
The Anthocyanin Accumulation Related ZmBZ1, Facilitates Seedling Salinity Stress Tolerance via ROS Scavenging
 , ,
, ,  and
and 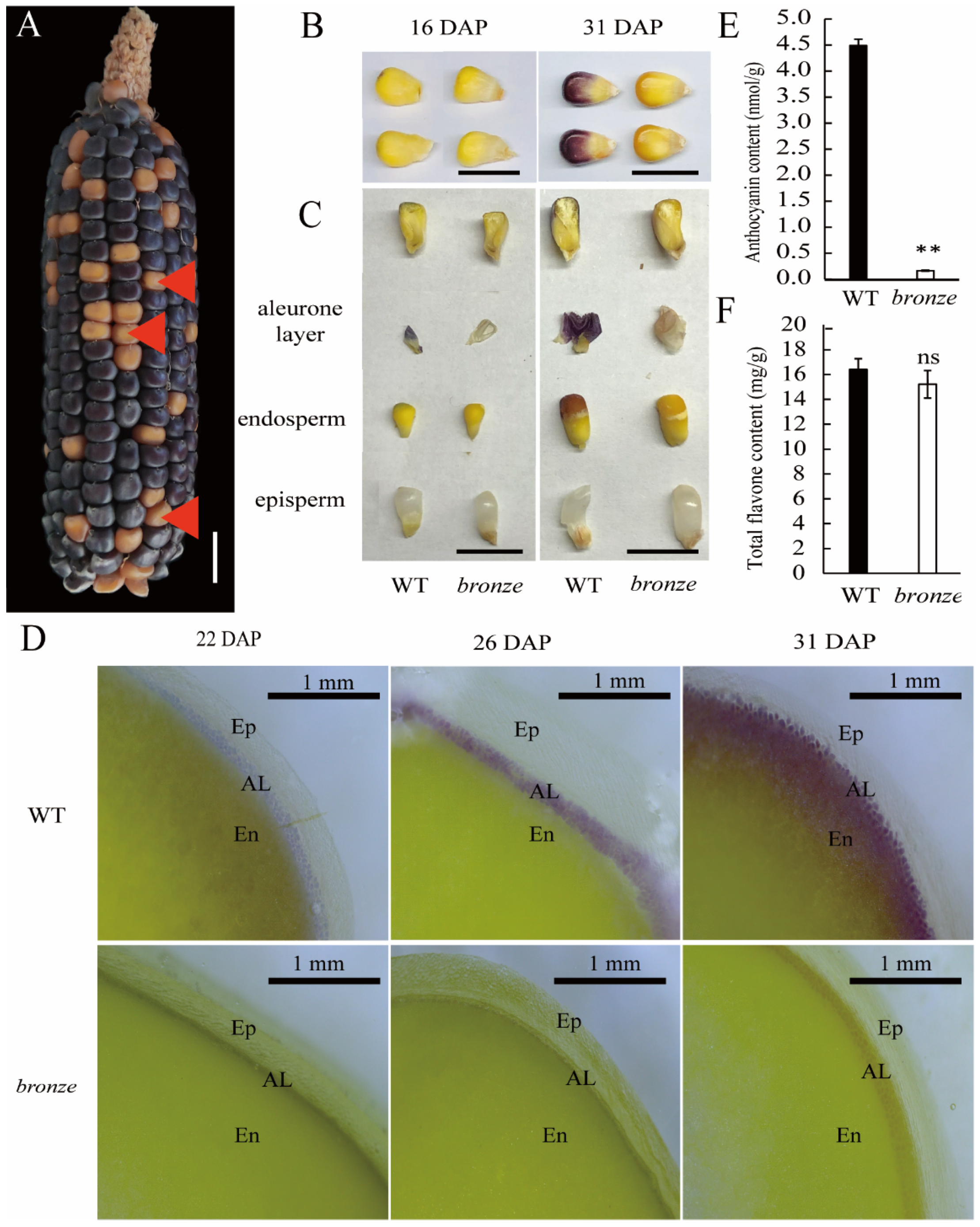{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. A New Isolated Maize Bronze Mutant Deposits Pigment in Organs of Seeds, Anther and Stem
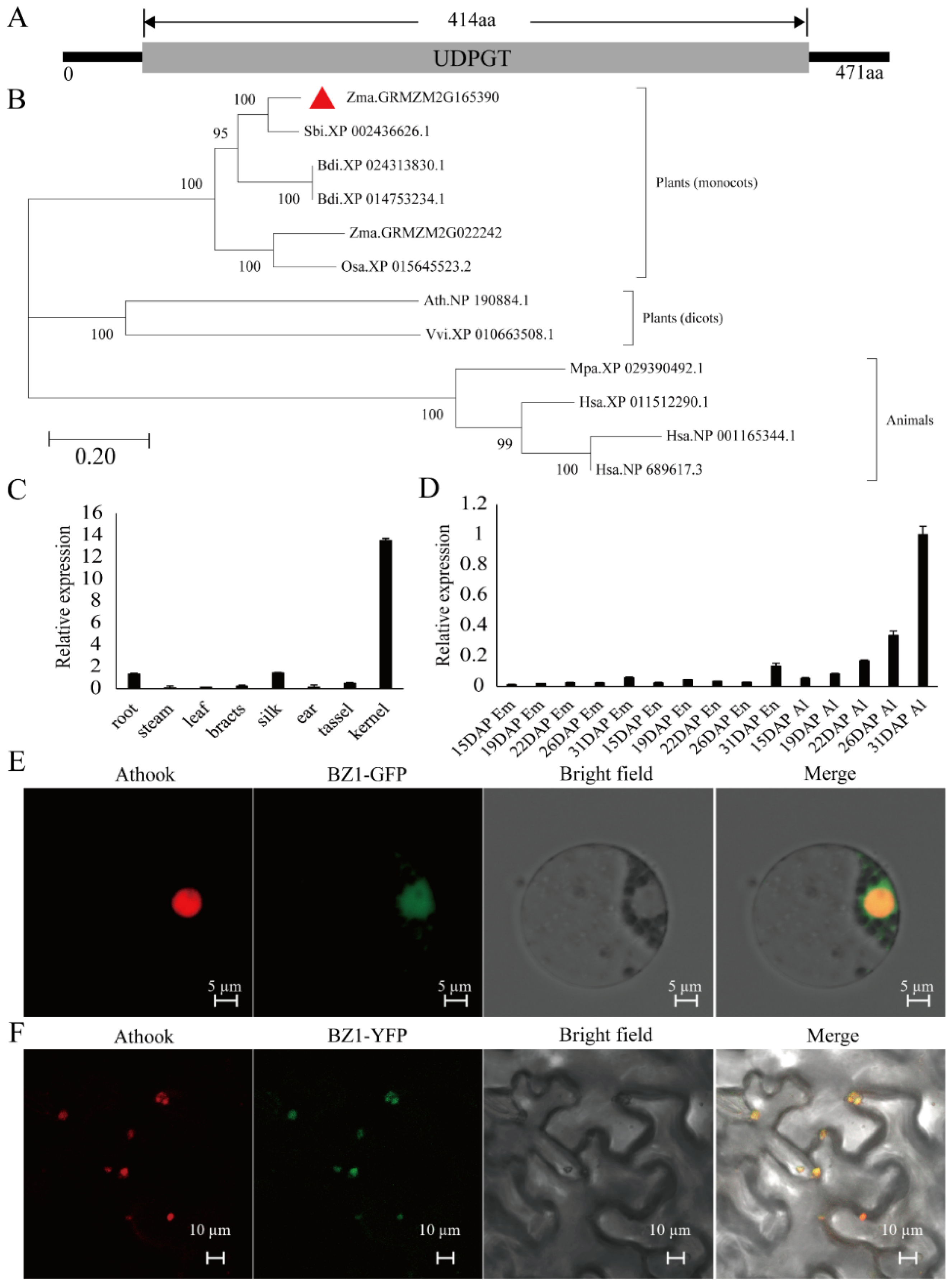
2.2. Mapping and Cloning of ZmBZ1 in Modified W22
2.3. The Nucleus-Located ZmBZ1 (Anthocyanidin 3-O-glucosyltransferase) Is Tissue-Specific Expressed in Seed Aleurone Layer
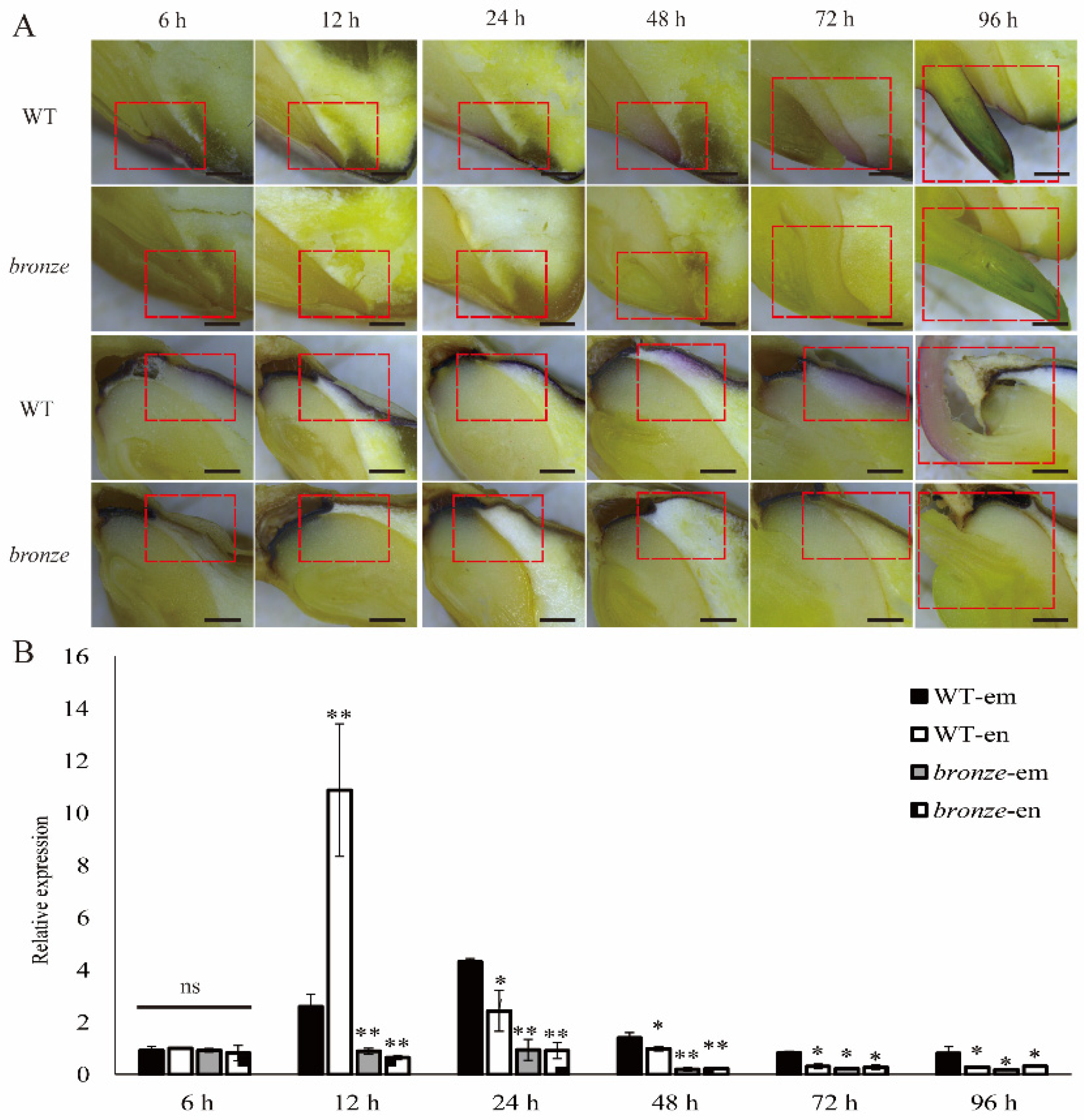
2.4. Spatio-Temporal Expression Pattern of ZmBZ1 Is Corresponded with Anthocyanin Movement during Seeds Germinating
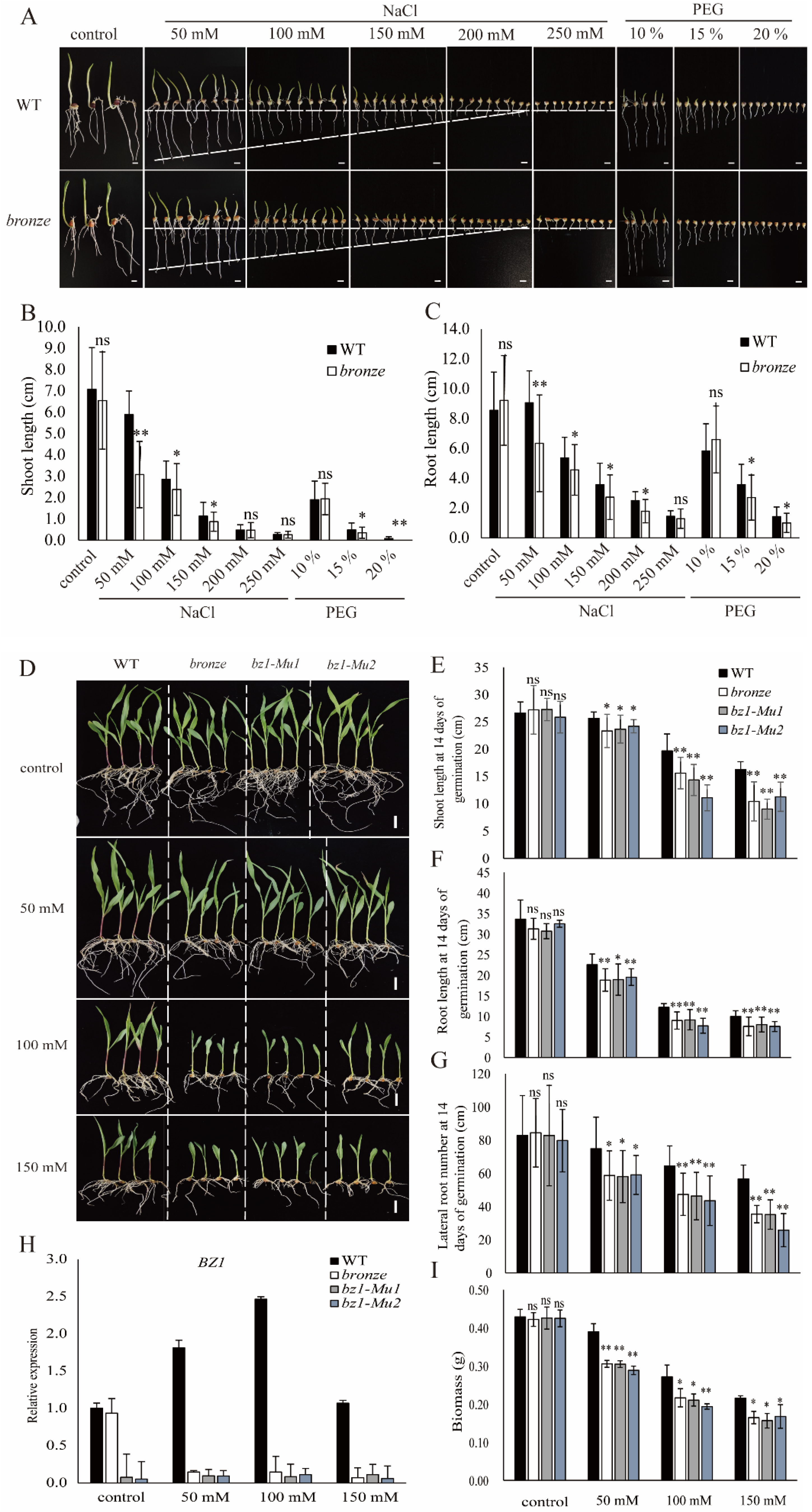
2.5. ZmBZ1 Is Essential for Stress Tolerance and Was Strongly Induced by Salinity Stress
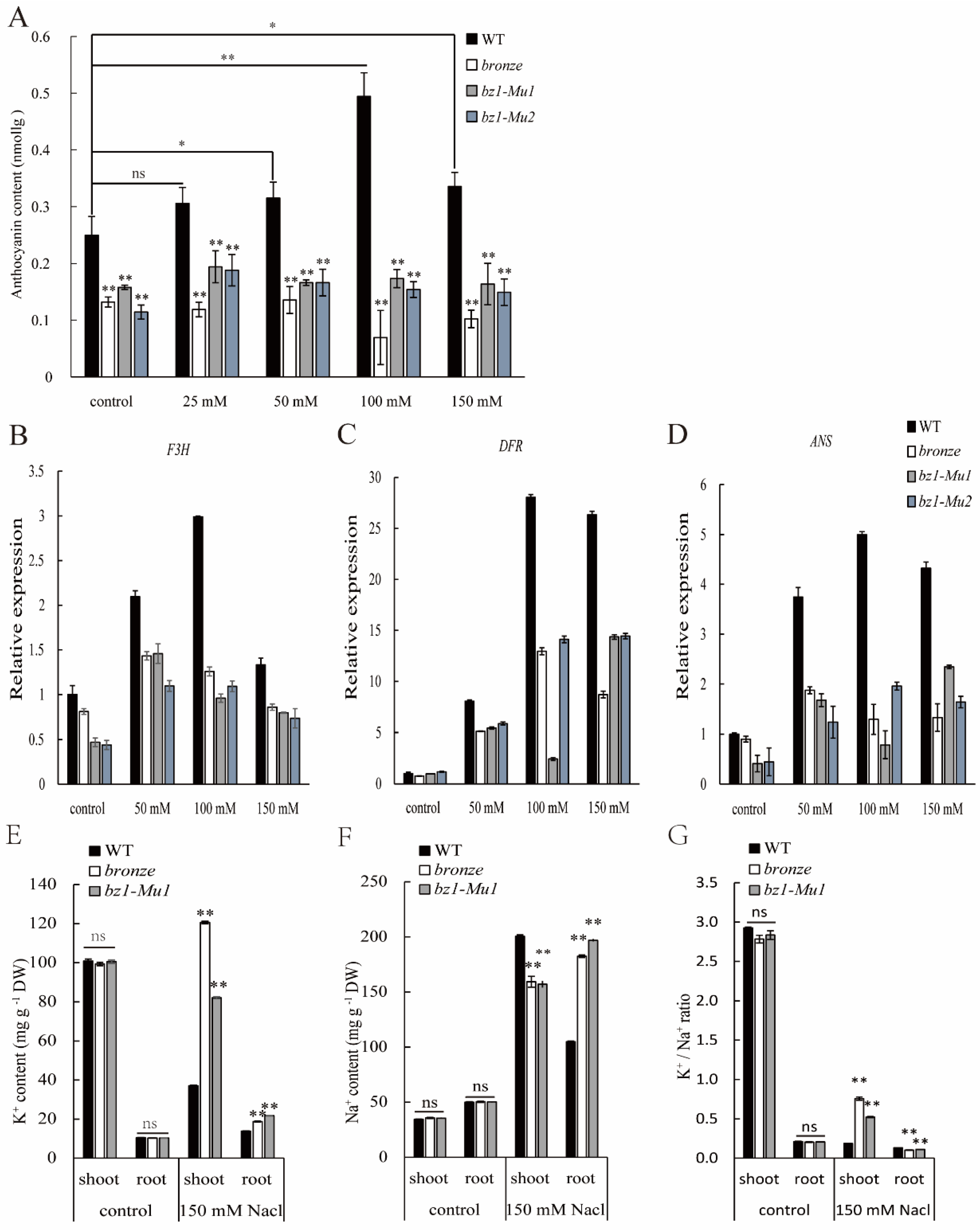
2.6. ZmBZ1 Promoted Seedling Salinity Stress Tolerance by Regulating of Anthocyanin Accumulation
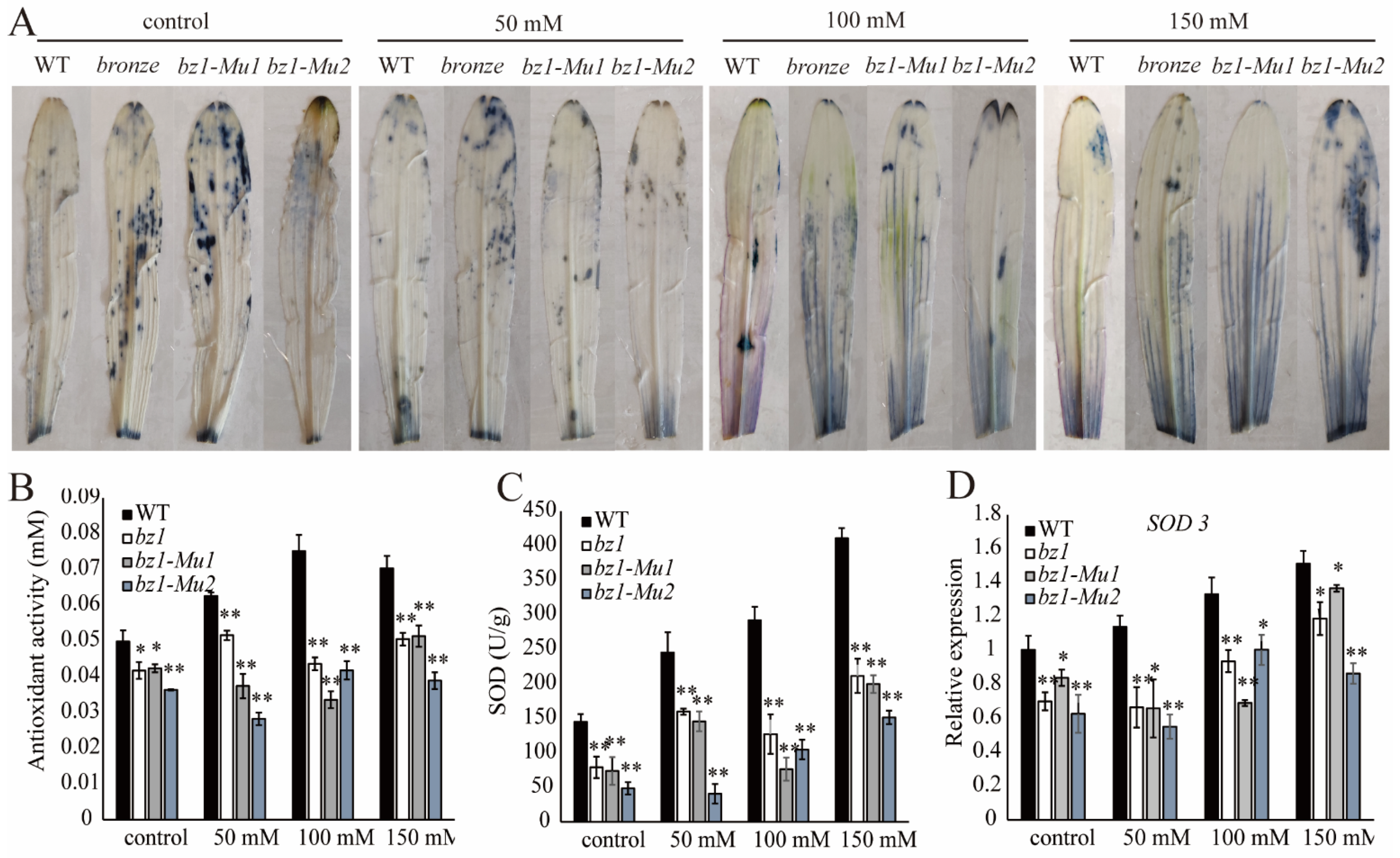
2.7. ZmBZ1 Enhanced ROS Scavenging under Salinity Stress
3. Discussion
3.1. ZmBZ1 Enhanced the Salinity Stress Tolerance in Maize through Atypical ROS Scavenging Way
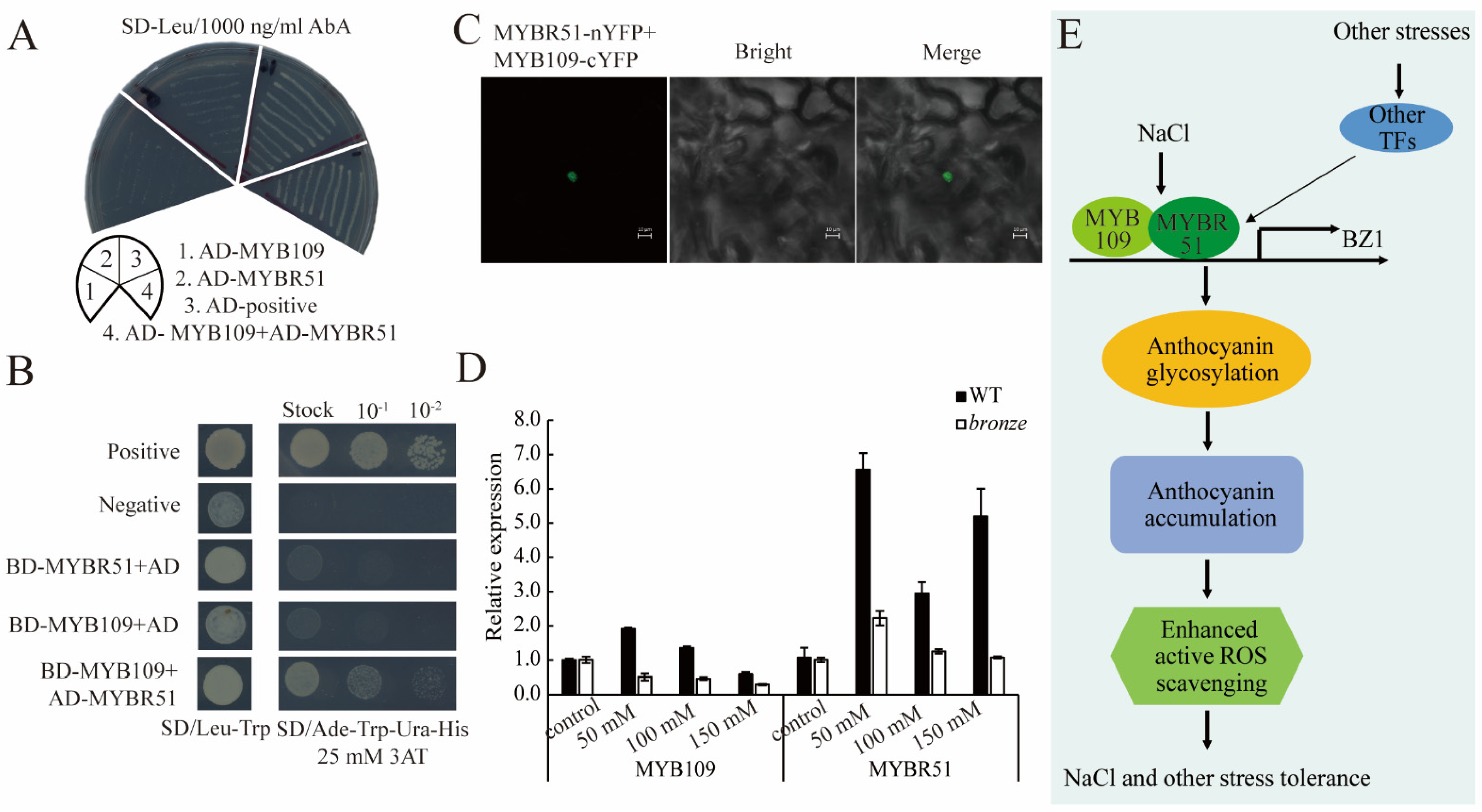
3.2. Anthocyanins Accumulation Was Induced by Extreme Environments and MYB109 and MYBR51
4. Materials and Methods
4.1. Plant Materials
4.2. Sections and Microscopy Observation
4.3. BSR-Seq and Fine Mapping Analysis
4.4. RNA Extraction and qRT-PCR Analysis
4.5. Subcellular Localization
4.6. Phylogenetic Analysis
4.7. Assays for Stress Treatment
4.8. Quantification of Na+ and K+ Concentration
4.9. Quantification of Anthocyanins
4.10. NBT and DAB Staining
4.11. Determination of Antioxidant Capacity
4.12. Y1H, Y2H and BiFC Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Lee, W.J.; Vu, T.T.; Jeong, C.Y.; Hong, S.W.; Lee, H. High accumulation of anthocyanins via the ectopic expression of AtDFR confers significant salt stress tolerance in Brassica napus L. Plant Cell Rep. 2017, 36, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yang, G.; Zhang, J.; Wang, Y.; Zhang, T.; Wang, N.; Jiang, S.; Zhang, Z.; Chen, X. Overexpression of a repressor MdMYB15L negatively regulates anthocyanin and cold tolerance in red-fleshed callus. Biochem. Bioph. Res. Co. 2018, 500, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Hanly, A.; Karagiannis, J.; Lu, Q.S.M.; Tian, L.; Hannoufa, A. Characterization of the Role of SPL9 in Drought Stress Tolerance in Medicago sativa. Int. J. Mol. Sci. 2020, 21, 6003. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Q.; Yang, G.; Zhang, C.; Dong, H.; Liu, Y.; Yin, R.; Lin, L. Pivotal roles of Tomato photoreceptor SlUVR8 in seedling development and UV-B stress tolerance. Biochem. Bioph. Res. Co. 2020, 522, 177–183. [Google Scholar] [CrossRef]
- Lin, Y.; Fan, L.; He, J.; Wang, Z.; Yin, Y.; Cheng, Y.; Li, Z. Anthocyanins contribute to fruit defense against postharvest green mold. Postharvest Biol. Tec. 2021, 181, 111661. [Google Scholar] [CrossRef]
- Yang, M.; Koo, S.I.; Song, W.O.; Chun, O.K. Food Matrix Affecting Anthocyanin Bioavailability: Review. Curr. Med. Chem. 2011, 18, 291–300. [Google Scholar] [CrossRef]
- Dong, X.X.; Braun, E.L.E.L.; Grotewold, E.E. Functional Conservation of Plant Secondary Metabolic Enzymes Revealed by Complementation of Arabidopsis Flavonoid Mutants with Maize Genes1. Plant Physiol. 2001, 127, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Wisman, E.; Hartmann, U.; Sagasser, M.; Baumann, E.; Palme, K.; Hahlbrock, K.; Saedler, H.; Weisshaar, B. Knock-out mutants from an En-1 mutagenized Arabidopsis thaliana population generate phenylpropanoid biosynthesis phenotypes. Proc. Natl. Acad. Sci. USA 1998, 95, 12432–12437. [Google Scholar] [CrossRef] [Green Version]
- Meyer, P.; Heidmann, I.; Forkmann, G.; Saedler, H. A New Petunia Flower Color Generated By Transformation of a Mutant with a Maize Gene. Nature 1987, 330, 677–678. [Google Scholar] [CrossRef]
- Nakatsuka, T.; Nishihara, M.; Mishiba, K.; Yamamura, S. Two different mutations are involved in the formation of white-flowered gentian plants. Plant Sci. 2005, 169, 949–958. [Google Scholar] [CrossRef]
- Kobayashi, S.; Ishimaru, M.; Ding, C.K.; Yakushiji, H.; Goto, N. Comparison of UDP-glucose:flavonoid 3-O-glucosyltransferase (UFGT) gene sequences between white grapes (Vitis vinifera) and their sports with red skin. Plant Sci. 2001, 160, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Mano, H.; Ogasawara, F.; Sato, K.; Higo, H.; Minobe, Y. Isolation of a Regulatory Gene of Anthocyanin Biosynthesis in Tuberous Roots of Purple-Fleshed Sweet Potato. Plant Physiol. 2007, 143, 1252–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, J.; Derynck, M.R.; Li, X.; Telmer, P.; Marsolais, F.; Dhaubhadel, S. A single-repeat MYB transcription factor, GmMYB176, regulates CHS8 gene expression and affects isoflavonoid biosynthesis in soybean. Plant J. 2010, 62, 1019–1034. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.M.; Heine, G.F.; Irani, N.G.; Feller, A.; Kim, M.; Matulnik, T.; Chandler, V.L.; Grotewold, E. Different Mechanisms Participate in the R-dependent Activity of the R2R3 MYB Transcription Factor C1. J. Biol. Chem. 2004, 279, 48205–48213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, N.A.; Glover, B.J. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005, 10, 63–70. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The Roles of Reactive Oxygen Metabolism in Drought: Not So Cut and Dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Liu, C.; Tang, D.; Yan, L.; Wang, D.; Yang, Y.; Gui, J.; Zhao, X.; Li, L.; Tang, X.; et al. The Receptor-Like Cytoplasmic Kinase STRK1 Phosphorylates and Activates CatC, Thereby Regulating H2O2 Homeostasis and Improving Salt Tolerance in Rice. Plant Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef] [Green Version]
- Schneider, J.R.; Caverzan, A.; Chavarria, G. Water deficit stress, ROS involvement, and plant performance. Arch. Agron. Soil Sci. 2019, 65, 1160–1181. [Google Scholar] [CrossRef]
- Song, Q.; Wang, X.; Liu, Y.; Brestic, M.; Yang, X. StLTO1, a lumen thiol oxidoreductase in Solanum tuberosum L., enhances the cold resistance of potato plants. Plant Sci. 2022, 325, 111481. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ying, Y.; Chen, J.; Wang, X. Transgenic Arabidopsis overexpressing Mn-SOD enhanced salt-tolerance. Plant Sci. 2004, 167, 671–677. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, K.; Zheng, Y.; Wang, Y.; Wang, J.; Liao, H. Disruption of AtWNK8 Enhances Tolerance of Arabidopsis to Salt and Osmotic Stresses via Modulating Proline Content and Activities of Catalase and Peroxidase. Int. J. Mol. Sci. 2013, 14, 7032–7047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmond, C.B.; Grace, S.C. Perspectives on Photoinhibition and Photorespiration in The Field-Quintessential Inefficiencies of The Light and Dark Reactions of Photosynthesis. J. Exp. Bot. 1995, 46, 1351–1362. [Google Scholar] [CrossRef]
- Addi, M.; Elbouzidi, A.; Abid, M.; Tungmunnithum, D.; Elamrani, A.; Hano, C. An Overview of Bioactive Flavonoids from Citrus Fruits. Appl. Sci. 2022, 12, 29. [Google Scholar] [CrossRef]
- Bhatia, C.; Pandey, A.; Gaddam, S.R.; Hoecker, U.; Trivedi, P.K. Low Temperature-Enhanced Flavonol Synthesis Requires Light-Associated Regulatory Components in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 2099–2112. [Google Scholar] [CrossRef] [Green Version]
- Guan, L.; Dai, Z.; Wu, B.; Wu, J.; Merlin, I.; Hilbert, G.; Renaud, C.; Gomès, E.; Edwards, E.; Li, S.; et al. Anthocyanin biosynthesis is differentially regulated by light in the skin and flesh of white-fleshed and teinturier grape berries. Planta 2016, 243, 23–41. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Yamamoto, T.; Ikeda, H.; Sugihara, R.; Kaihori, H.; Kawabata, M.; Suzuki, S. Effects of constantly high soil water content on vegetative growth and grape quality in Japan with high rainfall during grapevine growing season. Folia Hortic. 2020, 32, 135–145. [Google Scholar] [CrossRef]
- Gu, R.; Li, L.; Liang, X.; Wang, Y.; Fan, T.; Wang, Y.; Wang, J. The ideal harvest time for seeds of hybrid maize (Zea mays L.) XY335 and ZD958 produced in multiple environments. Sci. Rep. 2017, 7, 17537. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wu, X.; Wang, H.; Feng, X.; Yan, S.; Wu, S.; Liu, J.; Yao, X.; Bai, A.; Zhao, H.; et al. Defective mitochondrial function by mutation in Thick Aleurone 1 encoding a mitochondrion-targeted single-stranded DNA-binding protein leads to increased aleurone cell layers and improved nutrition in rice. Mol. Plant 2022, 15, 1638–1639. [Google Scholar] [CrossRef]
- Ming, H.; Wang, Q.; Wu, Y.; Liu, H.; Zheng, L.; Zhang, G. Transcriptome analysis reveals the mechanism of anthocyanidins biosynthesis during grains development in purple corn (Zea mays L.). J. Plant Physiol. 2021, 257, 153328. [Google Scholar] [CrossRef] [PubMed]
- Roth, B.A.; Goff, S.A.; Klein, T.M.; Fromm, M.E. C1-Dependent and R-dependent Expression of the Maize BZ1 Gene Requires Sequences with Homology to Mammalian MYB and MYC Binding-Sites. Plant Cell 1991, 3, 317–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, D.A.; Wang, L.J.; Ding, C.H.; Xu, Z.H. Auxin distribution and transport during embryogenesis and seed germination of Arabidopsis. Cell Res. 2001, 11, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.; Zhang, F.; Zhang, G.; Jiang, X.; Yu, H.; Hou, B. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. Plant J. 2017, 89, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivas-Aguirre, F.; Rodrigo-García, J.; Martínez-Ruiz, N.; Cárdenas-Robles, A.; Mendoza-Díaz, S.; Álvarez-Parrilla, E.; González-Aguilar, G.; de la Rosa, L.; Ramos-Jiménez, A.; Wall-Medrano, A. Cyanidin-3-O-glucoside: Physical-Chemistry, Foodomics and Health Effects. Molecules 2016, 21, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Liu, S.; Zhao, Z.; You, L.; Harrison, M.D.; Zhang, Z. Enzymatic acylation of cyanidin-3-glucoside with fatty acid methyl esters improves stability and antioxidant activity. Food Chem. 2021, 343, 128482. [Google Scholar] [CrossRef]
- Al-Aghabary, K.; Zhu, Z.; Shi, Q.H. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr. 2004, 27, 2101–2115. [Google Scholar] [CrossRef]
- Wang, Y.H.; Zhang, L.R.; Zhang, L.L.; Xing, T.; Peng, J.Z.; Sun, S.L.; Chen, G.; Wang, X.J. A novel stress-associated protein SbSAP14 from Sorghum bicolor confers tolerance to salt stress in transgenic rice. Mol. Breed. 2013, 32, 437–449. [Google Scholar] [CrossRef]
- Wang, T.; Xia, X.; Zhao, M.; Tian, Q.; Zhang, W. Expression of a Medicago falcata small GTPase gene, MfARL1 enhanced tolerance to salt stress in Arabidopsis thaliana. Plant Physiol. Bioch. 2013, 63, 227–235. [Google Scholar] [CrossRef]
- Zhou, X.; Li, J.; Wang, Y.; Liang, X.; Zhang, M.; Lu, M.; Guo, Y.; Qin, F.; Jiang, C. The classical SOS pathway confers natural variation of salt tolerance in maize. New Phytol. 2022, 236, 479–494. [Google Scholar] [CrossRef]
- Wu, X.X.; Li, J.; Wu, X.D.; Liu, Q.; Wang, Z.K.; Liu, S.S.; Li, S.N.; Ma, Y.L.; Sun, J.; Zhao, L.; et al. Ectopic expression of Arabidopsis thaliana Na+(K+)/H+ antiporter gene, AtNHX5, enhances soybean salt tolerance. Genet. Mol. Res. 2016, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Huang, R.F. The phytohormonal regulation of Na+/K+ and reactive oxygen species homeostasis in rice salt response. Mol. Breed. 2020, 40, 1–13. [Google Scholar] [CrossRef]
- Yan, F.; Wei, H.; Ding, Y.; Li, W.; Chen, L.; Ding, C.; Tang, S.; Jiang, Y.; Liu, Z.; Li, G. Melatonin enhances Na+/K+ homeostasis in rice seedlings under salt stress through increasing the root H+-pump activity and Na+/K+ transporters sensitivity to ROS/RNS. Environ. Exp. Bot. 2021, 182, 104328. [Google Scholar] [CrossRef]
- Taha, R.; Mills, D.; Heimer, Y.; Tal, M. The relation between low K+/Na+ ratio and salt-tolerance in the wild tomato species Lycopersicon pennellii. J. Plant Physiol. 2000, 157, 59–64. [Google Scholar] [CrossRef]
- Bian, S.M.; Jin, D.H.; Sun, G.Q.; Shan, B.H.; Zhou, H.N.; Wang, J.Y.; Zhai, L.L.; Li, X.Y. Characterization of the soybean R2R3-MYB transcription factor GmMYB81 and its functional roles under abiotic stresses. Gene 2020, 753, 144803. [Google Scholar] [CrossRef]
- Wei, Q.H.; Chen, R.; Wei, X.; Liu, Y.H.; Zhao, S.J.; Yin, X.P.; Xie, T. Genome-wide identification of R2R3-MYB family in wheat and functional characteristics of the abiotic stress responsive gene TaMYB344. BMC Genom. 2020, 21, 1–16. [Google Scholar] [CrossRef]
- Zhao, K.; Cheng, Z.H.; Guo, Q.; Yao, W.J.; Liu, H.J.; Zhou, B.R.; Jiang, T.B. Characterization of the Poplar R2R3-MYB Gene Family and Over-Expression of PsnMYB108 Confers Salt Tolerance in Transgenic Tobacco. Front. Plant Sci. 2020, 11, 1548. [Google Scholar] [CrossRef]
- Shen, Y.; Sun, T.; Pan, Q.; Anupol, N.; Chen, H.; Shi, J.; Liu, F.; Deqiang, D.; Wang, C.; Zhao, J.; et al. RrMYB5- and RrMYB10-regulated flavonoid biosynthesis plays a pivotal role in feedback loop responding to wounding and oxidation inRosa rugosa. Plant Biotechnol. J. 2019, 17, 2078–2095. [Google Scholar] [CrossRef] [Green Version]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Yang, F.; Li, W.; Jiang, N.; Yu, H.D.; Morohashi, K.; Ouma, W.Z.; Morales-Mantilla, D.E.; Gomez-Cano, F.A.; Mukundi, E.; Prada-Salcedo, L.D.; et al. A Maize Gene Regulatory Network for Phenolic Metabolism. Mol. Plant 2017, 10, 498–515. [Google Scholar] [CrossRef]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Mehrtens, F.; Niehaus, K.; Weisshaar, B. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.; Man, C.; Xie, Y.; Yan, J.; Chu, J.; Huang, J. A Crucial Role of GA-Regulated Flavonol Biosynthesis in Root Growth of Arabidopsis. Mol. Plant 2019, 12, 521–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, P.J.; Alfenito, M.R.; Walbot, V. Impact of Low-Temperature Stress on General Phenylpropanoid and Anthocyann Pathways-Enhancement of Transcript Abundance and Anthocyanin Pigmentation in Maize Seedlings. Planta 1994, 194, 541–549. [Google Scholar] [CrossRef]
- Beathard, C.; Mooney, S.; Al-Saharin, R.; Goyer, A.; Hellmann, H. Characterization of Arabidopsis thaliana R2R3 S23 MYB Transcription Factors as Novel Targets of the Ubiquitin Proteasome-Pathway and Regulators of Salt Stress and Abscisic Acid Response. Front. Plant Sci. 2021, 12, 629208. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.C.; Leitão, C.A.E. Utilisation of Toluidine blue O pH 4.0 and histochemical inferences in plant sections obtained by free-hand. Protoplasma 2020, 257, 993–1008. [Google Scholar] [CrossRef]
- Liu, S.; Yeh, C.; Tang, H.M.; Nettleton, D.; Schnable, P.S.; Hazen, S.P. Gene mapping via bulked segregant RNA-Seq (BSR-Seq). PLoS ONE 2012, 7, e36406. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zhang, J.; Wang, J.; Xie, Y.; Cui, Y.; Du, X.; Li, L.; Fu, J.; Liu, Y.; Wang, J.; et al. Small kernel 501 (smk501) encodes the RUBylation activating enzyme E1 subunit ECR1 (E1 C-TERMINAL RELATED 1) and is essential for multiple aspects of cellular events during kernel development in maize. New Phytol. 2021, 230, 2337–2354. [Google Scholar] [CrossRef]
- Shahbazi, E.; Arzani, A.; Saeidi, G. Effects of NaCl Treatments on Seed Germination and Antioxidant Activity of Canola (Brassica napus L.) Cultivars. Bangl. J. Bot. 2011, 40, 67–73. [Google Scholar] [CrossRef]
- Vaz-de-Melo, A.; Santos, L.; Finoto, E.L.; Dias, D.; Alvarenga, E.M. Germination and Vigor of Popcorn Seeds Submitted to Thermal and Waterstress. Biosci. J. 2012, 28, 229–687. [Google Scholar]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Cao, Y.; Wang, Z.; Wang, Z.Q.; Shi, J.; Liang, X.; Song, W.; Chen, Q.; Lai, J.; Jiang, C. A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na+ exclusion and salt tolerance in maize. N. Phytol. 2018, 217, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, B.; David, L.; Donca, R.; Chisbora, C. Degradation Kinetics of Anthocyanins from Crude Ethanolic Extract from Sour Cherries. Stud. Univ. Babes-Bolyai Chem. 2011, 56, 189–194. [Google Scholar]
- Xiong, J.; Zhang, W.; Zheng, D.; Xiong, H.; Feng, X.; Zhang, X.; Wang, Q.; Wu, F.; Xu, J.; Lu, Y. ZmLBD5 Increases Drought Sensitivity by Suppressing ROS Accumulation in Arabidopsis. Plants 2022, 11, 1382. [Google Scholar] [CrossRef] [PubMed]
- Avan, A.N.; Cekic, S.D.; Uzunboy, S.; Apak, R. Spectrophotometric Determination of Phenolic Antioxidants in the Presence of Thiols and Proteins. Int. J. Mol. Sci. 2016, 17, 1325. [Google Scholar] [CrossRef] [PubMed]
- Jung, S. Effect of chlorophyll reduction in Arabidopsis thaliana by methyl jasmonate or norflurazon on antioxidant systems. Plant Physiol. Bioch. 2004, 42, 225–231. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, X.H.; Xiong, Y.P.; Niu, M.Y.; Zhang, Y.Y.; Yan, H.F.; Li, Y.; Zhang, X.H.; Ma, G.H. Identification and Functional Analysis of SabHLHs in Santalum album L. Life 2022, 12, 1017. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, D.; Peng, Y.; Cai, M.; Liang, Z.; Yuan, Z.; Du, X.; Wang, J.; Schnable, P.S.; Gu, R.; et al. The Anthocyanin Accumulation Related ZmBZ1, Facilitates Seedling Salinity Stress Tolerance via ROS Scavenging. Int. J. Mol. Sci. 2022, 23, 16123. https://doi.org/10.3390/ijms232416123
Wang J, Li D, Peng Y, Cai M, Liang Z, Yuan Z, Du X, Wang J, Schnable PS, Gu R, et al. The Anthocyanin Accumulation Related ZmBZ1, Facilitates Seedling Salinity Stress Tolerance via ROS Scavenging. International Journal of Molecular Sciences. 2022; 23(24):16123. https://doi.org/10.3390/ijms232416123
Chicago/Turabian StyleWang, Jie, Delin Li, Yixuan Peng, Minghao Cai, Zhi Liang, Zhipeng Yuan, Xuemei Du, Jianhua Wang, Patrick S. Schnable, Riliang Gu, and et al. 2022. "The Anthocyanin Accumulation Related ZmBZ1, Facilitates Seedling Salinity Stress Tolerance via ROS Scavenging" International Journal of Molecular Sciences 23, no. 24: 16123. https://doi.org/10.3390/ijms232416123