
Overexpression of a Cinnamyl Alcohol Dehydrogenase-Coding Gene, GsCAD1, from Wild Soybean Enhances Resistance to Soybean Mosaic Virus
 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
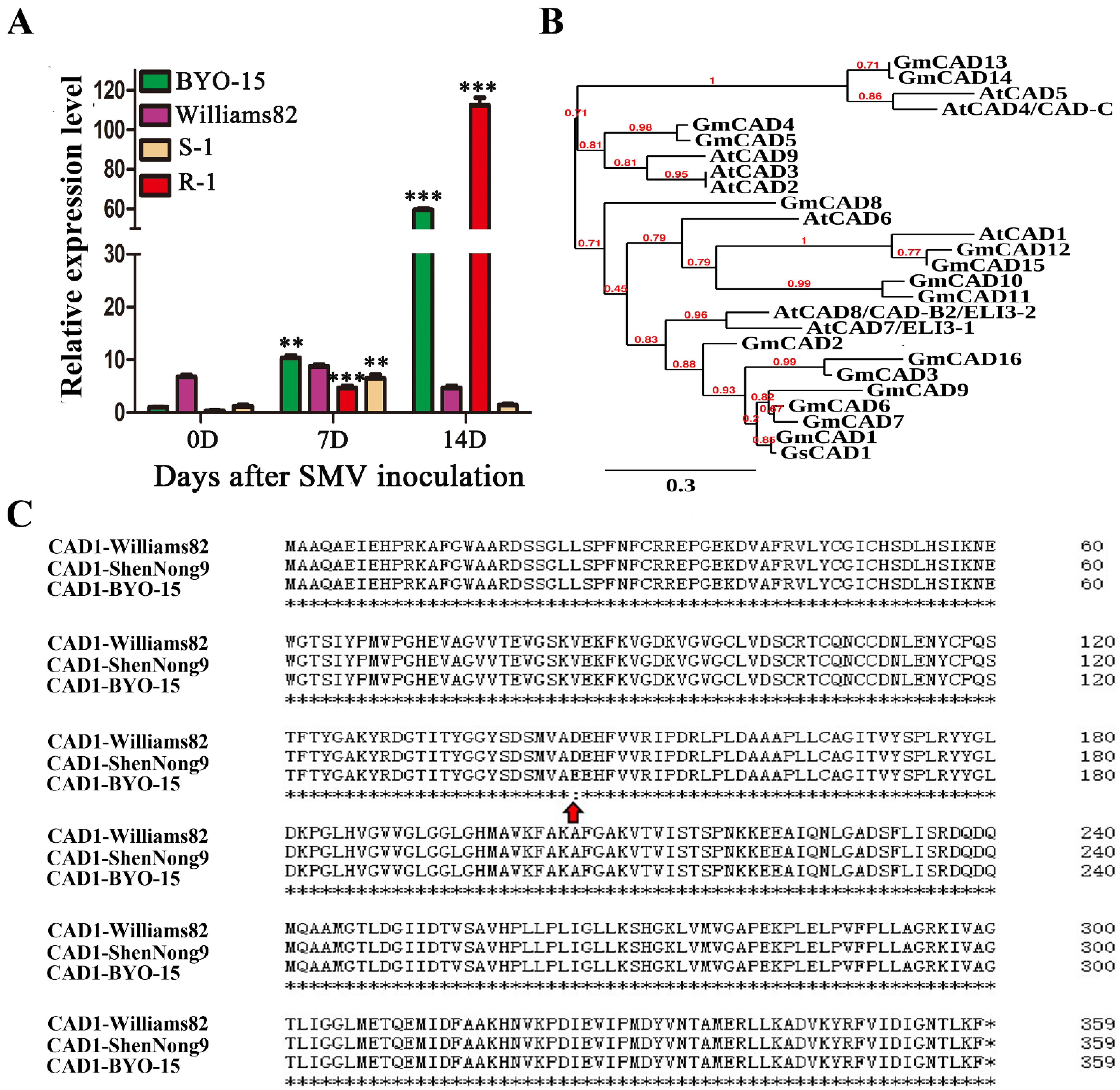
2.1. Identification of GsCAD1 as a Possible anti-SMV Gene
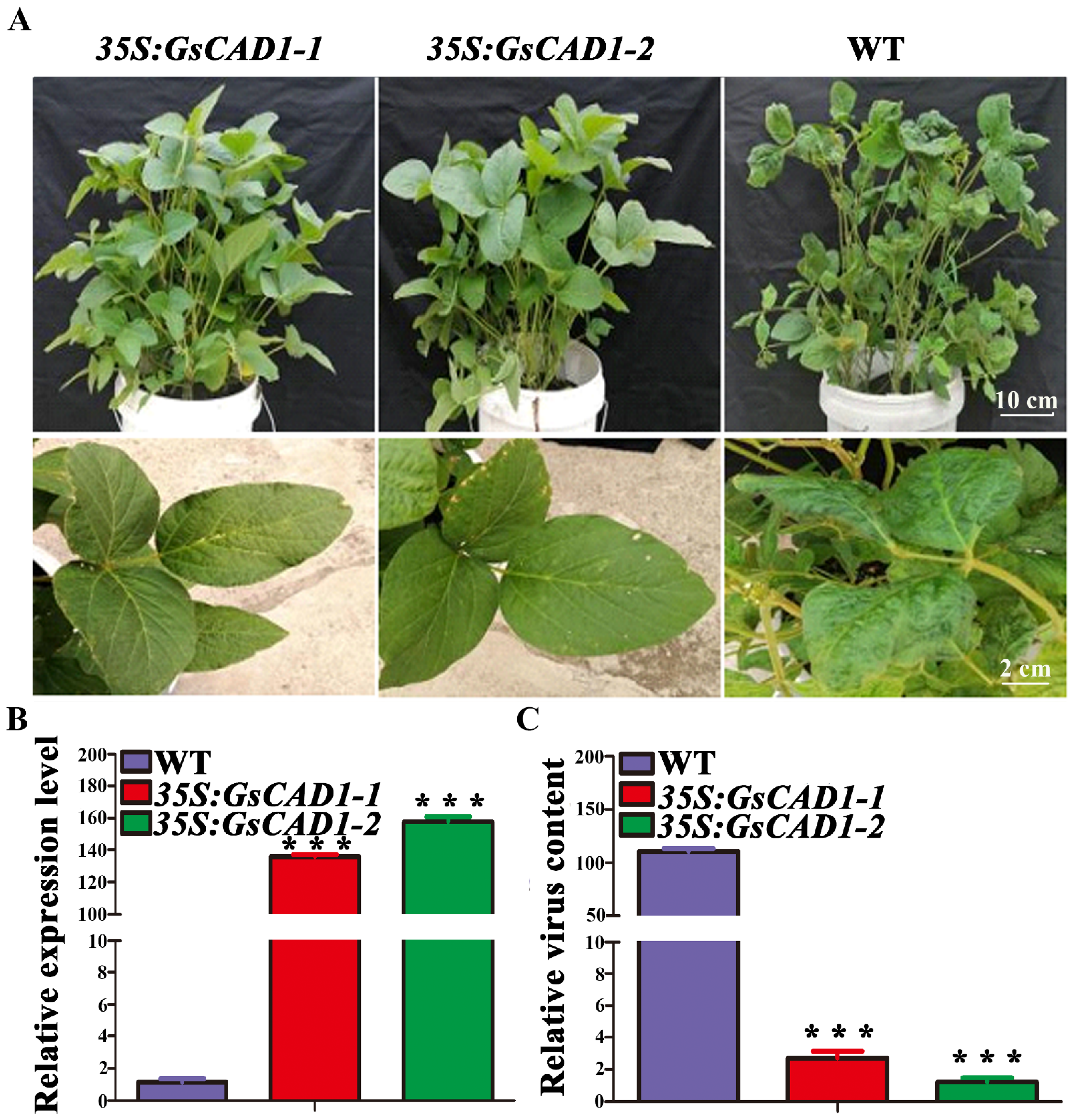
2.2. Overexpression of GsCAD1 in Soybean Increased Resistance to SMV
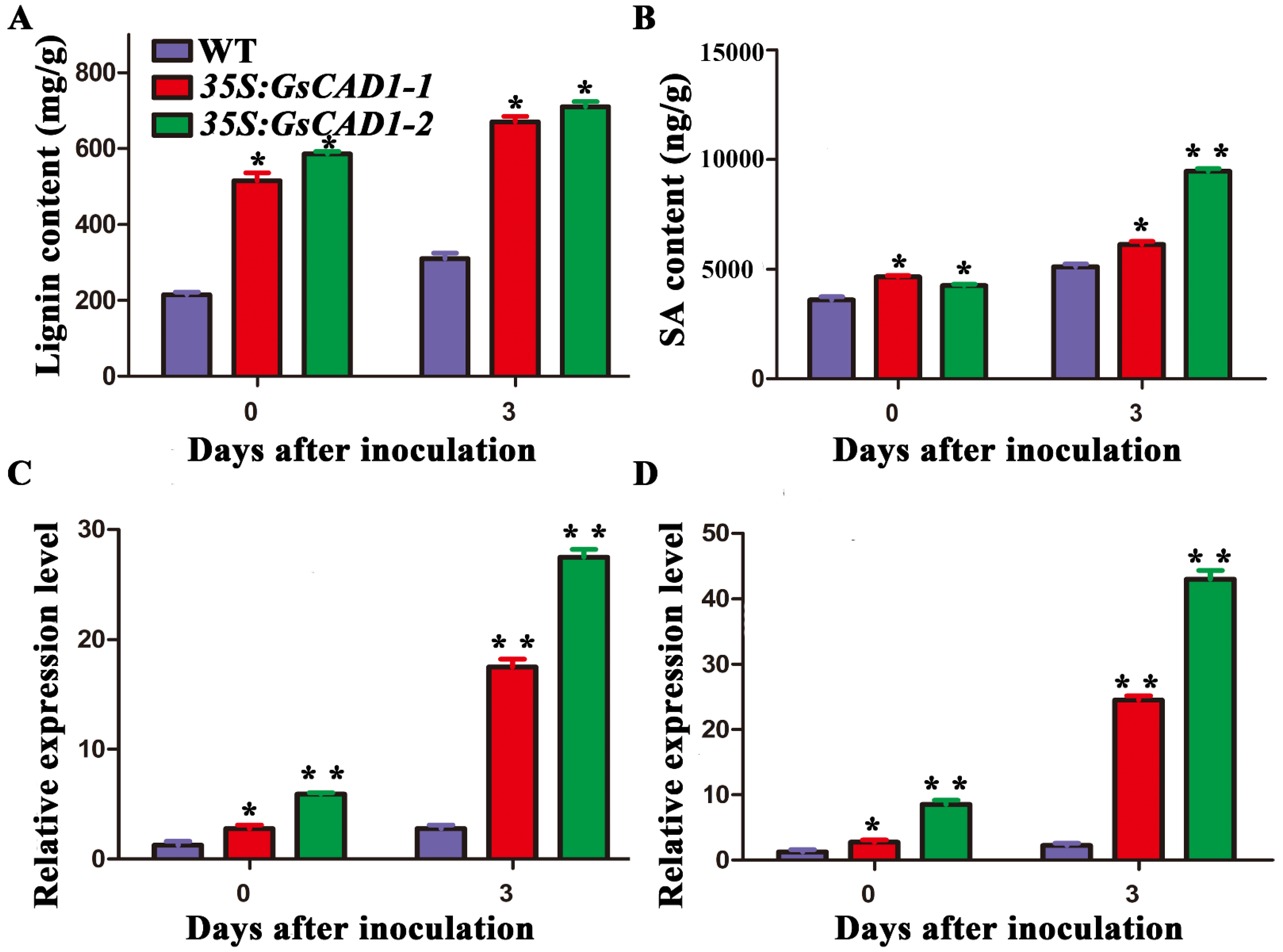
2.3. GsCAD1-Overexpressing Transgenic Soybean Lines Increased the Lignin Content before and after SMV Inoculation
2.4. GsCAD1-Overexpressing Transgenic Soybean Lines Showed Increased Salicylic Acid Contents before and after Inoculation with SMV
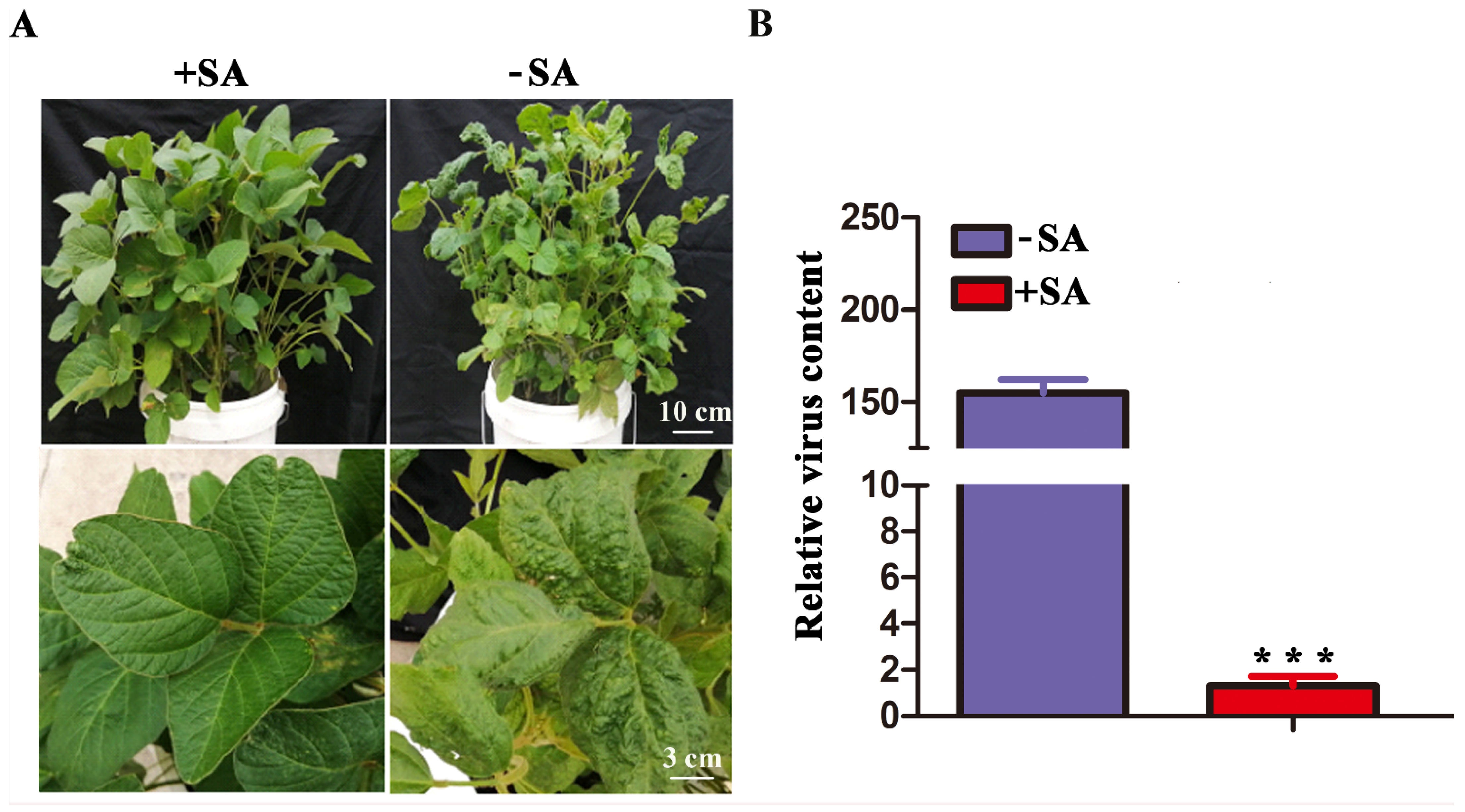
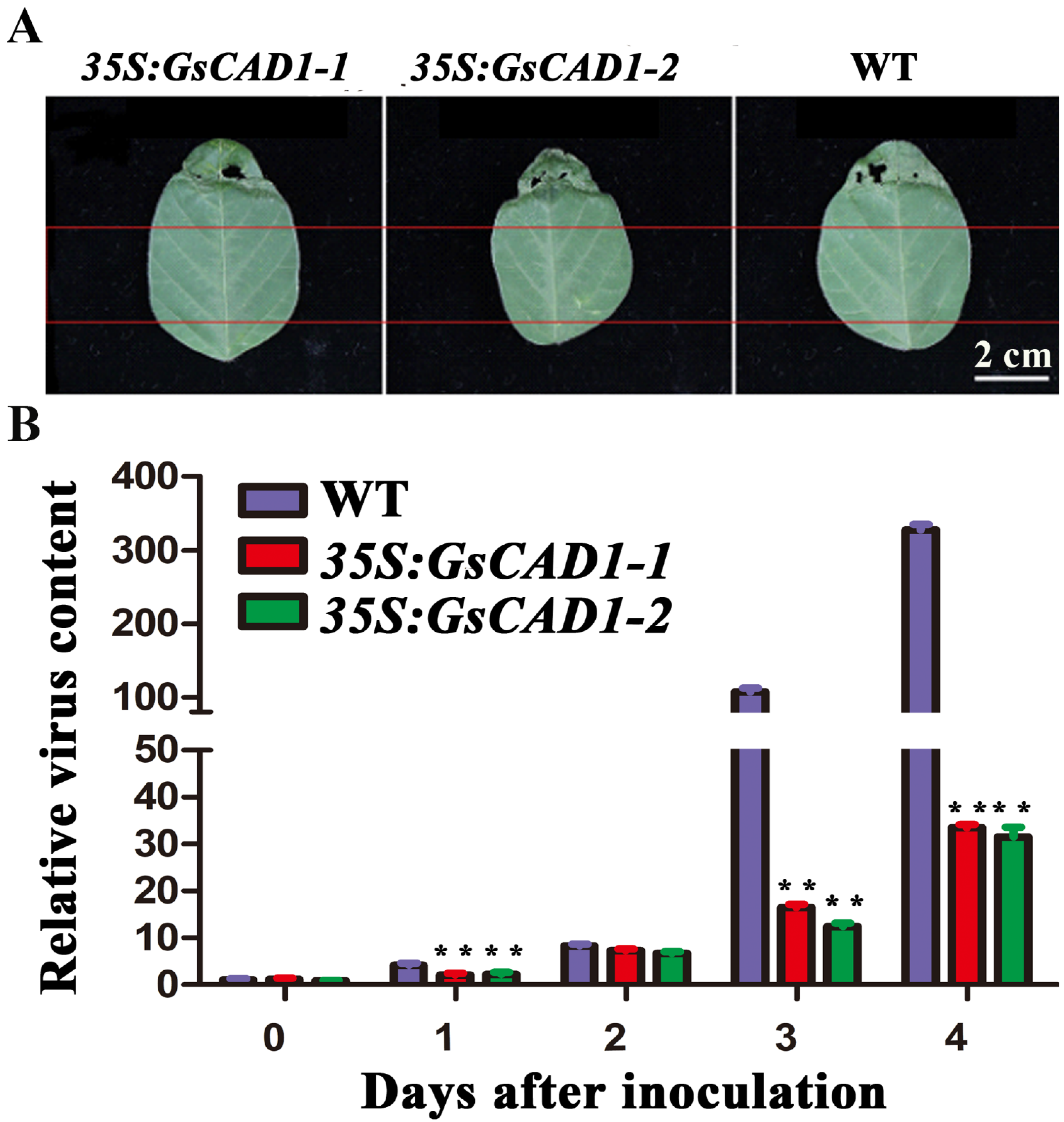
2.5. GsCAD1-Overexpression Affected the Accumulation of SMV in Soybean Leaves
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Virus and SMV Inoculation
4.3. Transcriptome Profiling
4.4. Phylogenetic Analysis of CAD Genes
4.5. Gene Clone and Vector Construction
4.6. Soybean Transformation and Screening of Transgenic Plants
4.7. Quantitative RT-PCR (qRT-PCR)
4.8. SMV Resistance Assay
4.9. Salicylic Acid Treatment
4.10. Measurement of SA and Lignin Contents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the world 2. soybean—Worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Cui, X.; Chen, X.; Wang, A. Detection, Understanding and Control of Soybean Mosaic Virus; InTech: Shanghai, China, 2011; pp. 335–354. [Google Scholar]
- Zheng, C.P.; Chen, P.Y.; Gergerich, R. Effect of temperature on the expression of necrosis in soybean infected with soybean mosaic virus. Crop Sci. 2005, 45, 916. [Google Scholar] [CrossRef]
- Hill, J.H.; Whitham, S.A. Control of virus diseases in soybeans. Adv. Virus Res. 2014, 90, 355–390. [Google Scholar] [PubMed]
- Cho, E.K.; Goodman, R.M. Strains of soybean mosaic virus: Classification based on virulence in resistant soybean cultivars. Phytopathology 1979, 69, 467–470. [Google Scholar] [CrossRef]
- Cho, E.K.; Goodman, R.M. Evaluation of resistance in soybeans to Soybean mosaic virus strains1. Crop Sci. 1982, 22, 1133–1136. [Google Scholar] [CrossRef]
- Li, K.; Yang, Q.H.; Zhi, H.J.; Gai, J.Y. Identification and distribution of soybean mosaic virus strains in southern China. Plant Dis. 2010, 94, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gai, J.; Pu, Z. Classification and distribution of strain groups of soybean mosaic virus in middle and lower Huang-huai and Changjiang valleys. Soybean Sci. 2003, 22, 102–107. [Google Scholar]
- Gore, M.A.; Hayes, A.J.; Jeong, S.C.; Yue, Y.G.; Buss, G.R.; Saghai Maroof, M.A. Mapping tightly linked genes controlling potyvirus infection at the Rsv1 and Rpv1 region in soybean. Genome 2002, 45, 592–599. [Google Scholar] [CrossRef] [Green Version]
- Hwang, T.Y.; Moon, J.K.; Yu, S.; Yang, K.; Mohankumar, S.; Yu, Y.H.; Lee, Y.H.; Kim, H.S.; Kim, H.M.; Saghai Maroof, M.A.; et al. Application of comparative genomics in developing molecular markers tightly linked to the virus resistance gene Rsv4 in soybean. Genome 2006, 49, 380–388. [Google Scholar] [CrossRef]
- Jeong, S.C.; Kristipati, S.; Hayes, A.J.; Maughan, P.J.; Noffsinger, S.L.; Gunduz, I.; Buss, G.R.; Saghai Maroof, M.A. Genetic and sequence analysis of markers tightly linked to the resistance gene, Rsv3. Crop Sci. 2002, 42, 265. [Google Scholar]
- Chen, P.; Ma, G.; Buss, G.R.; Gunduz, I.; Roane, C.W.; Tolin, S.A. Inheritance and allelism tests of Raiden soybean for resistance to soybean mosaic virus. J. Hered. 2001, 92, 51–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunduz, I.; Buss, G.R.; Chen, P.; Tolin, S.A. Characterization of SMV resistance genes in Tousan 140 and Hourei soybean. Crop Sci. 2002, 42, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.J.; Ma, G.; Buss, G.R.; Saghai Maroof, M.A. Molecular marker mapping of Rsv4, a gene conferring resistance to all known strains of soybean mosaic virus. Crop Sci. 2000, 40, 1434. [Google Scholar] [CrossRef]
- Rui, R.; Liu, S.; Karthikeyan, A.; Wang, T.; Niu, H.P.; Yin, J.L.; Yang, Y.H.; Yang, Q.H.; Zhi, H.J.; Li, K. Fine-mapping and identification of a novel locus Rsc15 underlying soybean resistance to Soybean mosaic virus. Theor. Appl. Genet. 2007, 130, 2395–2410. [Google Scholar] [CrossRef]
- Zhou, L.; He, H.L.; Liu, R.F.; Han, Q.; Shou, H.X.; Liu, B. Overexpression of GmAKT2 potassium channel enhances resistance to soybean mosaic virus. BMC Plant Biol. 2014, 14, 154. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.K.; Kwon, S.J.; Cho, W.K.; Choi, H.S.; Kim, K.H. Type 2c protein phosphatase is a key regulator of antiviral extreme resistance limiting virus spread. Sci. Rep. 2014, 4, 5905. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; He, H.L.; Ma, L.P.; Yu, X.Q.; Mi, Q.; Pang, J.S.; Tang, G.X.; Liu, B. Overexpression of a GmCnx1 gene enhanced activity of nitrate reductase and aldehyde oxidase, and boosted mosaic virus resistance in soybean. PLoS ONE 2015, 10, e0124273. [Google Scholar] [CrossRef] [Green Version]
- Luan, H.X.; Shine, M.B.; Cui, X.Y.; Chen, X.; Ma, N.; Kachroo, O.; Zhi, H.J.; Kachroo, A. The potyviral p3 protein targets eEF1a to promote the unfolded protein response and viral pathogenesis. Plant Physiol. 2016, 172, 221–234. [Google Scholar] [CrossRef] [Green Version]
- He, H.L.; Yang, X.D.; Xun, H.W.; Lou, X.; Li, S.Z.; Zhang, Z.B.; Jiang, L.L.; Dong, Y.S.; Wang, S.C.; Liu, B. Over-expression of GmSN1 enhances virus resistance in Arabidopsis and soybean. Plant Cell Rep. 2017, 36, 1441–1455. [Google Scholar] [CrossRef]
- Xun, H.W.; Yang, X.D.; He, H.L.; Wang, M.; Guo, P.; Wang, Y.; Pang, J.S.; Dong, Y.S.; Feng, X.Z.; Wang, S.C.; et al. Over-expression of GmKR3, a TIR-NBS-LRR type R gene, confers resistance to multiple viruses in soybean. Plant Mol. Biol. 2019, 99, 95–111. [Google Scholar] [CrossRef]
- Luan, H.X.; Niu, H.P.; Luo, J.Y.; Zhi, H.J. Soybean cytochrome b5 Is a restriction factor for soybean mosaic virus. Viruses 2019, 11, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.P.; Du, H.Y.; Wang, J.; Pu, Y.X.; Yang, C.Y.; Yan, R.J.; Yang, H.; Cheng, H.; Yu, D.Y. Multiplex CRISPR/Cas9-mediated metabolic engineering increases soya bean isoflavone content and resistance to soya bean mosaic virus. Plant Biotechnol. J. 2021, 18, 1384–1395. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yan, J.; Luo, Z.H.; Gao, S.N.; Teng, W.L.; Zhan, Y.H.; Qiu, L.J.; Zheng, H.K.; Li, W.B.; Han, Y.P. GmST1, which encodes a sulfotransferase, confers resistance to soybean mosaic virus strains G2 and G3. Plant Cell Environ. 2021, 44, 2777–2792. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.L.; Wang, L.Q.; Jin, T.T.; Nie, Y.; Liu, H.; Qiu, Y.L.; Yang, Y.H.; Li, B.W.; Zhang, J.J.; Wang, D.G.; et al. A cell wall-localized NLR confers resistance to soybean mosaic virus by recognizing viral-encoded cylindrical inclusion protein. Mol. Plant 2021, 14, 1881–1900. [Google Scholar] [CrossRef]
- Widyasari, K.; Tran, P.; Shin, J.; Son, H.; Kim, K.H. Overexpression of purple acid phosphatase GmPAP2.1 confers resistance to Soybean mosaic virus in a susceptible soybean cultivar. J. Exp. Bot. 2021, 73, 1623–1642. [Google Scholar] [CrossRef]
- Ren, Q.Y.; Jiang, H.; Xiang, W.Y.; Nie, Y.; Xue, S.; Zhi, H.J.; Li, K.; Gai, J. A MADS-box gene is involved in soybean resistance to multiple soybean mosaic virus strains. Crop J. 2022, 10, 802–808. [Google Scholar] [CrossRef]
- Lange, B.M.; Lapierre, C.; Sandermann, H.J. Elicitor-induced spruce stress lignin (structural similarity to early developmental lignins). Plant Physiol. 1995, 108, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.H. Regulation of lignification in defense. In Genes Involved in Plant Defense. Plant Gene Research; Boller, T., Meins, F., Eds.; Springer: Vienna, Austria, 1992; pp. 327–352. [Google Scholar]
- Cheong, Y.H.; Chang, H.S.; Gupta, R.; Wang, X.; Zhu, T.; Luan, S. Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in Arabidopsis. Plant Physiol. 2002, 129, 661–677. [Google Scholar] [CrossRef] [Green Version]
- Schenk, P.M.; Kazan, K.; Anderson, J.P.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef] [Green Version]
- Tronchet, M.; Balague, C.; Kroj, T.; Jouanin, L.; Roby, D. Cinnamyl alcohol dehydrogenases-C and D, key enzymes in lignin biosynthesis, play an essential role in disease resistance in Arabidopsis. Mol. Plant Pathol. 2010, 11, 83–92. [Google Scholar] [CrossRef]
- Sibou, R.; Eudes, A.; Mouille, G.; Pollet, B.; Lapierre, C.; Jouanin, L.; Seguin, A. Cinnamyl alcohol dehydrogenase-C and -D are the primary genes involved in lignin biosynthesis in the floral stem of Arabidopsis. Plant Cell 2005, 17, 2059–2076. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.M.; Ran, J.H.; Wang, X.Q. Evolution of the cinnamyl/sinapyl alcohol dehydrogenase (CAD/SAD) gene family: The emergence of real lignin is associated with the origin of Bona Fide CAD. J. Mol. Evol. 2010, 71, 202–218. [Google Scholar] [CrossRef]
- Kutsuki, H.; Shimada, M.; Higuchi, T. Regulatory role of cinnamyl alcohol dehydrogenase in the formation of guaiacyl and syringyl lignins. Phytochemistry 1982, 21, 19–23. [Google Scholar] [CrossRef]
- Proietti, S.; Bertini, L.; Falconieri, G.S.; Baccelli, I.; Timperio, A.M.; Caruso, C. A metabolic profiling analysis revealed a primary metabolism reprogramming in Arabidopsis glyI4 loss-of-function mutant. Plants 2021, 10, 2464. [Google Scholar] [CrossRef]
- Wei, R.; Luo, M.Y.; Shan, T.L.; Wei, X.N.; Du, L.P.; Xu, H.J.; Zhang, Z.Y. A wheat cinnamyl alcohol dehydrogenase TaCAD12 contributes to host resistance to the sharp eyespot disease. Front. Plant Sci. 2016, 7, 1723. [Google Scholar]
- Park, H.L.; Kim, T.L.; Bhoo, S.H.; Lee, T.H.; Lee, S.W.; Cho, M.H. Biochemical characterization of the rice cinnamyl alcohol dehydrogenase gene family. Molecules 2018, 23, 2659. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Lin, B.; Wang, H.; Li, X.M.; Yang, F.F.; Ding, X.H.; Chu, Z.H. Natural variation in ZmFBL41 confers banded leaf and sheath blight resistance in maize. Nat. Genet. 2019, 51, 1540–1548. [Google Scholar] [CrossRef]
- Xie, M.; Chung, C.Y.; Li, M.W.; Wong, F.L.; Wang, X.; Liu, A.L.; Wang, Z.L.; Leung, A.K.; Wong, T.H.; Tong, S.W.; et al. A reference-grade wild soybean genome. Nat. Commun. 2019, 10, 1216. [Google Scholar] [CrossRef] [Green Version]
- Xian, P.Q.; Cai, Z.D.; Cheng, Y.B.; Lin, R.B.; Lian, T.X.; Ma, Q.B.; Nian, H. Wild Soybean Oxalyl-CoA synthetase degrades oxalate and affects the tolerance to cadmium and aluminum stresses. Int. J. Mol. Sci. 2020, 21, 8869. [Google Scholar] [CrossRef]
- Jin, T.; Sun, Y.Y.; Shan, Z.; He, J.B.; Wang, N.; Gai, J.Y.; Li, Y. Natural variation in the promoter of GsERD15B affects salt tolerance in soybean. Plant Biotechnol. J. 2021, 19, 1155–1169. [Google Scholar] [CrossRef]
- Sun, X.L.; Cai, X.X.; Yin, K.D.; Gu, L.W.; Shen, Y.; Hu, B.S.; Wang, Y.; Chen, Y.; Zhu, Y.M.; Jia, B.W.; et al. Wild soybean SNARE proteins BET1s mediate the subcellular localization of the cytoplasmic receptor-like kinases CRCK1s to modulate salt stress responses. Plant J. 2021, 105, 771–785. [Google Scholar] [CrossRef]
- Song, S.; Wang, J.; Yang, X.; Zhang, X.; Xin, X.; Liu, C.; Zou, J.; Cheng, X.; Zhang, N.; Hu, Y.; et al. GsRSS3L, a candidate gene underlying soybean resistance to seedcoat mottling derived from wild soybean (Glycine soja Sieb. and Zucc). Int. J. Mol. Sci. 2022, 23, 7577. [Google Scholar] [CrossRef]
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef]
- Yuan, W.; Jiang, T.; Du, K.T.; Chen, H.; Cao, Y.Y.; Xie, J.P.; Li, M.F.; Carr, P.J.; Wu, B.M.; Fan, Z.F.; et al. Maize phenylalanine ammonia-lyases contribute to resistance to sugarcane mosaic virus infection, most likely through positive regulation of salicylic acid accumulation. Mol. Plant Pathol. 2019, 20, 1365–1378. [Google Scholar] [CrossRef] [Green Version]
- Hajimorad, M.R.; Domier, L.L.; Tolin, S.A.; Whitham, S.A.; Saghai Maroof, M.A. Soybean mosaic virus: A successful potyvirus with a wide distribution but restricted natural host range. Mol. Plant Pathol. 2018, 19, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Li, B.S.; Zheng, X.Y.; Li, J.G.; Yang, M.; Dong, X.N.; He, G.M.; An, C.C.; Deng, X.W. Salicylic acid biosynthesis is enhanced and contributes to increased biotrophic pathogen resistance in Arabidopsis hybrids. Nat. Commun. 2015, 6, 7309. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Z.; Fang, Y.; Pang, H.X. The current status of the soybean-soybean mosaic virus (SMV) pathosystem. Front. Microbiol. 2016, 7, 1906. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Zhang, L.; Huang, Z.P.; Wang, D.G.; Hu, G.Y. Transcription sequencing and its application on discovering the gene resources of wild soybean. Soybean Sci. 2013, 121, 25–30. [Google Scholar]
- Shine, M.B.; Yang, J.W.; EI-Habbak, M.; Nagyabhyru, P.; Fu, D.Q.; Navarre, D.; Ghabrial, S.; Kachroo, P.; Kachroo, A. Cooperative functioning between phenylalanine ammonia lyase and isochorismate synthase activities contributes to salicylic acid biosynthesis in soybean. New Phytol. 2016, 212, 627–636. [Google Scholar] [CrossRef] [Green Version]
- Young, N.D.; Tanksley, S.D. RFLP analysis of the size of chromosomal segments retained around the Tm-2 locus of tomato during backcross breeding. Theor. Appl. Genet. 1989, 77, 353–359. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zhao, Z.Q.; Yang, C.Y.; Zhang, M.C.; Yan, L.; Qiu, L.J.; Liu, C.J. Recombination frequencies between cultivated soybean (Glycine max) and its wild relative Glycine soja based on molecular marker analysis. Afr. J. Biotechnol. 2013, 12, 3522–3527. [Google Scholar]
- Ding, P.T.; Ding, Y.L. Stories of salicylic acid: A plant defense hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Abdelkefi, H.; Sugliani, M.; Ke, H.; Harchouni, S.; Soubigou-Taconnat, L.; Citerne, S.; Mouille, G.; Fakhfakh, H.; Robaglia, C.; Field, B. Guanosine tetraphosphate modulates salicylic acid signalling and the resistance of Arabidopsis thaliana to Turnip mosaic virus. Mol. Plant Pathol. 2017, 11, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Bi, M.M.; Li, X.Y.; Yan, X.; Liu, D.; Gao, G.; Zhu, P.F.; Mao, H.Y. Chrysanthemum WRKY15-1 promotes resistance to Puccinia horiana Henn via the salicylic acid signaling pathway. Hortic. Res. 2021, 8, 6. [Google Scholar] [CrossRef]
- Venturuzzi, A.L.; Rodriguez, M.C.; Conti, G.; Leone, M.; Caro, M.D.P.; Montecchia, J.F.; Zavallo, D.; Asurmendi, S. Negative modulation of SA signaling components by the capsid protein of tobacco mosaic virus is required for viral long-distance movement. Plant J. 2021, 106, 896–912. [Google Scholar] [CrossRef]
- Mao, C.Y.; Shan, S.Q.; Huang, Y.; Jiang, C.; Zhang, H.H.; Li, Y.J.; Chen, J.P.; Wei, Z.Y.; Sun, Z.T. The hypervariable N-terminal of soybean mosaic virus P1 protein influences its pathogenicity and host defense responses. Phytopathol. Res. 2022, 4, 10. [Google Scholar] [CrossRef]
- Jan, R.; Khan, M.A.; Asaf, S.; Lubna; Lee, I.-J.; Kim, K.-M. Over-expression of chorismate mutase enhances the accumulation of salicylic acid, lignin, and antioxidants in response to the white-backed planthopper in rice plants. Antioxidants 2021, 10, 1680. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one fastq preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Sudhir, K.; Glen, S.; Koichiro, T. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Paz, M.M.; Shou, H.; Guo, Z.; Zhang, X.; Banerjee, A.K.; Wang, K. Assessment of conditions affecting agrobacterium-mediated soybean transformation using the cotyledonary node explant. Euphytica 2014, 136, 167–179. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xun, H.; Qian, X.; Wang, M.; Yu, J.; Zhang, X.; Pang, J.; Wang, S.; Jiang, L.; Dong, Y.; Liu, B. Overexpression of a Cinnamyl Alcohol Dehydrogenase-Coding Gene, GsCAD1, from Wild Soybean Enhances Resistance to Soybean Mosaic Virus. Int. J. Mol. Sci. 2022, 23, 15206. https://doi.org/10.3390/ijms232315206
Xun H, Qian X, Wang M, Yu J, Zhang X, Pang J, Wang S, Jiang L, Dong Y, Liu B. Overexpression of a Cinnamyl Alcohol Dehydrogenase-Coding Gene, GsCAD1, from Wild Soybean Enhances Resistance to Soybean Mosaic Virus. International Journal of Molecular Sciences. 2022; 23(23):15206. https://doi.org/10.3390/ijms232315206
Chicago/Turabian StyleXun, Hongwei, Xueyan Qian, Meng Wang, Jiaxin Yu, Xue Zhang, Jinsong Pang, Shucai Wang, Lili Jiang, Yingshan Dong, and Bao Liu. 2022. "Overexpression of a Cinnamyl Alcohol Dehydrogenase-Coding Gene, GsCAD1, from Wild Soybean Enhances Resistance to Soybean Mosaic Virus" International Journal of Molecular Sciences 23, no. 23: 15206. https://doi.org/10.3390/ijms232315206