling, T.; Eder, C. Ion channel expression in resting and activated microglia of hippocampal slices from juvenile mice. Brain Res. 2007, 1186, 21–28. [Google Scholar] [CrossRef]
- Schilling, T.; Eder, C. Microglial K + channel expression in young adult and aged mice. Glia 2014, 63, 664–672. [Google Scholar] [CrossRef]
- Buttigieg, J.; Eftekharpour, E.; Karimi-Abdolrezaee, S.; Fehlings, M.G. Molecular and electrophysiological evidence for the expression of BK channels in oligodendroglial precursor cells. Eur. J. Neurosci. 2011, 34, 538–547. [Google Scholar] [CrossRef]
- Larson, V.A.; Zhang, Y.; Bergles, D.E. Electrophysiological properties of NG2 þ cells: Matching physiological studies with gene expression profiles. Brain Res. 2015, 15, 1853. [Google Scholar]
- Jäkel, S.; Agirre, E.; Falcão, A.M.; van Bruggen, D.; Lee, K.W.; Knuesel, I.; Malhotra, D.; Ffrench-Constant, C.; Williams, A.; Castelo-Branco, G. Altered human oligodendrocyte heterogeneity in multiple sclerosis. Nature 2019, 566, 543–547. [Google Scholar] [CrossRef]
- Valverde, M.A.; Rojas, P.; Amigo, J.; Cosmelli, D.; Orio, P.; Bahamonde, M.I.; Mann, G.E.; Vergara, C.; Latorre, R. Acute Activation of Maxi-K Channels (hSlo) by Estradiol Binding to the β Subunit. Science 1999, 285, 1929–1931. [Google Scholar] [CrossRef]
- King, J.T.; Lovell, P.V.; Rishniw, M.; Kotlikoff, M.I.; Zeeman, M.L.; McCobb, D.P. β2 and β4 Subunits of BK Channels Confer Differential Sensitivity to Acute Modulation by Steroid Hormones. J. Neurophysiol. 2006, 95, 2878–2888. [Google Scholar] [CrossRef]
- Teyler, T.J.; Vardaris, R.M.; Lewis, D.; Rawitch, A.B. Gonadal Steroids: Effects on Excitability of Hippocampal Pyramidal Cells. Science 1980, 209, 1017–1018. [Google Scholar] [CrossRef]
- Oberlander, J.G.; Woolley, C.S. 17β-Estradiol Acutely Potentiates Glutamatergic Synaptic Transmission in the Hippocampus through Distinct Mechanisms in Males and Females. J. Neurosci. 2016, 36, 2677–2690. [Google Scholar] [CrossRef]
- Soutar, C.N.; Grenier, P.; Patel, A.; Kabitsis, P.P.; Olmstead, M.C.; Bailey, C.D.C.; Dringenberg, H.C. Brain-Generated 17β-Estradiol Modulates Long-Term Synaptic Plasticity in the Primary Auditory Cortex of Adult Male Rats. Cereb. Cortex 2021, 32, 2140–2155. [Google Scholar] [CrossRef]
- Hoshi, T.; Tian, Y.; Xu, R.; Heinemann, S.H.; Hou, S. Mechanism of the modulation of BK potassium channel complexes with different auxiliary subunit compositions by the omega-3 fatty acid DHA. Proc. Natl. Acad. Sci. USA 2013, 110, 4822–4827. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhou, D.; Zhang, P.; Moczydlowski, E.G.; Haddad, G.G. β-subunit-dependent modulation of hSlo BK current by arachidonic acid. J. Neurophysiol. 2007, 97, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Duerson, K.; White, R.E.; Jiang, F.; Schonbrunn, A.; Armstrong, D.L. Somatostatin stimulates BK(Ca) channels in rat pituitary tumor cells through lipoxygenase metabolites of arachidonic acid. Neuropharmacology 1996, 35, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Barlow, R.S.; El-Mowafy, A.M.; White, A.R.E. H2O2 opens BK(Ca) channels via the PLA2-arachidomic acid signaling cascade in coronary artery smooth muscle. Am. J. Physiol.-Hear. Circ. Physiol. 2000, 279, H475–H483. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, A.I.; Panasiuk, O.; Okhai, I.; Montecucco, F.; Brandt, K.J.; Mach, F. Direct activation of Ca2+ and voltage-gated potassium channels of large conductance by anandamide in endothelial cells does not support the presence of endothelial atypical cannabinoid receptor. Eur. J. Pharmacol. 2017, 805, 14–24. [Google Scholar] [CrossRef]
- Sade, H.; Muraki, K.; Ohya, S.; Hatano, N.; Imaizumi, Y. Activation of large-conductance, Ca2+-activated K+ channels by cannabinoids. Am. J. Physiol.-Cell Physiol. 2006, 290, C77–C86. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Wu, Y.; Zheng, Q.; Chen, M.; Qian, Z.; Wei, C.; Han, J.; Liu, Z.; Ren, W.; et al. Cannabinoids-induced peripheral analgesia depends on activation of BK channels. Brain Res. 2019, 1711, 23–28. [Google Scholar] [CrossRef]
- Bailey, C.S.; Moldenhauer, H.J.; Park, S.M.; Keros, S.; Meredith, A.L. KCNMA1-linked channelopathy. J. Gen. Physiol. 2019, 151, 1173–1189. [Google Scholar] [CrossRef]
- Miller, J.P.; Moldenhauer, H.J.; Keros, S.; Meredith, A.L. An emerging spectrum of variants and clinical features in KCNMA1-linked channelopathy. Channels 2021, 15, 447–464. [Google Scholar] [CrossRef]
- Du, W.; Bautista, J.F.; Yang, H.; Diez-Sampedro, A.; You, S.-A.; Wang, L.; Kotagal, P.; Lüders, H.O.; Shi, J.; Cui, J.; et al. Calcium-sensitive potassium channelopathy in human epilepsy and paroxysmal movement disorder. Nat. Genet. 2005, 37, 733–738. [Google Scholar] [CrossRef]
- Mi Park, S.; Roache, C.E.; Iffland, P.H., II; Moldenhauer, H.J.; Matychak, K.K.; Plante, A.E. BK channel properties correlate with neurobehavioral severity in three KCNMA1-linked channelopathy mouse models. Elife 2022, 11, e77953. [Google Scholar] [CrossRef]
- Li, X.; Poschmann, S.; Chen, Q.; Fazeli, W.; Oundjian, N.J.; Snoeijen-Schouwenaars, F.M.; Fricke, O.; Kamsteeg, E.-J.; Willemsen, M.; Wang, Q.K. De novo BK channel variant causes epilepsy by affecting voltage gating but not Ca2+ sensitivity. Eur. J. Hum. Genet. 2018, 26, 220–229. [Google Scholar] [CrossRef]
- Whitmire, L.E.; Ling, L.; Bugay, V.; Carver, C.M.; Timilsina, S.; Chuang, H.-H.; Jaffe, D.B.; Shapiro, M.S.; Cavazos, J.; Brenner, R. Downregulation of KCNMB4 expression and changes in BK channel subtype in hippocampal granule neurons following seizure activity. PLoS ONE 2017, 12, e0188064. [Google Scholar] [CrossRef]
- Lorenz, S.; Heils, A.; Kasper, J.M.; Sander, T. Allelic association of a truncation mutation of theKCNMB3 gene with idiopathic generalized epilepsy. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2007, 144B, 10–13. [Google Scholar] [CrossRef]
- Chen, S.R.; Cai, Y.Q.; Pan, H.L. Plasticity and emerging role of BKCa channels in nociceptive control in neuropathic pain. J Neurochem. 2009, 110, 352–362. [Google Scholar] [CrossRef]
- Furukawa, N.; Takasusuki, T.; Fukushima, T.; Hori, Y. Presynaptic large-conductance calcium-activated potassium channels control synaptic transmission in the superficial dorsal horn of the mouse. Neurosci. Lett. 2008, 444, 79–82. [Google Scholar] [CrossRef]
- Sarantopoulos, C.D.; McCallum, J.B.; Rigaud, M.; Fuchs, A.; Kwok, W.-M.; Hogan, Q.H. Opposing effects of spinal nerve ligation on calcium-activated potassium currents in axotomized and adjacent mammalian primary afferent neurons. Brain Res. 2007, 1132, 84–99. [Google Scholar] [CrossRef]
- Zhang, F.-X.; Gadotti, V.M.; Souza, I.A.; Chen, L.; Zamponi, G.W. BK Potassium Channels Suppress Cavα2δ Subunit Function to Reduce Inflammatory and Neuropathic Pain. Cell Rep. 2018, 22, 1956–1964. [Google Scholar] [CrossRef]
- Imari, K.; Harada, Y.; Zhang, J.; Mori, Y.; Hayashi, Y. KCNMB3 in spinal microglia contributes to the generation and maintenance of neuropathic pain in mice. Int. J. Mol. Med. 2019, 44, 1585–1593. [Google Scholar] [CrossRef]
- Typlt, M.; Mirkowski, M.; Azzopardi, E.; Ruettiger, L.; Ruth, P.; Schmid, S. Mice with Deficient BK Channel Function Show Impaired Prepulse Inhibition and Spatial Learning, but Normal Working and Spatial Reference Memory. PLoS ONE 2013, 8, e81270. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Carvalho-De-Souza, J.L.; Wei, C.; Carrasquel-Ursulaez, W.; Lorenzo, Y.; Gonzalez, N.; Kubota, T.; Staisch, J.; Hain, T.; Petrossian, N.; et al. Loss-of-function BK channel mutation causes impaired mitochondria and progressive cerebellar ataxia. Proc. Natl. Acad. Sci. USA 2020, 117, 6023–6034. [Google Scholar] [CrossRef] [PubMed]
- Kratschmer, P.; Lowe, S.A.; Buhl, E.; Chen, K.F.; Kullmann, D.M.; Pittman, A.; Hodge, J.J.L.; Jepson, J.E.C. Impaired Pre-Motor Circuit Activity and Movement in a Drosophila Model of KCNMA1-Linked Dyskinesia. Mov. Disord. 2021, 36, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhao, J.; Ardiel, E.L.; Hall, Q.; Nurrish, S.; Kaplan, J.M. Shank promotes action potential repolarization by recruiting BK channels to calcium microdomains. Elife 2022, 11, e75140. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.-Y.; Klyachko, V.A. Channelopathies in fragile X syndrome. Nat. Rev. Neurosci. 2021, 22, 275–289. [Google Scholar] [CrossRef]
- Hébert, B.; Pietropaolo, S.; Même, S.; Laudier, B.; Laugeray, A.; Doisne, N.; Quartier, A.; Lefeuvre, S.; Got, L.; Cahard, D.; et al. Rescue of fragile X syndrome phenotypes in Fmr1KO mice by a BKCa channel opener molecule. Orphanet. J. Rare Dis. 2014, 9, 124. [Google Scholar] [CrossRef]
- Edwards, T.; Rickard, N. Pharmaco-behavioural evidence indicating a complex role for ryanodine receptor calcium release channels in memory processing for a passive avoidance task. Neurobiol. Learn. Mem. 2006, 86, 1–8. [Google Scholar] [CrossRef]
- Matthews, E.A.; Weible, A.P.; Shah, S.; Disterhoft, J.F. The BK-mediated fAHP is modulated by learning a hippocampus-dependent task. Proc. Natl. Acad. Sci. USA 2008, 105, 15154–15159. [Google Scholar] [CrossRef]
- Matthews, E.A.; Disterhoft, J.F. Blocking the BK channel impedes acquisition of trace eyeblink conditioning. Learn. Mem. 2009, 16, 106–109. [Google Scholar] [CrossRef]
- Ryan, D.P.; Henzel, K.S.; Pearson, B.L.; Siwek, M.E.; Papazoglou, A.; Guo, L.; Paesler, K.; Yu, M.; Müller, R.; Xie, K.; et al. A paternal methyl donor-rich diet altered cognitive and neural functions in offspring mice. Mol. Psychiatry 2017, 23, 1345–1355. [Google Scholar] [CrossRef]
- Choi, T.-Y.; Lee, S.-H.; Kim, S.-J.; Jo, Y.; Park, C.-S.; Choi, S.-Y. BK channel blocker paxilline attenuates thalidomide-caused synaptic and cognitive dysfunctions in mice. Sci. Rep. 2018, 8, 17653. [Google Scholar] [CrossRef]
- Beecham, G.W.; Schnetz-Boutaud, N.; Haines, J.L.; Pericak-Vance, M.A. CALHM1Polymorphism is not Associated with Late-onset Alzheimer Disease. Ann. Hum. Genet. 2009, 73, 379–381. [Google Scholar] [CrossRef]
- Beecham, G.W.; Hamilton, K.; Naj, A.C.; Martin, E.R.; Huentelman, M.; Myers, A.J.; Corneveaux, J.J.; Hardy, J.; Vonsattel, J.-P.; Younkin, S.G.; et al. Genome-Wide Association Meta-analysis of Neuropathologic Features of Alzheimer’s Disease and Related Dementias. PLOS Genet. 2014, 10, e1004606. [Google Scholar] [CrossRef]
- Ye, H.; Jalini, S.; Mylvaganam, S.; Carlen, P. Activation of large-conductance Ca2+-activated K+ channels depresses basal synaptic transmission in the hippocampal CA1 area in APP (swe/ind) TgCRND8 mice. Neurobiol. Aging 2010, 31, 591–604. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ueta, Y.; Wang, L.; Yamamoto, R.; Inoue, N.; Inokuchi, K.; Aiba, A.; Yonekura, H.; Kato, N. Suppression of a neocortical potassium channel activity by intracellular amyloid-β and its rescue with homer1a. J. Neurosci. 2011, 31, 11100–11109. [Google Scholar] [CrossRef] [Green Version]
- Gavello, D.; Calorio, C.; Franchino, C.; Cesano, F.; Carabelli, V.; Carbone, E.; Marcantoni, A. Early Alterations of Hippocampal Neuronal Firing Induced by Abeta42. Cereb. Cortex 2018, 28, 433–446. [Google Scholar] [CrossRef]
- Wang, L.; Kang, H.; Li, Y.; Shui, Y.; Yamamoto, R.; Sugai, T.; Kato, N. Cognitive recovery by chronic activation of the large-conductance calcium-activated potassium channel in a mouse model of Alzheimer’s disease. Neuropharmacology 2015, 92, 8–15. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CNS Area and Neuronal Type | Site of Neuronal Expression | Function | Subunit and/or Pharmacology | References |
|---|---|---|---|---|
| Prefrontal cortex pyramidal neurons | Pre- (AIS and axon) and postsynaptic (soma) | Limit AP broadening and allow high-frequency firing. | IBTX-sensitive | [151,152] |
| Pax-sensitive | [153] | |||
| Visual cortex pyramidal neurons | Postsynaptic (basal dendrites) | Reduce EPSPs in small-headed spines. | α subunit, Chtx-sensitive | [25] |
| Somatosensorial layer 5 pyramidal neurons | Postsynaptic (soma and dendrites) | Reduce influx of ions through NMDAR and increase threshold for plasticity. Modulate dendritic Na+ and Ca2+ spikes. | IBTX and Pax-sensitive | [154] |
| Pax-sensitive | [112,155,156] | |||
| Hippocampal CA1 pyramidal neuron | Pre- (axon) and Postsynaptic (soma and apical dendrites) | Limit repetitive firing of dendritic calcium spikes. AP repolarization and fAHP. | α subunit | [141,157] |
| Chtx-sensitive | [158,159] | |||
| Pax and IBTX-sensitive | [160] | |||
| IBTX-sensitive | [137,142] | |||
| Hippocampal CA3 pyramidal neuron | Pre- (Axonal terminals) and postsynaptic (soma) | Decrease glutamate release. Regulate neuronal excitability. | α subunit | [161] |
| β4 subunit | [15,162] | |||
| Chtx-sensitive | [163] | |||
| IBTX-sensitive | [23] | |||
| IBTX and Pax-sensitive | [24] | |||
| Hippocampal dentate gyrus granule cells | Pre- (mossy fiber boutons and perforant path terminals) and postsynaptic (soma and dendrites) | Control perforant path induce EPSP. Regulate excitability. At mossy fiber boutons contribute to AP repolarization when Kv channels are blocked. | α subunit | [161] |
| β2 and β4 subunits | [164] | |||
| β4 subunit | [72] | |||
| Pax-, Chtx- and IBTX-sensitive | [99] | |||
| Pax-sensitive | [27] | |||
| Piriform cortex pyramidal neuron | Postsynaptic (soma) | Reduce excitability modulating odor perception. | Pax-sensitive | [165] |
| Anterior cingulate cortex pyramidal neurons | Pre- (axonal terminals) and postsynaptic (soma) | Reduce hyperexcitability induced by neuropathic pain state. | β4 subunit. | [73] |
| Somatosensorial SOM+ interneurons | Pre- (axon) and postsynaptic (soma) | Contribute to AP repolarization. | IBTX-sensitive | [166] |
| Amygdala central nucleus gabaergic neurons | Presynaptic | Limit GABA release. | β2 and β4 subunits | [12] |
| Substantia nigra pars reticulata gabaergic neurons | Postsynaptic (soma) | Contribute to hyperpolarization. | Chtx- and Pax-sensitive | [167] |
| Substantia nigra dopaminergic neurons | Postsynaptic (soma) | Not tested. | IBTX-senstive | [134] |
| Infragranular pyramidal neurons | Postsynaptic (soma and apical dendrites) | Not tested. | Chtx-sensitive | [135] |
| Lateral habenula nucleus neurons | Postsynaptic (soma) | fAHP increased by corticotropin-releasing factor activation. | IBTX-sensitive | [168] |
| Medial habenula nucleus neurons | Not described | Not tested. | LRRC55 γ subunit | [169] |
| Cerebellum Purkinje neurons | Presynaptic (paranodal junctions of axons) Postsynaptic (soma and dendrites) | Decrease dendritic Ca2+ spikes avoiding DSE propagation. Regulate afterdepolarization. In myelinated axons allow high-fidelity firing of AP. | α subunit | [161] |
| β4 subunit | [170] | |||
| Penitrem A sensitive. | [171] | |||
| IBTX-sensitive | [172] | |||
| IBTX–sensitive + IBTX-insensitive component | [138] | |||
| IBTX and Pax-sensitive. | [102] | |||
| Pax-sensitive | [173] | |||
| Cerebellar basket cells | Presynaptic (axonal terminals) | Not tested. | α subunit | [161] |
| Cerebellar granule cells | Postsynaptic (soma) | Contribute to hyperpolarization and AHP. | IBTX-sensitive | [170] |
| Suprachiasmatic nucleus neurons | Postsynaptic (soma) | Increase neuronal firing at night but decrease firing during day. | β4 subunit | [174] |
| β2 subunit | [86] | |||
| Supraoptic nucleus neurons | Pre- and postsynaptic (dendrites, soma and nerve terminals) | β4 but not β1 allows potentiation by ethanol. | β1 subunit in soma and dendrites; β4 in nerve terminals | [79] |
| Medial vestibular nucleus neurons | Postsynaptic (soma) | Contribute to excitability. Can be activated by CaMKII and inhibited by PKC. | Not tested | [125] |
| Dorsal cochlear nucleus cartwheel cells | Postsynaptic (soma and dendrites) | Control EPSP. | IBTX-sensitive | [113,175] |
| Retinal ganglion cells | Postsynaptic (soma) | Regulate cell excitability. | IBTX-sensitive | [176] |
| A17 amacrine cells | Postsynaptic (varicosities) | Limit Cav channel activation and GABA release. | β2 subunit, IBTX-sensitive | [22] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancatén-González, C.; Segura, I.; Alvarado-Sánchez, R.; Chávez, A.E.; Latorre, R. Ca2+- and Voltage-Activated K+ (BK) Channels in the Nervous System: One Gene, a Myriad of Physiological Functions. Int. J. Mol. Sci. 2023, 24, 3407. https://doi.org/10.3390/ijms24043407
Ancatén-González C, Segura I, Alvarado-Sánchez R, Chávez AE, Latorre R. Ca2+- and Voltage-Activated K+ (BK) Channels in the Nervous System: One Gene, a Myriad of Physiological Functions. International Journal of Molecular Sciences. 2023; 24(4):3407. https://doi.org/10.3390/ijms24043407
Chicago/Turabian StyleAncatén-González, Carlos, Ignacio Segura, Rosangelina Alvarado-Sánchez, Andrés E. Chávez, and Ramon Latorre. 2023. "Ca2+- and Voltage-Activated K+ (BK) Channels in the Nervous System: One Gene, a Myriad of Physiological Functions" International Journal of Molecular Sciences 24, no. 4: 3407. https://doi.org/10.3390/ijms24043407