
F4-Neuroprostane Effects on Human Sperm
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. DNA Integrity Assessment with Acridine Orange (AO) Staining
2.2. Mitochondrial Membrane Potential (MMP) Assessment with JC-1 Labeling
2.3. Pisum Sativum Agglutinin (PSA) Evaluation
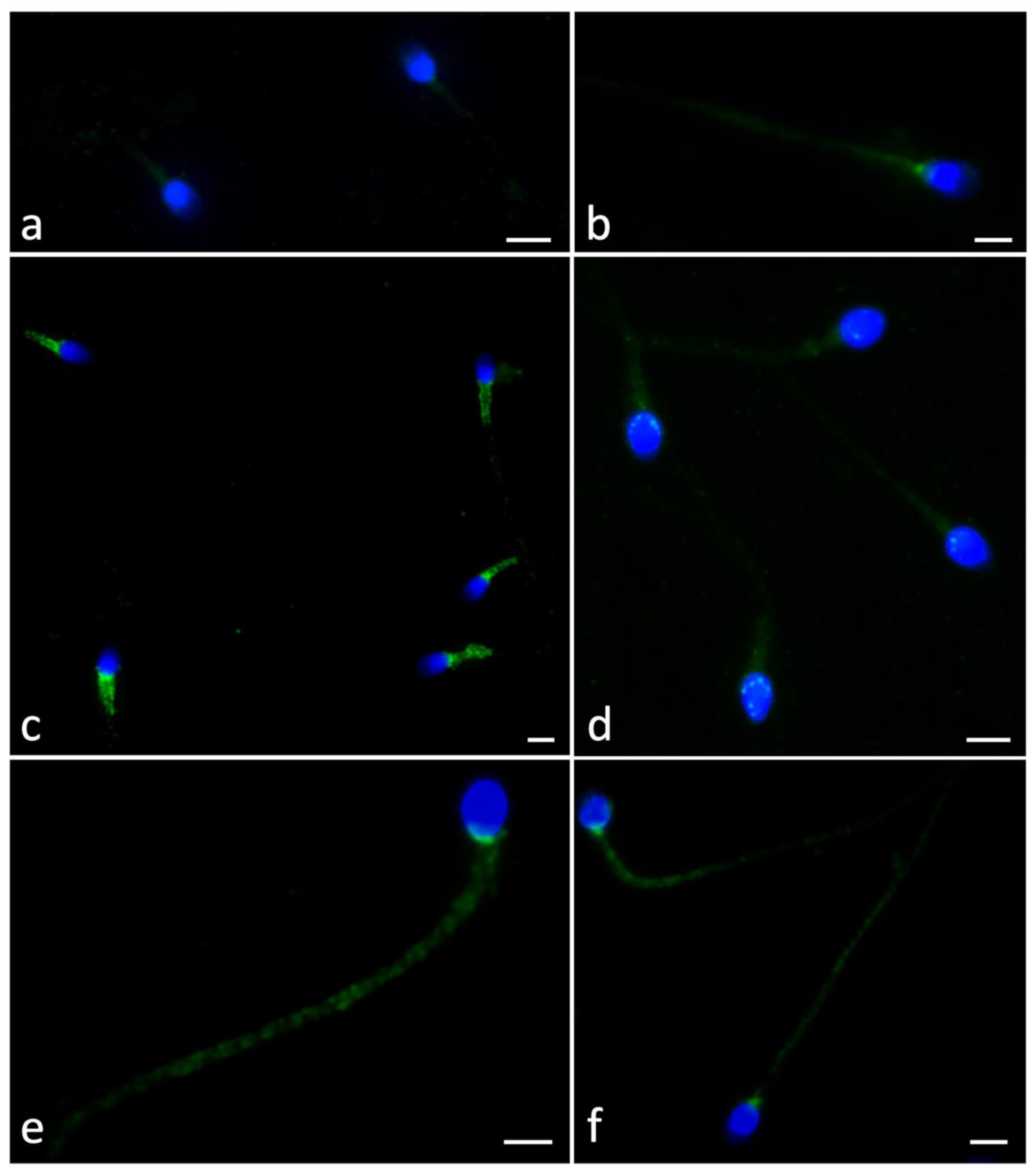
2.4. Ryanodine Receptor Localization
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Samples
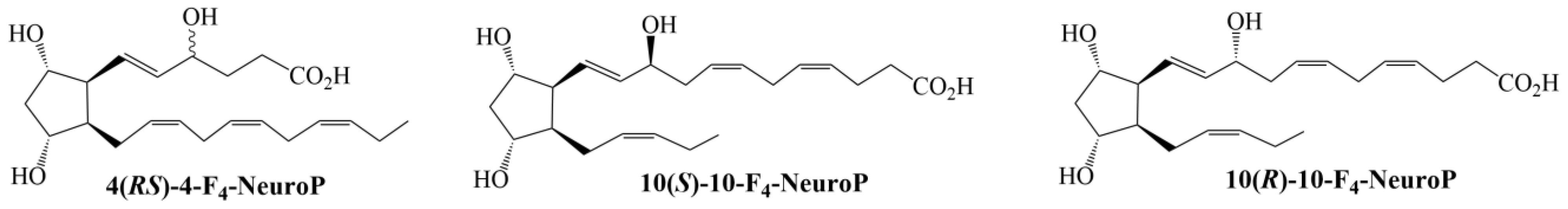
5.2. Chemical Structures of 4(RS)-4-F4t-NeuroPs, 10(S)-10-F4t-NeuroP and 10(R)-10-F4t-NeuroP
5.3. Swim-Up Technique on Human Semen Samples and Incubation in Presence of F4-NeuroPs
- -
- 7 ng F4-NeuroPs for 2 h
- -
- 7 ng F4-NeuroPs for 4 h
5.4. Acridine Orange Assay
5.5. JC-1 Dye
5.6. Pisum Sativum Agglutinin
5.7. Immunofluorescence of Ryanodine Receptor
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Milne, G.L.; Dai, Q.; Roberts, L.J. The isoprostanes—25 years later. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 433–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, O.S.; Galano, J.M.; Pavlickova, T.; Révol-Cavalier, T.J.; Vigor, C.; Lee, J.C.Y.; Oger, C.; Durand, T. Moving forward with isoprostanes, neuroprostanes and phytoprostanes—Where are we know? Essays Biochem. 2020, 64, 463–484. [Google Scholar] [PubMed]
- Geng, X.; Galano, J.-M.; Oger, C.; Sun, G.Y.; Durand, T.; Lee, J.C. Neuroprotective effects of DHA-derived peroxidation product 4(RS)-4-F4t-neuroprostane on microglia. Free. Radic. Biol. Med. 2022, 185, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.; Morel, A.; Saso, L.; Saluk, J. Isoprostanes and Neuroprostanes as Biomarkers of Oxidative Stress in Neurodegenerative Diseases. Oxidative Med. Cell. Longev. 2014, 2014, 572491. [Google Scholar] [CrossRef] [PubMed]
- Signorini, C.; De Felice, C.; Galano, J.-M.; Oger, C.; Leoncini, S.; Cortelazzo, A.; Ciccoli, L.; Durand, T.; Hayek, J.; Lee, J.C.-Y. Isoprostanoids in Clinical and Experimental Neurological Disease Models. Antioxidants 2018, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Syta-Krzyżanowska, A.; Jarocka-Karpowicz, I.; Kochanowicz, J.; Turek, G.; Rutkowski, R.; Gorbacz, K.; Mariak, Z.; Skrzydlewska, E. F2-isoprostanes and F4-neuroprostanes as markers of intracranial aneurysm development. Adv. Clin. Exp. Med. 2018, 27, 673–680. [Google Scholar] [CrossRef]
- Gladine, C.; Newman, J.W.; Durand, T.; Pedersen, T.L.; Galano, J.-M.; Demougeot, C.; Berdeaux, O.; Pujos-Guillot, E.; Mazur, A.; Comte, B. Lipid Profiling following Intake of the Omega 3 Fatty Acid DHA Identifies the Peroxidized Metabolites F4-Neuroprostanes as the Best Predictors of Atherosclerosis Prevention. PLoS ONE 2014, 9, e89393. [Google Scholar] [CrossRef]
- Roy, J.; Oger, C.; Thireau, J.; Roussel, J.; Mercier-Touzet, O.; Faure, D.; Pinot, E.; Farah, C.; Taber, D.F.; Cristol, J.P.; et al. Nonenzymatic lipid mediators neuroprostanes exert the anti-arrhythmic properties of docosahexenoic acid. Free. Radic. Biol. Med. 2015, 86, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Roy, J.; Fauconnier, J.; Oger, C.; Demion, M.; Farah, C.; Angbault-Prouteau, C.; Bideaux, P.; Scheuermann, V.; Bultel-Poncé, V.; Lee, J.C.Y.; et al. Non-enzymatic oxidized metabolite of DHA, 4(RS)-4-F4t neuroprostane protects the heart against reperfusion injuries. Free. Radic. Biol. Med. 2017, 102, 229–239. [Google Scholar] [CrossRef]
- Seet, R.C.; Lee, J.C.-Y.; Chan, B.P.; Sharma, V.K.; Teoh, H.-L.; Venketasubramanian, N.; Lim, E.C.; Chong, W.-L.; Looi, W.-F.; Huang, S.-H.; et al. Oxidative Damage in Ischemic Stroke Revealed Using Multiple Biomarkers. Stroke 2011, 42, 2326–2329. [Google Scholar] [CrossRef]
- Galano, J.-M.; Lee, Y.Y.; Oger, C.; Vigor, C.; Vercauteren, J.; Durand, T.; Giera, M.; Lee, J.C.-Y. Isoprostanes, neuroprostanes and phytoprostanes: An overview of 25 years of research in chemistry and biology. Prog. Lipid Res. 2017, 68, 83–108. [Google Scholar] [CrossRef] [PubMed]
- Medina, S.; De Miguel-Elízaga, I.; Oger, C.; Galano, J.-M.; Durand, T.; Martínez-Villanueva, M.; Castillo, M.L.G.-D.; Villegas-Martínez, I.; Ferreres, F.; Martínez-Hernández, P.; et al. Dihomo-isoprostanes—Nonenzymatic metabolites of AdA—Are higher in epileptic patients compared to healthy individuals by a new ultrahigh pressure liquid chromatography–triple quadrupole–tandem mass spectrometry method. Free Radic. Biol. Med. 2015, 79, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Longini, M.; Moretti, E.; Signorini, C.; Noto, D.; Iacoponi, F.; Collodel, G. Relevance of seminal F2-dihomo-IsoPs, F2-IsoPs and F4-NeuroPs in idiopathic infertility and varicocele. Prostaglandins Other Lipid Mediat. 2020, 149, 106448. [Google Scholar] [CrossRef] [PubMed]
- Signorini, C.; Moretti, E.; Noto, D.; Mattioli, S.; Castellini, C.; Pascarelli, N.; Durand, T.; Oger, C.; Galano, J.-M.; De Felice, C.; et al. F4-Neuroprostanes: A Role in Sperm Capacitation. Life 2021, 11, 655. [Google Scholar] [CrossRef]
- Deng, S.-L.; Sun, T.-C.; Wang, J.-H.; Wang, X.-X.; Liu, X.-M.; Zhang, C.-L.; Hao, C.-F.; Ma, W.-Z.; Liu, Y.-X. Effects of sperm proteins on fertilization in the female reproductive tract. Front. Biosci. 2019, 24, 735–749. [Google Scholar] [CrossRef]
- Vaquer, C.C.; Suhaiman, L.; Pavarotti, M.A.; De Blas, G.A.; Belmonte, S.A. Ceramide induces a multicomponent intracellular calcium increase triggering the acrosome secretion in human sperm. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2020, 1867, 118704. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; WHO Press: Geneva, Switzerland, 2021. Available online: https://www.who.int/publications/i/item/9789240030787 (accessed on 27 July 2021).
- Gallo, A.; Esposito, M.; Tosti, E.; Boni, R. Sperm Motility, Oxidative Status, and Mitochondrial Activity: Exploring Correlation in Different Species. Antioxidants 2021, 10, 1131. [Google Scholar] [CrossRef]
- Agnihotri, S.K.; Agrawal, A.K.; Hakim, B.A.; Vishwakarma, A.L.; Narender, T.; Sachan, R.; Sachdev, M. Mitochondrial membrane potential (MMP) regulates sperm motility. Vitr. Cell. Dev. Biol.-Anim. 2016, 52, 953–960. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, W.; Zou, P.; Jiang, F.; Zeng, Y.; Chen, Q.; Sun, L.; Yang, H.; Zhou, N.; Wang, X.; et al. Mitochondrial functionality modifies human sperm acrosin activity, acrosome reaction capability and chromatin integrity. Hum. Reprod. 2019, 34, 3–11. [Google Scholar] [CrossRef]
- Galano, J.-M.; Roy, J.; Durand, T.; Lee, J.C.-Y.; Le Guennec, J.-Y.; Oger, C.; Demion, M. Biological activities of non-enzymatic oxygenated metabolites of polyunsaturated fatty acids (NEO-PUFAs) derived from EPA and DHA: New anti-arrhythmic compounds? Mol. Asp. Med. 2018, 64, 161–168. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Galano, J.; Leung, H.H.; Balas, L.; Oger, C.; Durand, T.; Lee, J.C. Nonenzymatic oxygenated metabolite of docosahexaenoic acid, 4(RS)-4-F4t-neuroprostane, acts as a bioactive lipid molecule in neuronal cells. FEBS Lett. 2020, 594, 1797–1808. [Google Scholar] [CrossRef] [PubMed]
- Molnár, P.J.; Dér, B.; Borsodi, K.; Balla, H.; Borbás, Z.; Molnár, K.; Ruisanchez, E.; Kenessey, I.; Horváth, A.; Keszthelyi, A.; et al. Isoprostanes evoke contraction of the murine and human detrusor muscle via activation of the thromboxane prostanoid TP receptor and Rho kinase. Am. J. Physiol. Physiol. 2021, 320, F537–F547. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Espinosa, P.; López-Huedo, A.; Robles-Gómez, L.; Huerta-Retamal, N.; Aizpurua, J.; Gómez-Torres, M.J. Characterization of Human Spermatic Subpopulations by ConA-Binding Sites and Tyrosine Phosphorylation during in vitro Capacitation and Acrosome Reaction. Cells Tissues Organs 2021, 210, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sosa, C.; Pavarotti, M.; Zanetti, M.; Zoppino, F.C.M.; De Blas, G.; Mayorga, L. Kinetics of human sperm acrosomal exocytosis. Mol. Hum. Reprod. 2014, 21, 244–254. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, C.; de Lamirande, E.; Gagnon, C. Phosphorylation of the Arginine-X-X-(Serine/Threonine) motif in human sperm proteins during capacitation: Modulation and protein kinase A dependency. Mol. Hum. Reprod. 2004, 10, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Bragado, M.; Garcia-Marin, L.; Calle-Guisado, V.; de Llera, A.; Martin-Hidalgo, D.; Mijares, J.; Gil, M.; Alvarez, I. AMP-activated kinase in human spermatozoa: Identification, intracellular localization, and key function in the regulation of sperm motility. Asian J. Androl. 2017, 19, 707–714. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Cardona, C.; Moody, M.A.; Simpson, A.J.; Mendoza, R.; Seaman, E.; Travis, A.J. Timing of sperm capacitation varies reproducibly among men. Mol. Reprod. Dev. 2018, 85, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Martínez, G.C.; Aldana, A.; Carneiro, J.; Treviño, C.L.; Darszon, A. Acrosomal alkalinization occurs during human sperm capacitation. Mol. Hum. Reprod. 2022, 28, gaac005. [Google Scholar] [CrossRef]
- Gaburjakova, J.; Krejciova, E.; Gaburjakova, M. Multisite phosphorylation of the cardiac ryanodine receptor: A random or coordinated event? Pflüg. Arch.-Eur. J. Physiol. 2020, 472, 1793–1807. [Google Scholar] [CrossRef]
- Zhou, Y.; Ru, Y.; Wang, C.; Wang, S.; Zhou, Z.; Zhang, Y. Tripeptidyl Peptidase II Regulates Sperm Function by Modulating Intracellular Ca2+ Stores via the Ryanodine Receptor. PLoS ONE 2013, 8, e66634. [Google Scholar] [CrossRef]
- Collodel, G.; Moretti, E.; Noto, D.; Corsaro, R.; Signorini, C. Oxidation of Polyunsaturated Fatty Acids as a Promising Area of Research in Infertility. Antioxidants 2022, 11, 1002. [Google Scholar] [CrossRef] [PubMed]
- Zerbinati, C.; Caponecchia, L.; Rago, R.; Leoncini, E.; Bottaccioli, A.G.; Ciacciarelli, M.; Pacelli, A.; Salacone, P.; Sebastianelli, A.; Pastore, A.; et al. Fatty acids profiling reveals potential candidate markers of semen quality. Andrology 2016, 4, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, Y.; Aksoy, H.; Altınkaynak, K.; Aydın, H.R.; Özkan, A. Sperm fatty acid composition in subfertile men. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soto, J.C.; Domingo, J.C.; Cordobilla, B.; Nicolás, M.; Fernández, L.; Albero, P.; Gadea, J.; Landeras, J. Dietary supplementation with docosahexaenoic acid (DHA) improves seminal antioxidant status and decreases sperm DNA fragmentation. Syst. Biol. Reprod. Med. 2016, 62, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oger, C.; Brinkmann, Y.; Bouazzaoui, S.; Durand, T.; Galano, J.-M. Stereocontrolled Access to Isoprostanes via a Bicyclo[3.3.0]octene Framework. Org. Lett. 2008, 10, 5087–5090. [Google Scholar] [CrossRef] [PubMed]
- Oger, C.; Bultel-Poncé, V.; Guy, A.; Balas, L.; Rossi, J.-C.; Durand, T.; Galano, J.-M. The Handy Use of Brown’s P2-Ni Catalyst for a Skipped Diyne Deuteration: Application to the Synthesis of a [D4]-Labeled F4t-Neuroprostane. Chem.-A Eur. J. 2010, 16, 13976–13980. [Google Scholar] [CrossRef] [PubMed]
- Tejada, R.I.; Mitchell, J.C.; Norman, A.; Marik, J.J.; Friedman, S. A test for the practical evaluation of male fertility by acridine orange (AO) fluorescence. Fertil. Steril. 1984, 42, 87–91. [Google Scholar] [CrossRef]
- Pascarelli, N.A.; Moretti, E.; Terzuoli, G.; Lamboglia, A.; Renieri, T.; Fioravanti, A.; Collodel, G. Effects of gold and silver nanoparticles in cultured human osteoarthritic chondrocytes. J. Appl. Toxicol. 2013, 33, 1506–1513. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control (2 h) | F4-NeuroPs (2 h) | Control (4 h) | F4-NeuroPs (4 h) | |
|---|---|---|---|---|
| Sperm progressive motility% | 44 (42–46) | 56 (52–64) | 42 (40–43) | 44 (42–48) |
| Statistics | p = 0.02 | p = 0.119 | ||
| Control 2 h Red Spot | F4-NeuroPs 2 h Red Spot | Control 2 h Red Spot Strong Signal | F4-NeuroPs 2 h Red Spot Strong Signal | Control 4 h Red Spot | F4-NeuroPs 4 h Red Spot | Control 4 h Red Spot Strong Signal | F4-NeuroPs 4 h Red Spot Strong Signal | |
|---|---|---|---|---|---|---|---|---|
| J-C1-labeled sperm% | 68 (65–74) | 78 (74–79) | 26 (24–28) | 50 (48–52) | 44 (42–46) | 42 (42–43) | 18 (16–18) | 20 (18–21) |
| Statistics | p = 0.012 | p = 0.002 | p = 0.173 (ns) | p = 0.042 | ||||
| Control 2 h (C2) | F4-NeuroPs 2 h (F4-2) | Control 4 h (C4) | F4-NeuroPs 4 h (F4-4) | Statistics | |
|---|---|---|---|---|---|
| sperm with intact acrosome % | 82 (81–83.50) | 56 (55–58) | 58 (57–59) | 15 (14.50–15.50) | C2 vs. F4-2, p = 0.002 C4 vs. F4-4, p = 0.002 C2 vs. C4, p = 0.001 F4-2 vs. F4-4, p = 0.001 |
| sperm with partially reacted acrosome % | 12 (11–14) | 18 (17–19) | 22 (21–22) | 44 (43–44.50) | C2 vs. F4-2, p = 0.263 C4 vs. F4-4, p = 0.001 C2 vs. C4, p = 0.091 F4-2 vs. F4-4, p = 0.001 |
| sperm with totally reacted acrosome % | 5 (4.50–5.50) | 24 (23–24) | 20 (20–21) | 40 (40–42) | C2 vs. F4-2, p = 0.001 C4 vs. F4-4, p = 0.003 F4-2 vs. F4-4, p = 0.005 |
| Control (2 h) | F4-NeuroPs (2 h) | Control (4 h) | F4-NeuroPs (4 h) | Statistics | |
|---|---|---|---|---|---|
| sperm with diffuse fair signal% | 62 (60.50–63.50) | 10 (10–13) | 23 (22–24) | 10 (8.50–13) | C2 vs. F4-2, p = 0.001 C4 vs. F4-4, p = 0.001 C2 vs. C4, p = 0.001 F4-2 vs.F4-4, p = 0.981 |
| sperm with absence of signal% | 23 (20.25–25.75) | 10 (10–13) | 12 (10.50–13.50) | 14 (10.75–15) | C2 vs. F4-2, p = 0.001 C4 vs. F4-4, p = 0.865 C2 vs. C4, p = 0.001 F4-2 vs.F4-4, p = 0.527 |
| sperm with mitochondrial signal % | 6 (4.25–9.25) | 70 (68–73.50) | 38 (36.50–39.50) | 25 (25–25.75) | C2 vs. F4-2, p = 0.001 C4 vs. F4-4, p = 0.001 C2 vs. C4, p = 0.001 F4-2 vs.F4-4, p = 0.001 |
| sperm with dotted signal in tail | 8 (8–9.50) | 9 (6.50–11.50) | 27 (26–28) | 53.50 (50.50–56.50) | C2 vs. F4-2, p = 0.999 C4 vs. F4-4, p = 0.001 C2 vs. C4, p = 0.001 F4-2 vs.F4-4, p = 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moretti, E.; Signorini, C.; Noto, D.; Corsaro, R.; Micheli, L.; Durand, T.; Oger, C.; Galano, J.M.; Collodel, G. F4-Neuroprostane Effects on Human Sperm. Int. J. Mol. Sci. 2023, 24, 935. https://doi.org/10.3390/ijms24020935
Moretti E, Signorini C, Noto D, Corsaro R, Micheli L, Durand T, Oger C, Galano JM, Collodel G. F4-Neuroprostane Effects on Human Sperm. International Journal of Molecular Sciences. 2023; 24(2):935. https://doi.org/10.3390/ijms24020935
Chicago/Turabian StyleMoretti, Elena, Cinzia Signorini, Daria Noto, Roberta Corsaro, Lucia Micheli, Thierry Durand, Camille Oger, Jean Marie Galano, and Giulia Collodel. 2023. "F4-Neuroprostane Effects on Human Sperm" International Journal of Molecular Sciences 24, no. 2: 935. https://doi.org/10.3390/ijms24020935