
Promotion of Melanoma Cell Proliferation by Cyclic Straining through Regulatory Morphogenesis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
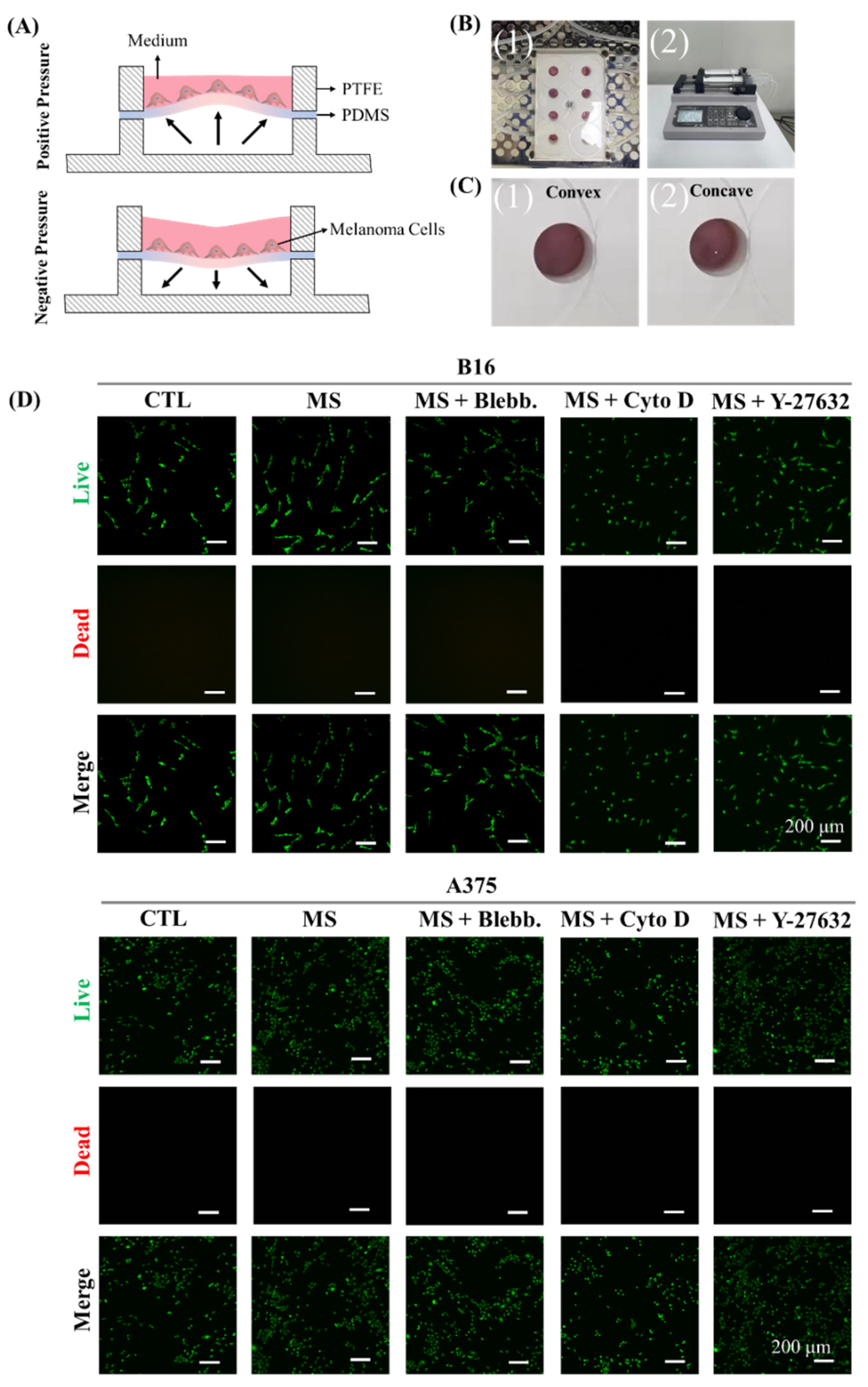
2.1. Cell Culture System for Cyclic Straining
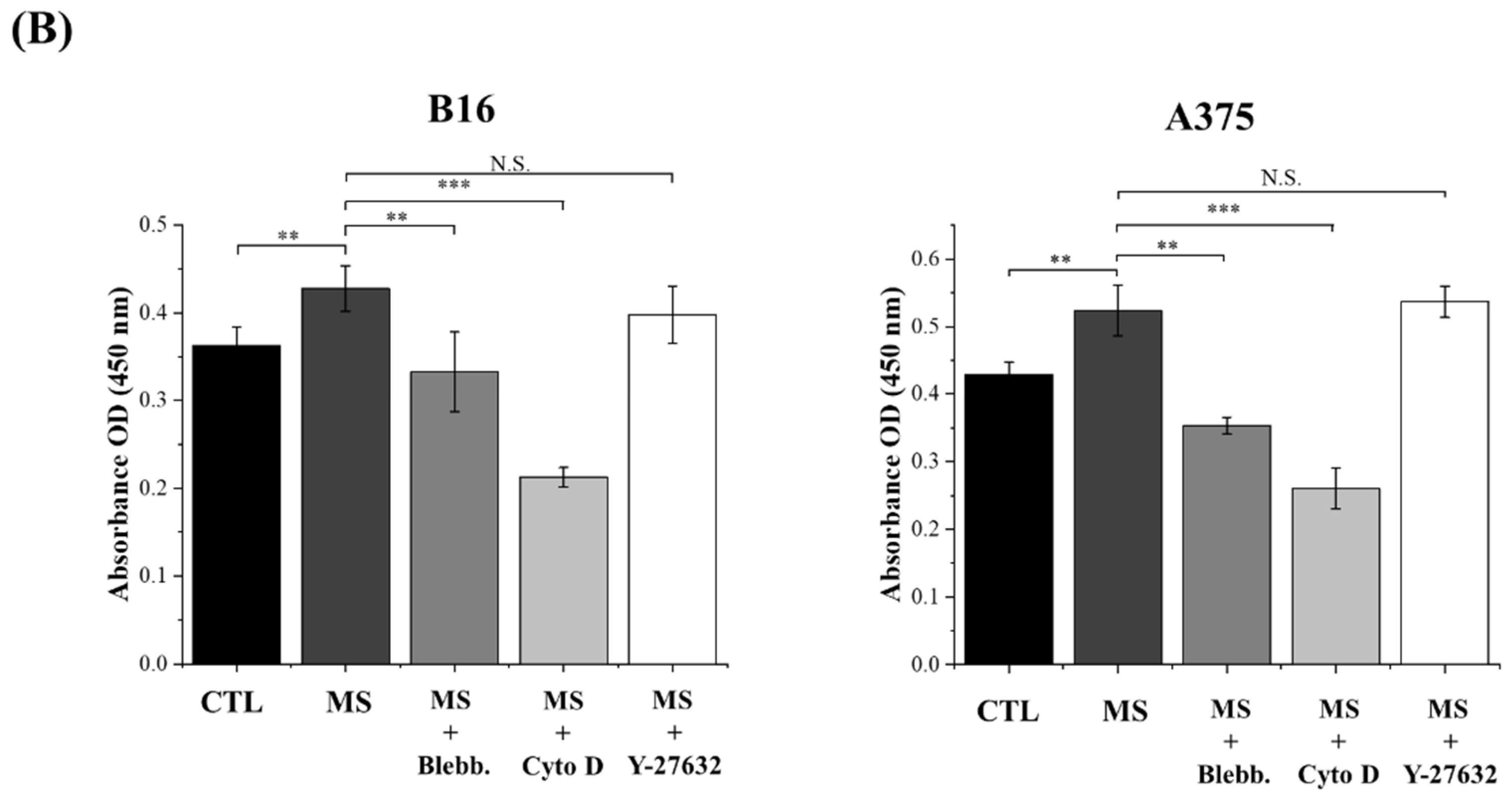
2.2. Melanoma Cell Proliferation
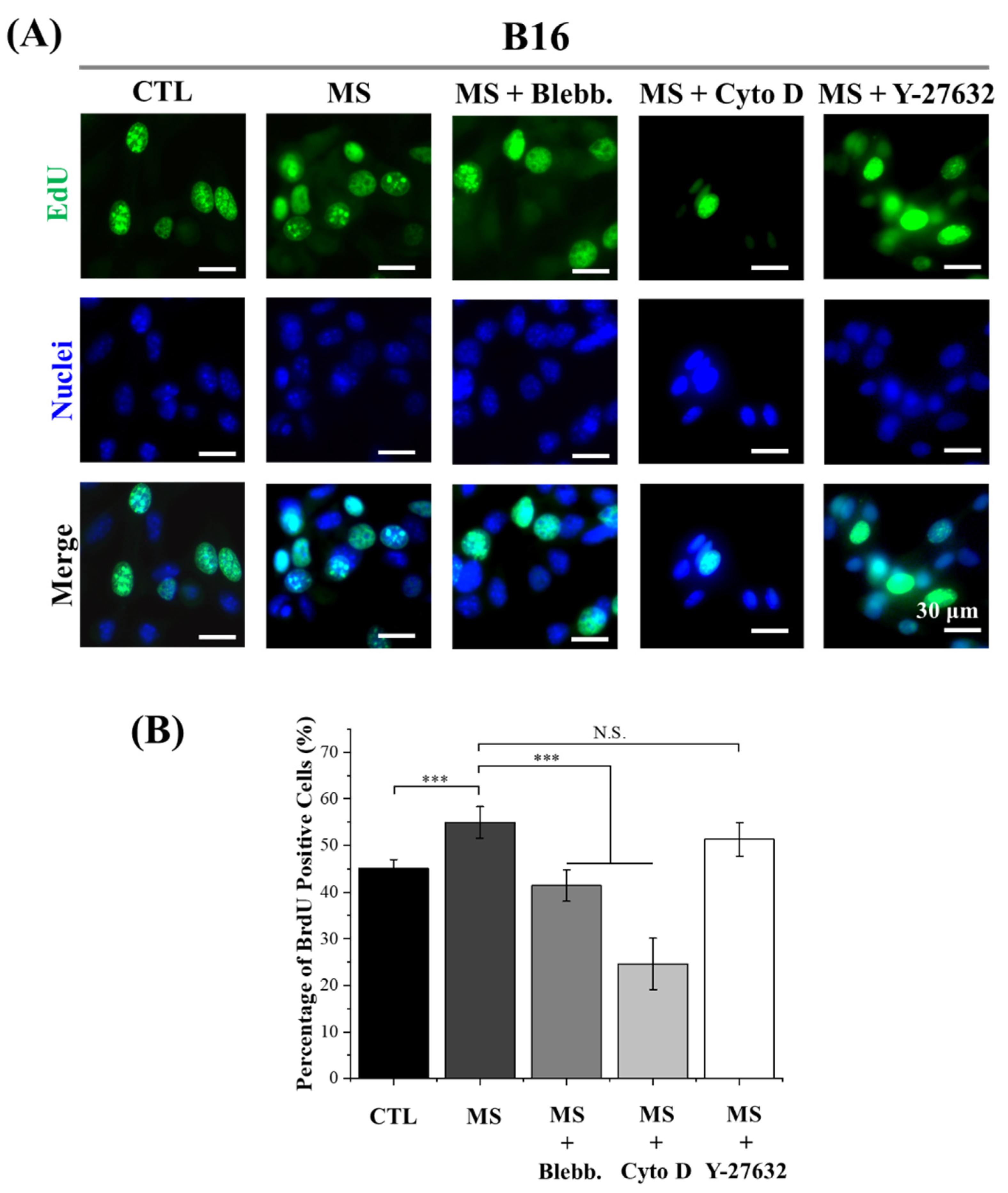
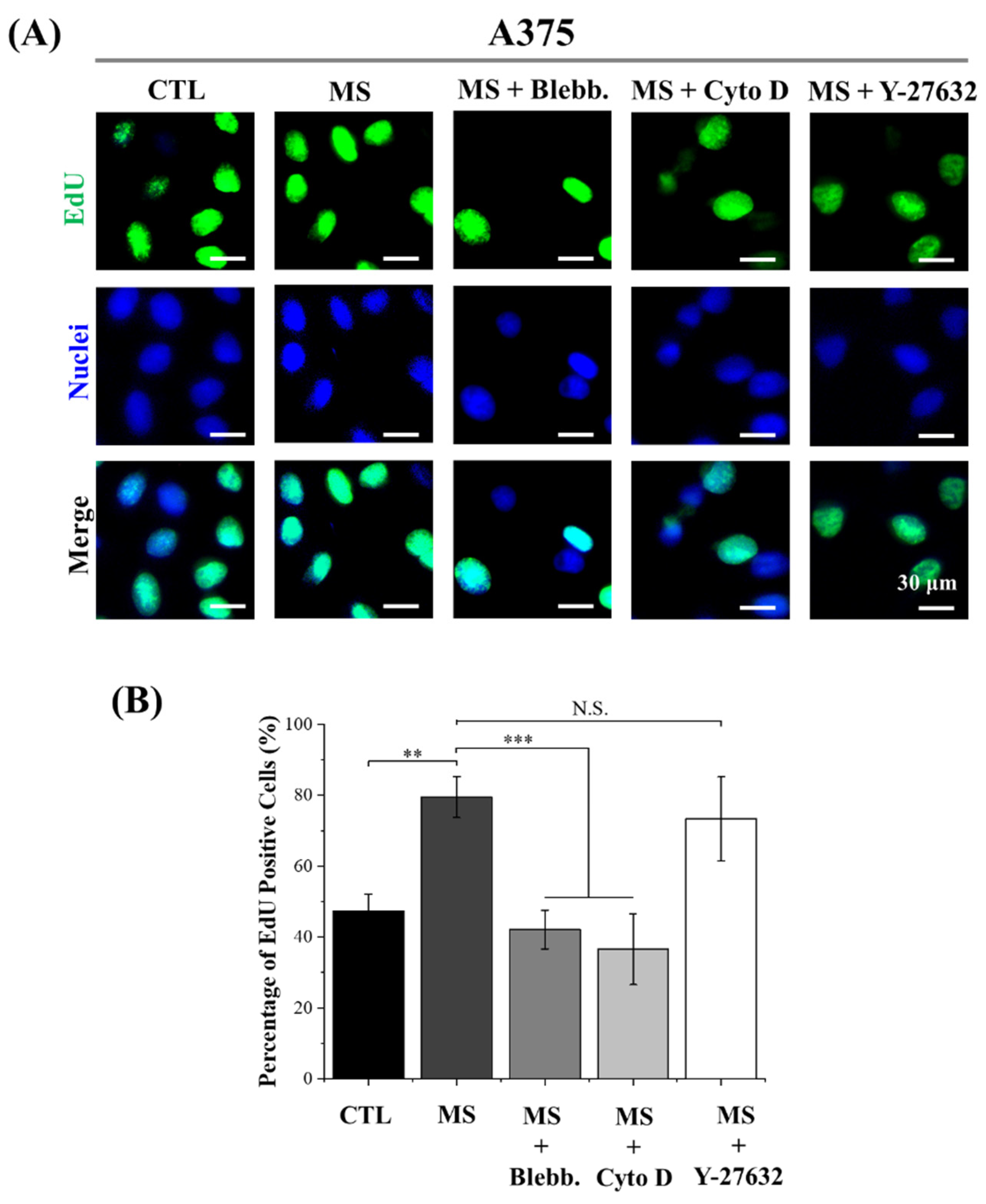
2.3. DNA Synthesis Activity
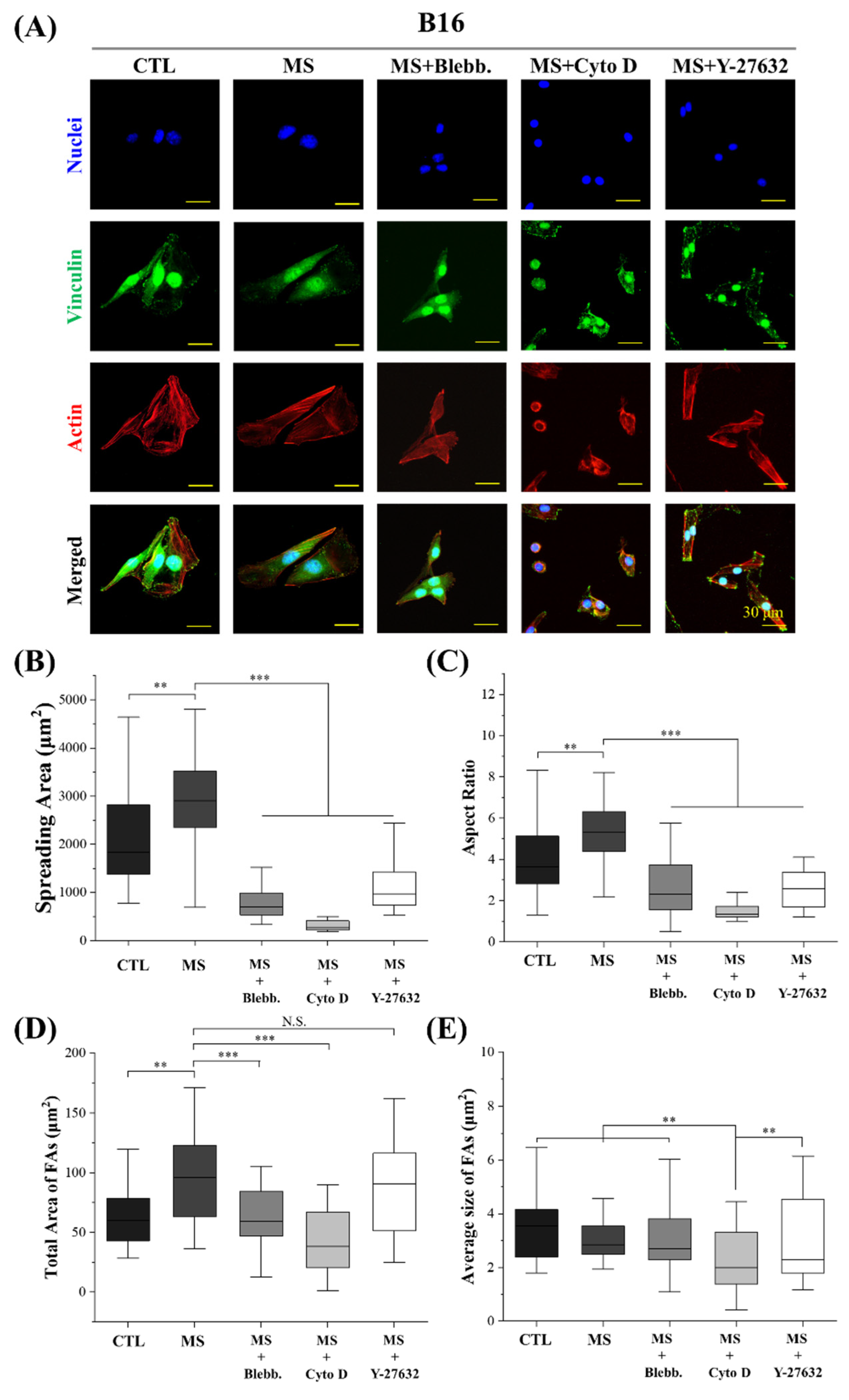
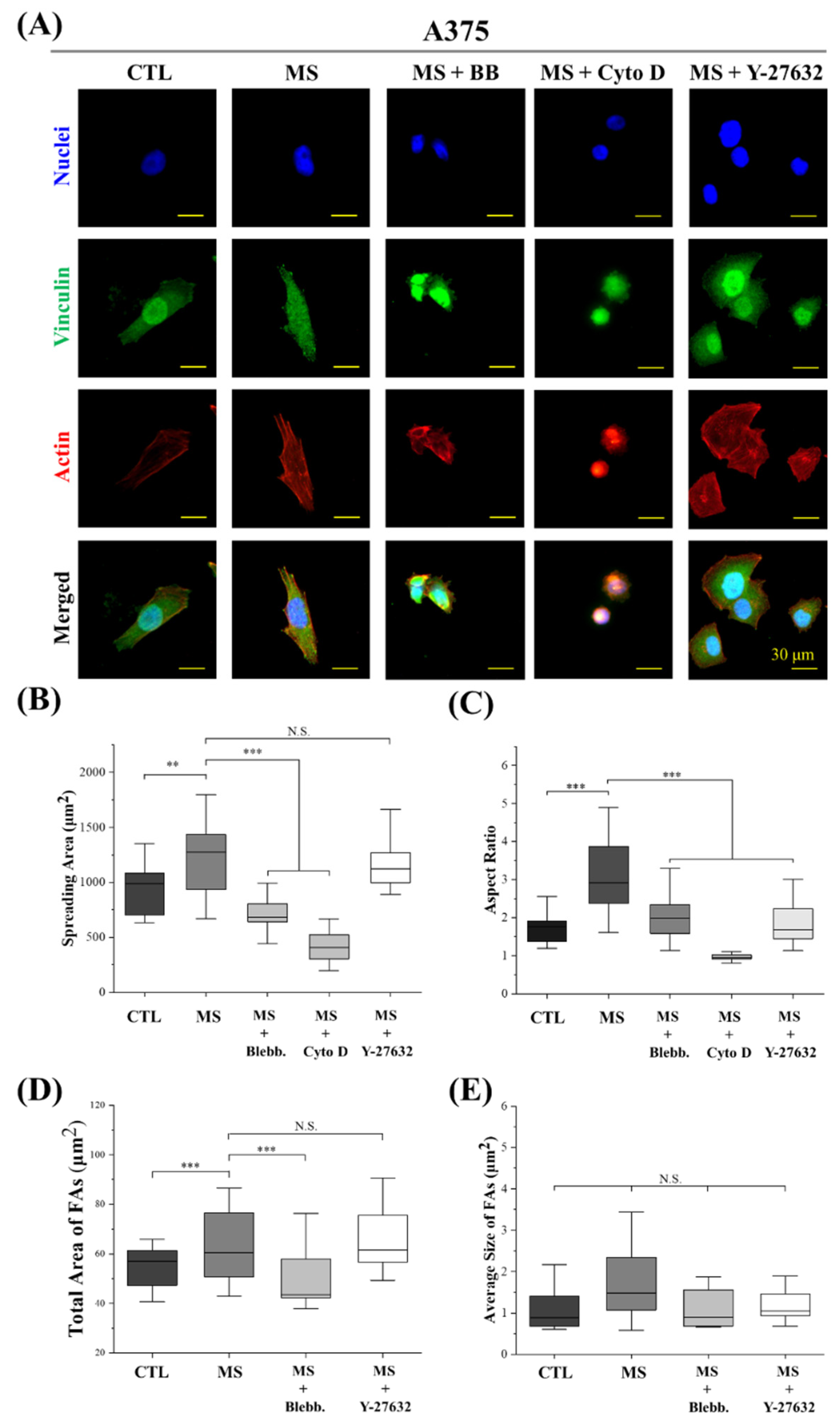
2.4. Cell Morphology and FAs
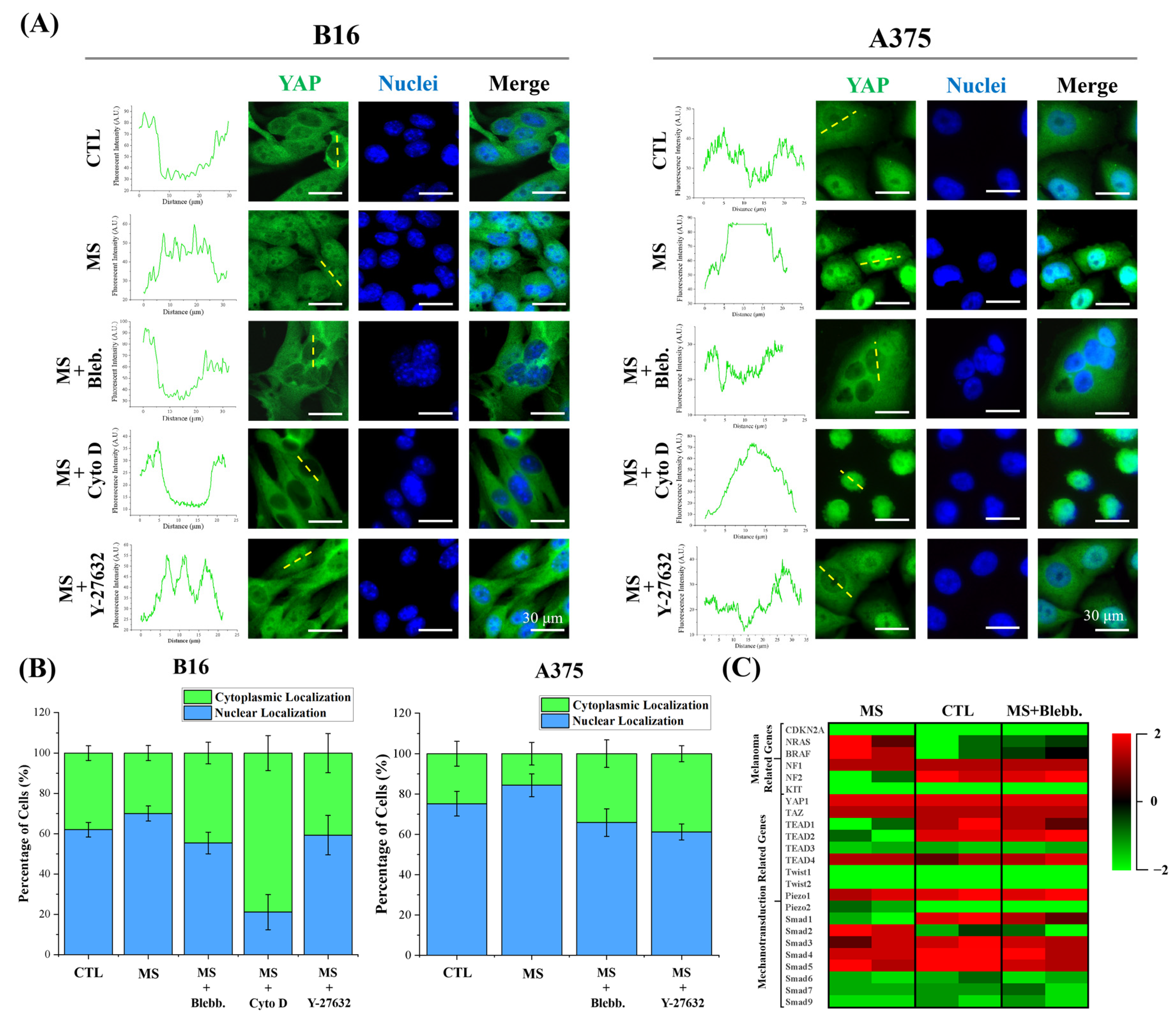
2.5. YAP Activity and Melanoma Gene Expression
3. Materials and Methods
3.1. Design and Preparation of Cell Culture System
3.2. Cell Culture
3.3. Belbbistatin, Cytochalasin D and Y-27632 Treatment
3.4. Cell Viability Analysis
3.5. Actin, Vinculin, and Nuclei Staining
3.6. CCK-8 and EdU Incorporation Assay
3.7. YAP and Nuclei Staining
3.8. Eukaryotic Transcriptome Sequencing and Differentially Gene Expression Analyses
3.9. Statistical Analysis
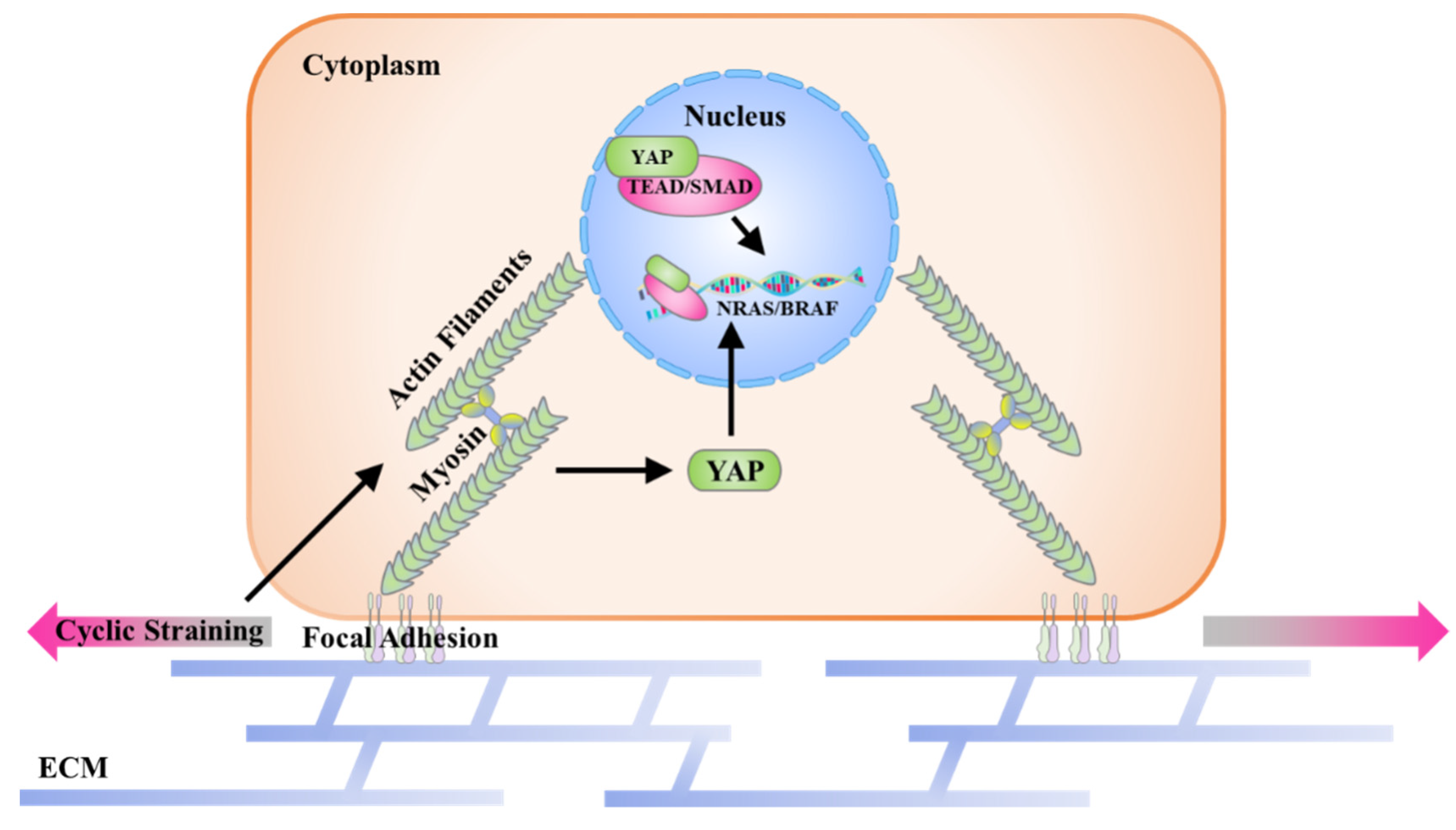
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schadendorf, D.; van Akkooi, A.C.J.; Berking, C.; Griewank, K.G.; Gutzmer, R.; Hauschild, A.; Stang, A.; Roesch, A.; Ugurel, S. Melanoma. Lancet 2018, 392, 971–984. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Ma, S. Racial differences in six major subtypes of melanoma: Descriptive epidemiology. BMC Cancer 2016, 16, 691. [Google Scholar] [CrossRef] [Green Version]
- Mounessa, J.S.; Caravaglio, J.V.; Dellavalle, R.P. Comparison of Regional and State Differences in Melanoma Rates in the United States: 2003 vs. 2013. JAMA Dermatol. 2017, 153, 345–347. [Google Scholar] [CrossRef] [PubMed]
- Neugut, A.I.; Kizelnik-Freilich, S.; Ackerman, C. Black-white differences in risk for cutaneous, ocular, and visceral melanomas. Am. J. Public Health 1994, 84, 1828–1829. [Google Scholar] [CrossRef] [Green Version]
- Bulliard, J.-L. Site-specific risk of cutaneous malignant melanoma and pattern of Sun exposure in New Zealand. Int. J. Cancer 2000, 85, 627–632. [Google Scholar] [CrossRef]
- Khan, A.Q.; Travers, J.B.; Kemp, M.G. Roles of UVA radiation and DNA damage responses in melanoma pathogenesis. Environ. Mol. Mutagen. 2018, 59, 438–460. [Google Scholar] [CrossRef] [PubMed]
- Budden, T.; Bowden, N.A. The Role of Altered Nucleotide Excision Repair and UVB-Induced DNA Damage in Melanomagenesis. Int. J. Mol. Sci. 2013, 14, 1132–1151. [Google Scholar] [CrossRef] [Green Version]
- Shain, A.H.; Yu, R.; Yeh, I.; Benhamida, J.; Kovalyshyn, I.; Sriharan, A.; Talevich, E.; Dummer, R.; North, J.; Pincus, L.; et al. Abstract 2372: The genetic evolution of melanoma. Cancer Res. 2016, 76, 2372. [Google Scholar] [CrossRef]
- Chen, L.; Jin, S. Trends in mortality rates of cutaneous melanoma in East Asian populations. PeerJ 2016, 4, e2809. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.W.-C. Acral Melanoma: A Unique Disease in Asia. JAMA Dermatol. 2013, 149, 1272–1273. [Google Scholar] [CrossRef] [PubMed]
- Goydos, J.S.; Shoen, S.L. Acral Lentiginous Melanoma. In Melanoma; Kaufman, H.L., Mehnert, J.M., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 321–329. [Google Scholar]
- Haugh, A.M.; Zhang, B.; Quan, V.L.; Garfield, E.M.; Bubley, J.A.; Kudalkar, E.; Verzi, A.E.; Walton, K.; VandenBoom, T.; Merkel, E.A.; et al. Distinct Patterns of Acral Melanoma Based on Site and Relative Sun Exposure. J. Investig. Dermatol. 2018, 138, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Moon, K.R.; Choi, Y.D.; Kim, J.M.; Jin, S.; Shin, M.-H.; Shim, H.-J.; Lee, J.-B.; Yun, S.J. Genetic Alterations in Primary Acral Melanoma and Acral Melanocytic Nevus in Korea: Common Mutated Genes Show Distinct Cytomorphological Features. J. Investig. Dermatol. 2018, 138, 933–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, A.; Touzet, S.; Dalle, S.; Ronger-Savlé, S.; Balme, B.; Thomas, L. Acral lentiginous melanoma: A clinicoprognostic study of 126 cases. Br. J. Dermatol. 2006, 155, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, A.; Omodaka, T.; Okuyama, R. Melanomas and Mechanical Stress Points on the Plantar Surface of the Foot. N. Engl. J. Med. 2016, 374, 2404–2406. [Google Scholar] [CrossRef] [Green Version]
- Sheen, Y.-S.; Liao, Y.-H.; Lin, M.-H.; Chen, J.-S.; Liau, J.-Y.; Tseng, Y.-J.; Lee, C.-H.; Chang, Y.-L.; Chu, C.-Y. A clinicopathological analysis of 153 acral melanomas and the relevance of mechanical stress. Sci. Rep. 2017, 7, 5564. [Google Scholar] [CrossRef]
- Costello, C.M.; Pittelkow, M.R.; Mangold, A.R. Acral Melanoma and Mechanical Stress on the Plantar Surface of the Foot. N. Engl. J. Med. 2017, 377, 395–396. [Google Scholar] [CrossRef]
- Gong, H.Z.; Zhang, S.; Zheng, H.Y.; Qu, T.; Li, J. The role of mechanical stress in the formation of plantar melanoma: A retrospective analysis of 72 chinese patients with plantar melanomas and a meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Rambhia, P.H.; Stojanov, I.J.; Arbesman, J. Predominance of oral mucosal melanoma in areas of high mechanical stress. J. Am. Acad. Dermatol. 2019, 80, 1133–1135. [Google Scholar] [CrossRef] [Green Version]
- Orr, A.W.; Helmke, B.P.; Blackman, B.R.; Schwartz, M.A. Mechanisms of Mechanotransduction. Dev. Cell 2006, 10, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Prauzner-Bechcicki, S.; Raczkowska, J.; Madej, E.; Pabijan, J.; Lukes, J.; Sepitka, J.; Rysz, J.; Awsiuk, K.; Bernasik, A.; Budkowski, A.; et al. PDMS substrate stiffness affects the morphology and growth profiles of cancerous prostate and melanoma cells. J. Mech. Behav. Biomed. Mater. 2015, 41, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Morikura, T.; Miyata, S. Effect of Mechanical Compression on Invasion Process of Malignant Melanoma Using In Vitro Three-Dimensional Cell Culture Device. Micromachines 2019, 10, 666. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Kim, H.; Min, K.I.; Kim, C.; Kwon, Y.; Zheng, Z.; Kim, Y.; Park, H.-S.; Ju, Y.S.; Roh, M.R.; et al. Weight-bearing activity impairs nuclear membrane and genome integrity via YAP activation in plantar melanoma. Nat. Commun. 2022, 13, 2214. [Google Scholar] [CrossRef]
- Wang, N. Review of cellular mechanotransduction. J. Phys. D Appl. Phys. 2017, 50, 233002. [Google Scholar] [CrossRef]
- Schwartz, M.A.; DeSimone, D.W. Cell adhesion receptors in mechanotransduction. Curr. Opin. Cell Biol. 2008, 20, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Ciobanasu, C.; Faivre, B.; Le Clainche, C. Actin dynamics associated with focal adhesions. Int. J. Cell Biol. 2012, 2012, 941292. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Kim, J. Regulation of Hippo signaling by actin remodeling. BMB Rep. 2018, 51, 151–156. [Google Scholar] [CrossRef]
- Oka, T.; Sudol, M.J.G.t.C. Nuclear localization and pro-apoptotic signaling of YAP2 require intact PDZ-binding motif. Genes Cells 2009, 14, 607–615. [Google Scholar] [CrossRef]
- Miskolczi, Z.; Smith, M.P.; Rowling, E.J.; Ferguson, J.; Barriuso, J.; Wellbrock, C. Collagen abundance controls melanoma phenotypes through lineage-specific microenvironment sensing. Oncogene 2018, 37, 3166–3182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeichen, J.; van Griensven, M.; Bosch, U. The Proliferative Response of Isolated Human Tendon Fibroblasts to Cyclic Biaxial Mechanical Strain. Am. J. Sports Med. 2000, 28, 888–892. [Google Scholar] [CrossRef]
- Lee, W.-C.C.; Maul, T.M.; Vorp, D.A.; Rubin, J.P.; Marra, K.G. Effects of uniaxial cyclic strain on adipose-derived stem cell morphology, proliferation, and differentiation. Biomech. Model. Mechanobiol. 2007, 6, 265–273. [Google Scholar] [CrossRef]
- Chin, L.; Xia, Y.; Discher, D.E.; Janmey, P.A. Mechanotransduction in cancer. Curr. Opin. Chem. Eng. 2016, 11, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Amano, M.; Nakayama, M.; Kaibuchi, K. Rho-kinase/ROCK: A key regulator of the cytoskeleton and cell polarity. Cytoskeleton 2010, 67, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Chang, F.; Zhang, Y.; Mi, J.; Zhou, Q.; Bai, F.; Xu, X.; Fisher, D.E.; Sun, Q.; Wu, X. ROCK inhibitor enhances the growth and migration of BRAF-mutant skin melanoma cells. Cancer Sci. 2018, 109, 3428–3437. [Google Scholar] [CrossRef]
- Zhang, S.; Cao, S.; Gong, M.; Zhang, W.; Zhang, W.; Zhu, Z.; Wu, S.; Yue, Y.; Qian, W.; Ma, Q.; et al. Mechanically activated ion channel Piezo1 contributes to melanoma malignant progression through AKT/mTOR signaling. Cancer Biol. Ther. 2022, 23, 336–347. [Google Scholar] [CrossRef]
- Della Pietra, A.; Mikhailov, N.; Giniatullin, R. The Emerging Role of Mechanosensitive Piezo Channels in Migraine Pain. Int. J. Mol. Sci. 2020, 21, 696. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Liu, C.; Zhang, D.; Men, H.; Huo, L.; Geng, Q.; Wang, S.; Gao, Y.; Zhang, W.; Zhang, Y.; et al. Mechanosensitive ion channel Piezo1 promotes prostate cancer development through the activation of the Akt/mTOR pathway and acceleration of cell cycle. Int. J. Oncol. 2019, 55, 629–644. [Google Scholar] [CrossRef] [Green Version]
- Elliott, H.; Fischer, R.S.; Myers, K.A.; Desai, R.A.; Gao, L.; Chen, C.S.; Adelstein, R.S.; Waterman, C.M.; Danuser, G. Myosin II controls cellular branching morphogenesis and migration in three dimensions by minimizing cell-surface curvature. Nat. Cell Biol. 2015, 17, 137–147. [Google Scholar] [CrossRef]
- Chisholm, R.L.; Firtel, R.A. Insights into morphogenesis from a simple developmental system. Nat. Rev. Mol. Cell Biol. 2004, 5, 531–541. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, X.; Wang, Y.; Hu, X.; Kawazoe, N.; Yang, Y.; Chen, G. Influence of Cell Spreading Area on the Osteogenic Commitment and Phenotype Maintenance of Mesenchymal Stem Cells. Sci. Rep. 2019, 9, 6891. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, X.; Huang, T.C.; Hu, X.; Kawazoe, N.; Tsai, W.B.; Yang, Y.; Chen, G. Regulation of mesenchymal stem cell functions by micro-nano hybrid patterned surfaces. J. Mater. Chem. B 2018, 6, 5424–5434. [Google Scholar] [CrossRef]
- Wang, Y.; Yoshitomi, T.; Kawazoe, N.; Yang, Y.; Chen, G. Micropattern-Controlled Cell Density and Its Effect on Gene Transfection of Mesenchymal Stem Cells. Adv. Healthc. Mater. 2022, 9, 2101978. [Google Scholar] [CrossRef]
- Huang, S.; Su, Q.; Hou, X.; Han, K.; Ma, S.; Xu, B.; Yang, Y. Influence of Colonies’ Morphological Cues on Cellular Uptake Capacity of Nanoparticles. Front. Bioeng. Biotechnol. 2022, 10, 922159. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, X.; Hu, X.; Kawazoe, N.; Yang, Y.; Chen, G. Influence of Cell Morphology on Mesenchymal Stem Cell Transfection. ACS Appl. Mater. Interfaces 2019, 11, 1932–1941. [Google Scholar] [CrossRef]
- Amores de Sousa, M.C.; Rodrigues, C.A.V.; Ferreira, I.A.F.; Diogo, M.M.; Linhardt, R.J.; Cabral, J.M.S.; Ferreira, F.C. Functionalization of Electrospun Nanofibers and Fiber Alignment Enhance Neural Stem Cell Proliferation and Neuronal Differentiation. Front. Bioeng. Biotechnol. 2020, 8, 580135. [Google Scholar] [CrossRef]
- Heinrich, M.A.; Alert, R.; LaChance, J.M.; Zajdel, T.J.; Košmrlj, A.; Cohen, D.J. Size-dependent patterns of cell proliferation and migration in freely-expanding epithelia. eLife 2020, 9, e58945. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Y.; Wang, X.; Yoshitomi, T.; Kawazoe, N.; Yang, Y.; Chen, G. Micropattern-controlled chirality of focal adhesions regulates the cytoskeletal arrangement and gene transfection of mesenchymal stem cells. Biomaterials 2021, 271, 120751. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Y.; Wang, X.; Kawazoe, N.; Yang, Y.; Chen, G. The varied influences of cell adhesion and spreading on gene transfection of mesenchymal stem cells on a micropatterned substrate. Acta Biomater. 2021, 125, 100–111. [Google Scholar] [CrossRef]
- Low, B.C.; Pan, C.Q.; Shivashankar, G.V.; Bershadsky, A.; Sudol, M.; Sheetz, M. YAP/TAZ as mechanosensors and mechanotransducers in regulating organ size and tumor growth. FEBS Lett. 2014, 588, 2663–2670. [Google Scholar] [CrossRef] [Green Version]
- Bittner, M.; Meltzer, P.; Chen, Y.; Jiang, Y.; Seftor, E.; Hendrix, M.; Radmacher, M.; Simon, R.; Yakhini, Z.; Ben-Dor, A.; et al. Molecular classification of cutaneous malignant melanoma by gene expression profiling. Nature 2000, 406, 536–540. [Google Scholar] [CrossRef] [Green Version]
- Wellbrock, C.; Rana, S.; Paterson, H.; Pickersgill, H.; Brummelkamp, T.; Marais, R. Oncogenic BRAF Regulates Melanoma Proliferation through the Lineage Specific Factor MITF. PLoS ONE 2008, 3, e2734. [Google Scholar] [CrossRef]
- Kwong, L.N.; Costello, J.C.; Liu, H.; Jiang, S.; Helms, T.L.; Langsdorf, A.E.; Jakubosky, D.; Genovese, G.; Muller, F.L.; Jeong, J.H.; et al. Oncogenic NRAS signaling differentially regulates survival and proliferation in melanoma. Nat. Med. 2012, 18, 1503–1510. [Google Scholar] [CrossRef] [Green Version]
- Nissan, M.H.; Pratilas, C.A.; Jones, A.M.; Ramirez, R.; Won, H.; Liu, C.; Tiwari, S.; Kong, L.; Hanrahan, A.J.; Yao, Z.; et al. Loss of NF1 in Cutaneous Melanoma Is Associated with RAS Activation and MEK Dependence. Cancer Res. 2014, 74, 2340–2350. [Google Scholar] [CrossRef] [Green Version]
- Horzum, U.; Ozdil, B.; Pesen-Okvur, D. Step-by-step quantitative analysis of focal adhesions. MethodsX 2014, 1, 56–59. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Chen, Z.; Hou, X.; Han, K.; Xu, B.; Zhang, M.; Ding, S.; Wang, Y.; Yang, Y. Promotion of Melanoma Cell Proliferation by Cyclic Straining through Regulatory Morphogenesis. Int. J. Mol. Sci. 2022, 23, 11884. https://doi.org/10.3390/ijms231911884
Huang S, Chen Z, Hou X, Han K, Xu B, Zhang M, Ding S, Wang Y, Yang Y. Promotion of Melanoma Cell Proliferation by Cyclic Straining through Regulatory Morphogenesis. International Journal of Molecular Sciences. 2022; 23(19):11884. https://doi.org/10.3390/ijms231911884
Chicago/Turabian StyleHuang, Siyuan, Zhu Chen, Xiaoqiang Hou, Kuankuan Han, Bingshe Xu, Miao Zhang, Shukai Ding, Yongtao Wang, and Yingjun Yang. 2022. "Promotion of Melanoma Cell Proliferation by Cyclic Straining through Regulatory Morphogenesis" International Journal of Molecular Sciences 23, no. 19: 11884. https://doi.org/10.3390/ijms231911884