
Ssc-miR-92b-3p Regulates Porcine Trophoblast Cell Proliferation and Migration via the PFKM Gene

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Expression Profile of ssc-miR-92b-3p
2.2. The Role of miR-92b-3p in Porcine Trophoblast Cells
2.3. Ssc-miR-92b-3p Directly Targets the 3’-UTR of PFKM in PTr2
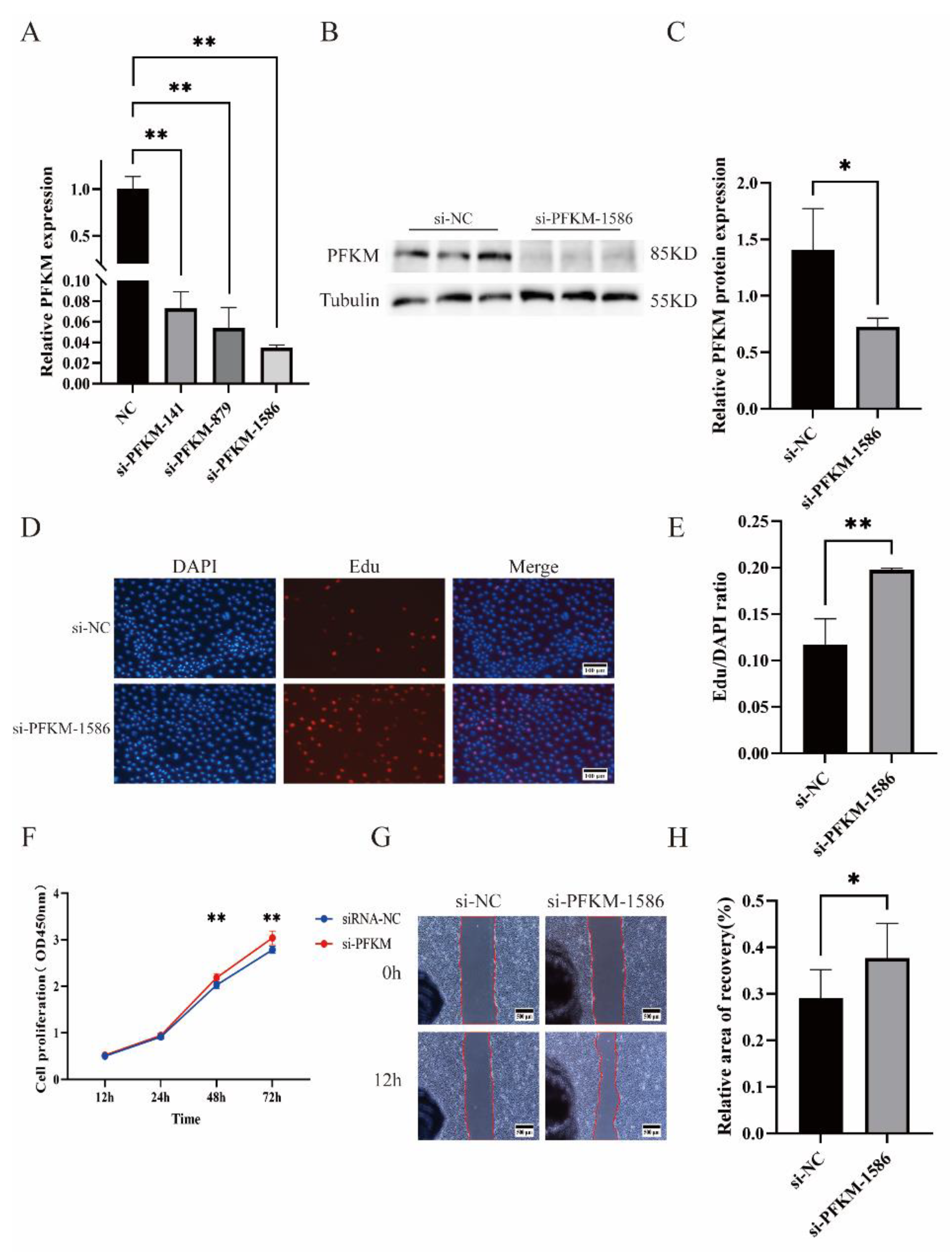
2.4. The Role of PFKM in PTr2
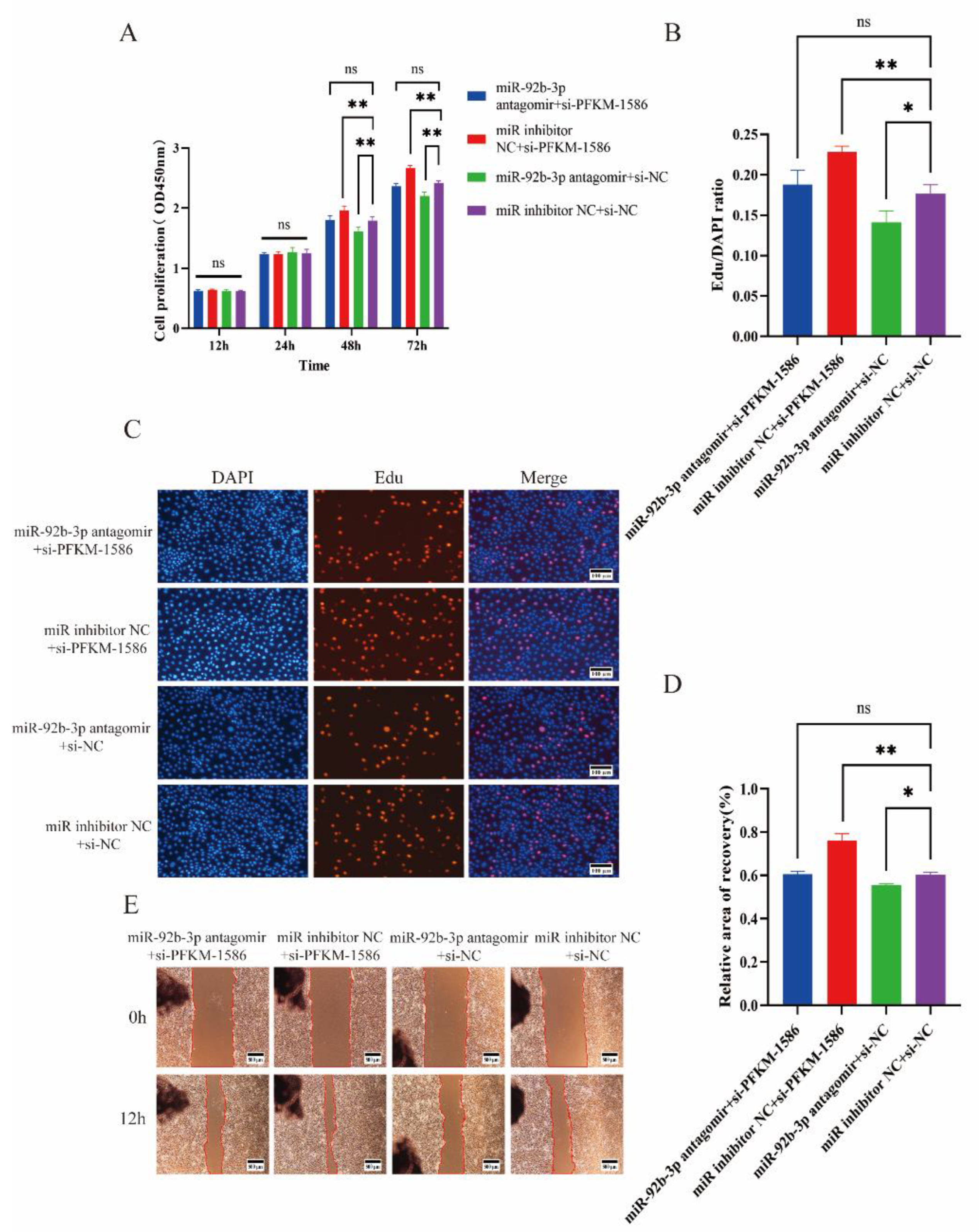
2.5. Si-PFKM Rescued the Influence of miR-92b-3p Antagomir in PTr2
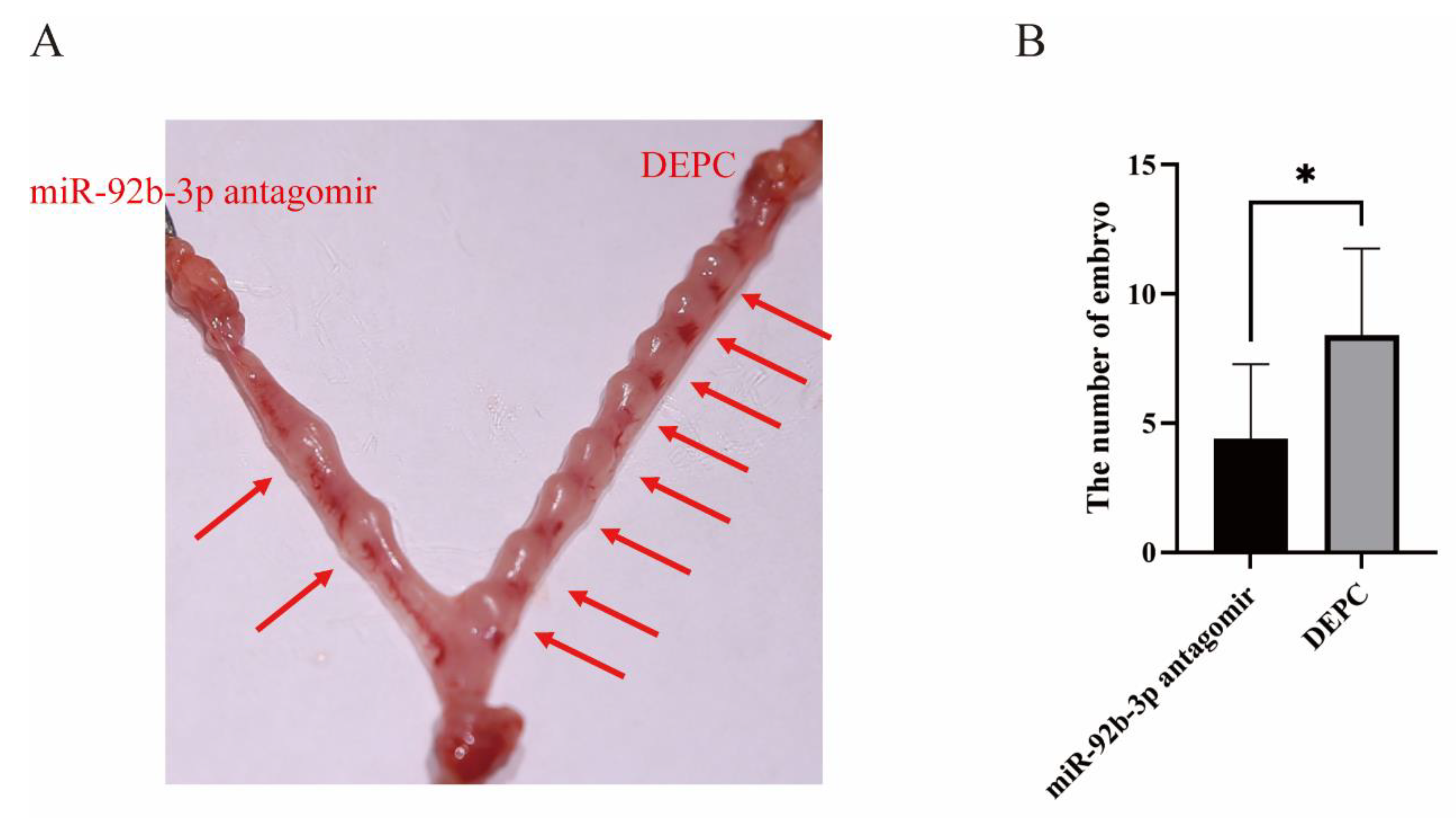
2.6. Inhibition of miR-92b-3p Hindered Mouse Embryo Implantation in a Mouse Model
3. Discussion
4. Materials and Methods
4.1. Sample Tissue Collection
4.2. FISH Assay
4.3. Cell Culture and Transfection
4.4. RNA Extraction and RT-qPCR
4.5. Cell Counting Kit-8 (CCK-8) Assay
4.6. EdU Assay
4.7. Cell Wound-Healing Assay
4.8. Western Blotting
4.9. Dual-Luciferase Reporter Gene Assay
4.10. Intrauterine Injection in Mice
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bidarimath, M.; Tayade, C. Pregnancy and spontaneous fetal loss: A pig perspective. Mol. Reprod. Dev. 2017, 84, 856–869. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.W.; Ashworth, M.D.; Stein, D.R.; Couture, O.P.; Tuggle, C.K.; Geisert, R.D. Identification of differential gene expression during porcine conceptus rapid trophoblastic elongation and attachment to uterine luminal epithelium. Physiol. Genom. 2009, 36, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E. Early pregnancy: Concepts, challenges, and potential solutions. Anim. Front. 2013, 3, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Bazer, F.W. Conceptus signals for establishment and maintenance of pregnancy. Anim. Reprod. Sci. 2004, 82–83, 537–550. [Google Scholar] [CrossRef]
- Bidarimath, M.; Khalaj, K.; Kridli, R.T.; Kan, F.W.K.; Koti, M.; Tayade, C. Extracellular vesicle mediated intercellular communication at the porcine maternal-fetal interface: A new paradigm for conceptus-endometrial cross-talk. Sci. Rep. 2017, 7, 40476. [Google Scholar] [CrossRef] [Green Version]
- Weckman, A.M.; Ngai, M.; Wright, J.; McDonald, C.R.; Kain, K.C. The Impact of Infection in Pregnancy on Placental Vascular Development and Adverse Birth Outcomes. Front. Microbiol. 2019, 10, 1924. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.J.; Jensen, S.S.; Lim, J.W.E. Proteomic profiling of exosomes: Current perspectives. Proteomics 2008, 8, 4083–4099. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Veerman, R.E.; Teeuwen, L.; Czarnewski, P.; Akpinar, G.G.; Sandberg, A.; Cao, X.; Pernemalm, M.; Orre, L.M.; Gabrielsson, S.; Eldh, M. Molecular evaluation of five different isolation methods for extracellular vesicles reveals different clinical applicability and subcellular origin. J. Extracell. Vesicles 2021, 10, e12128. [Google Scholar] [CrossRef]
- Ogawa, Y.; Miura, Y.; Harazono, A.; Kanai-Azuma, M.; Akimoto, Y.; Kawakami, H.; Yamaguchi, T.; Toda, T.; Endo, T.; Tsubuki, M.; et al. Proteomic Analysis of Two Types of Exosomes in Human Whole Saliva. Biol. Pharm. Bull. 2011, 34, 13–23. [Google Scholar] [CrossRef]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [Green Version]
- Poliakov, A.; Spilman, M.; Dokland, T.; Amling, C.L.; Mobley, J.A. Structural heterogeneity and protein composition of exosome-like vesicles (prostasomes) in human semen. Prostate 2009, 69, 159–167. [Google Scholar] [CrossRef]
- Hu, Q.; Zang, X.; Ding, Y.; Gu, T.; Shi, J.; Li, Z.; Cai, G.; Liu, D.; Wu, Z.; Hong, L. Porcine uterine luminal fluid-derived extracellular vesicles improve conceptus-endometrial interaction during implantation. Theriogenology 2021, 178, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Caby, M.-P.; Lankar, D.; Vincendeau-Scherrer, C.; Raposo, G.; Bonnerot, C. Exosomal-like vesicles are present in human blood plasma. Int. Immunol. 2005, 17, 879–887. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nature reviews. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Cai, G.; Meng, F.; Xu, Z.; He, Y.; Hu, Q.; Zheng, E.; Huang, S.; Xu, Z.; Gu, T.; et al. Deep-Sequencing Identification of MicroRNA Biomarkers in Serum Exosomes for Early Pig Pregnancy. Front. Genet. 2020, 11, 536. [Google Scholar] [CrossRef] [PubMed]
- Yadava, S.M.; Feng, A.; Parobchak, N.; Wang, B.; Rosen, T. miR-15b-5p promotes expression of proinflammatory cytokines in human placenta by inhibiting Apelin signaling pathway. Placenta 2021, 104, 8–15. [Google Scholar] [CrossRef]
- Hua, R.; Wang, Y.; Lian, W.; Li, W.; Xi, Y.; Xue, S.; Kang, T.; Lei, M. Small RNA-seq analysis of extracellular vesicles from porcine uterine flushing fluids during peri-implantation. Gene 2021, 766, 145117. [Google Scholar] [CrossRef]
- Hirschhaeuser, F.; Sattler, U.G.A.; Mueller-Klieser, W. Lactate: A Metabolic Key Player in Cancer. Cancer Res. 2011, 71, 6921–6925. [Google Scholar] [CrossRef] [Green Version]
- Zamudio, S.; Torricos, T.; Fik, E.; Oyala, M.; Echalar, L.; Pullockaran, J.; Tutino, E.; Martin, B.; Belliappa, S.; Balanza, E.; et al. Hypoglycemia and the origin of hypoxia-induced reduction in human fetal growth. PLoS ONE 2010, 5, e8551. [Google Scholar] [CrossRef] [PubMed]
- Kalisch-Smith, J.I.; Simmons, D.G.; Pantaleon, M.; Moritz, K.M. Sex differences in rat placental development: From pre-implantation to late gestation. Biol. Sex Differ. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Che, L.; Gao, K.; Wang, L.; Yang, X.; Wen, X.; Jiang, Z.; Wu, D. Effects of Dietary Taurine Supplementation to Gilts during Late Gestation and Lactation on Offspring Growth and Oxidative Stress. Animals 2019, 9, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.; Xiao, Y.; Chen, Y.; Wu, X.; Zhang, G.; Wang, Q.; Xie, C. Effects of catechins on litter size, reproductive performance and antioxidative status in gestating sows. Anim. Nutr. 2015, 1, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Anastasiou, D.; Poulogiannis, G.; Asara, J.M.; Boxer, M.B.; Jiang, J.K.; Shen, M.; Bellinger, G.; Sasaki, A.T.; Locasale, J.W.; Auld, D.S.; et al. Inhibition of Pyruvate Kinase M2 by Reactive Oxygen Species Contributes to Cellular Antioxidant Responses. Science 2011, 334, 1278–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaskar, S.U.; Chu, A. Intrauterine Growth Restriction: Hungry for an Answer. Physiology 2016, 31, 131–146. [Google Scholar] [CrossRef] [Green Version]
- Reliszko, Z.P.; Gajewski, Z.; Kaczmarek, M.M. Signs of embryo-maternal communication: miRNAs in the maternal serum of pregnant pigs. Reproduction 2017, 154, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Li, L.; Li, B.; Fang, X.; Ren, S. Long form leptin receptor and SNP effect on reproductive traits during embryo attachment in Suzhong sows. Anim. Reprod. Sci. 2016, 168, 57–65. [Google Scholar] [CrossRef]
- Fu, Y.; Li, L.; Fang, X.; Li, B.; Zhao, W.; Zhou, L.; Ren, S. Investigation of Eph-ephrin A1 in the regulation of embryo implantation in sows. Reprod. Domest. Anim. 2018, 53, 1563–1574. [Google Scholar] [CrossRef]
- Viganò, P.; Mangioni, S.; Pompei, F.; Chiodo, I. Maternal-conceptus Cross Talk—A Review. Placenta 2003, 24, S56–S61. [Google Scholar] [CrossRef]
- Zhou, Y.; Xia, M.; Cui, C.; Wei, H.; Jiang, S.; Peng, J. Circulating Exosomal miR-221 from Maternal Obesity Inhibits Angiogenesis via Targeting Angptl2. Int. J. Mol. Sci. 2021, 22, 10343. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Tan, Q.; Liang, J.; Cao, D.; Wang, S.; Liang, J.; Chen, K.; Wang, Z. Placental trophoblast cell-derived exosomal microRNA-1290 promotes the interaction between endometrium and embryo by targeting LHX6. Mol. Ther.-Nucleic Acids 2021, 26, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Q.; Xie, T.; Yuan, M.; Sheng, X.; Qi, X.; Xing, K.; Liu, F.; Guo, Y.; Xiao, L.; et al. Exosomes from bovine endometrial epithelial cells ensure trophoblast cell development by miR-218 targeting secreted frizzled related protein 2. J. Cell. Physiol. 2020, 236, 4565–4579. [Google Scholar] [CrossRef]
- Wu, Z.B.; Cai, L.; Lin, S.J.; Lu, J.L.; Yao, Y.; Zhou, L.F. The miR-92b functions as a potential oncogene by targeting on Smad3 in glioblastomas. Brain Res. 2013, 1529, 16–25. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, Z.; Wei, H.; Wu, S.; Wang, X.; Xiao, J. Promotion of tumour proliferation, migration and invasion bymiR-92b in targeting RECK in osteosarcoma. Clin. Sci. 2016, 130, 921–930. [Google Scholar] [CrossRef]
- Sengupta, S.; Nie, J.; Wagner, R.J.; Yang, C.; Stewart, R.; Thomson, J.A. MicroRNA 92b Controls the G1/S Checkpoint Gene p57 in Human Embryonic Stem Cells. Stem Cells 2009, 27, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Johnson, G.A. Pig blastocyst–uterine interactions. Differentiation 2014, 87, 52–65. [Google Scholar] [CrossRef]
- Tan, Q.; Shi, S.; Liang, J.; Cao, D.; Wang, S.; Wang, Z. Endometrial cell-derived small extracellular vesicle miR-100-5p promotes functions of trophoblast during embryo implantation. Mol. Ther. Nucleic Acids 2021, 23, 217–231. [Google Scholar] [CrossRef]
- Hasawi, N.A.; Alkandari, M.F.; Luqmani, Y.A. Phosphofructokinase: A mediator of glycolytic flux in cancer progression. Crit. Rev. Oncol./Hematol. 2014, 92, 312–321. [Google Scholar] [CrossRef]
- White, C.E.; Piper, E.L.; Noland, P.R. Conversion of glucose to fructose in the fetal pig. J. Anim. Sci. 1979, 48, 585. [Google Scholar] [CrossRef] [Green Version]
- Yi, W.; Clark, P.M.; Mason, D.E.; Keenan, M.C.; Hill, C.; Goddard, W.A.; Peters, E.C.; Driggers, E.M.; Hsieh-Wilson, L.C. Phosphofructokinase 1 Glycosylation Regulates Cell Growth and Metabolism. Science 2012, 337, 975–980. [Google Scholar] [CrossRef]
- Ali, A.; Bouma, G.J.; Anthony, R.V.; Winger, Q.A. The Role of LIN28-let-7-ARID3B Pathway in Placental Development. Int. J. Mol. Sci. 2020, 21, 3637. [Google Scholar] [CrossRef]
- Jaeger, L.A.; Spiegel, A.K.; Ing, N.H.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C. Functional Effects of Transforming Growth Factor β on Adhesive Properties of Porcine Trophectoderm. Endocrinology 2005, 146, 3933–3942. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhou, C.; Meng, F.; Hu, Q.; Ding, Y.; Wang, X.; Gu, T.; Li, Z.; Wu, Z.; Hong, L.; et al. Ssc-miR-92b-3p Regulates Porcine Trophoblast Cell Proliferation and Migration via the PFKM Gene. Int. J. Mol. Sci. 2022, 23, 16138. https://doi.org/10.3390/ijms232416138
Wang Y, Zhou C, Meng F, Hu Q, Ding Y, Wang X, Gu T, Li Z, Wu Z, Hong L, et al. Ssc-miR-92b-3p Regulates Porcine Trophoblast Cell Proliferation and Migration via the PFKM Gene. International Journal of Molecular Sciences. 2022; 23(24):16138. https://doi.org/10.3390/ijms232416138
Chicago/Turabian StyleWang, Yongzhong, Chen Zhou, Fanming Meng, Qun Hu, Yue Ding, Xiaoliang Wang, Ting Gu, Zicong Li, Zhenfang Wu, Linjun Hong, and et al. 2022. "Ssc-miR-92b-3p Regulates Porcine Trophoblast Cell Proliferation and Migration via the PFKM Gene" International Journal of Molecular Sciences 23, no. 24: 16138. https://doi.org/10.3390/ijms232416138