
Silencing DTX3L Inhibits the Progression of Cervical Carcinoma by Regulating PI3K/AKT/mTOR Signaling Pathway
 ,
,
Abstract
:1. Introduction
2. Results
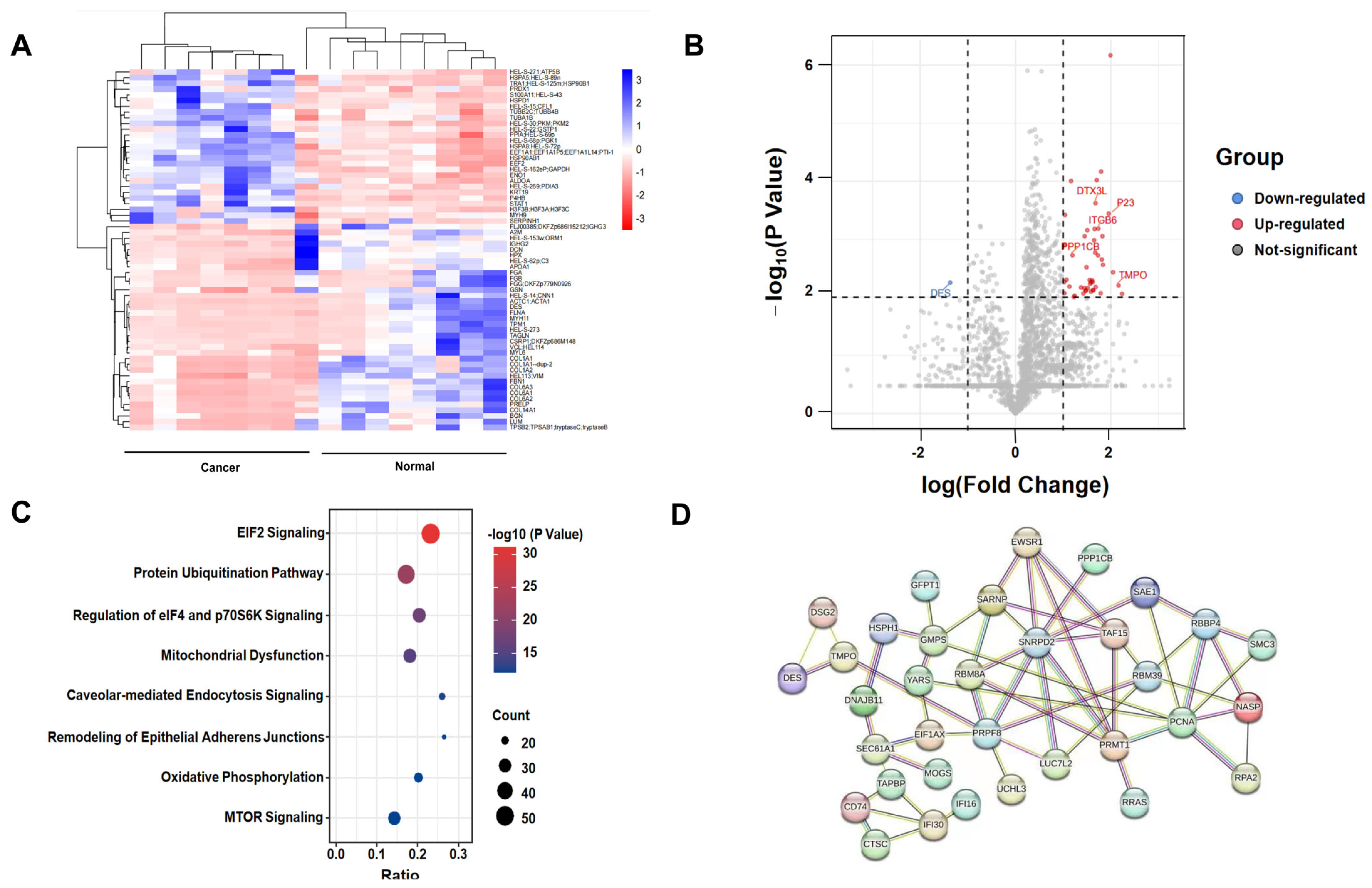
2.1. Protein Identification by Label-Free Quantitative Proteomics
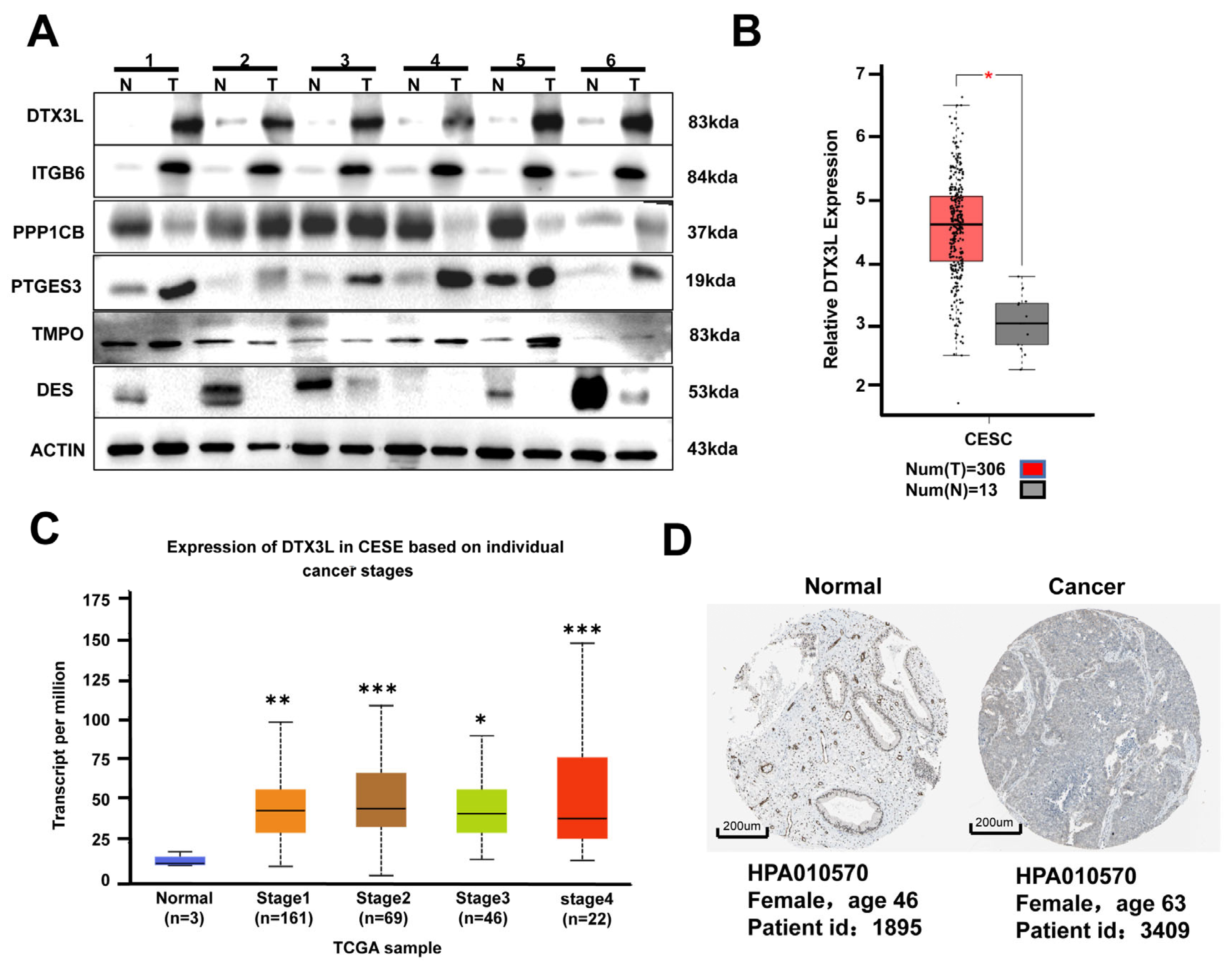
2.2. DTX3L Is Overexpressed in CC Tissues
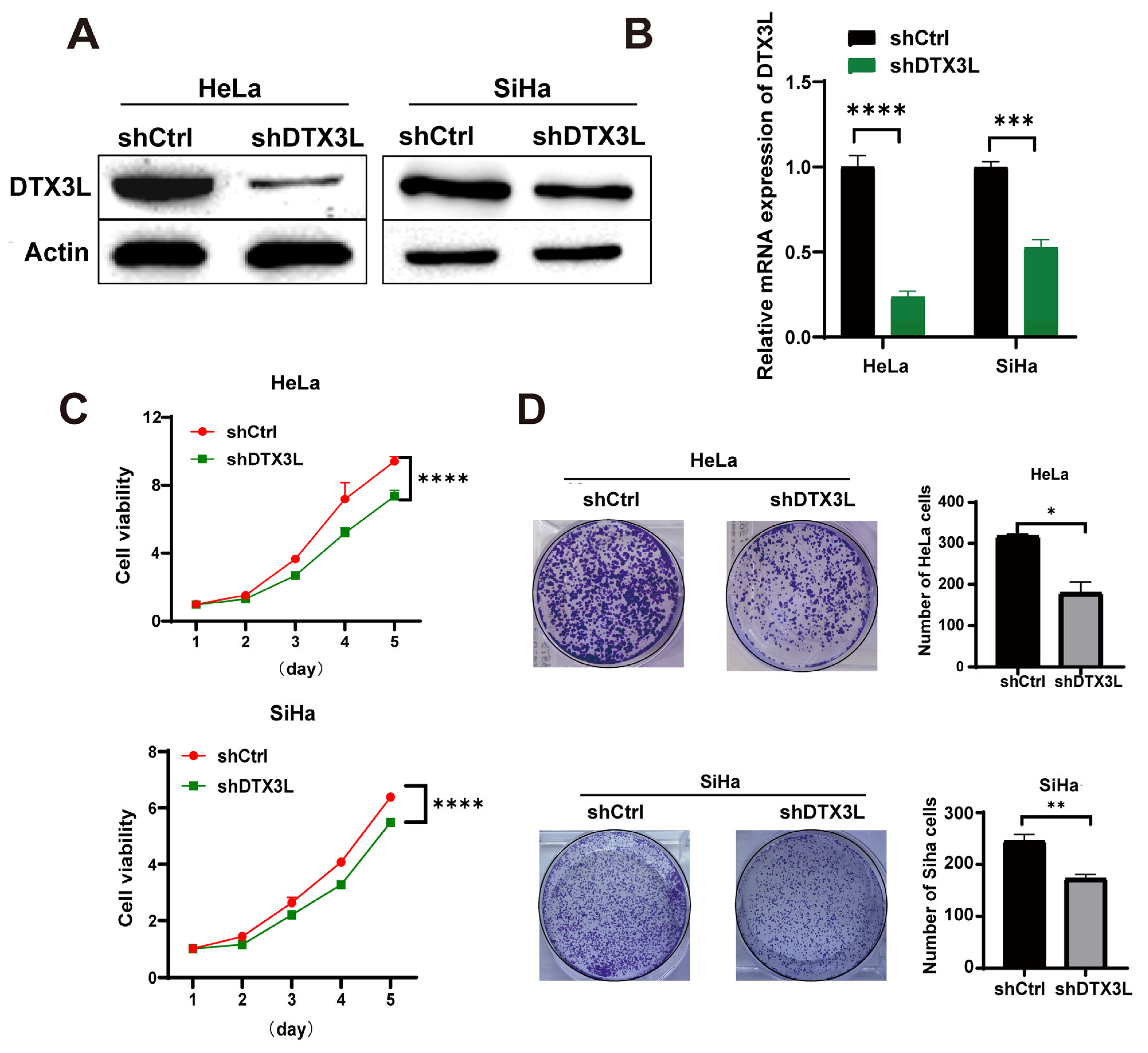
2.3. Knockdown of DTX3L Suppresses Proliferation of CC Cells
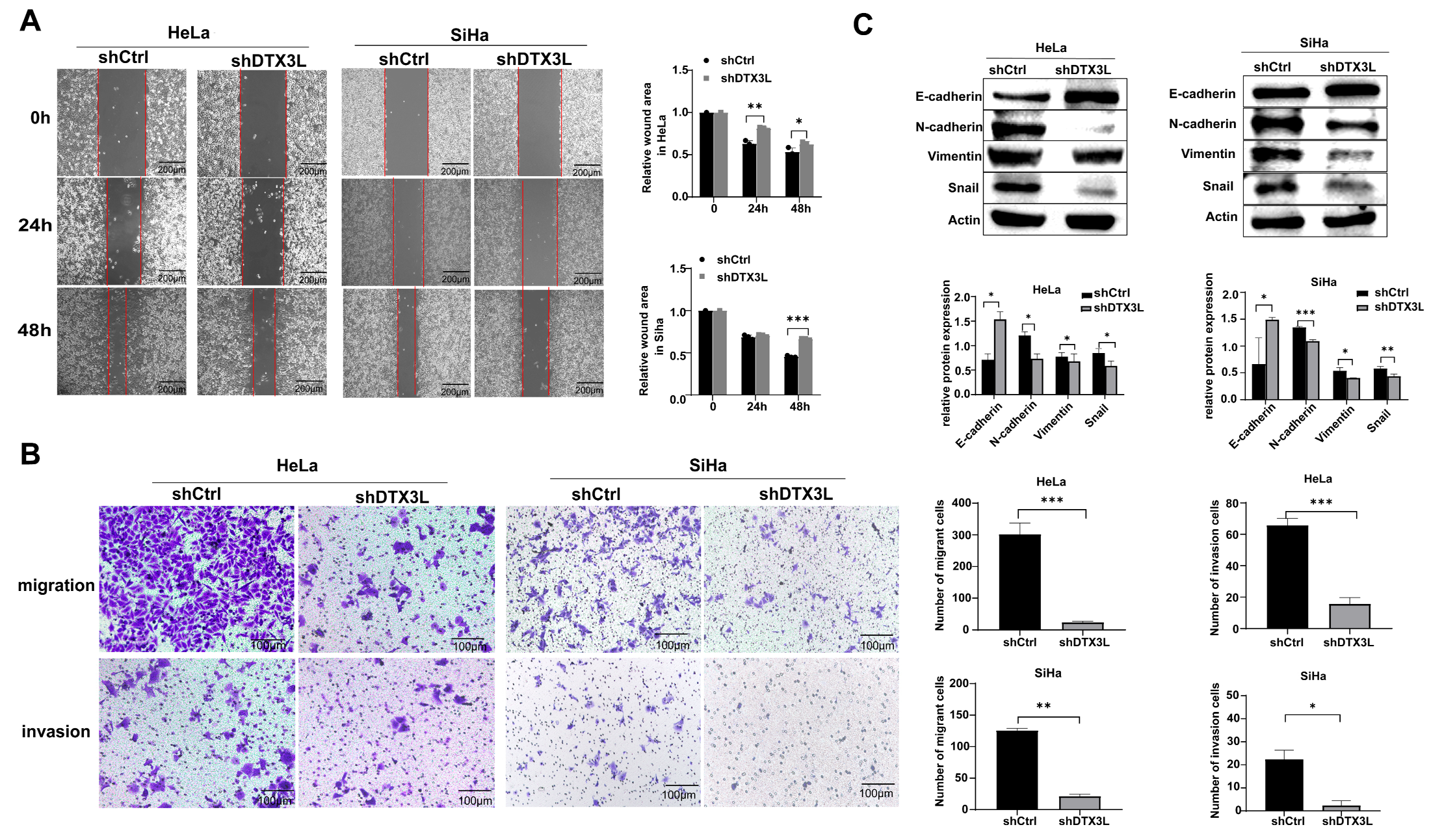
2.4. Knockdown of DTX3L Suppresses Migration and Invasion of CC Cells
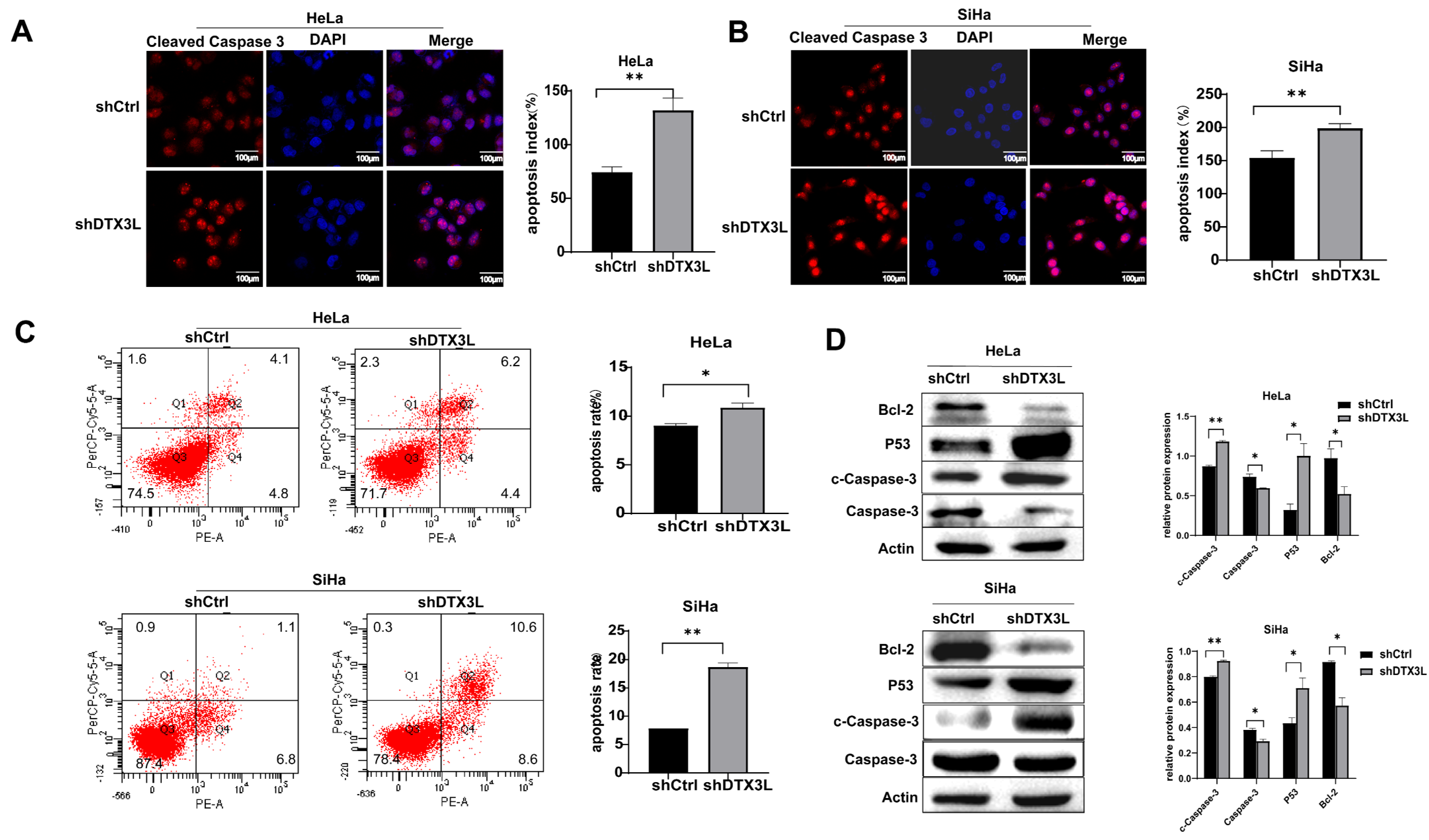
2.5. Knockdown of DTX3L Promotes Apoptosis in CC Cells
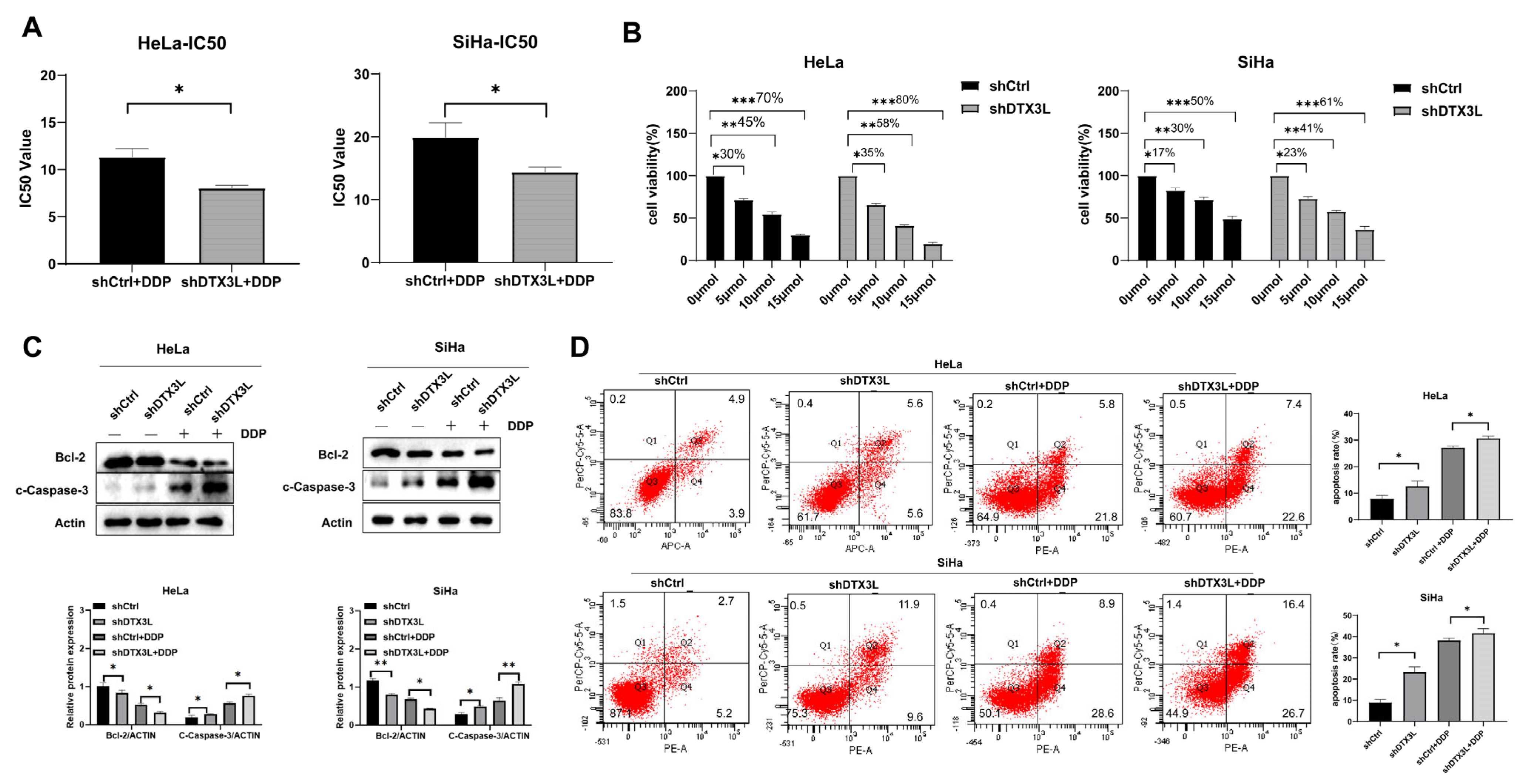
2.6. Combination of Silencing DTX3L and Cisplatin Treatment Enhances Apoptosis of CC Cells
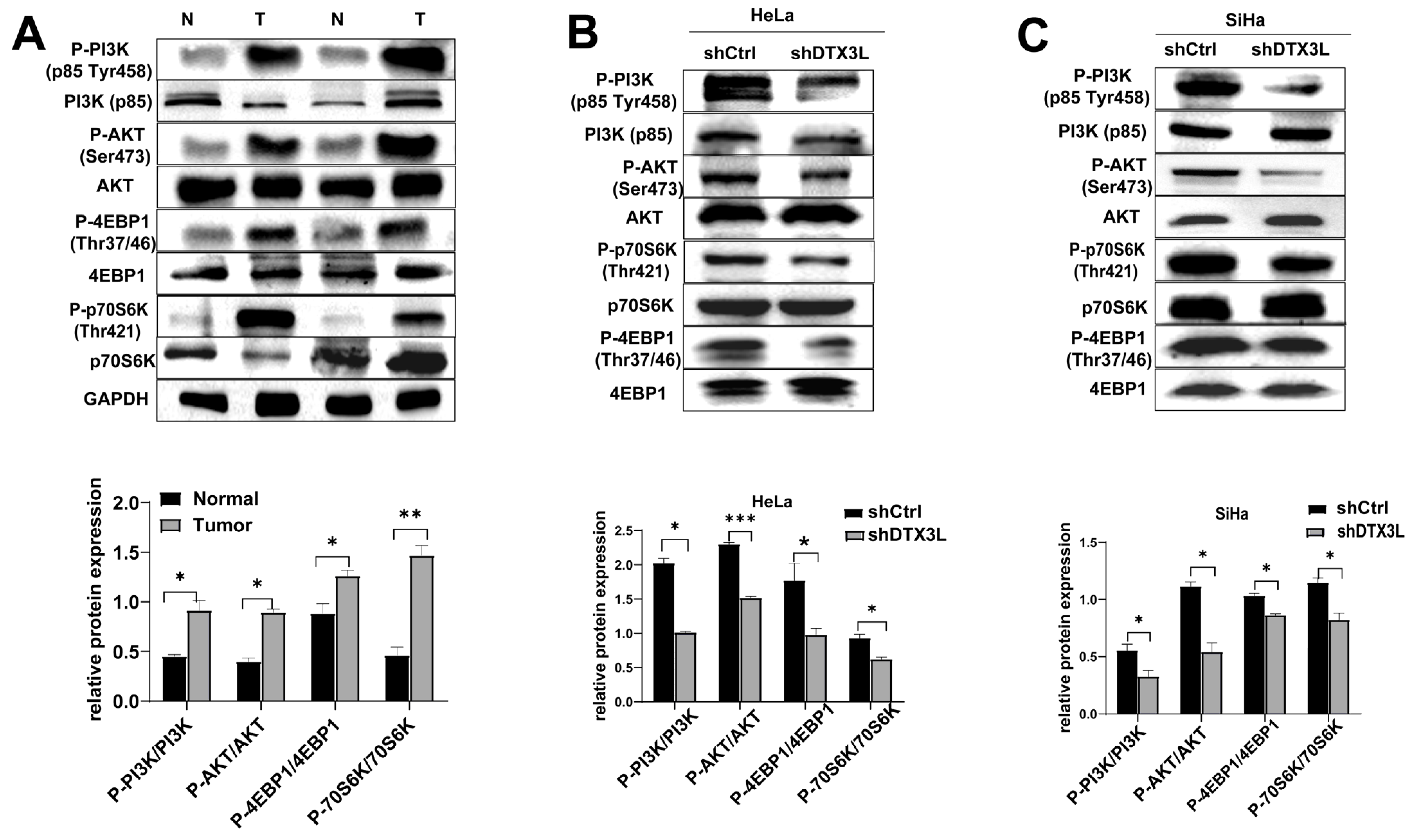
2.7. Knockdown of DTX3L Inhibits PI3K/AKT/mTOR Pathway in CC Cells
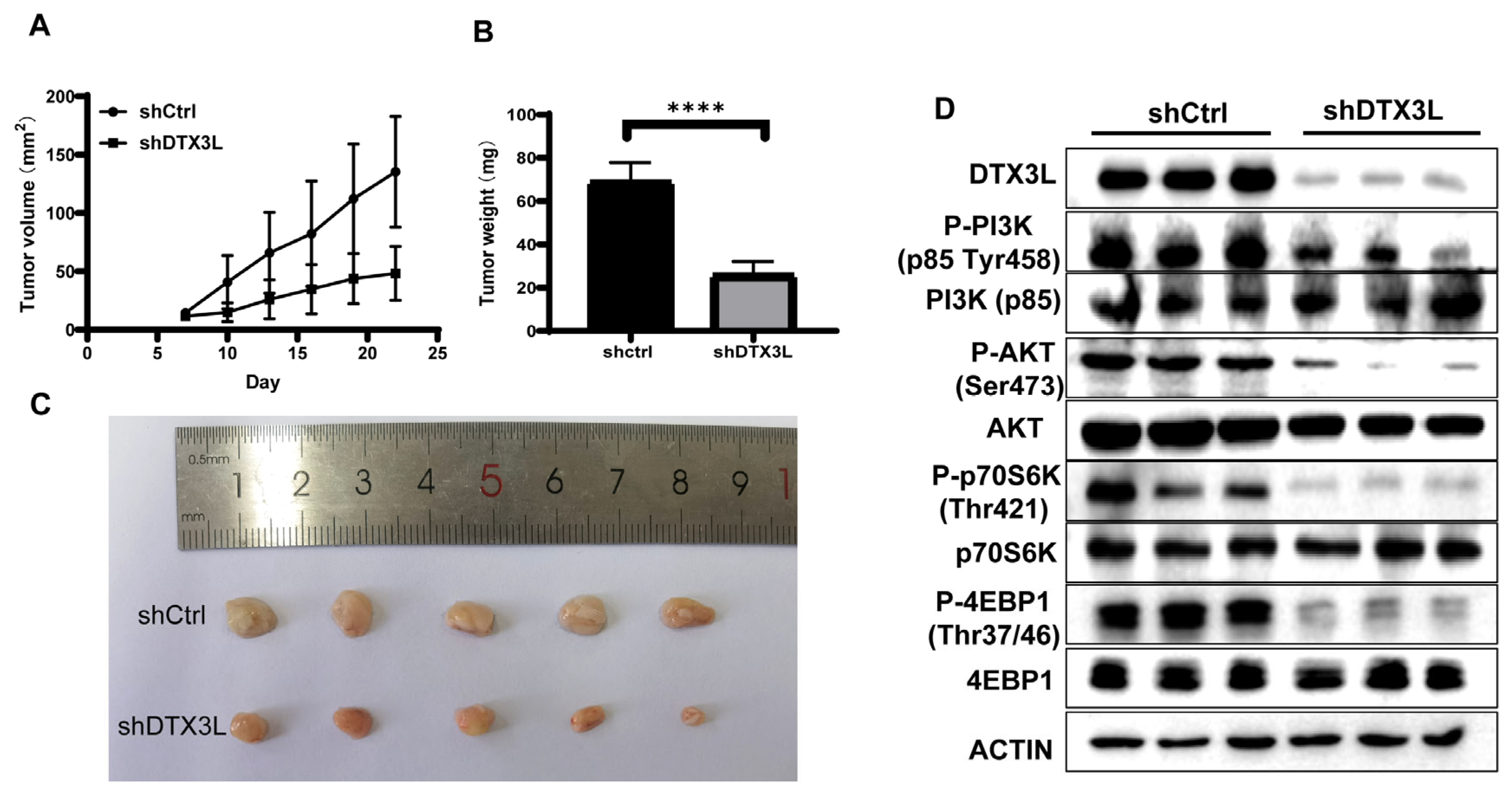
2.8. Silencing DTX3L Inhibits Tumorigenesis In Vivo
3. Discussion
4. Materials and Methods
4.1. Patient Specimens
4.2. LC-MS/MS Analysis
4.3. Proteomic Data Analysis
4.4. Protein–Protein Interaction (PPI) Network Construction
4.5. Real-Time Quantitative PCR (RT-PCR)
4.6. Western Blot
4.7. Cell Culture and Transfection
4.8. CCK-8 Assays
4.9. Wound-Healing Assay
4.10. Colony-Formation Assay
4.11. Transwell Assay
4.12. Flow Cytometry Was Used to Analyze Apoptosis
4.13. Immunofluorescence
4.14. Subcutaneous Xenograft Experiments
4.15. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martínez-Rodríguez, F.; Limones-González, J.E.; Mendoza-Almanza, B.; Esparza-Ibarra, E.L.; Gallegos-Flores, P.I.; Ayala-Luján, J.L.; Godina-González, S.; Salinas, E.; Mendoza-Almanza, G. Understanding Cervical Cancer through Proteomics. Cells 2021, 10, 1854. [Google Scholar] [CrossRef] [PubMed]
- Castle, P.E.; Einstein, M.H.; Sahasrabuddhe, V.V. Cervical cancer prevention and control in women living with human immunodeficiency virus. CA Cancer J. Clin. 2021, 71, 505–526. [Google Scholar] [CrossRef] [PubMed]
- Ramogola-Masire, D.; Luckett, R.; Dreyer, G. Progress and challenges in human papillomavirus and cervical cancer in southern Africa. Curr. Opin. Infect. Dis. 2022, 35, 49–54. [Google Scholar] [CrossRef]
- Wang, R.; Pan, W.; Jin, L.; Huang, W.; Li, Y.; Wu, D.; Gao, C.; Ma, D.; Liao, S. Human papillomavirus vaccine against cervical cancer: Opportunity and challenge. Cancer Lett. 2020, 471, 88–102. [Google Scholar] [CrossRef]
- Gadducci, A.; Cosio, S. Neoadjuvant Chemotherapy in Locally Advanced Cervical Cancer: Review of the Literature and Perspectives of Clinical Research. Anticancer Res. 2020, 40, 4819–4828. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ren, B.; Shi, Y.; Gao, H.; Wang, J.; Xin, Y.; Huang, B.; Liao, S.; Yang, Y.; Xu, Z.; et al. Notch1 inhibition enhances DNA damage induced by cisplatin in cervical cancer. Exp. Cell Res. 2019, 376, 27–38. [Google Scholar] [CrossRef]
- Liu, J.; Yang, C.; Zhang, H.; Hu, W.; Bergquist, J.; Wang, H.; Deng, T.; Yang, X.; Zhang, C.; Zhu, Y.; et al. Quantitative proteomics approach reveals novel biomarkers and pathological mechanism of keloid. Proteom. Clin Appl. 2022, 16, e2100127. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, D.; Li, M.; Wei, X.; Liu, S.; Zhao, M.; Liu, C.; Wang, X.; Jiang, X.; Li, X.; et al. Quantitative Proteomics of TRAMP Mice Combined with Bioinformatics Analysis Reveals That PDGF-B Regulatory Network Plays a Key Role in Prostate Cancer Progression. J. Proteom. Res. 2018, 17, 2401–2411. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, Y.; Men, T.; Jiang, X.; Yang, C.; Li, H.; Wei, X.; Yan, D.; Feng, G.; Yang, J.; et al. Quantitative proteomic analysis of gastric cancer tissue reveals novel proteins in platelet-derived growth factor b signaling pathway. Oncotarget 2017, 8, 22059–22075. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Ye, M.; Chen, X.; Zhao, H.; Hasim, A.; Guo, X. Discovery and validation of FBLN1 and ANT3 as potential biomarkers for early detection of cervical cancer. Cancer Cell Int. 2021, 21, 125. [Google Scholar] [CrossRef]
- Xia, C.; Yang, F.; He, Z.; Cai, Y. iTRAQ-based quantitative proteomic analysis of the inhibition of cervical cancer cell invasion and migration by metformin. Biomed. Pharmacother. 2020, 123, 109762. [Google Scholar] [CrossRef] [PubMed]
- Güzel, C.; Govorukhina, N.I.; Wisman, G.B.A.; Stingl, C.; Dekker, L.J.; Klip, H.G.; Hollema, H.; Guryev, V.; Horvatovich, P.L.; van der Zee, A.G.; et al. Proteomic alterations in early stage cervical cancer. Oncotarget 2018, 9, 18128–18147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Sun, X.; He, J.; Liu, Z. Functions and Molecular Mechanisms of Deltex Family Ubiquitin E3 Ligases in Development and Disease. Front. Cell Dev. Biol. 2021, 9, 706997. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.M.; Rajski, M.; Sivasankaran, B.; Moncayo, G.; Hemmings, B.A.; Merlo, A. Deltex-1 activates mitotic signaling and proliferation and increases the clonogenic and invasive potential of U373 and LN18 glioblastoma cells and correlates with patient survival. PLoS ONE 2013, 8, e57793. [Google Scholar] [CrossRef] [Green Version]
- Hakmé, A.; Huber, A.; Dollé, P.; Schreiber, V. The macroPARP genes Parp-9 and Parp-14 are developmentally and differentially regulated in mouse tissues. Dev. Dyn. 2008, 237, 209–215. [Google Scholar] [CrossRef]
- Holleman, J.; Marchese, A. The ubiquitin ligase deltex-3l regulates endosomal sorting of the G protein-coupled receptor CXCR4. Mol. Biol. Cell. 2014, 25, 1892–1904. [Google Scholar] [CrossRef]
- Liang, H.; Huang, C. Identification of tumor microenvironment-related genes in lower-grade gliomas by mining TCGA database. Transl. Cancer Res. 2020, 9, 4583–4595. [Google Scholar] [CrossRef]
- Shen, Y.; Sun, Y.; Zhang, L.; Liu, H. Effects of DTX3L on the cell proliferation, adhesion, and drug resistance of multiple myeloma cells. Tumour Biol. 2017, 39, 1010428317703941. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, S.B.; Frommel, S.C.; Camicia, R.; Winkler, H.C.; Santoro, R.; Hassa, P.O. DTX3L and ARTD9 inhibit IRF1 expression and mediate in cooperation with ARTD8 survival and proliferation of metastatic prostate cancer cells. Mol. Cancer 2014, 13, 125. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Xu, R.; Zhu, L.; Cheng, X.; Wang, Z.; Manis, J.; Shipp, M.A. BAL1 and its partner E3 ligase, BBAP, link Poly(ADP-ribose) activation, ubiquitylation, and double-strand DNA repair independent of ATM, MDC1, and RNF8. Mol. Cell. Biol. 2013, 33, 845–857. [Google Scholar] [CrossRef]
- Zheng, X.; Zhu, Y.; Wang, X.; Hou, Y.; Fang, Y. Silencing of ITGB6 inhibits the progression of cervical carcinoma via regulating JAK/STAT3 signaling pathway. Ann. Transl. Med. 2021, 9, 803. [Google Scholar] [CrossRef] [PubMed]
- Micalizzi, D.S.; Farabaugh, S.M.; Ford, H.L. Epithelial-mesenchymal transition in cancer: Parallels between normal development and tumor progression. J. Mammary Gland Biol. Neoplasia 2010, 15, 117–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Rajczewski, A.T.; Jagtap, P.D.; Griffin, T.J. An overview of technologies for MS-based proteomics-centric multi-omics. Expert Rev. Proteom. 2022, 19, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Kim, H.S.; Bojmar, L.; Gyan, K.E.; Cioffi, M.; Hernandez, J.; Zambirinis, C.P.; Rodrigues, G.; Molina, H.; Heissel, S.; et al. Extracellular Vesicle and Particle Biomarkers Define Multiple Human Cancers. Cell 2020, 182, 1044–1061.e18. [Google Scholar] [CrossRef]
- Harris, C.A.; Andryuk, P.J.; Cline, S.W.; Mathew, S.; Siekierka, J.J.; Goldstein, G. Structure and mapping of the human thymopoietin (TMPO) gene and relationship of human TMPO beta to rat lamin-associated polypeptide 2. Genomics 1995, 28, 198–205. [Google Scholar] [CrossRef]
- Sun, X.; Xin, S.; Li, W.; Zhang, Y.; Ye, L. Discovery of Notch Pathway-Related Genes for Predicting Prognosis and Tumor Microenvironment Status in Bladder Cancer. Front. Genet. 2022, 13, 928778. [Google Scholar] [CrossRef]
- Takeyama, K.; Aguiar, R.C.; Gu, L.; He, C.; Freeman, G.J.; Kutok, J.L.; Aster, J.C.; Shipp, M.A. The BAL-binding protein BBAP and related Deltex family members exhibit ubiquitin-protein isopeptide ligase activity. J. Biol. Chem. 2003, 278, 21930–21937. [Google Scholar] [CrossRef] [Green Version]
- Juszczynski, P.; Kutok, J.L.; Li, C.; Mitra, J.; Aguiar, R.C.; Shipp, M.A. BAL1 and BBAP are regulated by a gamma interferon-responsive bidirectional promoter and are overexpressed in diffuse large B-cell lymphomas with a prominent inflammatory infiltrate. Mol. Cell. Biol. 2006, 26, 5348–5359. [Google Scholar] [CrossRef] [Green Version]
- Bolis, M.; Paroni, G.; Fratelli, M.; Vallerga, A.; Guarrera, L.; Zanetti, A.; Kurosaki, M.; Garattini, S.K.; Gianni, M.; Lupi, M.; et al. All-Trans Retinoic Acid Stimulates Viral Mimicry, Interferon Responses and Antigen Presentation in Breast-Cancer Cells. Cancers 2020, 12, 1169. [Google Scholar] [CrossRef]
- Xu, P.; Tao, X.; Zhao, C.; Huang, Q.; Chang, H.; Ban, N.; Bei, Y.; Xia, X.; Shen, C.; Wang, K.; et al. DTX3L is upregulated in glioma and is associated with glioma progression. Int. J. Mol. Med. 2017, 40, 491–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alausa, A.; Victor, U.C.; Fadahunsi, O.S.; Owolabi, N.; Adeniji, A.; Olatinwo, M.; Ogunlana, A.T.; Olaleke, B.; Balogun, T.A.; Ogundepo, S.; et al. Checkpoints and immunity in cancers: Role of GNG12. Pharmacol. Res. 2022, 180, 106242. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, N.; Li, S.; Yu, H.; Wang, Y.; Liu, Z.; Han, L.; Tang, Z. Synergistic Therapy for Cervical Cancer by Codelivery of Cisplatin and JQ1 Inhibiting Plk1-Mutant Trp53 Axis. Nano Lett. 2021, 21, 2412–2421. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: A novel therapeutic strategy. Semin. Cancer Biol. 2022, 80, 1–17. [Google Scholar] [CrossRef]
- Alzahrani, A.S. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Z.; Wang, L. Uncover the Nuclear Proteomic Landscape with Enriched Nuclei Followed by Label-Free Quantitative Mass Spectrometry. Methods Mol. Biol. 2021, 2297, 115–124. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | 5′-3′Sequence |
|---|---|---|
| DTX3L | F | TGAGTCCTTTGGCACCAT |
| R | GGCAAGTATGCAGTTCGC | |
| GAPDH | F | CCCTTCATTGACCTCAACTACATG |
| R | TGGGATTTCCATTGATGACAAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.; Hu, Y.; Pei, Y.; Li, R.; Xu, F.; Chi, X.; Mi, J.; Bergquist, J.; Lu, L.; Zhang, L.; et al. Silencing DTX3L Inhibits the Progression of Cervical Carcinoma by Regulating PI3K/AKT/mTOR Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 861. https://doi.org/10.3390/ijms24010861
Hu W, Hu Y, Pei Y, Li R, Xu F, Chi X, Mi J, Bergquist J, Lu L, Zhang L, et al. Silencing DTX3L Inhibits the Progression of Cervical Carcinoma by Regulating PI3K/AKT/mTOR Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(1):861. https://doi.org/10.3390/ijms24010861
Chicago/Turabian StyleHu, Wei, Yaorui Hu, Yao Pei, Rongrong Li, Fuyi Xu, Xiaodong Chi, Jia Mi, Jonas Bergquist, Lu Lu, Luping Zhang, and et al. 2023. "Silencing DTX3L Inhibits the Progression of Cervical Carcinoma by Regulating PI3K/AKT/mTOR Signaling Pathway" International Journal of Molecular Sciences 24, no. 1: 861. https://doi.org/10.3390/ijms24010861