
Influence of Sex and Strain on Hepatic and Adipose Tissue Trace Element Concentrations and Gene Expression in C57BL/6J and DBA/2J High Fat Diet Models

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
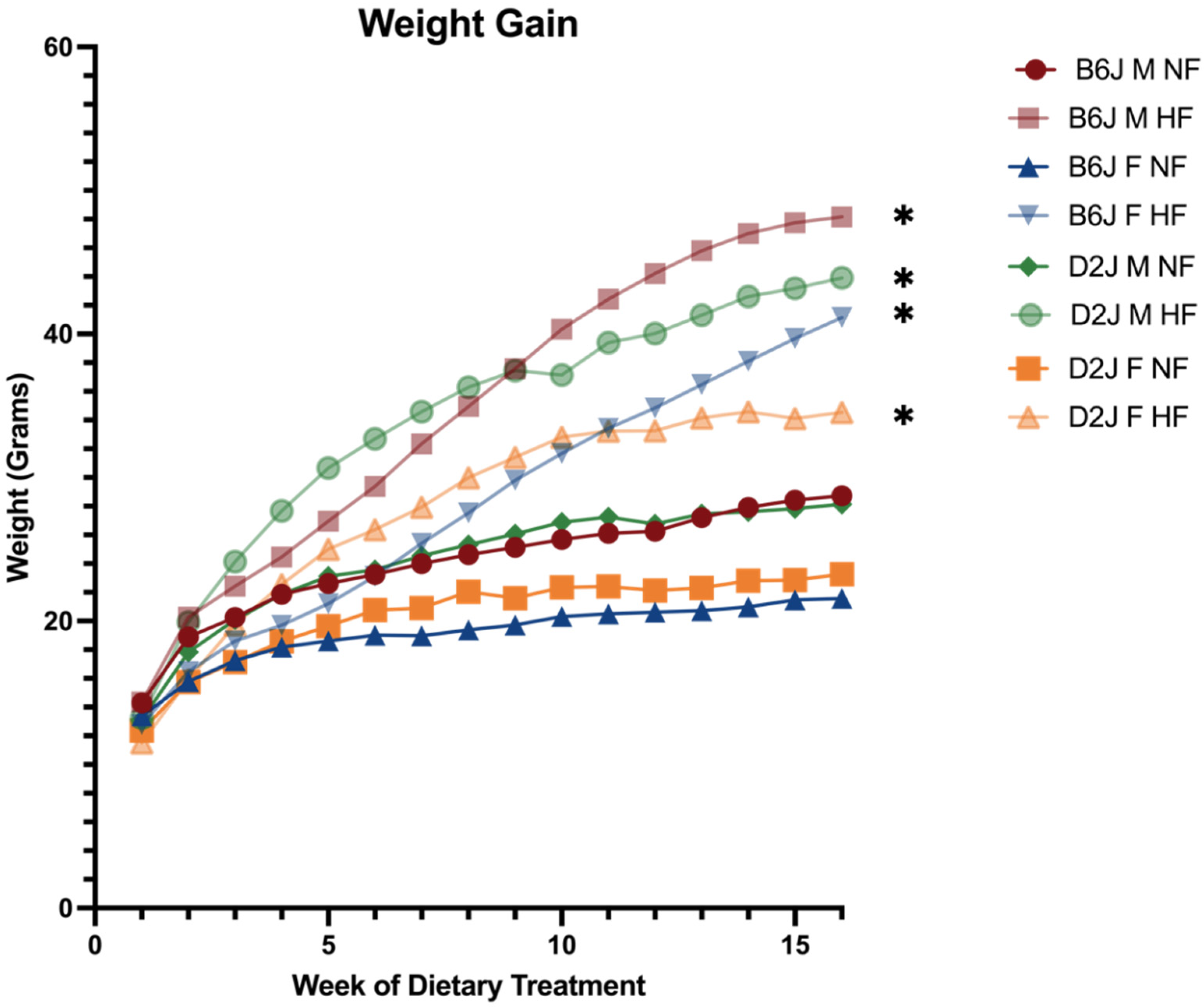
2.1. Weight Gain and Feed Efficiency
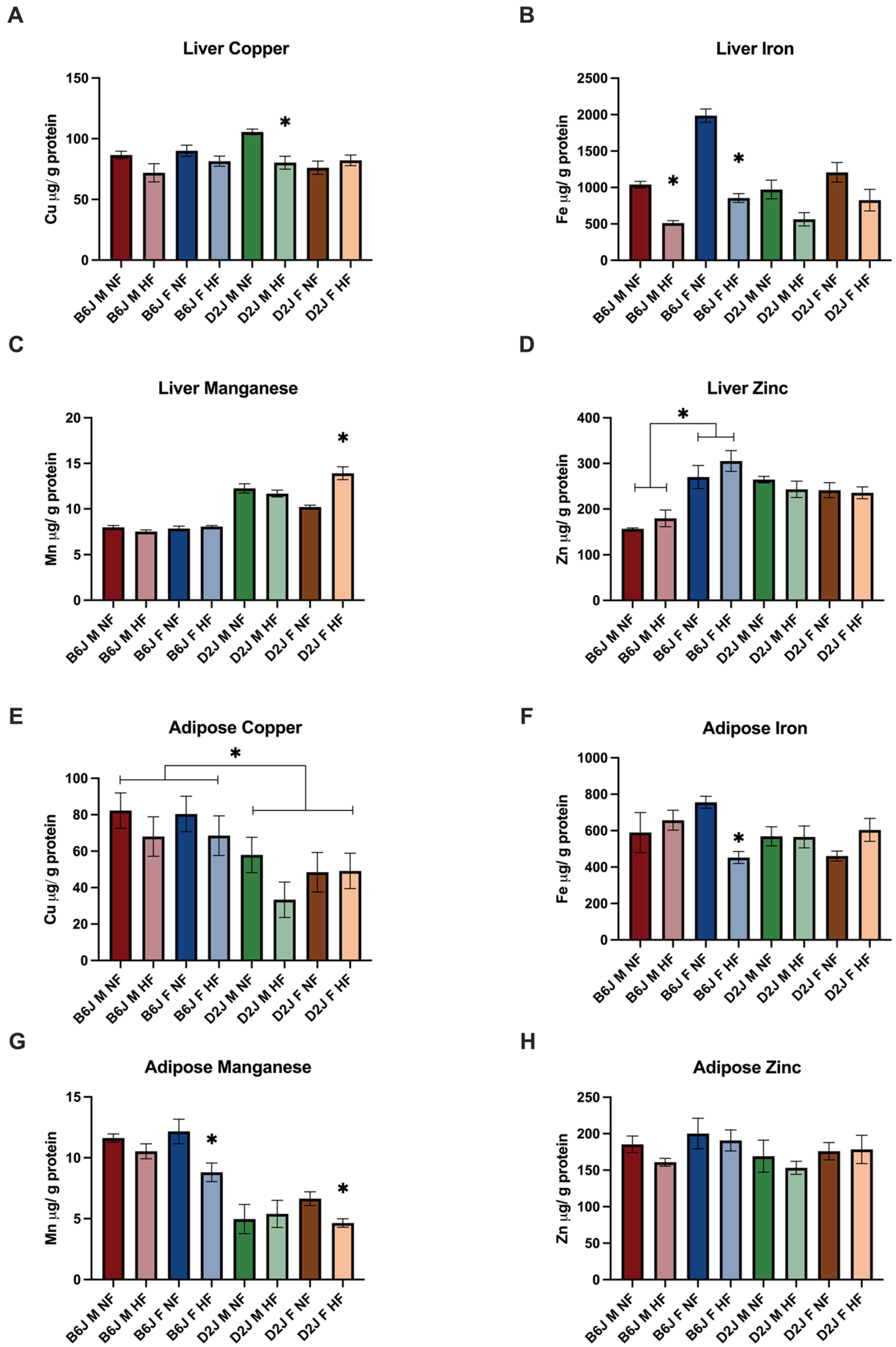
2.2. Effect of Diet, Sex, and Strain on Trace Element Concentration
2.2.1. Liver
2.2.2. Adipose Tissue
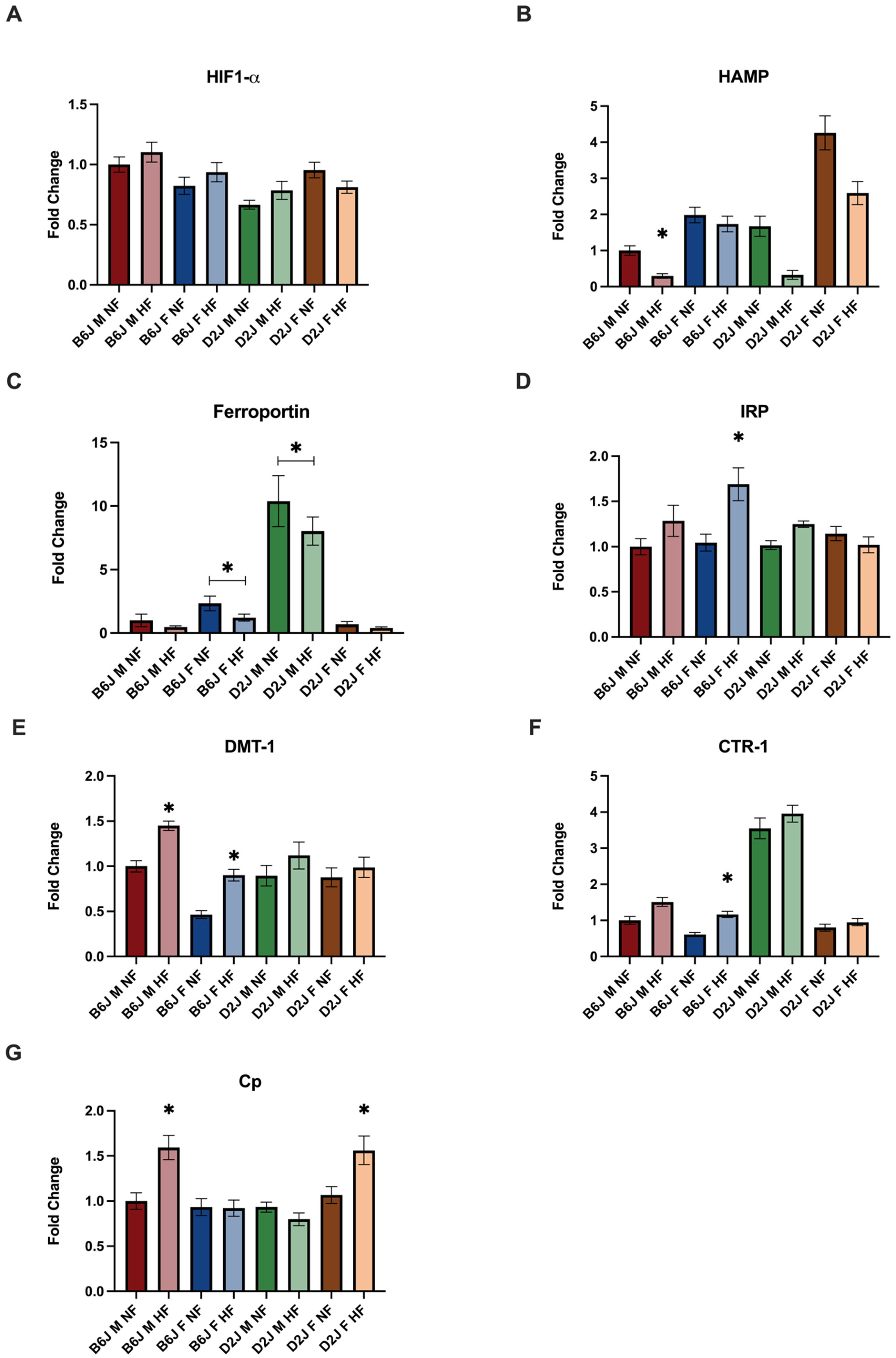
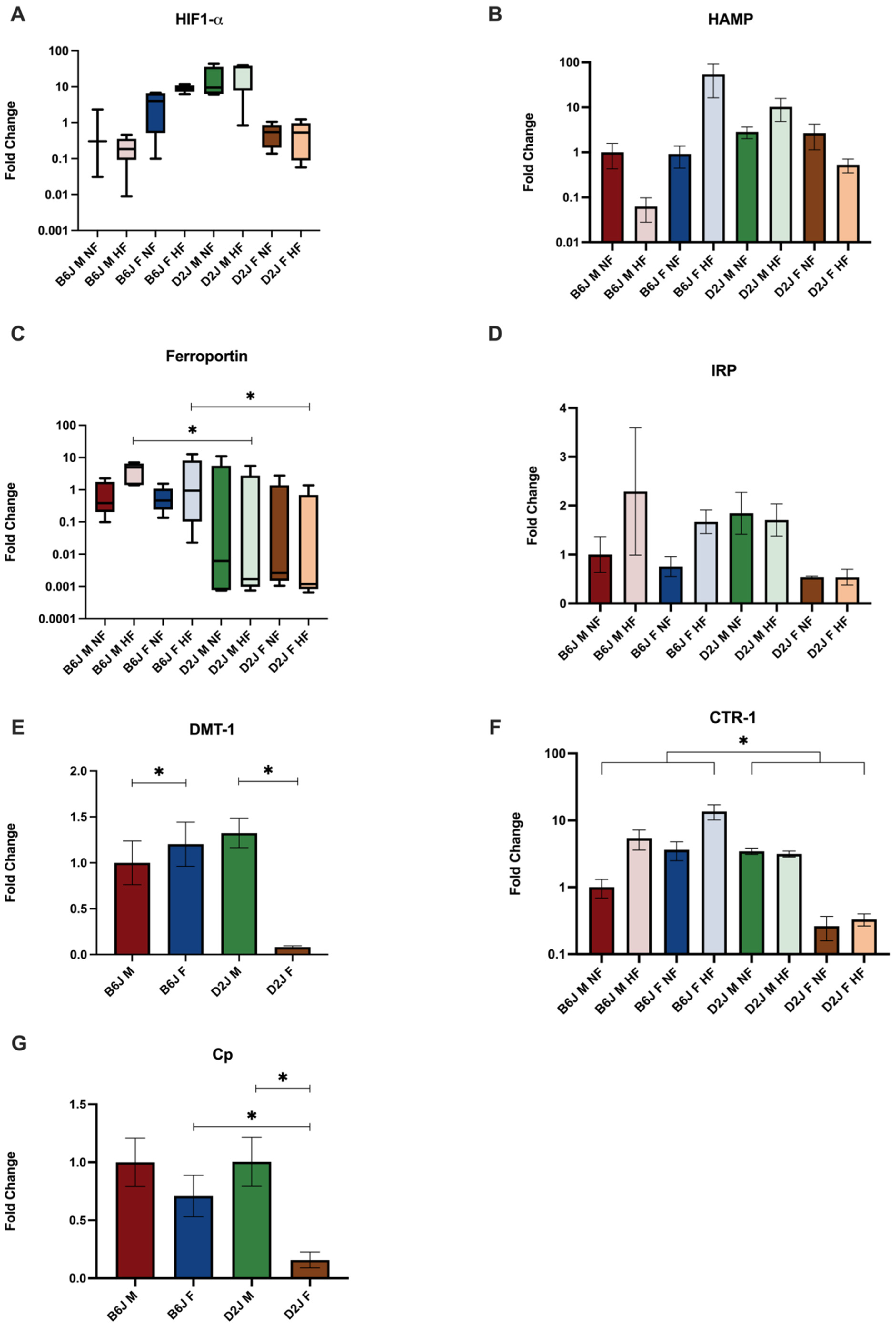
2.3. Effect of Diet, Sex, and Strain on the Gene Expression of Trace Element Homeostatic Proteins
2.3.1. Liver
2.3.2. Adipose Tissue
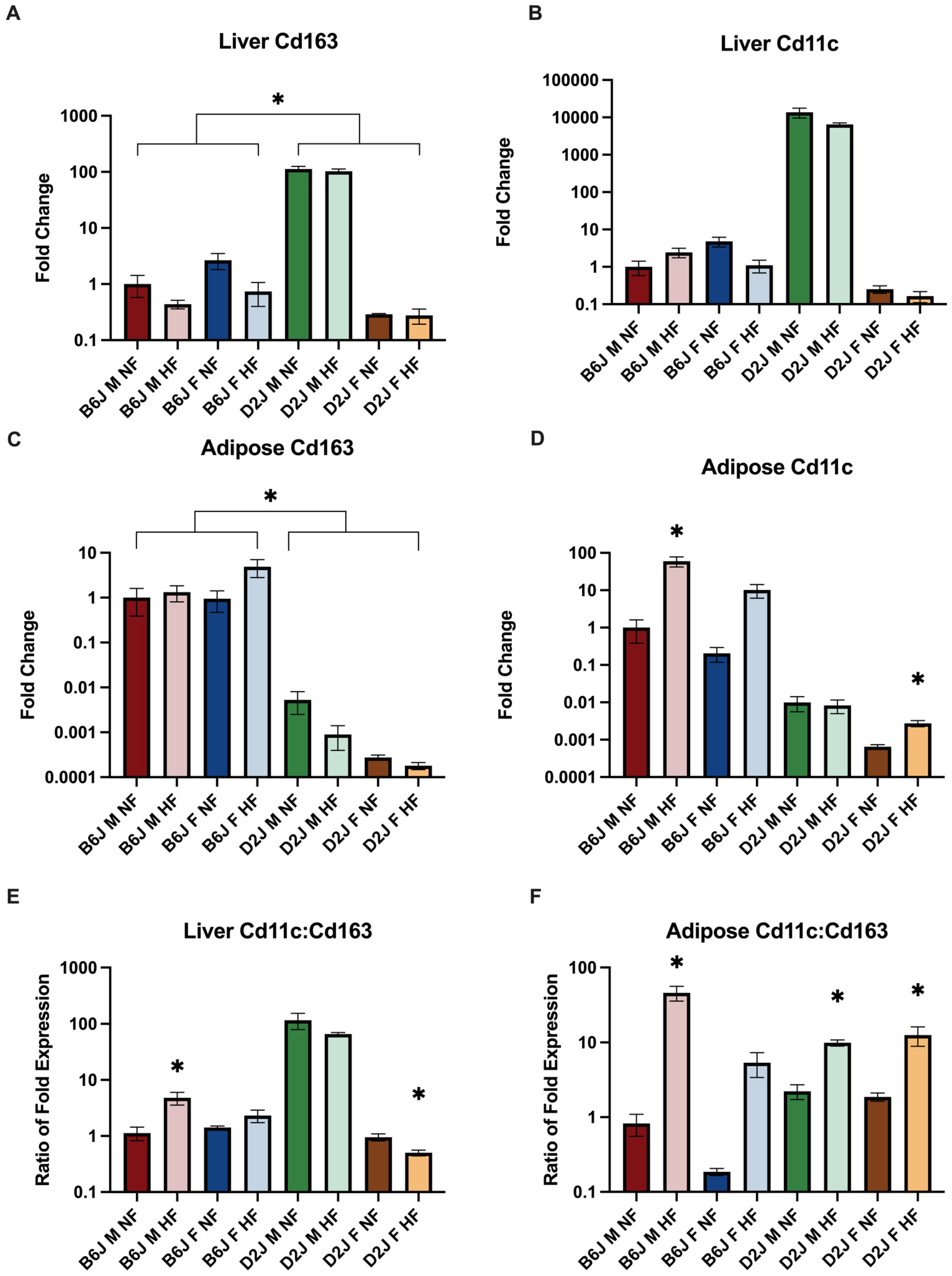
2.4. Effect of Diet, Sex, and Strain on the Gene Expression of Repair (M2-like) and Pro-inflammatory (M1-like) Macrophages
2.4.1. Liver
2.4.2. Adipose Tissue
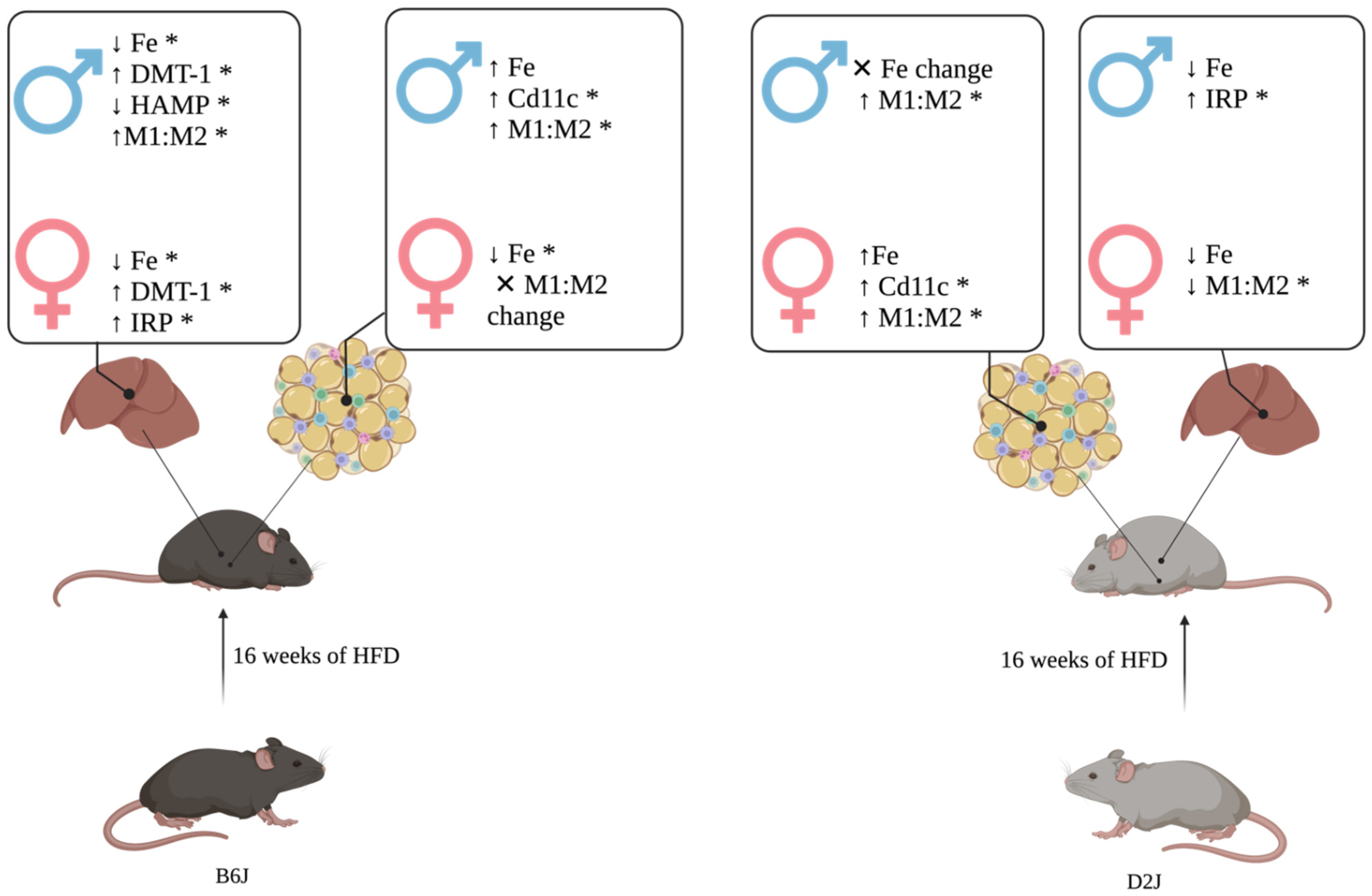
3. Discussion
3.1. HFD Impacted Trace Element Concentrations Differentially Due to Sex and Strain
3.2. Sex and Strain Impacted Relative Expression of Iron-Related Genes in Liver and Adipose Tissue
3.3. Macrophage Polarization May Explain Sex and Strain Differences in Adipose Iron Concentrations
4. Methods
4.1. Animals and Diet
4.2. Tissue Collection
4.3. Trace Element Concentration Analysis
4.4. RNA Isolation and cDNA Synthesis
4.5. Real Time Polymerase Chain Reaction (RT-PCR)
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fryar, C.D.; Carroll, M.D.; Afful, J. Prevalence of Obesity and Severe Obesity among Adults: United States, 2017–2018; NCHS: Hyattsville, MD, USA, 2020; p. 8. [Google Scholar]
- Martin-Jiménez, C.A.; Gaitán-Vaca, D.M.; Echeverria, V.; González, J.; Barreto, G.E. Relationship Between Obesity, Alzheimer’s Disease, and Parkinson’s Disease: An Astrocentric View. Mol. Neurobiol. 2017, 54, 7096–7115. [Google Scholar] [CrossRef] [PubMed]
- Pi-Sunyer, X. The Medical Risks of Obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Tinkov, A.A.; Gatiatulina, E.R.; Popova, E.V.; Polyakova, V.S.; Skalnaya, A.A.; Agletdinov, E.F.; Nikonorov, A.A.; Skalny, A.V. Early High-Fat Feeding Induces Alteration of Trace Element Content in Tissues of Juvenile Male Wistar Rats. Biol. Trace Elem. Res. 2017, 175, 367–374. [Google Scholar] [CrossRef]
- Totten, M.S.; Pierce, D.M.; Erikson, K.M. The Influence of Sex and Strain on Trace Element Dysregulation in the Brain Due to Diet-Induced Obesity. J. Trace Elem. Med. Biol. 2021, 63, 126661. [Google Scholar] [CrossRef] [PubMed]
- Birkenfeld, A.L.; Shulman, G.I. Non Alcoholic Fatty Liver Disease, Hepatic Insulin Resistance and Type 2 Diabetes. Hepatology 2014, 59, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Chen, M.; Liu, G.; Xu, E.; Chen, H. Ablation of Hephaestin and Ceruloplasmin Results in Iron Accumulation in Adipocytes and Type 2 Diabetes. FEBS Lett. 2018, 592, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Tinkov, A.A.; Sinitskii, A.I.; Popova, E.V.; Nemereshina, O.N.; Gatiatulina, E.R.; Skalnaya, M.G.; Skalny, A.V.; Nikonorov, A.A. Alteration of Local Adipose Tissue Trace Element Homeostasis as a Possible Mechanism of Obesity-Related Insulin Resistance. Med. Hypotheses 2015, 85, 343–347. [Google Scholar] [CrossRef]
- Sensi, S.L.; Granzotto, A.; Siotto, M.; Squitti, R. Copper and Zinc Dysregulation in Alzheimer’s Disease. Trends Pharmacol. Sci. 2018, 39, 1049–1063. [Google Scholar] [CrossRef]
- Gatiatulina, E.R.; Popova, E.V.; Polyakova, V.S.; Skalnaya, A.A.; Agletdinov, E.F.; Nikonorov, A.A.; Skalny, A.V.; Tinkov, A.A. Evaluation of Tissue Metal and Trace Element Content in a Rat Model of Non-Alcoholic Fatty Liver Disease Using ICP-DRC-MS. J. Trace Elem. Med. Biol. 2017, 39, 91–99. [Google Scholar] [CrossRef]
- Skjørringe, T.; Møller, L.; Moos, T. Impairment of Interrelated Iron- and Copper Homeostatic Mechanisms in Brain Contributes to the Pathogenesis of Neurodegenerative Disorders. Front. Pharmacol. 2012, 3, 169. [Google Scholar] [CrossRef]
- Ye, Q.; Park, J.E.; Gugnani, K.; Betharia, S.; Pino-Figueroa, A.; Kim, J. Influence of Iron Metabolism on Manganese Transport and Toxicity. Metallomics 2017, 9, 1028–1046. [Google Scholar] [CrossRef] [PubMed]
- Clayton, J.A. Applying the New SABV (Sex as a Biological Variable) Policy to Research and Clinical Care. Physiol. Behav. 2018, 187, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Norris, K.M.; Okie, W.; Kim, W.K.; Adhikari, R.; Yoo, S.; King, S.; Pazdro, R. A High-Fat Diet Differentially Regulates Glutathione Phenotypes in the Obesity-Prone Mouse Strains DBA/2J, C57BL/6J, and AKR/J. Nutr. Res. 2016, 36, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Chang, G.Q.; Dourmashkin, J.T.; Leibowitz, S.F. Distinct Phenotypes of Obesity-Prone AKR/J, DBA2J and C57BL/6J Mice Compared to Control Strains. Int. J. Obes. 2006, 30, 50–59. [Google Scholar] [CrossRef] [Green Version]
- West, D.B.; Boozer, C.N.; Moody, D.L.; Atkinson, R.L. Dietary Obesity in Nine Inbred Mouse Strains. Am. J. Physiol. 1992, 262, R1025–R1032. [Google Scholar] [CrossRef]
- Montgomery, M.K.; Hallahan, N.L.; Brown, S.H.; Liu, M.; Mitchell, T.W.; Cooney, G.J.; Turner, N. Mouse Strain-Dependent Variation in Obesity and Glucose Homeostasis in Response to High-Fat Feeding. Diabetologia 2013, 56, 1129–1139. [Google Scholar] [CrossRef]
- Totten, M.S.; Pierce, D.M.; Erikson, K.M. Diet-Induced Obesity Disrupts Trace Element Homeostasis and Gene Expression in the Olfactory Bulb. Nutrients 2020, 12, 3909. [Google Scholar] [CrossRef]
- Totten, M.S.; Wallace, C.W.; Pierce, D.M.; Fordahl, S.C.; Erikson, K.M. The Impact of a High-Fat Diet on Physical Activity and Dopamine Neurochemistry in the Striatum Is Sex and Strain Dependent in C57BL/6J and DBA/2J Mice. Nutr. Neurosci. 2021, 1–15. [Google Scholar] [CrossRef]
- Rodriguez, L.A.; Plomin, R.; Blizard, D.A.; Jones, B.C.; McClearn, G.E. Alcohol Acceptance, Preference, and Sensitivity in Mice. Alcohol. Clin. Exp. Res. 1994, 18, 1416–1422. [Google Scholar] [CrossRef]
- Martins, A.C.; López-Granero, C.; Ferrer, B.; Tinkov, A.A.; Skalny, A.V.; Paoliello, M.M.B.; Aschner, M. BXD Recombinant Inbred Mice as a Model to Study Neurotoxicity. Biomolecules 2021, 11, 1762. [Google Scholar] [CrossRef]
- Blizard, D.A. Recombinant-Inbred Strains: General Methodological Considerations Relevant to the Study of Complex Characters. Behav. Genet. 1992, 22, 621–633. [Google Scholar] [CrossRef] [PubMed]
- van der Heijden, R.A.; Sheedfar, F.; Morrison, M.C.; Hommelberg, P.P.; Kor, D.; Kloosterhuis, N.J.; Gruben, N.; Youssef, S.A.; de Bruin, A.; Hofker, M.H.; et al. High-Fat Diet Induced Obesity Primes Inflammation in Adipose Tissue Prior to Liver in C57BL/6j Mice. Aging 2015, 7, 256–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, I.S.; Heredia, F.P.d.; Wang, B.; Trayhurn, P. Cellular Hypoxia and Adipose Tissue Dysfunction in Obesity: Symposium on ‘Frontiers in Adipose Tissue Biology. Proc. Nutr. Soc. 2009, 68, 370–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheen, A.J.; Luyckx, F.H. Obesity and Liver Disease. Best Pract. Res. Clin. Endocrinol. Metab. 2002, 16, 703–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Li, P.; Huh, J.Y.; Hwang, I.J.; Lu, M.; Kim, J.I.; Ham, M.; Talukdar, S.; Chen, A.; Lu, W.J.; et al. Inflammation Is Necessary for Long-Term but Not Short-Term High-Fat Diet–Induced Insulin Resistance. Diabetes 2011, 60, 2474–2483. [Google Scholar] [CrossRef] [Green Version]
- Cai, D.; Yuan, M.; Frantz, D.F.; Melendez, P.A.; Hansen, L.; Lee, J.; Shoelson, S.E. Local and Systemic Insulin Resistance Resulting from Hepatic Activation of IKK-β and NF-ΚB. Nat. Med. 2005, 11, 183–190. [Google Scholar] [CrossRef]
- Hubler, M.J.; Peterson, K.R.; Hasty, A.H. Iron Homeostasis: A New Job for Macrophages in Adipose Tissue? Trends Endocrinol. Metab. TEM 2015, 26, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Andreux, P.A.; Williams, E.G.; Koutnikova, H.; Houtkooper, R.H.; Champy, M.-F.; Henry, H.; Schoonjans, K.; Williams, R.W.; Auwerx, J. Systems Genetics of Metabolism: The Use of the BXD Murine Reference Panel for Multiscalar Integration of Traits. Cell 2012, 150, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.; Kim, M.S.; Han, S.N. Diet-Induced Obesity Leads to Decreased Hepatic Iron Storage in Mice. Nutr. Res. 2011, 31, 915–921. [Google Scholar] [CrossRef]
- McLachlan, S.; Page, K.E.; Lee, S.-M.; Loguinov, A.; Valore, E.; Hui, S.T.; Jung, G.; Zhou, J.; Lusis, A.J.; Fuqua, B.; et al. Hamp1 MRNA and Plasma Hepcidin Levels Are Influenced by Sex and Strain but Do Not Predict Tissue Iron Levels in Inbred Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G511–G523. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Hepcidin and Disorders of Iron Metabolism. Annu. Rev. Med. 2011, 62, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin Regulates Cellular Iron Efflux by Binding to Ferroportin and Inducing Its Internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumerle, S.; Mathieu, J.R.R.; Delga, S.; Heinis, M.; Viatte, L.; Vaulont, S.; Peyssonnaux, C. Targeted Disruption of Hepcidin in the Liver Recapitulates the Hemochromatotic Phenotype. Blood 2014, 123, 3646–3650. [Google Scholar] [CrossRef] [Green Version]
- Moore Heslin, A.; O’Donnell, A.; Buffini, M.; Nugent, A.P.; Walton, J.; Flynn, A.; McNulty, B.A. Risk of Iron Overload in Obesity and Implications in Metabolic Health. Nutrients 2021, 13, 1539. [Google Scholar] [CrossRef]
- Sangkhae, V.; Nemeth, E. Regulation of the Iron Homeostatic Hormone Hepcidin. Adv. Nutr. 2017, 8, 126–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotardo, É.M.F.; dos Santos, A.N.; Miyashiro, R.A.; Gambero, S.; Rocha, T.; Ribeiro, M.L.; Gambero, A. Mice That Are Fed a High-Fat Diet Display Increased Hepcidin Expression in Adipose Tissue. J. Nutr. Sci. Vitaminol. 2013, 59, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Bekri, S.; Gual, P.; Anty, R.; Luciani, N.; Dahman, M.; Ramesh, B.; Iannelli, A.; Staccini-Myx, A.; Casanova, D.; Ben Amor, I.; et al. Increased Adipose Tissue Expression of Hepcidin in Severe Obesity Is Independent From Diabetes and NASH. Gastroenterology 2006, 131, 788–796. [Google Scholar] [CrossRef]
- Sonnweber, T.; Ress, C.; Nairz, M.; Theurl, I.; Schroll, A.; Murphy, A.T.; Wroblewski, V.; Witcher, D.R.; Moser, P.; Ebenbichler, C.F.; et al. High-Fat Diet Causes Iron Deficiency via Hepcidin-Independent Reduction of Duodenal Iron Absorption. J. Nutr. Biochem. 2012, 23, 1600–1608. [Google Scholar] [CrossRef]
- Citelli, M.; Fonte-Faria, T.; Nascimento-Silva, V.; Renovato-Martins, M.; Silva, R.; Luna, A.S.; Vargas da Silva, S.; Barja-Fidalgo, C. Obesity Promotes Alterations in Iron Recycling. Nutrients 2015, 7, 335–348. [Google Scholar] [CrossRef] [Green Version]
- Dupic, F.; Fruchon, S.; Bensaid, M.; Loreal, O.; Brissot, P.; Borot, N.; Roth, M.P.; Coppin, H. Duodenal MRNA Expression of Iron Related Genes in Response to Iron Loading and Iron Deficiency in Four Strains of Mice. Gut 2002, 51, 648–653. [Google Scholar] [CrossRef]
- Winn, N.C.; Volk, K.M.; Hasty, A.H. Regulation of Tissue Iron Homeostasis: The Macrophage “Ferrostat”. JCI Insight 2020, 5, e132964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corna, G.; Campana, L.; Pignatti, E.; Castiglioni, A.; Tagliafico, E.; Bosurgi, L.; Campanella, A.; Brunelli, S.; Manfredi, A.A.; Apostoli, P.; et al. Polarization Dictates Iron Handling by Inflammatory and Alternatively Activated Macrophages. Haematologica 2010, 95, 1814–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caslin, H.L.; Bhanot, M.; Bolus, W.R.; Hasty, A. Adipose Tissue Macrophages: Unique Polarization and Bioenergetics in Obesity. Immunol. Rev. 2020, 295, 101–113. [Google Scholar] [CrossRef]
- Orr, J.S.; Kennedy, A.; Anderson-Baucum, E.K.; Webb, C.D.; Fordahl, S.C.; Erikson, K.M.; Zhang, Y.; Etzerodt, A.; Moestrup, S.K.; Hasty, A.H. Obesity Alters Adipose Tissue Macrophage Iron Content and Tissue Iron Distribution. Diabetes 2014, 63, 421–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recalcati, S.; Locati, M.; Marini, A.; Santambrogio, P.; Zaninotto, F.; De Pizzol, M.; Zammataro, L.; Girelli, D.; Cairo, G. Differential Regulation of Iron Homeostasis during Human Macrophage Polarized Activation. Eur. J. Immunol. 2010, 40, 824–835. [Google Scholar] [CrossRef]
- Gabrielsen, J.S.; Gao, Y.; Simcox, J.A.; Huang, J.; Thorup, D.; Jones, D.; Cooksey, R.C.; Gabrielsen, D.; Adams, T.D.; Hunt, S.C.; et al. Adipocyte Iron Regulates Adiponectin and Insulin Sensitivity. J. Clin. Investig. 2012, 122, 3529–3540. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Navarrete, J.M.; Novelle, M.G.; Catalán, V.; Ortega, F.; Moreno, M.; Gomez-Ambrosi, J.; Xifra, G.; Serrano, M.; Guerra, E.; Ricart, W.; et al. Insulin Resistance Modulates Iron-Related Proteins in Adipose Tissue. Diabetes Care 2014, 37, 1092–1100. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, J.H.; Etzerodt, A.; Svendsen, P.; Moestrup, S.K. The Haptoglobin-CD163-Heme Oxygenase-1 Pathway for Hemoglobin Scavenging. Oxid. Med. Cell. Longev. 2013, 2013, 523652. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.A.; Bolus, W.R.; Hasty, A.H. A Decade of Progress in Adipose Tissue Macrophage Biology. Immunol. Rev. 2014, 262, 134–152. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity Induces a Phenotypic Switch in Adipose Tissue Macrophage Polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef]
- Prieur, X.; Mok, C.Y.L.; Velagapudi, V.R.; Núñez, V.; Fuentes, L.; Montaner, D.; Ishikawa, K.; Camacho, A.; Barbarroja, N.; O’Rahilly, S.; et al. Differential Lipid Partitioning Between Adipocytes and Tissue Macrophages Modulates Macrophage Lipotoxicity and M2/M1 Polarization in Obese Mice. Diabetes 2011, 60, 797–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.S.; Jung, D.Y.; Morel, C.; Lakhani, S.A.; Kim, J.K.; Flavell, R.A.; Davis, R.J. JNK Expression by Macrophages Promotes Obesity-Induced Insulin Resistance and Inflammation. Science 2013, 339, 218–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaul, M.E.; Bennett, G.; Strissel, K.J.; Greenberg, A.S.; Obin, M.S. Dynamic, M2-Like Remodeling Phenotypes of CD11c+ Adipose Tissue Macrophages During High-Fat Diet–Induced Obesity in Mice. Diabetes 2010, 59, 1171–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C.J. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The Aligned Rank Transform for Nonparametric Factorial Analyses Using Only Anova Procedures. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Vancouver, BC, Canada, 7–12 May 2011; pp. 143–146. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 22 April 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Sex | Diet | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Week | |||||||||||||||||||
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | Average | ||||
| B6J | Male | Normal Fat | 2.46 ± 0.12 | 7.53 ± 0.96 | 6.45 ± 1.52 | 15.66 ± 4.94 | 14.96 ± 1.73 | 13.15 ± 2.60 | 14.81 ± 0.47 | −0.40 ± 15.58 | 21.13 ± 9.09 | 28.40 ± 11.85 | 38.14 ± 64.80 | 10.99 ± 1.96 | 14.18 ± 1.36 | 57.41 ± 53.23 | 23.99 ± 20.88 | 17.93 ± 4.59 | |
| B6J | Male | High Fat | 2.05 ± 0.17 | 5.58 ± 0.72 | 5.78 ± 0.47 | 4.99 ± 0.35 | 5.37 ± 0.44 | 4.67 ± 0.29 | 5.40 ± 0.57 | 5.46 ± 0.54 | 5.28 ± 0.54 | 6.90 ± 0.77 | 8.56 ± 2.05 | 10.05 ± 2.80 | 12.22 ± 2.40 | 19.29 ± 2.12 | 17.08 ± 11.49 | 7.91 ± 0.46 | |
| B6J | Female | Normal Fat | 4.21 ± 0.26 | 6.36 ± 0.91 | 9.63 ± 1.33 | 22.44 ± 7.96 | 20.59 ± 5.51 | 93.79 ± 102.42 | 82.23 ± 83.65 | −71.24 ± 108.48 | 16.06 ± 3.00 | −2.42 ± 33.33 | −15.98 ± 70.66 | 12.11 ± 21.62 | −2.38 ± 29.74 | −2.74 ± 16.09 | −75.12 ± 111.54 | 6.50 ± 19.337 | |
| B6J | Female | High Fat | 3.00 ± 0.05 | 5.32 ± 0.80 | 10.75 ± 2.15 | 9.61 ± 1.68 | 7.66 ± 1.70 | 6.62 ± 1.89 | 6.43 ± 0.67 | 6.02 ± 0.16 | 7.89 ± 1.71 | 7.99 ± 1.43 | 10.24 ± 2.92 | 8.24 ± 0.70 | 8.42 ± 1.13 | 8.63 ± 1.65 | 9.32 ± 0.81 | 7.74 ± 0.54 | |
| D2J | Male | Normal Fat | 2.71 ± 0.14 | 6.54 ± 3.19 | 6.18 ± 1.12 | 9.39 ± 3.03 | −5.49 ± 23.71 | 11.33 ± 2.99 | 14.00 ± 0.98 | 15.13 ± 3.92 | 16.26 ± 6.95 | −57.69 ± 151.16 | −121.64 ± 110.97 | 28.06 ± 14.74 | 45.04 ± 69.20 | 2.18 ± 30.39 | 56.68 ± 24.26 | 1.91 ± 10.29 | |
| D2J | Male | High Fat | 1.98 ± 0.08 | 3.11 ± 0.18 | 3.76 ± 0.38 | 3.96 ± 0.40 | 6.05 ± 0.58 | 6.47 ± 0.72 | 7.37 ± 1.28 | 12.17 ± 3.49 | 7.32 ± 6.66 | 7.92 ± 3.12 | 36.27 ± 22.68 | 11.03 ± 2.32 | 11.93 ± 2.26 | 34.51 ± 17.07 | 34.21 ± 24.61 | 12.54 ± 1.88 | |
| D2J | Female | Normal Fat | 3.31 ± 0.35 | 7.57 ± 2.36 | 17.64 ± 15.66 | 9.51 ± 2.27 | 10.25 ± 3.28 | 12.37 ± 31.05 | 9.74 ± 3.26 | −26.82 ± 12.04 | 25.01 ± 18.88 | −35.43 ± 174.47 | −1.25 ± 31.56 | −27.71 ± 24.56 | 23.40 ± 9.28 | 15.94 ± 20.34 | −45.10 ± 46.62 | −0.11 ± 4.84 | |
| D2J | Female | High Fat | 2.62 ± 0.08 | 3.05 ± 0.32 | 4.19 ± 0.25 | 4.85 ± 0.35 | 11.66 ± 6.09 | 9.17 ± 3.26 | 6.33 ± 1.24 | 10.71 ± 4.50 | 8.93 ± 1.03 | −11.26 ± 34.95 | 25.11 ± 30.06 | 31.97 ± 23.42 | 3.90 ± 22.97 | −47.94 ± 26.12 | −4.89 ± 42.44 | 3.89 ± 4.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hagarty-Waite, K.A.; Totten, M.S.; Pierce, M.; Armah, S.M.; Erikson, K.M. Influence of Sex and Strain on Hepatic and Adipose Tissue Trace Element Concentrations and Gene Expression in C57BL/6J and DBA/2J High Fat Diet Models. Int. J. Mol. Sci. 2022, 23, 13778. https://doi.org/10.3390/ijms232213778
Hagarty-Waite KA, Totten MS, Pierce M, Armah SM, Erikson KM. Influence of Sex and Strain on Hepatic and Adipose Tissue Trace Element Concentrations and Gene Expression in C57BL/6J and DBA/2J High Fat Diet Models. International Journal of Molecular Sciences. 2022; 23(22):13778. https://doi.org/10.3390/ijms232213778
Chicago/Turabian StyleHagarty-Waite, Kristen A., Melissa S. Totten, Matthew Pierce, Seth M. Armah, and Keith M. Erikson. 2022. "Influence of Sex and Strain on Hepatic and Adipose Tissue Trace Element Concentrations and Gene Expression in C57BL/6J and DBA/2J High Fat Diet Models" International Journal of Molecular Sciences 23, no. 22: 13778. https://doi.org/10.3390/ijms232213778