
Biofilms in Chronic Wound Infections: Innovative Antimicrobial Approaches Using the In Vitro Lubbock Chronic Wound Biofilm Model

 , , ,
, , ,  and
and
Abstract
:1. Introduction
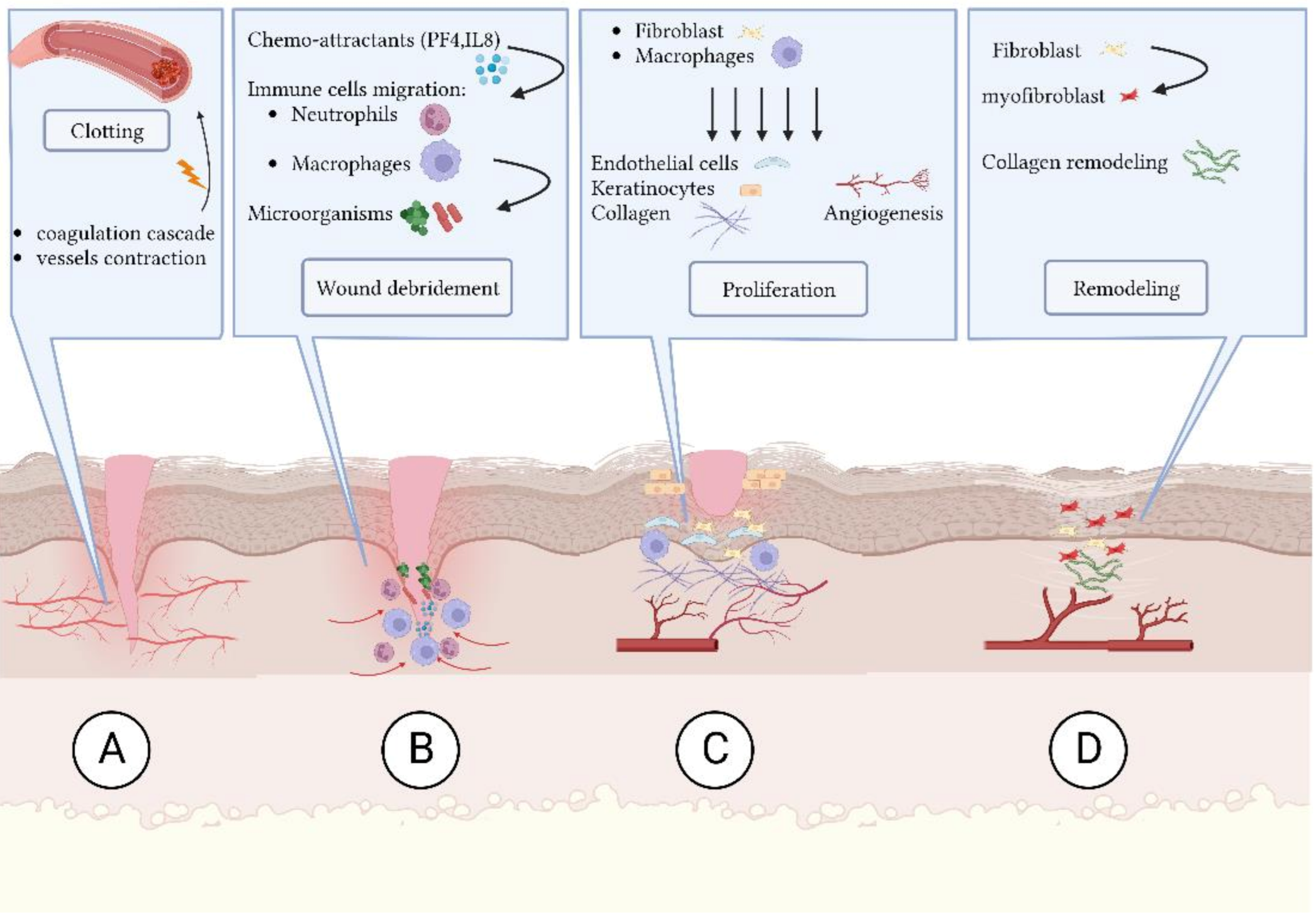
- (A)
- Coagulation and hemostasis: blood vessels contract and the coagulation cascade seals the injured area and minimizes blood loss [2];
- (B)
- (C)
- (D)
2. Clinical Significance of Chronic Wounds
2.1. Microbial Composition in Wound Site
2.2. Biofilms in Chronic Wounds
3. In Vitro Wound Models
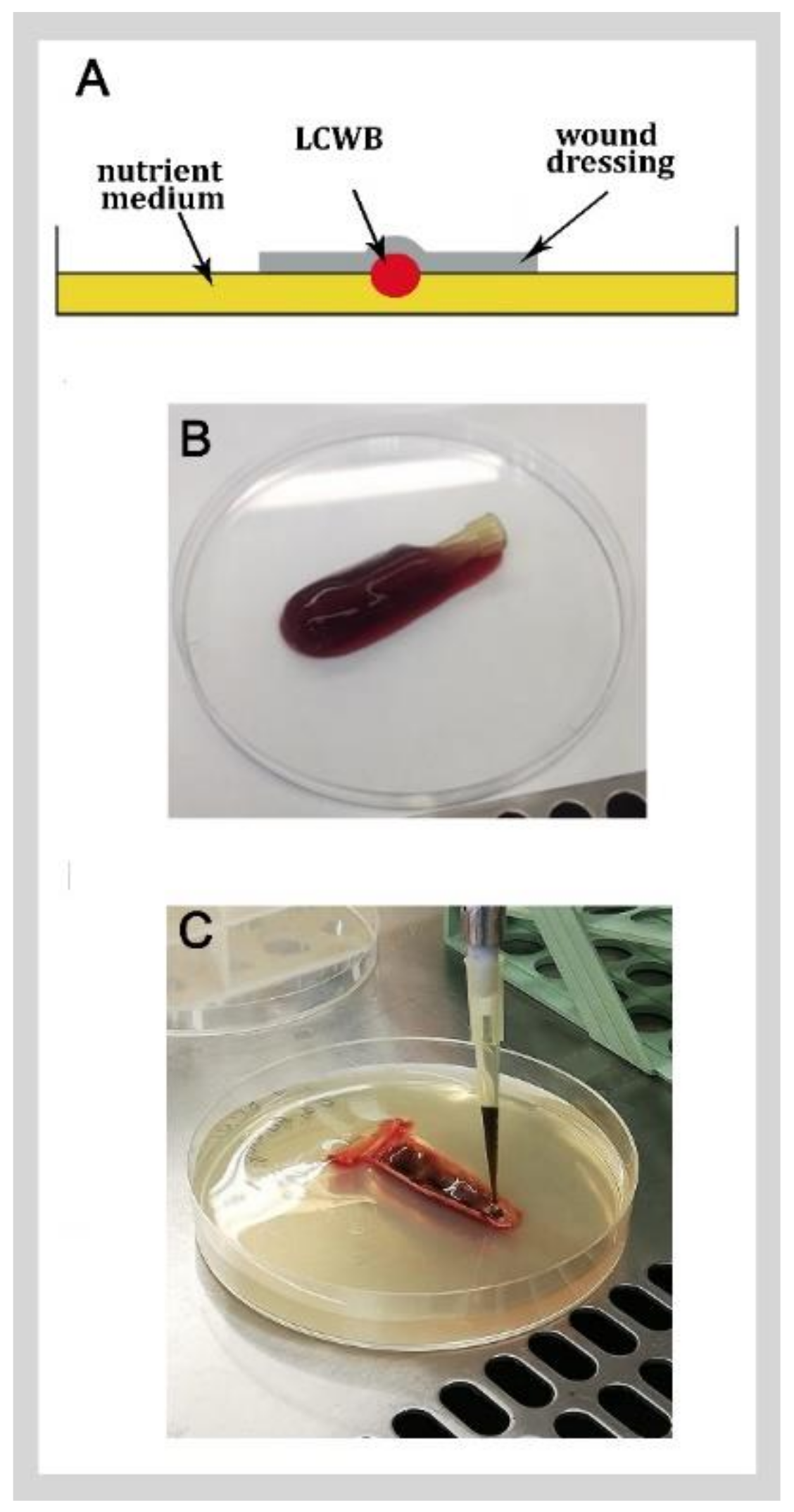
4. Lubbock Chronic Wound Biofilm (LCWB)
4.1. LCWB Models
4.2. LCWB as Model for Detection of Novel Strategies
5. Understanding Biofilms Using the “Zone Model”
6. Promising Therapies in Chronic Wounds
- -
- Ultrasonic debridement
- -
- Antiseptics and Antibiotics
- -
- Antimicrobial peptides
- -
- Photodynamic therapy
- -
- Biodegradable bacterial by-products
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falcone, M.; De Angelis, B.; Pea, F.; Scalise, A.; Stefani, S.; Tasinato, R.; Zanetti, O.; Paola, L.D. Challenges in the management of chronic wound infections. J. Glob. Antimicrob. Resist. 2021, 26, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Abazari, M.; Ghaffari, A.; Rashidzadeh, H.; Badeleh, S.M.; Maleki, Y. A Systematic Review on Classification, Identification, and Healing Process of Burn Wound Healing. Int. J. Low. Extrem. Wounds 2020, 21, 18–30. [Google Scholar] [CrossRef]
- McDaniel, J.C.; Roy, S.; Wilgus, T.A. Neutrophil activity in chronic venous leg ulcers-A target for therapy? Wound Repair Regen. 2013, 21, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Richmond, A. Chemokine Regulation of Neutrophil Infiltration of Skin Wounds. Adv. Wound Care 2015, 4, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Ellis, S.; Lin, E.J.; Tartar, D. Immunology of Wound Healing. Curr. Dermatol. Rep. 2018, 7, 350–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, D.L.; Trinder, S.; Labelle, M.; Rodriguez-Justo, M.; Hughes, S.; Holmes, A.M.; Scotton, C.J.; Porter, J.C. Platelet-derived transforming growth factor-β1 promotes keratinocyte proliferation in cutaneous wound healing. J. Tissue Eng. Regen. Med. 2020, 14, 645–649. [Google Scholar] [CrossRef] [Green Version]
- Ridiandries, A.; Tan, J.; Bursill, C. The Role of Chemokines in Wound Healing. Int. J. Mol. Sci. 2018, 19, 3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, S.; Koch, M.; Lee, Y.; Jung, F.; Blocki, A. An In Vitro Model of Angiogenesis during Wound Healing Provides Insights into the Complex Role of Cells and Factors in the Inflammatory and Proliferation Phase. Int. J. Mol. Sci. 2018, 19, 2913. [Google Scholar] [CrossRef] [Green Version]
- Cañedo-Dorantes, L.; Cañedo-Ayala, M. Skin Acute Wound Healing: A Comprehensive Review. Int. J. Inflamm. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. 2016, 58, 81–94. [Google Scholar] [CrossRef]
- Mervis, J.S.; Phillips, T.J. Pressure ulcers: Pathophysiology, epidemiology, risk factors, and presentation. J. Am. Acad. Dermatol. 2019, 81, 881–890. [Google Scholar] [CrossRef]
- Leren, L.; Johansen, E.; Eide, H.; Falk, R.S.; Juvet, L.K.; Ljoså, T.M. Pain in persons with chronic venous leg ulcers: A systematic review and meta-analysis. Int. Wound J. 2020, 17, 466–484. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.; Rocha, D.; Bezzerra, S.; Andrade, E.; Santos, A.; Nogueira, L. Quality of life of people with chronic wounds. ACTA Paul. Enferm. 2019, 32, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.; Hamm, R.L. Factors That Impair Wound Healing. J. Am. Coll. Clin. Wound Spéc. 2012, 4, 84–91. [Google Scholar] [CrossRef]
- Darvishi, S.; Tavakoli, S.; Kharaziha, M.; Girault, H.H.; Kaminski, C.F.; Mela, I. Advances in the Sensing and Treatment of Wound Biofilms. Angew. Chem. Int. Ed. 2022, 61, e202112218. [Google Scholar] [CrossRef]
- Rahim, K.; Saleha, S.; Zhu, X.; Huo, L.; Basit, A.; Franco, O.L. Bacterial Contribution in Chronicity of Wounds. Microb. Ecol. 2016, 73, 710–721. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.F.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti-Infect. Ther. 2015, 13, 605–613. [Google Scholar] [CrossRef]
- Wu, M.; Pan, H.; Leng, W.; Lei, X.; Chen, L.; Liang, Z. Distribution of Microbes and Drug Susceptibility in Patients with Diabetic Foot Infections in Southwest China. J. Diabetes Res. 2018, 2018, e9817308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahim, K.; Qasim, M.; Rahman, H.; Khan, T.A.; Ahmad, I.; Khan, N.; Ullah, A.; Basit, A.; Saleha, S. Antimicrobial resistance among aerobic biofilm producing bacteria isolated from chronic wounds in the tertiary care hospitals of Peshawar, Pakistan. J. Wound Care 2016, 25, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Krumkamp, R.; Oppong, K.; Hogan, B.; Strauss, R.; Frickmann, H.; Wiafe-Akenten, C.; Boahen, K.G.; Rickerts, V.; Smith, I.M.; Groß, U.; et al. Spectrum of antibiotic resistant bacteria and fungi isolated from chronically infected wounds in a rural district hospital in Ghana. PLoS ONE 2020, 15, e0237263. [Google Scholar] [CrossRef]
- Wong, S.Y.; Manikam, R.; Muniandy, S. Prevalence and antibiotic susceptibility of bacteria from acute and chronic wounds in Malaysian subjects. J. Infect. Dev. Ctries. 2015, 9, 936–944. [Google Scholar] [CrossRef] [Green Version]
- Wolcott, R.D.; Hanson, J.D.; Rees, E.J.; Koenig, L.D.; Phillips, C.; Wolcott, R.A.; Cox, S.B.; White, J.S. Analysis of the chronic wound microbiota of 2963 patients by 16S rDNA pyrosequencing. Wound Repair Regen. 2015, 24, 163–174. [Google Scholar] [CrossRef]
- Dowd, S.E.; Delton Hanson, J.; Rees, E.; Wolcott, R.D.; Zischau, A.M.; Sun, Y.; White, J.; Smith, D.M.; Kennedy, J.; Jones, C.E. Survey of fungi and yeast in polymicrobial infections in chronic wounds. J. Wound Care 2011, 20, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Kirketerp-Møller, K.; Jensen, P.Ø.; Fazli, M.; Madsen, K.G.; Pedersen, J.; Moser, C.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Distribution, Organization, and Ecology of Bacteria in Chronic Wounds. J. Clin. Microbiol. 2008, 46, 2717–2722. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Hochwalt, P.C.; Usui, M.L.; Underwood, R.A.; Singh, P.K.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Delayed wound healing in diabetic (db/db) mice with Pseudomonas aeruginosa biofilm challenge: A model for the study of chronic wounds. Wound Repair Regen. 2010, 18, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaarup, I.C.; Iversen, A.K.S.; Lichtenberg, M.; Bjarnsholt, T.; Jakobsen, T.H. Biofilm Survival Strategies in Chronic Wounds. Microorganisms 2022, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Gjødsbøl, K.; Christensen, J.J.; Karlsmark, T.; Jørgensen, B.; Klein, B.M.; Krogfelt, K.A. Multiple bacterial species reside in chronic wounds: A longitudinal study. Int. Wound J. 2006, 3, 225–231. [Google Scholar] [CrossRef]
- Price, L.B.; Liu, C.M.; Frankel, Y.M.; Melendez, J.H.; Aziz, M.; Buchhagen, J.; Contente-Cuomo, T.; Engelthaler, D.M.; Keim, P.S.; Ravel, J.; et al. Macroscale spatial variation in chronic wound microbiota: A cross-sectional study. Wound Repair Regen. 2010, 19, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percival, S.L.; Hill, K.E.; Malic, S.; Thomas, D.; Williams, D. Antimicrobial tolerance and the significance of persister cells in recalcitrant chronic wound biofilms. Wound Repair Regen. 2011, 19, 1–9. [Google Scholar] [CrossRef]
- Bessa, L.J.; Fazii, P.; Di Giulio, M.; Cellini, L. Bacterial isolates from infected wounds and their antibiotic susceptibility pattern: Some remarks about wound infection. Int. Wound J. 2015, 12, 47–52. [Google Scholar] [CrossRef]
- Conlon, B.P. Staphylococcus aureus chronic and relapsing infections: Evidence of a role for persister cells. Bioessays 2014, 36, 991–996. [Google Scholar] [CrossRef]
- Kwan, B.W.; Chowdhury, N.; Wood, T.K. Combatting bacterial infections by killing persister cells with mitomycin C. Environ. Microbiol. 2015, 17, 4406–4414. [Google Scholar] [CrossRef]
- Malone, M.; Bjarnsholt, T.; McBain, A.J.; James, G.A.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R.D. The prevalence of biofilms in chronic wounds: A systematic review and meta-analysis of published data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-K.; Cheng, N.-C.; Cheng, C.-M. Biofilms in Chronic Wounds: Pathogenesis and Diagnosis. Trends Biotechnol. 2019, 37, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Das, L.; Singh, Y. Quorum Sensing Inhibition: A Target for Treating Chronic Wounds. In Biotechnological Applications of Quorum Sensing Inhibitors; Springer: Singapore, 2018; pp. 111–126. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [Green Version]
- Kumar, L.; Patel, S.K.S.; Kharga, K.; Kumar, R.; Kumar, P.; Pandohee, J.; Kulshresha, S.; Harjai, K.; Chhibber, S. Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria. Molecules 2022, 27, 7584. [Google Scholar] [CrossRef]
- Nett, J.E.; Andes, D.R. Contributions of the Biofilm Matrix to Candida Pathogenesis. J. Fungi 2020, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.D.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Maisetta, G.; Batoni, G. Editorial: Interspecies Interactions: Effects on Virulence and Antimicrobial Susceptibility of Bacterial and Fungal Pathogens. Front. Microbiol. 2020, 11, 01922. [Google Scholar] [CrossRef]
- Wei, D.; Zhu, X.-M.; Chen, Y.-Y.; Li, X.-Y.; Chen, Y.-P.; Liu, H.-Y.; Zhang, M. Chronic wound biofilms: Diagnosis and therapeutic strategies. Chin. Med. J. 2019, 132, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.; Burgess, J.L.; Verpile, R.; Tomic-Canic, M.; Pastar, I. Novel Diagnostic Technologies and Therapeutic Approaches Targeting Chronic Wound Biofilms and Microbiota. Curr. Dermatol. Rep. 2022, 11, 60–72. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: Biotechnological applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef]
- Lu, J.; Cokcetin, N.N.; Burke, C.M.; Turnbull, L.; Liu, M.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Honey can inhibit and eliminate biofilms produced by Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 18160. [Google Scholar] [CrossRef] [Green Version]
- Di Lodovico, S.; Menghini, L.; Ferrante, C.; Recchia, E.; Castro-Amorim, J.; Gameiro, P.; Cellini, L.; Bessa, L.J. Hop Extract: An Efficacious Antimicrobial and Anti-biofilm Agent Against Multidrug-Resistant Staphylococci Strains and Cutibacterium acnes. Front. Microbiol. 2020, 11, 1852. [Google Scholar] [CrossRef] [PubMed]
- Thaarup, I.C.; Bjarnsholt, T. Current In Vitro Biofilm-Infected Chronic Wound Models for Developing New Treatment Possibilities. Adv. Wound Care 2021, 10, 91–102. [Google Scholar] [CrossRef]
- Besser, M.; Terberger, J.; Weber, L.; Ghebremedhin, B.; Naumova, E.A.; Arnold, W.H.; Stuermer, E.K. Impact of probiotics on pathogen survival in an innovative human plasma biofilm model (hpBIOM). J. Transl. Med. 2019, 17, 243. [Google Scholar] [CrossRef] [Green Version]
- Besser, M.; Dietrich, M.; Weber, L.; Rembe, J.D.; Stuermer, E.K. Efficacy of antiseptics in a novel 3-dimensional human plasma biofilm model (hpBIOM). Sci. Rep. 2020, 10, 4792. [Google Scholar] [CrossRef] [Green Version]
- Stuermer, E.K.; Plattfaut, I.; Dietrich, M.; Brill, F.; Kampe, A.; Wiencke, V.; Ulatowski, A.; Geffken, M.; Rembe, J.-D.; Naumova, E.A.; et al. In vitro Activity of Antimicrobial Wound Dressings on P. aeruginosa Wound Biofilm. Front. Microbiol. 2021, 12, 664030. [Google Scholar] [CrossRef]
- Besser, M.; Schaeler, L.; Plattfaut, I.; Brill, F.H.; Kampe, A.; Geffken, M.; Smeets, R.; Debus, E.S.; Stuermer, E.K. Pulsed low-intensity laser treatment stimulates wound healing without enhancing biofilm development in vitro. J. Photochem. Photobiol. B Biol. 2022, 233, 112504. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lorenzen, J.; Xu, Y.; Jonikaite, M.; Thaarup, I.C.; Bjarnsholt, T.; Kirketerp-Møller, K.; Thomsen, T.R. A novel chronic wound biofilm model sustaining coexistence of Pseudomonas aeruginosa and Staphylococcus aureus suitable for testing of antibiofilm effect of antimicrobial solutions and wound dressings. Wound Repair Regen. 2021, 29, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Dowd, S.E.; Smith, E.; Rhoads, D.; Wolcott, R.D. In vitro multispecies Lubbock chronic wound biofilm model. Wound Repair Regen. 2008, 16, 805–813. [Google Scholar] [CrossRef]
- Hotterbeekx, A.; Kumar-Singh, S.; Goossens, H.; Malhotra-Kumar, S. In vivo and In vitro Interactions between Pseudomonas aeruginosa and Staphylococcus spp. Front. Cell. Infect. Microbiol. 2017, 7, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.C.; Rice, A.; Sutton, B.; Gabrilska, R.; Wessel, A.K.; Whiteley, M.; Rumbaugh, K.P. Albumin Inhibits Pseudomonas aeruginosa Quorum Sensing and Alters Polymicrobial Interactions. Infect. Immun. 2017, 85, e00116-17. [Google Scholar] [CrossRef] [Green Version]
- DeLeon, S.; Clinton, A.; Fowler, H.; Everett, J.; Horswill, A.R.; Rumbaugh, K.P. Synergistic Interactions of Pseudomonas aeruginosa and Staphylococcus aureus in an In Vitro Wound Model. Infect. Immun. 2014, 82, 4718–4728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An In Vivo Polymicrobial Biofilm Wound Infection Model to Study Interspecies Interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef] [Green Version]
- Klein, P.; Sojka, M.; Kucera, J.; Matonohova, J.; Pavlik, V.; Nemec, J.; Kubickova, G.; Slavkovsky, R.; Szuszkiewicz, K.; Danek, P.; et al. A porcine model of skin wound infected with a polybacterial biofilm. Biofouling 2018, 34, 226–236. [Google Scholar] [CrossRef]
- Staneviciute, E.; Na’Amnih, W.; Kavaliauskas, P.; Prakapaite, R.; Ridziauskas, M.; Kevlicius, L.; Kirkliauskiene, A.; Zabulis, V.; Urboniene, J.; Triponis, V. New in vitro model evaluating antiseptics’ efficacy in biofilm-associated Staphylococcus aureus prosthetic vascular graft infection. J. Med. Microbiol. 2019, 68, 432–439. [Google Scholar] [CrossRef]
- Kucera, J.; Sojka, M.; Pavlik, V.; Szuszkiewicz, K.; Velebny, V.; Klein, P. Multispecies biofilm in an artificial wound bed—A novel model for in vitro assessment of solid antimicrobial dressings. J. Microbiol. Methods 2014, 103, 18–24. [Google Scholar] [CrossRef]
- Pouget, C.; Gustave, C.-A.; Ngba-Essebe, C.; Laurent, F.; Lemichez, E.; Tristan, A.; Sotto, A.; Dunyach-Rémy, C.; Lavigne, J.-P. Adaptation of Staphylococcus aureus in a Medium Mimicking a Diabetic Foot Environment. Toxins 2021, 13, 230. [Google Scholar] [CrossRef]
- Pouget, C.; Dunyach-Remy, C.; Bernardi, T.; Provot, C.; Tasse, J.; Sotto, A.; Lavigne, J.-P. A Relevant Wound-Like in vitro Media to Study Bacterial Cooperation and Biofilm in Chronic Wounds. Front. Microbiol. 2022, 13, 705479. [Google Scholar] [CrossRef]
- Pouget, C.; Dunyach-Remy, C.; Pantel, A.; Schuldiner, S.; Sotto, A.; Lavigne, J.-P. New Adapted In Vitro Technology to Evaluate Biofilm Formation and Antibiotic Activity Using Live Imaging under Flow Conditions. Diagnostics 2021, 11, 1746. [Google Scholar] [CrossRef]
- Di Lodovico, S.; Bacchetti, T.; D’Ercole, S.; Covone, S.; Petrini, M.; Di Giulio, M.; Di Fermo, P.; Diban, F.; Ferretti, G.; Cellini, L. Complex Chronic Wound Biofilms Are Inhibited in vitro by the Natural Extract of Capparis spinose. Front. Microbiol. 2022, 13, 832919. [Google Scholar] [CrossRef] [PubMed]
- Di Fermo, P.; Ciociola, T.; Di Lodovico, S.; D’Ercole, S.; Petrini, M.; Giovati, L.; Conti, S.; Di Giulio, M.; Cellini, L. Antimicrobial Peptide L18R Displays a Modulating Action against Inter-Kingdom Biofilms in the Lubbock Chronic Wound Biofilm Model. Microorganisms 2021, 9, 1779. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Smith, E.; Wolcott, R.; Dowd, S. Propagation of anaerobic bacteria within an aerobic multi-species chronic wound biofilm model. J. Wound Care 2009, 18, 426–431. [Google Scholar] [CrossRef]
- Dowd, S.; Sun, Y.; Smith, E.; Kennedy, J.; Jones, C.; Wolcott, R. Effects of biofilm treatments on the multi-species Lubbock chronic wound biofilm model. J. Wound Care 2009, 18, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Di Giulio, M.; Di Lodovico, S.; Fontana, A.; Traini, T.; Di Campli, E.; Pilato, S.; D’Ercole, S.; Cellini, L. Graphene Oxide affects Staphylococcus aureus and Pseudomonas aeruginosa dual species biofilm in Lubbock Chronic Wound Biofilm model. Sci. Rep. 2020, 10, 18525. [Google Scholar] [CrossRef]
- Sojka, M.; Valachova, I.; Bucekova, M.; Majtan, J. Antibiofilm efficacy of honey and bee-derived defensin-1 on multispecies wound biofilm. J. Med. Microbiol. 2016, 65, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Sanford, N.E.; Wilkinson, J.E.; Nguyen, H.; Diaz, G.; Wolcott, R. Efficacy of hyperbaric oxygen therapy in bacterial biofilm eradication. J. Wound Care 2018, 27, S20–S28. [Google Scholar] [CrossRef]
- Di Lodovico, S.; Diban, F.; Di Fermo, P.; Petrini, M.; Fontana, A.; Di Giulio, M.; Piattelli, A.; D’Ercole, S.; Cellini, L. Antimicrobial Combined Action of Graphene Oxide and Light Emitting Diodes for Chronic Wound Management. Int. J. Mol. Sci. 2022, 23, 6942. [Google Scholar] [CrossRef]
- Redman, W.K.; Welch, G.S.; Rumbaugh, K.P. Differential Efficacy of Glycoside Hydrolases to Disperse Biofilms. Front. Cell Infect. Microbiol. 2020, 10, 379. [Google Scholar] [CrossRef]
- Aunkor, M.T.H.; Raihan, T.; Prodhan, S.H.; Metselaar, H.S.C.; Malik, S.U.F.; Azad, A.K. Antibacterial Activity of Graphene Oxide Nanosheet against Multidrug Resistant Superbugs Isolated from Infected Patients. R. Soc. Open Sci. 2020, 7, 200640. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.U.; Fleming, D.F.; Wang, L.; Rumbaugh, K.P.; Gordon, V.D.; Christopher, G.F. Microrheology of Pseudomonas aeruginosa biofilms grown in wound beds. NPJ Biofilms Microbiomes 2022, 8, 1–9. [Google Scholar] [CrossRef]
- Pirlar, R.F.; Emaneini, M.; Beigverdi, R.; Banar, M.; van Leeuwen, W.B.; Jabalameli, F. Combinatorial effects of antibiotics and enzymes against dual-species Staphylococcus aureus and Pseudomonas aeruginosa biofilms in the wound-like medium. PLoS ONE 2020, 15, e0235093. [Google Scholar] [CrossRef]
- Versey, Z.; da Cruz Nizer, W.S.; Russell, E.; Zigic, S.; DeZeeuw, K.G.; Marek, J.E.; Overhage, J.; Cassol, E. Biofilm-Innate Immune Interface: Contribution to Chronic Wound Formation. Front. Immunol. 2021, 12, 648554. [Google Scholar] [CrossRef] [PubMed]
- Kirketerp-Møller, K.; Stewart, P.S.; Bjarnsholt, T. The zone model: A conceptual model for understanding the microenvironment of chronic wound infection. Wound Repair Regen. 2020, 28, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Schultz, G.; Bjarnsholt, T.; James, G.A.; Leaper, D.J.; McBain, A.J.; Malone, M.; Stoodley, P.; Swanson, T.; Tachi, M.; Wolcott, R.D.; et al. Consensus guidelines for the identification and treatment of biofilms in chronic nonhealing wounds. Wound Repair Regen. 2017, 25, 744–757. [Google Scholar] [CrossRef]
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Nakagami, G.; Kitamura, A.; Minematsu, T.; Kinoshita, M.; Suga, H.; Kurita, M.; Hayashi, C.; Kawasaki, A.; Sanada, H. Effectiveness of biofilm-based wound care system on wound healing in chronic wounds. Wound Repair Regen. 2019, 27, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Attinger, C.; Wolcott, R. Clinically Addressing Biofilm in Chronic Wounds. Adv. Wound Care 2012, 1, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Alves, P.J.; Barreto, R.T.; Barrois, B.M.; Gryson, L.G.; Meaume, S.; Monstrey, S.J. Update on the role of antiseptics in the management of chronic wounds with critical colonisation and/or biofilm. Int. Wound J. 2020, 18, 342–358. [Google Scholar] [CrossRef]
- Leaper, D.; Assadian, O.; Edmiston, C. Approach to chronic wound infections. Br. J. Dermatol. 2015, 173, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Lipsky, B.A.; Hoey, C. Topical Antimicrobial Therapy for Treating Chronic Wounds. Clin. Infect. Dis. 2009, 49, 1541–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalim, W.; Mwesigye, J.; Tungotyo, M.; Twinomujuni, S.S. Resistance pattern of infected chronic wound isolates and factors associated with bacterial resistance to third generation cephalosporins at Mbarara Regional Referral Hospital, Uganda. PLoS ONE 2021, 16, e0261264. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Batoni, G.; Maisetta, G.; Esin, S. Therapeutic Potential of Antimicrobial Peptides in Polymicrobial Biofilm-Associated Infections. Int. J. Mol. Sci. 2021, 22, 482. [Google Scholar] [CrossRef]
- Maisetta, G.; Grassi, L.; Esin, S.; Serra, I.; Scorciapino, M.A.; Rinaldi, A.C.; Batoni, G. The Semi-Synthetic Peptide Lin-SB056-1 in Combination with EDTA Exerts Strong Antimicrobial and Antibiofilm Activity against Pseudomonas aeruginosa in Conditions Mimicking Cystic Fibrosis Sputum. Int. J. Mol. Sci. 2017, 18, 1994. [Google Scholar] [CrossRef] [PubMed]
- Niculescu, A.-G.; Grumezescu, A. Photodynamic Therapy—An Up-to-Date Review. Appl. Sci. 2021, 11, 3626. [Google Scholar] [CrossRef]
- Nesi-Reis, V.; Lera-Nonose, D.S.S.L.; Oyama, J.; Lalucci, M.; Demarchi, I.G.; Aristides, S.M.A.; Teixeira, J.J.V.; Silveira, T.G.V.; Lonardoni, M.V.C. Contribution of photodynamic therapy in wound healing: A systematic review. Photodiagnosis Photodyn. Ther. 2017, 21, 294–305. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Shanmugam, R.; Lee, J.-K. Polyhydroxyalkanoates: Trends and advances toward biotechnological applications. Bioresour. Technol. 2021, 326, 124737. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment in LCWB | Pathogens in LCWB | Results | Reference |
|---|---|---|---|
| Bleach (6% sodium hypochlorite) triclosan gallium nitrate | P. aeruginosa E. faecalis S. aureus 24 h incubation | In planktonic cells, 1% bleach solution was required for 100% disinfection, while multispecies biofilm withstood 50% concentration; Triclosan inhibited biofilm formation at 1, 10 and 100 ppm concentrations with a selective inhibitory effect on S. aureus; Gallium inhibited biofilm formation at 1 μM with a selective inhibitory effect on P. aeruginosa. | Sun et al., 2008 [53] |
| Anaerobic bacteria integration in LCWB model | P. aeruginosa E. faecalis S. aureus | Clostridium perfringens, Peptoniphilus ivorii and Anaerococcus lactolyticus showed a slight growth in biofilm after 24 h and a greater integration of in biofilm after 48 h; Peptostreptococcus anaerobius showed integration in biofilm after both 24 and 48 h; The same result was detected with Finegoldia magna, which became dominant biofilm after 48 h. | Sun et al., 2009 [66] |
| Several biofilm effectors | P. aeruginosa E. faecalis S. aureus | 20% xylitol, 10% erythritol, 1000 µg/mL farnesol, 20 mM salicylic acid or 0.1% of either of the two gel formulations were able to inhibit biofilm formation | Dowd et al., 2009 [67] |
| Polyvinylpyrroli- done–iodine complex Cadexomer–iodine complex | P. aeruginosa E. faecalis S. aureus Bacillus subtilis 48 h incubation | Cadexomer–iodine (1.8 mg I2/cm2) reduced bacterial count in the biofilm (5 log reduction in S. aureus and B. subtilis, 9 log reduction in P. aeruginosa and E. faecalis); Both (PVP iodine and cadexomer-iodine) were ineffective at the concentration of 0.2 mg I2/cm2. | Kucera et al., 2014 [60] |
| Gentamicin Ciprofloxacin Tetracycline | P. aeruginosa S. aureus | The poly-microbial growth of these strains showed a synergistic effect regarding the antimicrobial tolerance in LCWB compared to the planktonic culture, showing the role of host derived matrix in anti-microbial tolerance enhancement; The deletion of the ica gene in S.aureus and the algD gene in P. aeruginosa decreased the tolerance of the coculture in comparison to the wild-type of both microorganisms. | DeLeon et al., 2014 [56] |
| Capparis spinose aqueous extract | P. aeruginosa S. aureus 48 h incubation | Capparis spinose aqueous extract reduced the LCWB formation by 97.32% and 99.67% for resistant S. aureus and P. aeruginosa strains, respectively. | Di Lodovico et al., 2022 [64] |
| Octenidine Dihydrochloride 0.1% Povidone–iodine 10% Chlorhexidine digluconate 0.02% | S. aureus monomicrobial biofilm in LCWB medium loaded into a prosthetic vascular graft infection (PVGI) model | All antiseptics demonstrated significant antimicrobial efficacy, decreasing colony counts, with the superiority of Octenidine against S. aureus biofilms grown on vascular graft (7 orders of magnitude CFU reduction); Chlorhexidine worked best against S. aureus biofilms integrity on glass coverslips and decreased the surface area covered with S. aureus from 73.75 to 10.55%. | Staneviciute et al., 2019 [59] |
| Graphene oxide 50 mg/L | P. aeruginosa S. aureus | Graphene oxide showed an antibiofilm effect disrupting the fibrin network and reducing the CFU/mg by up to 70.24% and 59.31% for S. aureus and P. aeruginosa, respectively. | Di Giulio et al., 2020 [68] |
| Manuka honey 100% Honeydew honey 100% Honey recombinant Defensin-1 (Def-1) 0.1 and 1 mg/mL | S. aureus, Streptococcus agalactiae, P. aeruginosa, E. faecalis | Both types of honey reduced cell viability of S. aureus (by a 4 log reduction), S. agalactiae (5 log reduction) and P. aeruginosa (5 log reduction) but showed no effect against E. faecalis; Def-1 reduced the viability of S. aureus (5 log reduction at both concentrations) and P. aeruginosa (2 log reduction at 0.1 mg/mL and 4 log reduction at 1 mg/mL); Def-1 inhibited biofilm formation of E. faecalis and S. agalactiae at both concentrations. | Sojka et al., 2016 [69] |
| Hyperbaric oxygen therapy (HBOT) | P. aeruginosa, E. faecalis, S. aureus | Treatment with HBOT showed a slight but significant reduction of the viability of the three bacterial species after 30 and 90 min of application in vitro. | Sanford et al., 2018 [70] |
| Antimicrobial Peptide L18R 100 µg/ml | S. aureus P. aeruginosa C. albicans | L18R showed antimicrobial activity against all strains in planktonic form (especially C. albicans); L18R reduced biofilm formation of C. albicans (97.19% and 98.81% reduction in biofilm biomass for early stage and mature biofilms, respectively), with less effect against the bacterial strains; L18R did not affect the dual and triadic poly-microbial biofilms. | Di Fermo et al., 2021 [65] |
| Graphene oxide (50 mg/L) and 5-aminolevulinic acid (ALAD) mediated photodynamic therapy (PDT) therapy | S. aureus P. aeruginosa | The most effective combination was graphene oxide (50 mg/L) application followed by ALAD-PDT (630 nm) with reductions in S. aureus (78.96%) and P. aeruginosa (85.67%); Additionally, the application of graphene oxide (50 mg/L) for 24 h incubation, followed by ALAD-PDT exhibited a CFU/mg reduction in P. aeruginosa (95.17%); Other conditions reduced the CFU count with different percentages. | Di Lodovico et al., 2022 [71] |
| Glycoside hydrolases, GH, (α-amilase and cellulase, to target the EPS of biofilm) | S. aureus SA31 P. aeruginosa PAO1 | After 48–96 h of growth, the dual species biofilm was treated for 1 h with either 1× PBS, 2.5% amylase, 2.5% cellulase, or both (5% GH); Amylase significantly dispersed both P. aeruginosa and S. aureus; The ability of cellulase to disperse S. aureus was completely abated as well as to disperse P. aeruginosa; One possible explanation for this result is that the activity of cellulase is inhibited by proteolytic blood components in the microcosm model and ex vivo tissue. | Redman et al., 2020 [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diban, F.; Di Lodovico, S.; Di Fermo, P.; D’Ercole, S.; D’Arcangelo, S.; Di Giulio, M.; Cellini, L. Biofilms in Chronic Wound Infections: Innovative Antimicrobial Approaches Using the In Vitro Lubbock Chronic Wound Biofilm Model. Int. J. Mol. Sci. 2023, 24, 1004. https://doi.org/10.3390/ijms24021004
Diban F, Di Lodovico S, Di Fermo P, D’Ercole S, D’Arcangelo S, Di Giulio M, Cellini L. Biofilms in Chronic Wound Infections: Innovative Antimicrobial Approaches Using the In Vitro Lubbock Chronic Wound Biofilm Model. International Journal of Molecular Sciences. 2023; 24(2):1004. https://doi.org/10.3390/ijms24021004
Chicago/Turabian StyleDiban, Firas, Silvia Di Lodovico, Paola Di Fermo, Simonetta D’Ercole, Sara D’Arcangelo, Mara Di Giulio, and Luigina Cellini. 2023. "Biofilms in Chronic Wound Infections: Innovative Antimicrobial Approaches Using the In Vitro Lubbock Chronic Wound Biofilm Model" International Journal of Molecular Sciences 24, no. 2: 1004. https://doi.org/10.3390/ijms24021004