
Low-Intensity Pulsed Ultrasound Alleviates Human Testicular Leydig Cell Senescence In Vitro
 ,
,
Abstract
:1. Introduction
2. Results
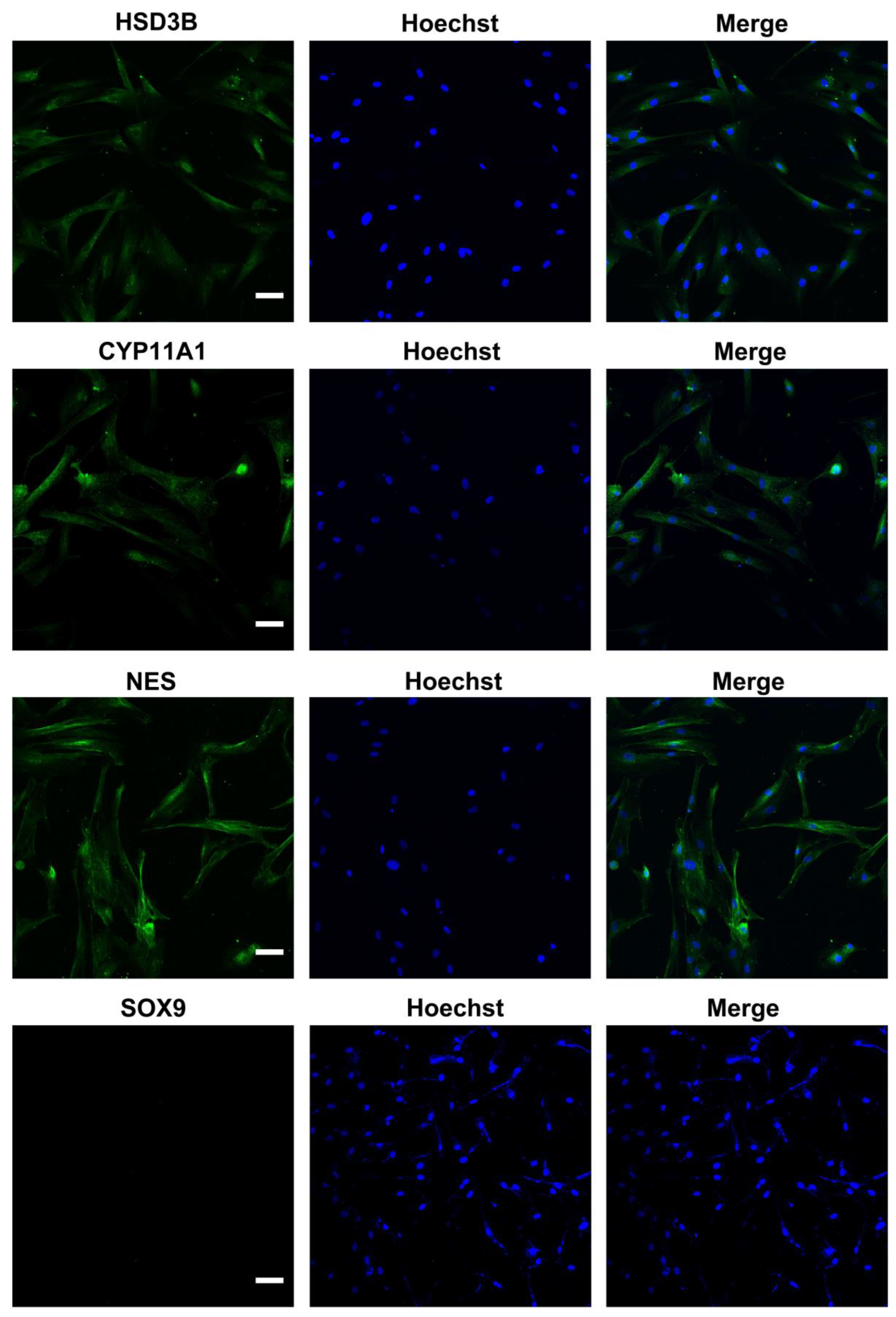
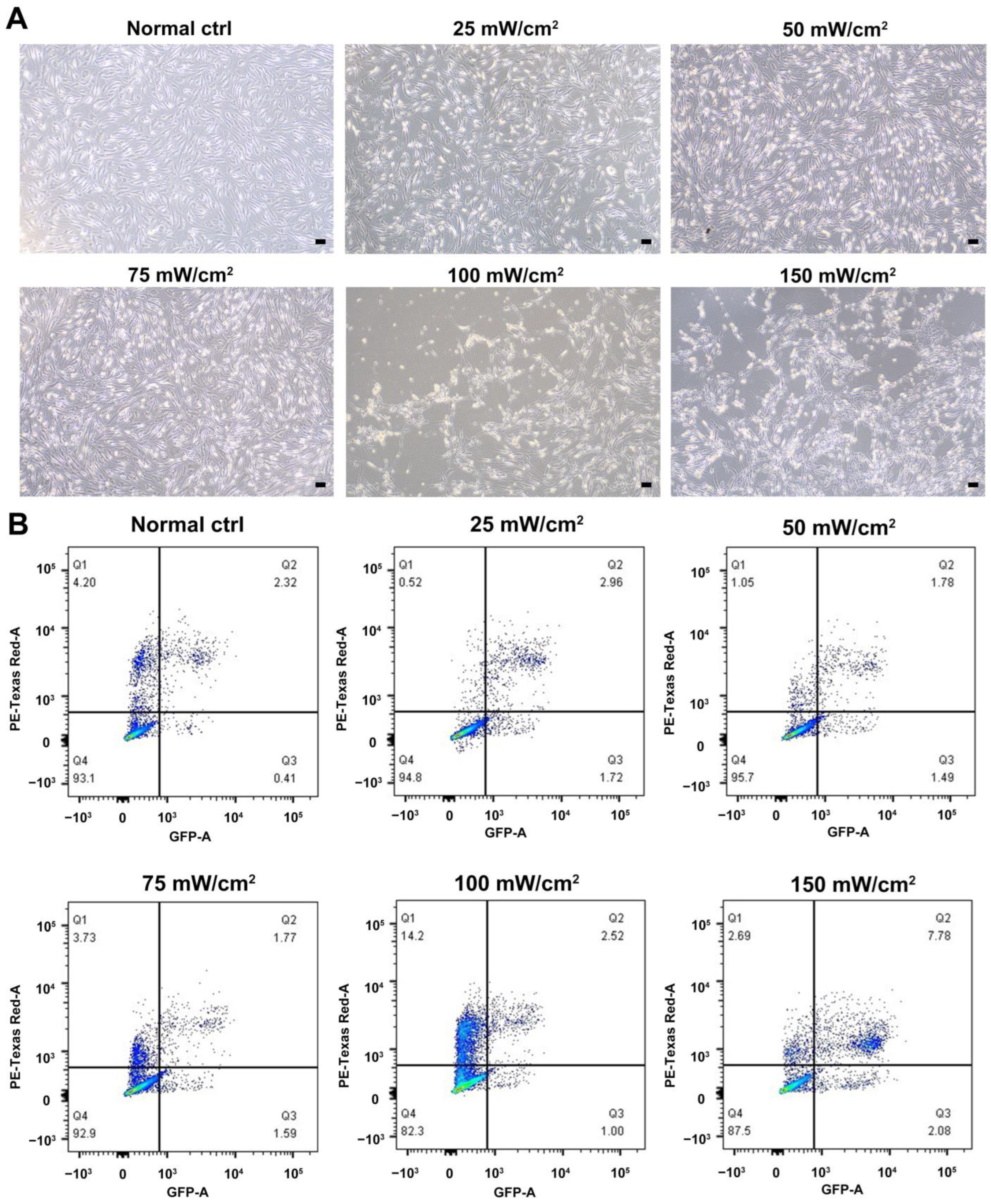
2.1. Identification of LCs and Biosafety Assessment of LIPUS
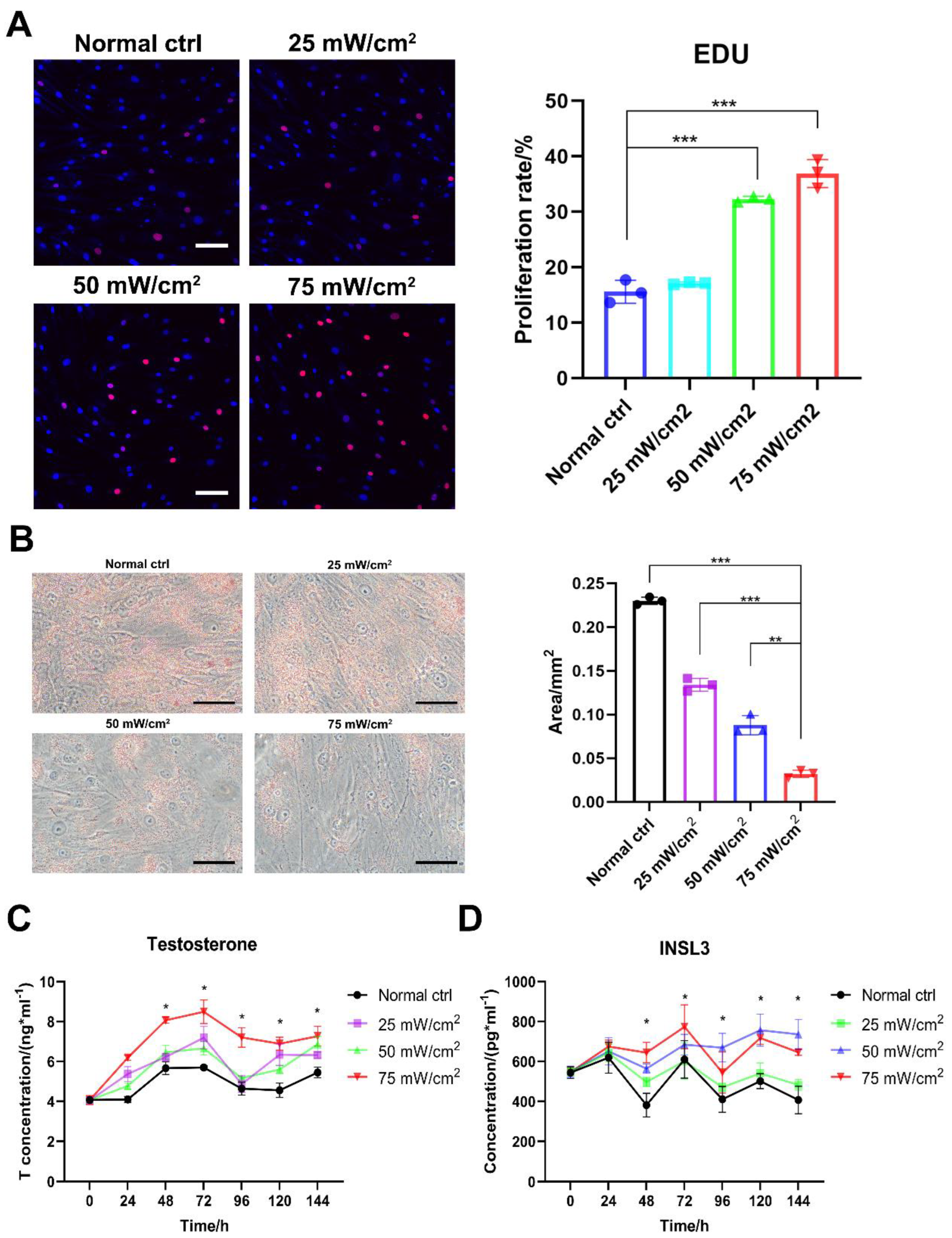
2.2. LIPUS Improves Proliferation and Secretion Function of Aged LCs
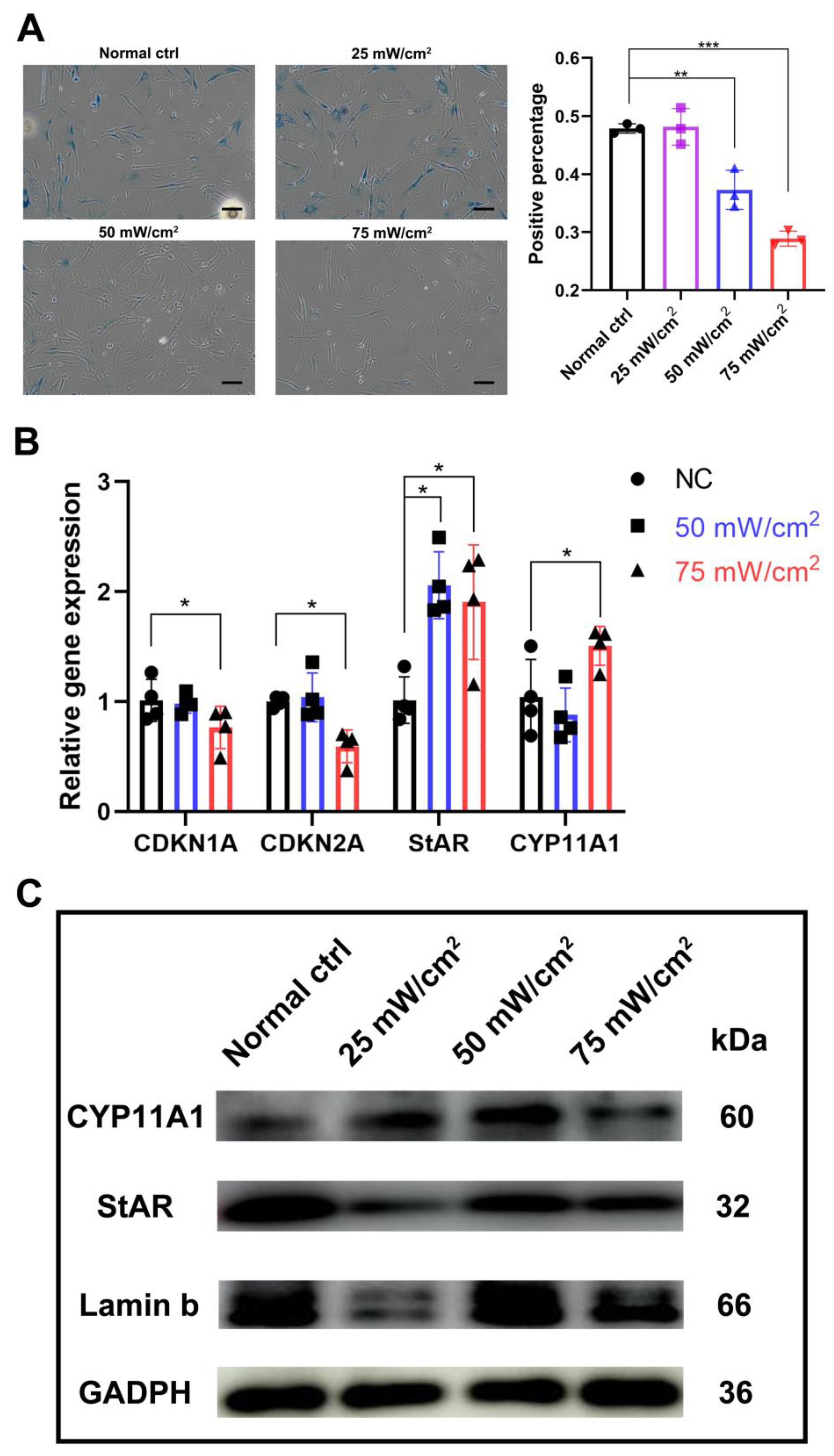
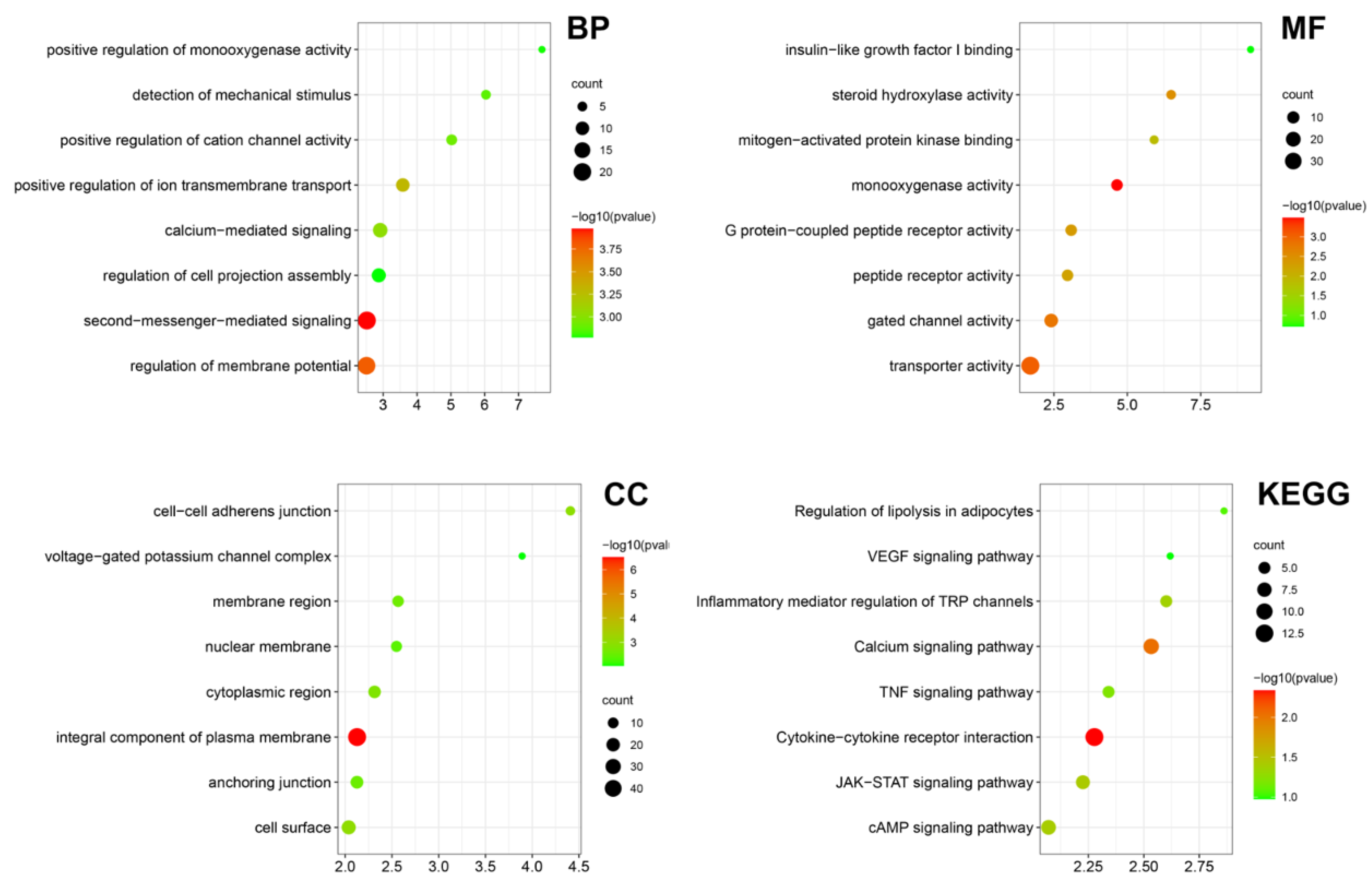
2.3. LIPUS Delayed Cellular Senescence and Testosterone-Related Enzyme Synthesis Pathways
2.4. LIPUS Possess the Effect of Alleviating Leydig Cell Senescence and Testosterone Production Pathways
3. Discussion
4. Materials and Methods
4.1. Donor Testicular Tissue
4.2. Isolation and Immunofluorescence Staining of Human LCs
4.3. LIPUS Treatment
4.4. Annexin-V and PI Staining and Flow Cytometry
4.5. Oil Red O Staining
4.6. β-Gal Staining
4.7. RNA Extraction, RT-PCR, and Real-Time qPCR
4.8. ELISA
4.9. RNA Sequencing
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Camacho, E.M.; Huhtaniemi, I.T.; O’Neill, T.W.; Finn, J.D.; Pye, S.R.; Lee, D.M.; Tajar, A.; Bartfai, G.; Boonen, S.; Casanueva, F.F.; et al. Age-associated changes in hypothalamic-pituitary-testicular function in middle-aged and older men are modified by weight change and lifestyle factors: Longitudinal results from the European Male Ageing Study. Eur. J. Endocrinol. 2013, 168, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Wen, X.; Ji, M.; Guan, X.; Chen, P.; Hao, X.; Chen, F.; Hu, Y.; Duan, P.; Ge, R.S.; et al. Differentiation of seminiferous tubule-associated stem cells into leydig cell and myoid cell lineages. Mol. Cell. Endocrinol. 2021, 525, 111179. [Google Scholar] [CrossRef]
- Mendis-Handagama, S.M. Peroxisomes and intracellular cholesterol trafficking in adult rat Leydig cells following Luteinizing hormone stimulation. Tissue Cell 2000, 32, 102–106. [Google Scholar] [CrossRef]
- Nie, X.; Munyoki, S.K.; Sukhwani, M.; Schmid, N.; Missel, A.; Emery, B.R.; DonorConnect; Stukenborg, J.B.; Mayerhofer, A.; Orwig, K.E.; et al. Single-cell analysis of human testis aging and correlation with elevated body mass index. Dev. Cell 2022, 57, 1160–1176.e1165. [Google Scholar] [CrossRef]
- Mularoni, V.; Esposito, V.; Di Persio, S.; Vicini, E.; Spadetta, G.; Berloco, P.; Fanelli, F.; Mezzullo, M.; Pagotto, U.; Pelusi, C.; et al. Age-related changes in human Leydig cell status. Hum. Reprod. 2020, 35, 2663–2676. [Google Scholar] [CrossRef]
- Xin, Z.; Lin, G.; Lei, H.; Lue, T.F.; Guo, Y. Clinical applications of low-intensity pulsed ultrasound and its potential role in urology. Transl. Androl. Urol. 2016, 5, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Monma, Y.; Shindo, T.; Eguchi, K.; Kurosawa, R.; Kagaya, Y.; Ikumi, Y.; Ichijo, S.; Nakata, T.; Miyata, S.; Matsumoto, A.; et al. Low-intensity pulsed ultrasound ameliorates cardiac diastolic dysfunction in mice: A possible novel therapy for heart failure with preserved left ventricular ejection fraction. Cardiovasc. Res. 2021, 117, 1325–1338. [Google Scholar] [CrossRef]
- Haddad, S.; Anselmo-Franci, J.A.; Petenusci, S.O.; Lamano-Carvalho, T.L. Effect of low-intensity pulsed ultrasound on prepubertal rat testis. Braz. J. Med. Biol. Res. 1991, 24, 697–700. [Google Scholar]
- Haddad, S.; Petenusci, S.O.; Franci, J.A.; Carvalho, T.L. Stimulation of prepubertal, pubertal and adult rat testis with low-intensity pulsed ultrasound. Arch. Int. Physiol. Biochim. Biophys. 1994, 102, 13–16. [Google Scholar] [CrossRef]
- Li, X.; Zhu, Q.; Wen, Z.; Yuan, K.; Su, Z.; Wang, Y.; Zhong, Y.; Ge, R.S. Androgen and Luteinizing Hormone Stimulate the Function of Rat Immature Leydig Cells Through Different Transcription Signals. Front. Endocrinol. 2021, 12, 599149. [Google Scholar] [CrossRef]
- Zirkin, B.R.; Papadopoulos, V. Leydig cells: Formation, function, and regulation. Biol. Reprod. 2018, 99, 101–111. [Google Scholar] [CrossRef]
- Li, X.; Wang, Z.; Jiang, Z.; Guo, J.; Zhang, Y.; Li, C.; Chung, J.; Folmer, J.; Liu, J.; Lian, Q.; et al. Regulation of seminiferous tubule-associated stem Leydig cells in adult rat testes. Proc. Natl. Acad. Sci. USA 2016, 113, 2666–2671. [Google Scholar] [CrossRef] [Green Version]
- Gaur, M.; Ramathal, C.; Reijo Pera, R.A.; Turek, P.J.; John, C.M. Isolation of human testicular cells and co-culture with embryonic stem cells. Reproduction 2018, 155, 153–166. [Google Scholar] [CrossRef]
- Tabuchi, Y.; Sugahara, Y.; Ikegame, M.; Suzuki, N.; Kitamura, K.; Kondo, T. Genes responsive to low-intensity pulsed ultrasound in MC3T3-E1 preosteoblast cells. Int. J. Mol. Sci. 2013, 14, 22721–22740. [Google Scholar] [CrossRef] [Green Version]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef]
- Surampudi, P.N.; Wang, C.; Swerdloff, R. Hypogonadism in the aging male diagnosis, potential benefits, and risks of testosterone replacement therapy. Int. J. Endocrinol. 2012, 2012, 625434. [Google Scholar] [CrossRef] [Green Version]
- Huhtaniemi, I. Late-onset hypogonadism: Current concepts and controversies of pathogenesis, diagnosis and treatment. Asian J. Androl. 2014, 16, 192–202. [Google Scholar] [CrossRef]
- Gentile, G.; Rizzo, M.; Bianchi, L.; Falcone, M.; Dente, D.; Ciletti, M.; Franceschelli, A.; Vagnoni, V.; Garofalo, M.; Schiavina, R.; et al. Testis Sparing Surgery of Small Testicular Masses: Retrospective Analysis of a Multicenter Cohort. J. Urol. 2020, 203, 760–766. [Google Scholar] [CrossRef]
- Gagliano-Juca, T.; Basaria, S. Testosterone replacement therapy and cardiovascular risk. Nat. Rev. Cardiol. 2019, 16, 555–574. [Google Scholar] [CrossRef]
- Santella, C.; Renoux, C.; Yin, H.; Yu, O.H.Y.; Azoulay, L. Testosterone Replacement Therapy and the Risk of Prostate Cancer in Men with Late-Onset Hypogonadism. Am. J. Epidemiol. 2019, 188, 1666–1673. [Google Scholar] [CrossRef]
- Aghazadeh, Y.; Zirkin, B.R.; Papadopoulos, V. Pharmacological regulation of the cholesterol transport machinery in steroidogenic cells of the testis. Vitam. Horm. 2015, 98, 189–227. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Chen, H.; Zirkin, B.R. Leydig cell aging: Steroidogenic acute regulatory protein (StAR) and cholesterol side-chain cleavage enzyme. J. Androl. 2001, 22, 149–156. [Google Scholar] [PubMed]
- Luo, L.; Chen, H.; Zirkin, B.R. Temporal relationships among testosterone production, steroidogenic acute regulatory protein (StAR), and P450 side-chain cleavage enzyme (P450scc) during Leydig cell aging. J. Androl. 2005, 26, 25–31. [Google Scholar] [PubMed]
- Ivell, R.; Anand-Ivell, R. Biology of insulin-like factor 3 in human reproduction. Hum. Reprod. Update. 2009, 15, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Anand-Ivell, R.; Heng, K.; Severn, K.; Antonio, L.; Bartfai, G.; Casanueva, F.F.; Huhtaniemi, I.T.; Giwercman, A.; Maggi, M.; O’Neill, T.W.; et al. Association of age, hormonal, and lifestyle factors with the Leydig cell biomarker INSL3 in aging men from the European Male Aging Study cohort. Andrology 2022, 10, 1328–1338. [Google Scholar] [CrossRef]
- Wilson, W.H.; O’Connor, O.A.; Czuczman, M.S.; LaCasce, A.S.; Gerecitano, J.F.; Leonard, J.P.; Tulpule, A.; Dunleavy, K.; Xiong, H.; Chiu, Y.L.; et al. Navitoclax, a targeted high-affinity inhibitor of BCL-2, in lymphoid malignancies: A phase 1 dose-escalation study of safety, pharmacokinetics, pharmacodynamics, and antitumour activity. Lancet Oncol. 2010, 11, 1149–1159. [Google Scholar] [CrossRef] [Green Version]
- Vizioli, M.G.; Liu, T.; Miller, K.N.; Robertson, N.A.; Gilroy, K.; Lagnado, A.B.; Perez-Garcia, A.; Kiourtis, C.; Dasgupta, N.; Lei, X.; et al. Mitochondria-to-nucleus retrograde signaling drives formation of cytoplasmic chromatin and inflammation in senescence. Genes Dev. 2020, 34, 428–445. [Google Scholar] [CrossRef]
- Farr, J.N.; Xu, M.; Weivoda, M.M.; Monroe, D.G.; Fraser, D.G.; Onken, J.L.; Negley, B.A.; Sfeir, J.G.; Ogrodnik, M.B.; Hachfeld, C.M.; et al. Targeting cellular senescence prevents age-related bone loss in mice. Nat. Med. 2017, 23, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, R.; Stahl, P.J.; Schlegel, P.N. Medical therapy for spermatogenic failure. Asian J. Androl. 2012, 14, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Bosland, M.C. Testosterone treatment is a potent tumor promoter for the rat prostate. Endocrinology 2014, 155, 4629–4633. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Freeman, G.; Cowling, B.J.; Schooling, C.M. Testosterone therapy and cardiovascular events among men: A systematic review and meta-analysis of placebo-controlled randomized trials. BMC Med. 2013, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Claps, F.; Pavan, N.; Umari, P.; Rizzo, M.; Barbone, F.; Giangreco, M.; Liguori, G.; Mir, C.M.; Bussani, R.; Trombetta, C. Incidence, predictive factors and survival outcomes of incidental prostate cancer in patients who underwent radical cystectomy for bladder cancer. Minerva Urol. Nephrol. 2021, 73, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xie, Y.; Chen, H.; Lv, L.; Yao, J.; Zhang, M.; Xia, K.; Feng, X.; Li, Y.; Liang, X.; et al. FOXO4-DRI alleviates age-related testosterone secretion insufficiency by targeting senescent Leydig cells in aged mice. Aging 2020, 12, 1272–1284. [Google Scholar] [CrossRef] [PubMed]
- Peak, T.C.; Haney, N.M.; Wang, W.; DeLay, K.J.; Hellstrom, W.J. Stem cell therapy for the treatment of Leydig cell dysfunction in primary hypogonadism. World J. Stem Cells 2016, 8, 306–315. [Google Scholar] [CrossRef]
- Li, L.; Papadopoulos, V. Advances in stem cell research for the treatment of primary hypogonadism. Nat. Rev. Urol. 2021, 18, 487–507. [Google Scholar] [CrossRef]
- Xia, S.J.; Chen, H.R.; Li, Z.; Li, X.C.; Zhi, E.L.; Tian, R.H.; Li, P.; Huang, Y.H.; Hu, J.L.; Qiu, Y. Observation on the effectiveness and safety of different frequencies of low-intensity pulsed ultrasonic mechanical force in the treatment of erectile dysfunction. Zhonghua Yi Xue Za Zhi 2020, 100, 1432–1436. [Google Scholar] [CrossRef]
- Lu, Z.; Lin, G.; Reed-Maldonado, A.; Wang, C.; Lee, Y.C.; Lue, T.F. Low-intensity Extracorporeal Shock Wave Treatment Improves Erectile Function: A Systematic Review and Meta-analysis. Eur. Urol. 2017, 71, 223–233. [Google Scholar] [CrossRef]
- Beattie, M.C.; Adekola, L.; Papadopoulos, V.; Chen, H.; Zirkin, B.R. Leydig cell aging and hypogonadism. Exp. Gerontol. 2015, 68, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Midzak, A.S.; Chen, H.; Papadopoulos, V.; Zirkin, B.R. Leydig cell aging and the mechanisms of reduced testosterone synthesis. Mol. Cell. Endocrinol. 2009, 299, 23–31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Old | Young | |
|---|---|---|
| Age | 64 | 28 |
| Estradiol | 25.00 pg/mg | 27.00 pg/mg |
| LH | 11.17 IU/L | 3.57 IU/L |
| FSH | 12.43 IU/L | 2.10 IU/L |
| Prolactin | 1.76 ng/ml | 7.58 ng/ml |
| Progesterone | <0.08 ug/L | 0.25 ug/L |
| Testosterone | 1.34 ug/L | 5.76 ug/L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.; Luo, J.; Xu, S.; Zhao, L.; Yao, C.; Xu, J.; Tian, R.; Zhi, E.; Huang, Y.; Xia, S.; et al. Low-Intensity Pulsed Ultrasound Alleviates Human Testicular Leydig Cell Senescence In Vitro. Int. J. Mol. Sci. 2023, 24, 418. https://doi.org/10.3390/ijms24010418
Han S, Luo J, Xu S, Zhao L, Yao C, Xu J, Tian R, Zhi E, Huang Y, Xia S, et al. Low-Intensity Pulsed Ultrasound Alleviates Human Testicular Leydig Cell Senescence In Vitro. International Journal of Molecular Sciences. 2023; 24(1):418. https://doi.org/10.3390/ijms24010418
Chicago/Turabian StyleHan, Sha, Jiaqiang Luo, Shuai Xu, Liangyu Zhao, Chencheng Yao, Junwei Xu, Ruhui Tian, Erlei Zhi, Yuhua Huang, Shujie Xia, and et al. 2023. "Low-Intensity Pulsed Ultrasound Alleviates Human Testicular Leydig Cell Senescence In Vitro" International Journal of Molecular Sciences 24, no. 1: 418. https://doi.org/10.3390/ijms24010418