
A New Transgenic Tool to Study the Ret Signaling Pathway in the Enteric Nervous System
 ,
,
Abstract
:1. Introduction
2. Results
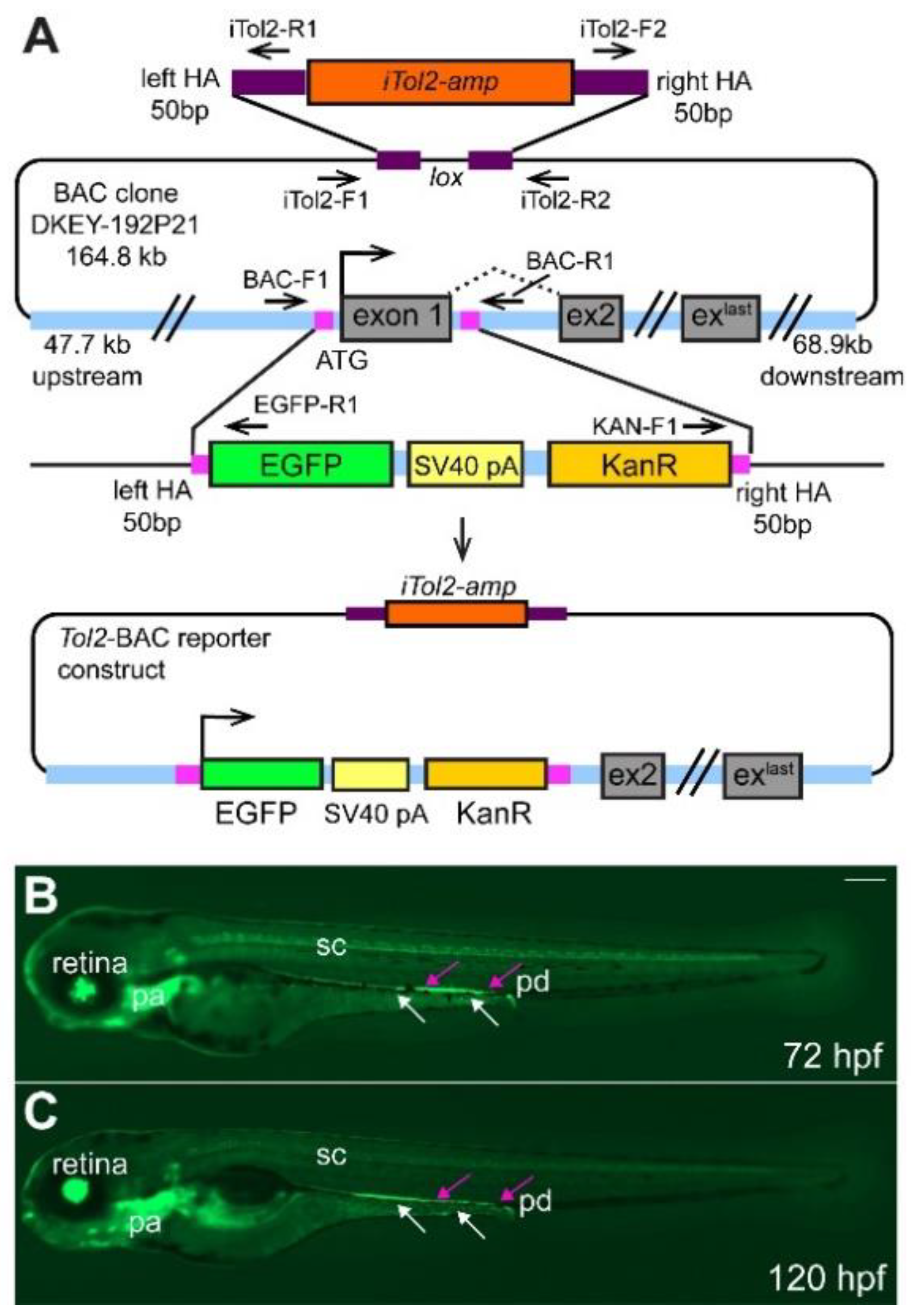
2.1. Generation of TgBAC(ret:EGFP)b1331 Transgenic Line
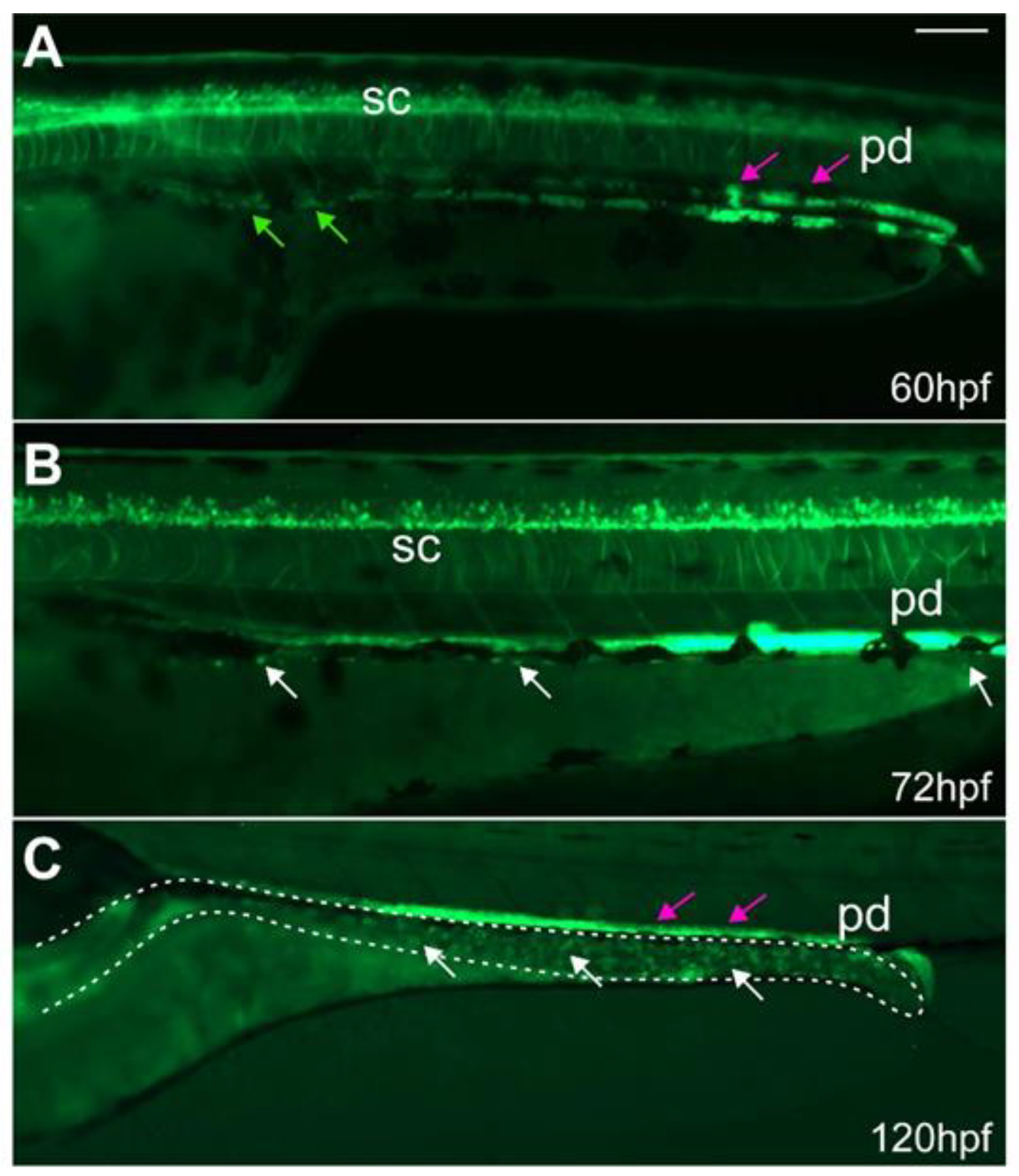
2.2. ret:GFP Is Expressed in Migrating Enteric Progenitor Cells and ENS Cells at Larval Stages
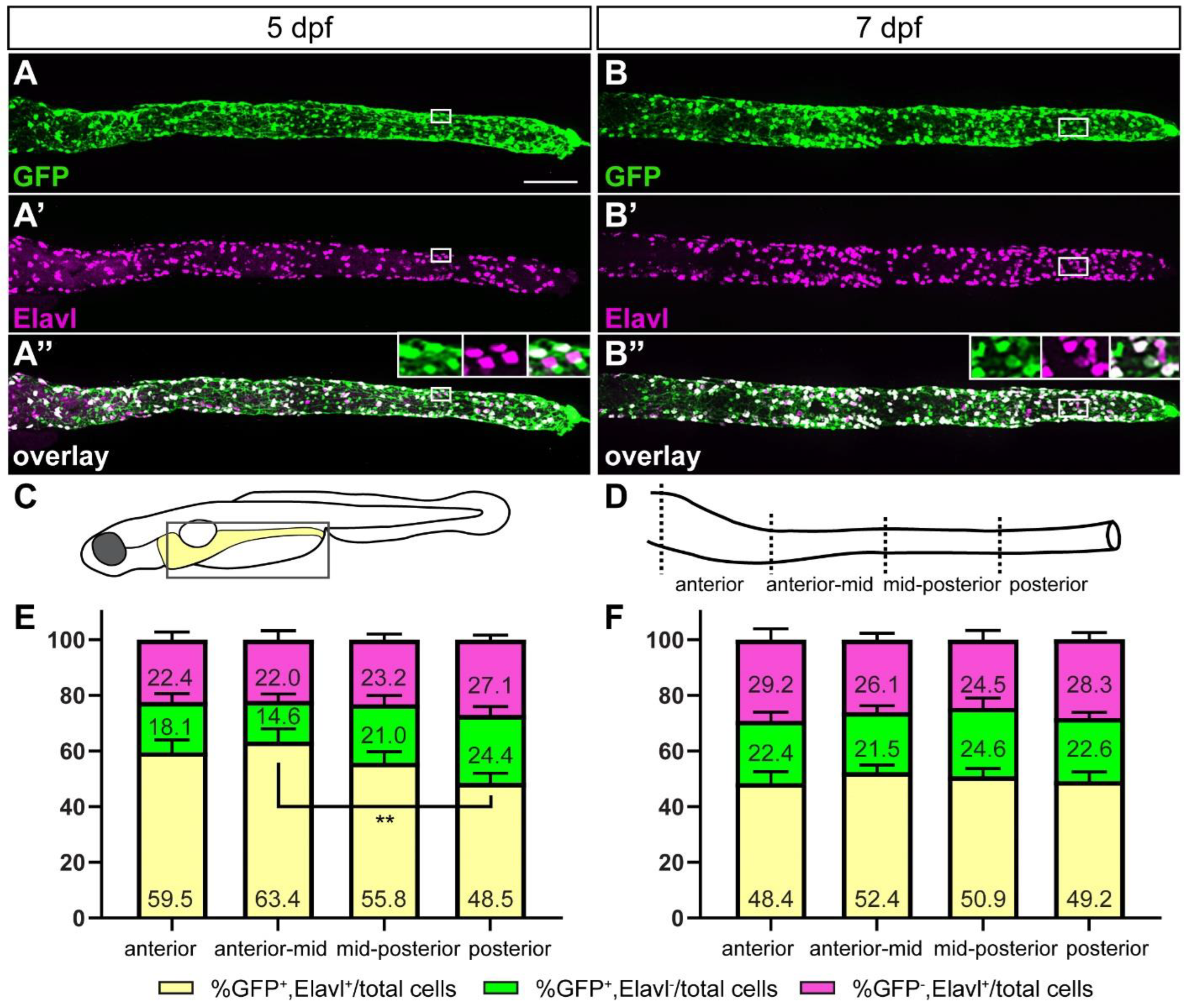
2.3. The Majority of ret:GFP Cells in the Gut Are ENS Neurons
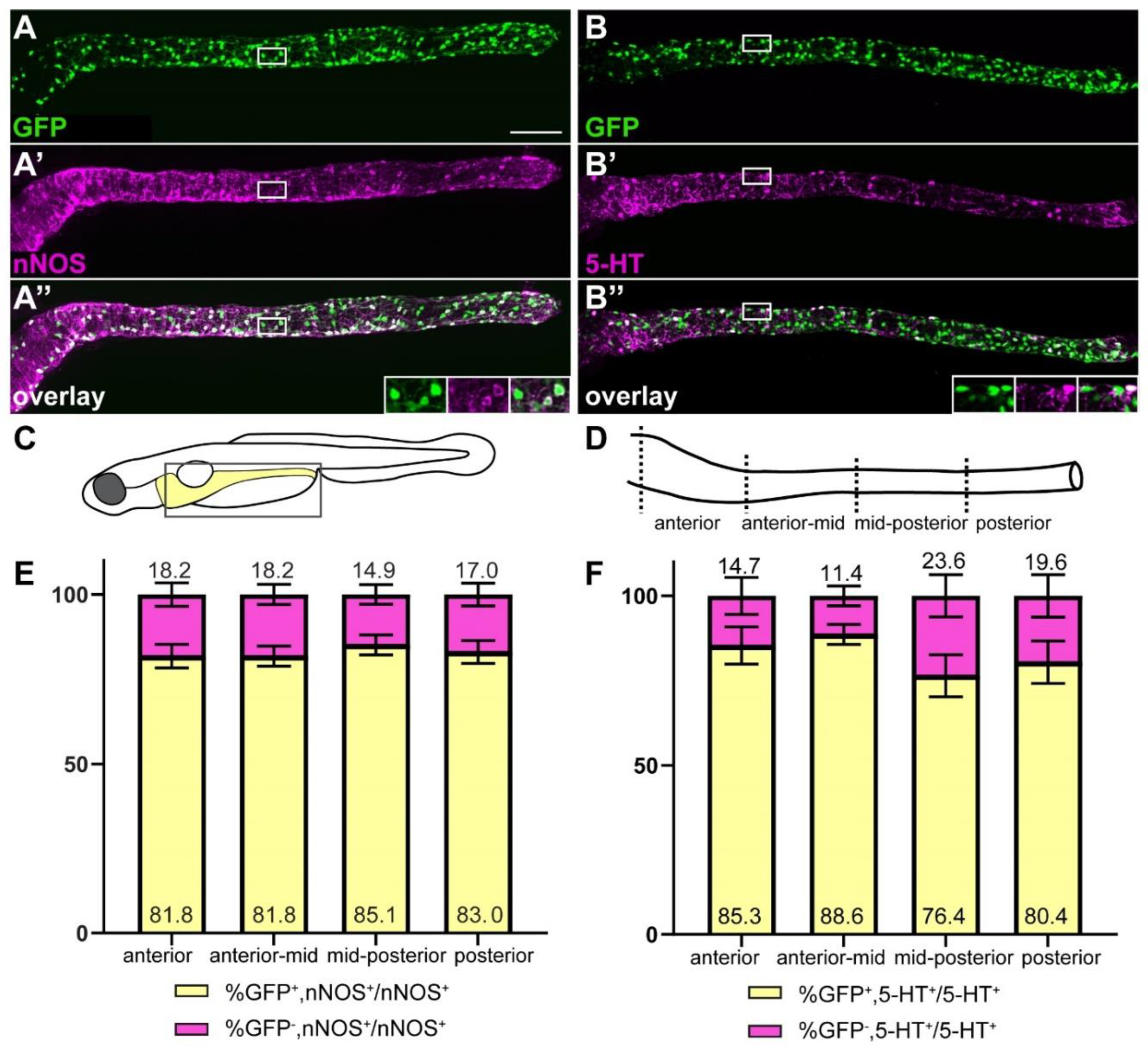
2.4. ret:GFP Is Expressed in Two Prominent Neuronal Subpopulations
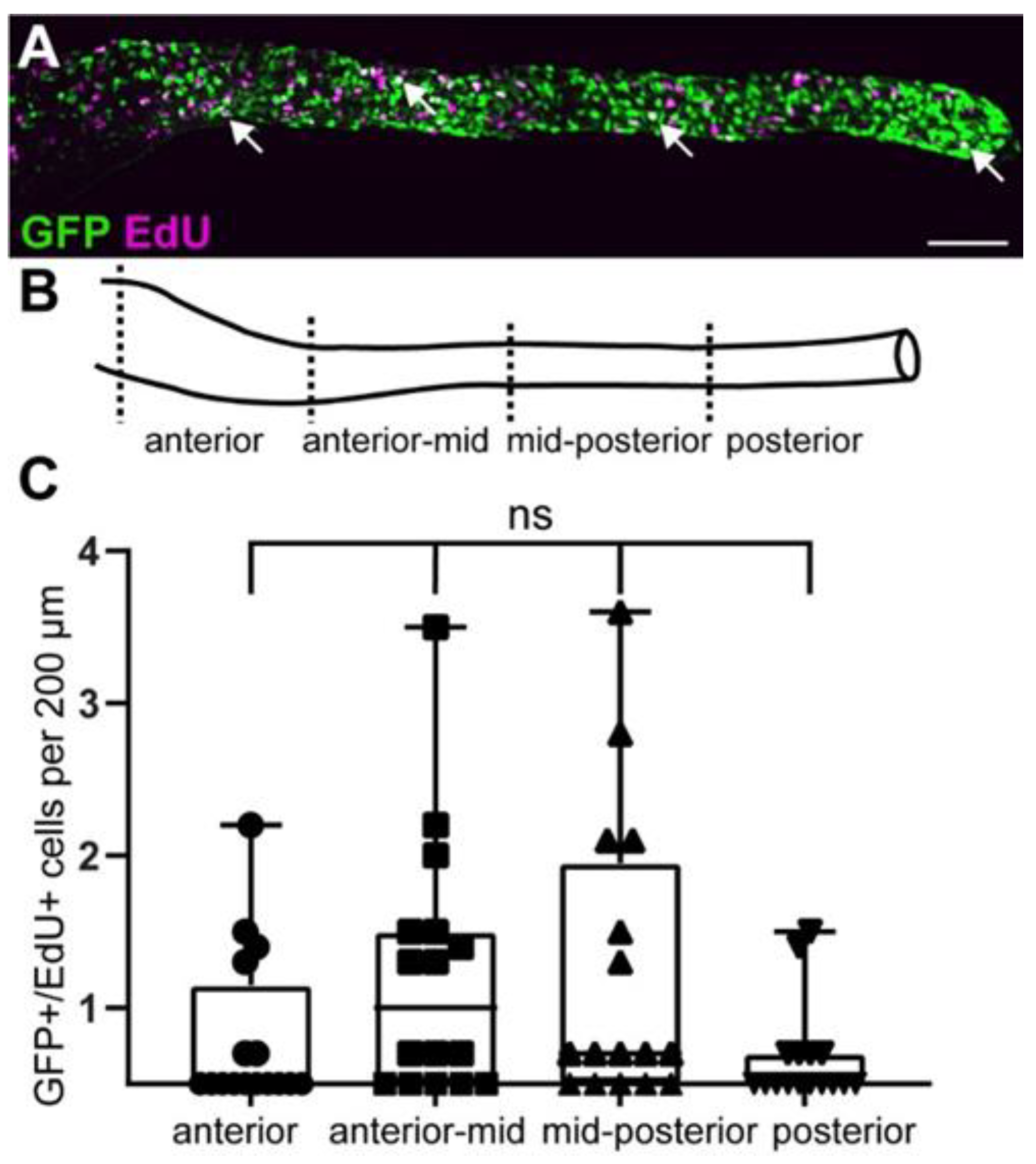
2.5. GFP+ Cells Include Proliferating EPCs
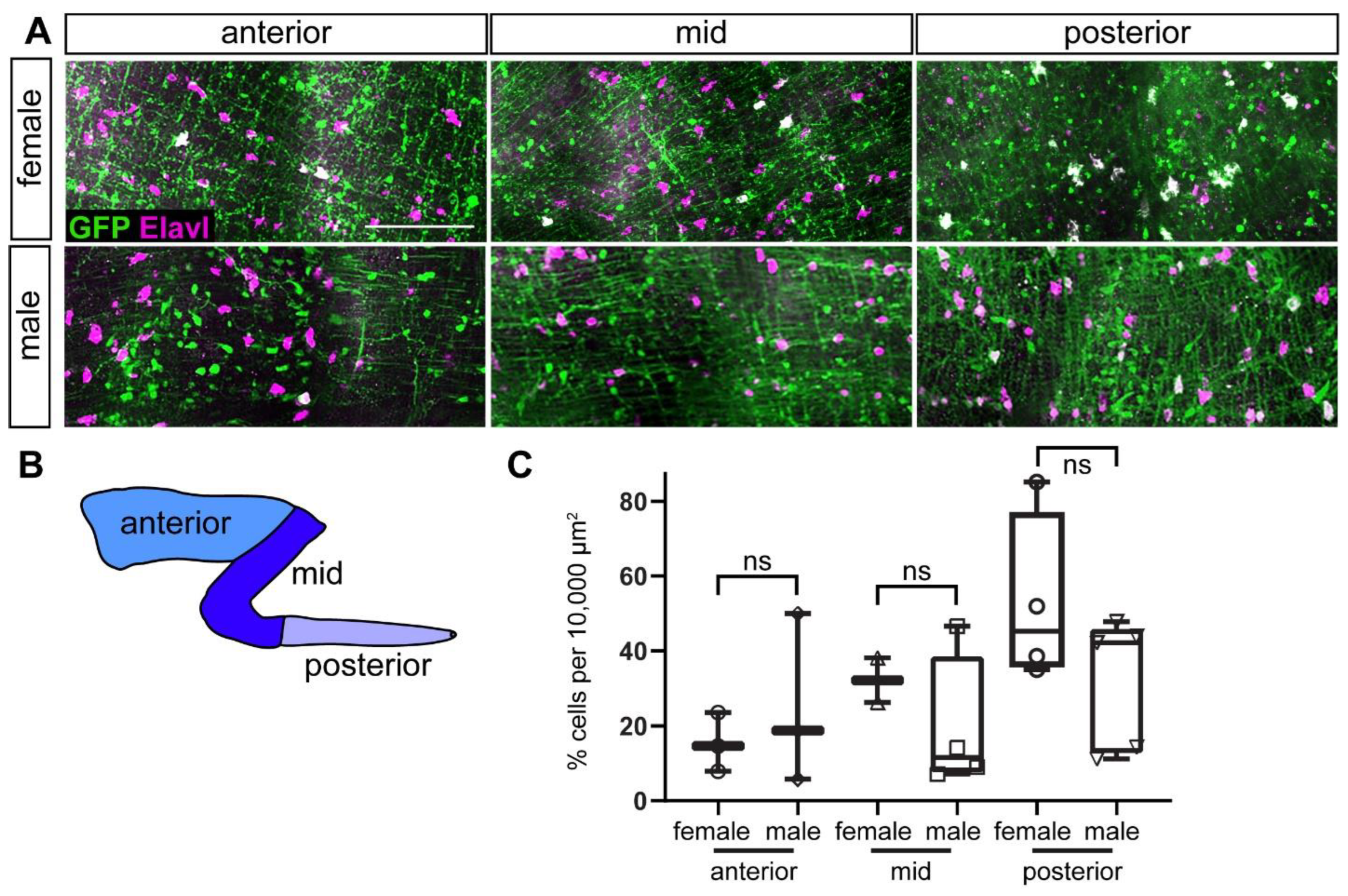
2.6. Adult ENS Neurons Express ret:GFP
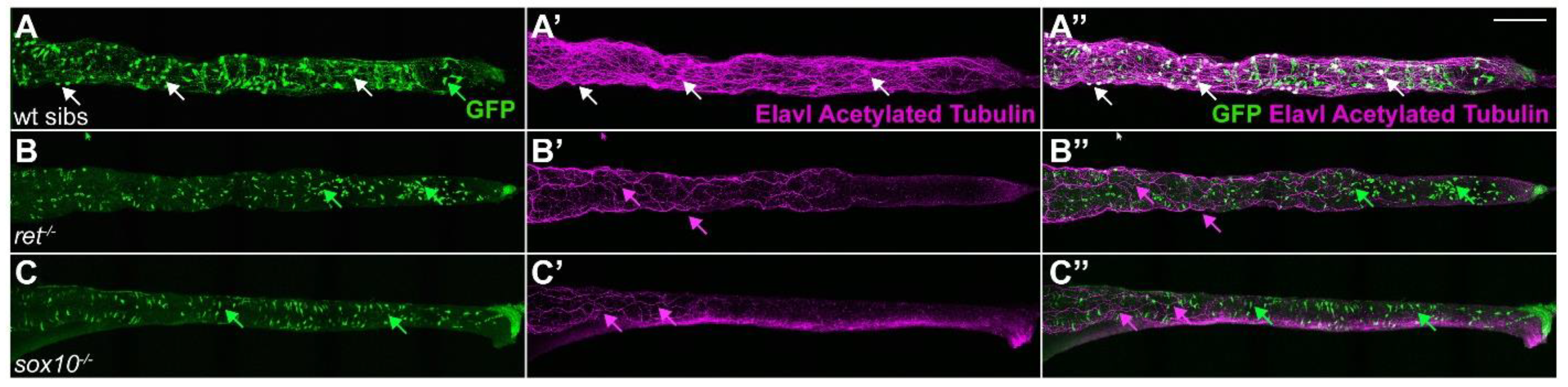
2.7. ret:GFP Positive ENS Cells Are Absent in ret and sox10 Mutants
3. Discussion
3.1. Most Enteric Neurons Are GFP Positive
3.2. Adult ENS Neurons Are GFP Positive
3.3. A Small Proportion of ENS Neurons Is GFP-Negative at Larval and Adult Stages
3.4. Enteric Progenitor Cells Are GFP-Positive at Larval Stages
4. Materials and Methods
4.1. Zebrafish Husbandry and Strains
4.2. Generation of Transgenic Line
4.3. Tissue Preparation
4.4. Immunostaining
4.5. In Situ Hybridization
4.6. EdU Staining
4.7. Image Acquisition
4.8. Image Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Ganz, J. Gut feelings: Studying enteric nervous system development, function, and disease in the zebrafish model system. Dev. Dyn. 2018, 247, 268–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romeo, G.; Ronchetto, P.; Luo, Y.; Barone, V.; Seri, M.; Ceccherini, I.; Pasini, B.; Bocciardi, R.; Lerone, M.; Kaariainen, H.; et al. Point mutations affecting the tyrosine kinase domain of the RET proto-oncogene in Hirschsprung’s disease. Nature 1994, 367, 377–378. [Google Scholar] [CrossRef] [PubMed]
- Edery, P.; Lyonnet, S.; Mulligan, L.M.; Pelet, A.; Dow, E.; Abel, L.; Holder, S.; Nihoul-Fekete, C.; Ponder, B.A.; Munnich, A. Mutations of the RET proto-oncogene in Hirschsprung’s disease. Nature 1994, 367, 378–380. [Google Scholar] [CrossRef]
- Klein, M.D.; Coran, A.G.; Wesley, J.R.; Drongowski, R.A. Hirschsprung’s disease in the newborn. J. Pediatr. Surg. 1984, 19, 370–374. [Google Scholar] [CrossRef]
- Amiel, J.; Lyonnet, S. Hirschsprung disease, associated syndromes, and genetics: A review. J. Med. Genet. 2001, 38, 729–739. [Google Scholar] [CrossRef]
- Tam, P.K.; Garcia-Barcelo, M. Genetic basis of Hirschsprung’s disease. Pediatr. Surg. Int. 2009, 25, 543–558. [Google Scholar] [CrossRef]
- Schuchardt, A.; D’Agati, V.; Larsson-Blomberg, L.; Costantini, F.; Pachnis, V. Defects in the kidney and enteric nervous system of mice lacking the tyrosine kinase receptor Ret. Nature 1994, 367, 380–383. [Google Scholar] [CrossRef]
- Shepherd, I.T.; Pietsch, J.; Elworthy, S.; Kelsh, R.N.; Raible, D.W. Roles for GFRalpha1 receptors in zebrafish enteric nervous system development. Development 2004, 131, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Ganz, J.; Baker, R.P.; Hamilton, M.K.; Melancon, E.; Diba, P.; Eisen, J.S.; Parthasarathy, R. Image velocimetry and spectral analysis enable quantitative characterization of larval zebrafish gut motility. Neurogastroenterol. Motil. 2018, 30, e13351. [Google Scholar] [CrossRef]
- Heanue, T.A.; Boesmans, W.; Bell, D.M.; Kawakami, K.; Vanden Berghe, P.; Pachnis, V. A Novel Zebrafish ret Heterozygous Model of Hirschsprung Disease Identifies a Functional Role for mapk10 as a Modifier of Enteric Nervous System Phenotype Severity. PLoS Genet. 2016, 12, e1006439. [Google Scholar] [CrossRef]
- Shepherd, I.; Eisen, J. Development of the zebrafish enteric nervous system. Methods Cell Biol. 2011, 101, 143–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, N.; Guyer, R.A.; Hotta, R.; Zhang, D.; Newgreen, D.F.; Halasy, V.; Kovacs, T.; Goldstein, A.M. RET overactivation leads to concurrent Hirschsprung disease and intestinal ganglioneuromas. Development 2020, 147, dev190900. [Google Scholar] [CrossRef] [PubMed]
- Durbec, P.L.; Larsson-Blomberg, L.B.; Schuchardt, A.; Costantini, F.; Pachnis, V. Common origin and developmental dependence on c-ret of subsets of enteric and sympathetic neuroblasts. Development 1996, 122, 349–358. [Google Scholar] [CrossRef]
- Taraviras, S.; Marcos-Gutierrez, C.V.; Durbec, P.; Jani, H.; Grigoriou, M.; Sukumaran, M.; Wang, L.C.; Hynes, M.; Raisman, G.; Pachnis, V. Signalling by the RET receptor tyrosine kinase and its role in the development of the mammalian enteric nervous system. Development 1999, 126, 2785–2797. [Google Scholar] [CrossRef]
- Taylor, C.R.; Montagne, W.A.; Eisen, J.S.; Ganz, J. Molecular fingerprinting delineates progenitor populations in the developing zebrafish enteric nervous system. Dev. Dyn. 2016, 245, 1081–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, H.M.; Ciampoli, D.; Hsuan, J.; Canty, A.J. Expression of Ret-, p75(NTR)-, Phox2a-, Phox2b-, and tyrosine hydroxylase-immunoreactivity by undifferentiated neural crest-derived cells and different classes of enteric neurons in the embryonic mouse gut. Dev. Dyn. 1999, 216, 137–152. [Google Scholar] [CrossRef]
- Young, H.M.; Hearn, C.J.; Ciampoli, D.; Southwell, B.R.; Brunet, J.F.; Newgreen, D.F. A single rostrocaudal colonization of the rodent intestine by enteric neuron precursors is revealed by the expression of Phox2b, Ret, and p75 and by explants grown under the kidney capsule or in organ culture. Dev. Biol. 1998, 202, 67–84. [Google Scholar] [CrossRef] [Green Version]
- Avantaggiato, V.; Dathan, N.A.; Grieco, M.; Fabien, N.; Lazzaro, D.; Fusco, A.; Simeone, A.; Santoro, M. Developmental expression of the RET protooncogene. Cell Growth Differ. 1994, 5, 305–311. [Google Scholar]
- Lo, L.; Anderson, D.J. Postmigratory neural crest cells expressing c-RET display restricted developmental and proliferative capacities. Neuron 1995, 15, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Tsuzuki, T.; Takahashi, M.; Asai, N.; Iwashita, T.; Matsuyama, M.; Asai, J. Spatial and temporal expression of the ret proto-oncogene product in embryonic, infant and adult rat tissues. Oncogene 1995, 10, 191–198. [Google Scholar]
- Sunardi, M.; Ito, K.; Enomoto, H. Live visualization of a functional RET-EGFP chimeric receptor in homozygous knock-in mice. Dev. Growth Differ. 2021, 63, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Morarach, K.; Mikhailova, A.; Knoflach, V.; Memic, F.; Kumar, R.; Li, W.; Ernfors, P.; Marklund, U. Diversification of molecularly defined myenteric neuron classes revealed by single cell RNA-sequencing. Nat. Neurosci. 2021, 24, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Lasrado, R.; Boesmans, W.; Kleinjung, J.; Pin, C.; Bell, D.; Bhaw, L.; McCallum, S.; Zong, H.; Luo, L.; Clevers, H.; et al. Lineage-dependent spatial and functional organization of the mammalian enteric nervous system. Science 2017, 356, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Kuil, L.E.; Chauhan, R.K.; Cheng, W.W.; Hofstra, R.M.W.; Alves, M.M. Zebrafish: A Model Organism for Studying Enteric Nervous System Development and Disease. Front. Cell Dev. Biol. 2021, 8, 629073. [Google Scholar] [CrossRef] [PubMed]
- Heanue, T.A.; Pachnis, V. Ret isoform function and marker gene expression in the enteric nervous system is conserved across diverse vertebrate species. Mech. Dev. 2008, 125, 687–699. [Google Scholar] [CrossRef]
- Fisher, S.; Grice, E.A.; Vinton, R.M.; Bessling, S.L.; McCallion, A.S. Conservation of RET regulatory function from human to zebrafish without sequence similarity. Science 2006, 312, 276–279. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, I.T.; Beattie, C.E.; Raible, D.W. Functional analysis of zebrafish GDNF. Dev. Biol. 2001, 231, 420–435. [Google Scholar] [CrossRef] [Green Version]
- Bisgrove, B.W.; Raible, D.W.; Walter, V.; Eisen, J.S.; Grunwald, D.J. Expression of c-ret in the zebrafish embryo: Potential roles in motoneuronal development. J. NeuroBiol. 1997, 33, 749–768. [Google Scholar] [CrossRef]
- Ye, L.; Bae, M.; Cassilly, C.D.; Jabba, S.V.; Thorpe, D.W.; Martin, A.M.; Lu, H.Y.; Wang, J.; Thompson, J.D.; Lickwar, C.R.; et al. Enteroendocrine cells sense bacterial tryptophan catabolites to activate enteric and vagal neuronal pathways. Cell Host Microbe 2021, 29, 179–196.e9. [Google Scholar] [CrossRef]
- Suster, M.L.; Abe, G.; Schouw, A.; Kawakami, K. Transposon-mediated BAC transgenesis in zebrafish. Nat. Protoc. 2011, 6, 1998–2021. [Google Scholar] [CrossRef]
- Snapp, E.L. Fluorescent proteins: A cell biologist’s user guide. Trends Cell Biol. 2009, 19, 649–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olden, T.; Akhtar, T.; Beckman, S.A.; Wallace, K.N. Differentiation of the zebrafish enteric nervous system and intestinal smooth muscle. Genesis 2008, 46, 484–498. [Google Scholar] [CrossRef] [PubMed]
- McCallum, S.; Obata, Y.; Fourli, E.; Boeing, S.; Peddie, C.J.; Xu, Q.; Horswell, S.; Kelsh, R.; Collinson, L.; Wilkinson, D.; et al. Enteric glia as a source of neural progenitors in adult zebrafish. eLife 2020, 9, e56086. [Google Scholar] [CrossRef]
- Roy-Carson, S.; Natukunda, K.; Chou, H.C.; Pal, N.; Farris, C.; Schneider, S.Q.; Kuhlman, J.A. Defining the transcriptomic landscape of the developing enteric nervous system and its cellular environment. BMC Genom. 2017, 18, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, A.G.T.; Baker, P.A.; Ibarra-Garcia-Padilla, R.; Moore, J.A.; Rivas, L.J.; Tallman, J.J.; Singleton, E.W.; Westheimer, J.L.; Corteguera, J.A.; Uribe, R.A. An atlas of neural crest lineages along the posterior developing zebrafish at single-cell resolution. eLife 2021, 10, e60005. [Google Scholar] [CrossRef]
- Chehrehasa, F.; Meedeniya, A.C.; Dwyer, P.; Abrahamsen, G.; Mackay-Sim, A. EdU, a new thymidine analogue for labelling proliferating cells in the nervous system. J. Neurosci. Methods 2009, 177, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelsh, R.N.; Eisen, J.S. The zebrafish colourless gene regulates development of non-ectomesenchymal neural crest derivatives. Development 2000, 127, 515–525. [Google Scholar] [CrossRef]
- Uyttebroek, L.; Shepherd, I.T.; Harrisson, F.; Hubens, G.; Blust, R.; Timmermans, J.P.; Van Nassauw, L. Neurochemical coding of enteric neurons in adult and embryonic zebrafish (Danio rerio). J. Comp. Neurol. 2010, 518, 4419–4438. [Google Scholar] [CrossRef] [Green Version]
- Landman, K.A.; Simpson, M.J.; Newgreen, D.F. Mathematical and experimental insights into the development of the enteric nervous system and Hirschsprung’s disease. Dev. Growth Differ. 2007, 49, 277–286. [Google Scholar] [CrossRef]
- Newgreen, D.F.; Zhang, D.; Cheeseman, B.L.; Binder, B.J.; Landman, K.A. Differential Clonal Expansion in an Invading Cell Population: Clonal Advantage or Dumb Luck? Cells Tissues Organs 2017, 203, 105–113. [Google Scholar] [CrossRef]
- Simpson, M.J.; Zhang, D.C.; Mariani, M.; Landman, K.A.; Newgreen, D.F. Cell proliferation drives neural crest cell invasion of the intestine. Dev. Biol. 2007, 302, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.M.; Schneider, S.; Smith-Edwards, K.M.; Mafra, F.; Leembruggen, A.J.L.; Gonzalez, M.V.; Kothakapa, D.R.; Anderson, J.B.; Maguire, B.A.; Gao, T.; et al. scRNA-Seq Reveals New Enteric Nervous System Roles for GDNF, NRTN, and TBX3. Cell Mol. Gastroenterol. Hepatol. 2021, 11, 1548–1592.e1. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, A.; Feinstein, L.; Sabel, S.; Rastelli, D.; Mezhibovsky, E.; Matthews, L.; Muppirala, A.; Robinson, A.; Gershon, M.D.; Rao, M. RET Signaling Persists in the Adult Intestine and Stimulates Motility by Limiting PYY Release from Enteroendocrine Cells. bioRxiv 2022. [Google Scholar] [CrossRef]
- Barrenschee, M.; Bottner, M.; Hellwig, I.; Harde, J.; Egberts, J.H.; Becker, T.; Wedel, T. Site-specific gene expression and localization of growth factor ligand receptors RET, GFRalpha1 and GFRalpha2 in human adult colon. Cell Tissue Res. 2013, 354, 371–380. [Google Scholar] [CrossRef]
- Rodrigues, D.M.; Li, A.Y.; Nair, D.G.; Blennerhassett, M.G. Glial cell line-derived neurotrophic factor is a key neurotrophin in the postnatal enteric nervous system. Neurogastroenterol. Motil. 2011, 23, e44–e56. [Google Scholar] [CrossRef]
- Bottner, M.; Harde, J.; Barrenschee, M.; Hellwig, I.; Vogel, I.; Ebsen, M.; Wedel, T. GDNF induces synaptic vesicle markers in enteric neurons. Neurosci. Res. 2013, 77, 128–136. [Google Scholar] [CrossRef]
- Wang, H.; Hughes, I.; Planer, W.; Parsadanian, A.; Grider, J.R.; Vohra, B.P.; Keller-Peck, C.; Heuckeroth, R.O. The timing and location of glial cell line-derived neurotrophic factor expression determine enteric nervous system structure and function. J. Neurosci. 2010, 30, 1523–1538. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.P.; Mohammadi, E.; Ligon, C.; Latorre, R.; Johnson, A.C.; Hoang, B.; Krull, D.; Ho, M.W.; Eidam, H.S.; DeMartino, M.P.; et al. Enteric RET inhibition attenuates gastrointestinal secretion and motility via cholinergic signaling in rat colonic mucosal preparations. Neurogastroenterol. Motil. 2019, 31, e13479. [Google Scholar] [CrossRef] [Green Version]
- Luesma, M.J.; Cantarero, I.; Alvarez-Dotu, J.M.; Santander, S.; Junquera, C. New insights into c-Ret signalling pathway in the enteric nervous system and its relationship with ALS. Biomed. Res. Int. 2014, 2014, 328348. [Google Scholar] [CrossRef] [Green Version]
- Elworthy, S.; Pinto, J.P.; Pettifer, A.; Cancela, M.L.; Kelsh, R.N. Phox2b function in the enteric nervous system is conserved in zebrafish and is sox10-dependent. Mech. Dev. 2005, 122, 659–669. [Google Scholar] [CrossRef]
- El-Nachef, W.N.; Bronner, M.E. De novo enteric neurogenesis in post-embryonic zebrafish from Schwann cell precursors rather than resident cell types. Development 2020, 147, dev186619. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Knight, R.D.; Mebus, K.; d’Angelo, A.; Yokoya, K.; Heanue, T.; Tubingen Screen, C.; Roehl, H. Ret signalling integrates a craniofacial muscle module during development. Development 2011, 138, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Parichy, D.M.; Elizondo, M.R.; Mills, M.G.; Gordon, T.N.; Engeszer, R.E. Normal table of postembryonic zebrafish development: Staging by externally visible anatomy of the living fish. Dev. Dyn. 2009, 238, 2975–3015. [Google Scholar] [CrossRef] [Green Version]
- DeLaurier, A.; Eames, B.F.; Blanco-Sanchez, B.; Peng, G.; He, X.; Swartz, M.E.; Ullmann, B.; Westerfield, M.; Kimmel, C.B. Zebrafish sp7:EGFP: A transgenic for studying otic vesicle formation, skeletogenesis, and bone regeneration. Genesis 2010, 48, 505–511. [Google Scholar] [CrossRef]
- Suster, M.L.; Kikuta, H.; Urasaki, A.; Asakawa, K.; Kawakami, K. Transgenesis in zebrafish with the tol2 transposon system. Methods Mol. Biol. 2009, 561, 41–63. [Google Scholar] [CrossRef]
- Suster, M.L.; Sumiyama, K.; Kawakami, K. Transposon-mediated BAC transgenesis in zebrafish and mice. BMC Genom. 2009, 10, 477. [Google Scholar] [CrossRef] [Green Version]
- Gupta, T.; Mullins, M.C. Dissection of organs from the adult zebrafish. J. Vis. Exp. 2010, 37, 1717. [Google Scholar] [CrossRef]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef]
- Quigley, I.K.; Turner, J.M.; Nuckels, R.J.; Manuel, J.L.; Budi, E.H.; MacDonald, E.L.; Parichy, D.M. Pigment pattern evolution by differential deployment of neural crest and post-embryonic melanophore lineages in Danio fishes. Development 2004, 131, 6053–6069. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage of ENS Development | hpf | ENS Cell Type | Method of Detection | Reference | |
|---|---|---|---|---|---|
| EPC migration a | 36 | EPCs c | RNA in situ hybridization | [8,15] | |
| neuronal differentiation b | 54 | EPCs c + neurons d | RNA in situ hybridization | [15] | |
| 60 | EPCs c + neurons d | RNA in situ hybridization | [15] | ||
| 68–70 | neurons e | scRNA-seq | [35] | ||
| 72 | neurons e | RNA in situ hybridization | [28] | ||
| 84 | neurons d | RNA in situ hybridization | [15] | ||
| 168 | neurons e | bulk RNA-seq experiment | [34] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandla, A.; Melancon, E.; Taylor, C.R.; Davidson, A.E.; Eisen, J.S.; Ganz, J. A New Transgenic Tool to Study the Ret Signaling Pathway in the Enteric Nervous System. Int. J. Mol. Sci. 2022, 23, 15667. https://doi.org/10.3390/ijms232415667
Bandla A, Melancon E, Taylor CR, Davidson AE, Eisen JS, Ganz J. A New Transgenic Tool to Study the Ret Signaling Pathway in the Enteric Nervous System. International Journal of Molecular Sciences. 2022; 23(24):15667. https://doi.org/10.3390/ijms232415667
Chicago/Turabian StyleBandla, Ashoka, Ellie Melancon, Charlotte R. Taylor, Ann E. Davidson, Judith S. Eisen, and Julia Ganz. 2022. "A New Transgenic Tool to Study the Ret Signaling Pathway in the Enteric Nervous System" International Journal of Molecular Sciences 23, no. 24: 15667. https://doi.org/10.3390/ijms232415667