
Anthocyanin Biosynthesis Induced by MYB Transcription Factors in Plants


College of Agriculture, Guangxi University, Nanning 530004, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(19), 11701; https://doi.org/10.3390/ijms231911701
Submission received: 16 September 2022
/
Revised: 28 September 2022
/
Accepted: 29 September 2022
/
Published: 2 October 2022
(This article belongs to the Special Issue Anthocyanins: Chemistry and Bioactivity 2.0)
Abstract
:Anthocyanins act as polyphenolic pigment that is ubiquitously found in plants. Anthocyanins play a role not only in health-promoting as an antioxidant, but also in protection against all kinds of abiotic and biotic stresses. Most recent studies have found that MYB transcription factors (MYB TFs) could positively or negatively regulate anthocyanin biosynthesis. Understanding the roles of MYB TFs is essential in elucidating how MYB TFs regulate the accumulation of anthocyanin. In the review, we summarized the signaling pathways medicated by MYB TFs during anthocyanin biosynthesis including jasmonic acid (JA) signaling pathway, cytokinins (CKs) signaling pathway, temperature-induced, light signal, 26S proteasome pathway, NAC TFs, and bHLH TFs. Moreover, structural and regulator genes induced by MYB TFs, target genes bound and activated or suppressed by MYB TFs, and crosstalk between MYB TFs and other proteins, were found to be vitally important in the regulation of anthocyanin biosynthesis. In this study, we focus on the recent knowledge concerning the regulator signaling and mechanism of MYB TFs on anthocyanin biosynthesis, covering the signaling pathway, genes expression, and target genes and protein expression.
1. Introduction
Anthocyanins act as natural water-soluble pigments which range from red to blue, existing as glycosides in combination with glucose or cellobiose molecules, which are widely distributed in the roots, leaves, flowers, fruits, and so on of many plants [1]. Anthocyanins are not only essential for plant performance, but also have beneficial effects on human health due to antioxidant, anti-mutagenic, and anti-carcinogenic capacities [2,3,4]. Therefore, how to improve the anthocyanins accumulation of plants has long been a focus of intense research.
The biosynthesis pathway of anthocyanins has been clearly clarified in plants [5]. Anthocyanin is biosynthesized by the phenylpropanoid pathway that is well known [5]. Briefly, the pathway starts with the condensation of one molecule of 4-coumaroyl-coenzyme A (CoA) and three molecules of malonyl-CoA, leading to naringenin chalcone [6]. This process is usually accompanied by CHS conduction, before the pathway diverges into side branches leading to different classes of flavonoids, including anthocyanins [7]. Anthocyanin accumulation is regulated by a series of structural genes [8]. Additionally, increasing studies indicate that many environmental factors affect anthocyanin biosynthesis during the past few years, including nutrient deficiency (nitrogen, phosphorus, sugars, magnesium, and CaCl2), light stress, UV-B irradiation, low temperature, wounding stress, the invasion of pathogenic bacteria, etc. [9,10,11,12,13,14,15]. Importantly, MYB transcription factors (TFs) are involved in the regulation of the anthocyanin biosynthesis pathway that has been identified. For example, in grapevine, VvMYBA1 and VvMYBA2 are identified that regulate the biosynthesis of anthocyanin [16]. In Arabidopsis, R2R3-MYB TFs including PAP1, PAP2, MYB113, and MYB114 as positive regulators are involved in the biosynthetic pathway of anthocyanin [17,18]. In apples, MdMYB1 has been shown to be responsible for controlling anthocyanin biosynthesis [19,20,21]. Continued research has shown that other TF families are involved in anthocyanin synthesis, including the MADS, NAC, HD-Zip, and ERF TFs, indicating that there are still many mechanisms of anthocyanin synthesis and regulation that are yet to be elucidated [22,23,24,25].
Most R2R3 MYB TFs involved in the control of anthocyanin biosynthesis are activators that enhance the expression of biosynthetic pathway genes, such as AtMYB75 (or PAP1), AtMYB90 (or PAP2), AtMYB113 and AtMYB114 in Arabidopsis [18,26], MdMYBA, MdMYB1, MdMYB10 and MdMYB110a in apple [21,27], SlMYB12 in tomato [28], and StAN1 in potato [29,30]. In addition to MYB activators, it has also been reported that MYB TFs can repress anthocyanin accumulation. There are two distinct classes of MYB TFs which negatively regulate anthocyanin accumulation: R3-MYB and R2R3-MYB repressors, which have one or two repeats of the MYB domain region, respectively. These R2R3 MYB repressors include AtMYB3/4/6 in Arabidopsis [31], MdMYB16/17/111 in apple [32], FaMYB1 in strawberry [33] and PhMYB4 and PhMYB17 in petunia [34]. The R3 MYB repressors have been reported in several plant species, including CAPRICE (CPC), TRIPTYCHON (TRY), and AtMYBL2 in Arabidopsis.
In view of the increasing interest in studies of MYB TFs in anthocyanin biosynthesis, this paper provides a critical review of the most recent work in the literature regarding various mechanisms associated with the anthocyanin biosynthesis of plants in which MYB TFs play a role. In addition, for genes/proteins, the expression of which is mediated by MYB TFs during the anthocyanin biosynthesis, are also briefly summarized. Therefore, this review may provide new ideas for future researchers studying the regulatory role of MYB TFs in the anthocyanin biosynthesis pathway. Additionally, it was an objective of this study to describe in detail the regulatory mechanisms that are relevant to the anthocyanin accumulation of plants in which MYB TFs are involved and further accelerate research into the use of MYB TFs in identifying new varieties and breeding new cultivars.
2. Effect of MYB TFs on the Anthocyanin Biosynthesis
2.1. MYB TFs Positively Regulate Anthocyanin Biosynthesis
It was reported that the MdMYB10 transcription factor had potential to enhance anthocyanin biosynthesis in apple fruit [21]; Table 1. Ban et al. [20] found that MdMYBA was responsible for controlling anthocyanin biosynthesis in apple skin. A novel MYB transcription factor was designated as MdMYB3 and found that it regulated the anthocyanin accumulation of apple fruits skin [35]. MYB110a, a paralog of MYB10, was responsible for red fruit-flesh phenotypes of apple [27]. Overexpression of MdMYB9 or MdMYB11 improved anthocyanin accumulation in apple calluses, suggesting that MdMYB9 or MdMYB11 acted as an important regulatory factor in anthocyanin biosynthesis [36]. Hu et al. [37] found that MdMYB1 controlled the positively anthocyanin accumulation in apples. MdMYB16 was also an important part of the regulatory network controlling the anthocyanin biosynthetic pathway in apple fruits [38]. The overexpression of MdMYBDL1 enhanced anthocyanin accumulation in apple calli, verifying that MdMYBDL1 functioned as a positive regulator of anthocyanin biosynthesis [39] (Table 1). Wang et al. [40] revealed that overexpressed MdMYB24L resulted in higher anthocyanin contents in the transgenic apple calli than in the wild-type control calli, manifesting that MdMYB24L helped the anthocyanin accumulation of apple as a positive factor. A novel Myb-related sequence, VlmybA1-3, was isolated from grape berry [16]. They also further confirmed the gene-regulated anthocyanin biosynthesis in grape berry skin as a positive factor [16]. Similarly, VvMYB5b isolated from grape berry belonged to the R2R3-MYB transcription factor [41]. Overexpression of VvMYB5b in tobacco resulted in the accumulation of anthocyanin, suggesting that the transcriptional mechanisms of VvMYB5b were closely linked to the regulation of the anthocyanin biosynthesis throughout grape berry development [41]. Myb factor, VvMybPA1 and VvMybPA2, promoted anthocyanin biosynthesis in grapevine indicated by Bogs et al. [42] and Terrier et al. [43]. Arabidopsis MYBL2, which encoded an R3-MYB-related protein, positively regulated anthocyanin biosynthesis reported by Dubos et al. [8]; Table 1. Overexpression of MYB112 positively affected the relative expression of key transcription factors of anthocyanin biosynthesis in Arabidopsis, verifying that MYB112 as a positive regulator promoted anthocyanin accumulation [44]. It has been reported that the encoded AN2, an R2R3-MYB transcription factor, was an important regulatory transcription factor in anthocyanin biosynthesis in potato tuber skin by regulating coordinately the expression of multiple anthocyanin biosynthetic genes [29]. Ballester et al. [28] demonstrated that the MYB12 transcription factor played a key role in regulating the anthocyanin biosynthesis in tomato fruit. SlMYB75-OE tomatoes improved anthocyanin content as a key regulator [45]. Yan et al. [46] indicated that SlAN2-like was responsible for anthocyanin biosynthesis in tomato fruits through the CRISPR/Cas9 approach. DEEP PURPLE (DPL) and PURPLE HAZE (PHZ) encoded members of the R2R3-MYB transcription factor family that regulated positively anthocyanin biosynthesis in petunia, and controlled anthocyanin production in vegetative tissues and contributed to floral pigmentation [34]; Table 1. The identified R2R3 MYB transcription factor PavMYB10.1 was involved in the anthocyanin biosynthesis pathway and determined the fruit skin colour in sweet cherry [47]. The isolated R2R3 MYB transcription factor was designated as DcMYB6 from a purple carrot cultivar and overexpression of DcMYB6 in Arabidopsis resulted in enhancing anthocyanin accumulation in both vegetative and reproductive tissues, revealing that DcMYB6 was involved in regulating anthocyanin biosynthesis in purple carrots [48]. Knockout of DcMYB7 in carrots with purple roots resulted in carrots with yellow roots using the CRISPR/Cas9 system; DcMYB7 promoted anthocyanin accumulation in carrot roots [49]. Overexpression of LcMYB5 resulted in enhanced biosynthesis of anthocyanins in tobacco and petunia, indicating that LcMYB5 was an R2R3 transcriptional factor which positively regulated anthocyanin biosynthesis [50] (Table 1). Anthocyanin content was increased when R2R3 MYB transcription factor MYB10 or MYB110 was over-expressed, suggesting that kiwifruit anthocyanin biosynthesis was dependent on characterized MYB transcription factors [51]. Over-expressed LrAN2 and LbAN2 induced anthocyanin accumulation in tobacco, indicating that LrAN2 and LbAN2 were closely related to anthocyanin biosynthesis Lycium ruthenicum and Lycium barbarum, respectively [52]. Overexpression of PsMYB114L and PsMYB12L exhibited significantly higher accumulation of anthocyanins in transgenic Arabidopsis plants, resulting in purple-red leaves. Over-expressed PsMYB114L and PsMYB12L in transgenic apple calli significantly increased the anthocyanins content and led to a change in the callus color to red. These results implied that PsMYB114L and PsMYB12L as positive regulators enhanced anthocyanins accumulation in penoy [53]. Wang et al. [54] indicated that PdMYB118 functions as an essential transcription factor regulating anthocyanin biosynthesis in poplar. The MYB6 overexpressing in transgenic poplar significantly enhanced accumulation of anthocyanin, demonstrating that MYB6 regulated anthocyanin biosynthesis as a positive factor [55]. PaMYB10 overexpression was consistent with the accumulation of anthocyanin in apricot [56]. The isolated RsMYB1, which encoded an R2R3-MYB transcription factor, accumulated high contents of anthocyanins in radish [57] (Table 1). Kim et al. [58] isolated an allele from RsMYB which was named RsMYB1Short, and found that RsMYB1Short failed to promote anthocyanin accumulation. CmMYB6 was proposed as a novel activator in Chrysanthemum anthocyanin biosynthesis [59]. GmMYB10 positively regulated anthocyanin biosynthesis in both purple kale and harvested purple kale [60]. LrMYB15 was involved in the positive regulation of anthocyanins in lily by stimulating transcription of anthocyanin biosynthesis genes [61]. LvMYB5-silencing in VIGS experiments significantly reduced anthocyanins accumulation in lily petals [62]. AaMYB2 positively related to anthocyanin biosynthesis in Anthurium spathes [63]. A R2R3 MYB transcription factor named BoPAP1 may play an important role in activating the anthocyanin accumulation in the purple kale [64]. Ectopic expression of EsAN2 in tobacco significantly enhanced the anthocyanin biosynthesis and accumulation, both in leaves and flowers, EsAN2 may regulate positively anthocyanin biosynthesis in Epimedium sagittatum [65]. AgMYB2 contained highly conserved R2R3 domain and two anthocyanin characteristic motifs, ANDV motif and KPRPR[S/T]F motif, and revealed that AgMYB2 could enhance anthocyanin biosynthesis and accumulation in celery [66]. The LAP1 MYB transcription factor induced a massive accumulation of anthocyanin pigments and the expression level of UGT78G1 that increased anthocyanin accumulation was strongly up-regulated by LAP1 in medicago during this process [67]. Anthocyanin production was positively regulated by MYB transcription factor VlmybA2 in tobacco and Arabidopsis [68]. The gene PcMYB10, encoding an R2R3 MYB transcription factor, was involved in the anthocyanin biosynthetic pathway regulation of tobacco and Arabidopsis [69] (Table 1). Transient overexpression of CsMYB33 and CsMYB78 activated anthocyanin biosynthesis in the leaves of Nicotiana benthamiana [70]. Over-expressed BoMYB increased the anthocyanidins accumulation in transgenic Arabidopsis, implying that BoMYB positively regulated the synthesis of anthocyanins in kale [71]. RNAi repression of AcMYB123 significantly decreased anthocyanin biosynthesis in kiwifruit [72] (Table 1). The higher expression level of NsMYB1 may cause higher anthocyanin accumulation in the black fruit in Nitraria sibirica Pall, suggesting that NsMYB1 positively induced anthocyanin accumulation [73]. The coloration of purple leaves in Dendrobium bigibbum was associated with MYB2, and transient overexpression of MYB2 significantly increased anthocyanin accumulation in tobacco [74] (Table 1).

{kind=link}
{kind=link}
Table 1.
Summary of MYB TFs that regulate anthocyanin biosynthesis in plants.
| Species | MYB TFs | Effect | References |
|---|---|---|---|
| Apple | MdMYB10 | Positive | [21] |
| Apple | MdMYBA | Positive | [20] |
| Apple | MdMYB3 | Positive | [35] |
| Apple | MYB110a | Positive | [27] |
| Apple | MdMYB9 MdMYB11 | Positive | [36] |
| Apple | MdMYB1 | Positive | [37] |
| Apple | MdMYB16 | Positive | [38] |
| Apple | MdMYBDL1 | Positive | [39] |
| Apple | MdMYB24L | Positive | [40] |
| Grape berry | VlmybA1-3 | Positive | [16] |
| Grape berry | VvMYB5b | Positive | [41] |
| Grapevine | VvMYBPA1 VvMYBPA2 | Positive | [42,43] |
| Arabidopsis | MYBl2 | Positive | [8] |
| Arabidopsis | MYB112 | Positive | [44] |
| Potato | AN2 | Positive | [29] |
| Tomato | MYB12 | Positive | [28] |
| Tomato | SlMYB75 | Positive | [45] |
| Tomato | SlAN2-like | Positive | [46] |
| Petunia | DPL PHZ | Positive | [34] |
| Sweet cherry | PavMYB10.1 | Positive | [47] |
| Carrot | DcMYB6 | Positive | [48] |
| Carrot | DcMYB7 | Positive | [49] |
| Petunia | LcMYB5 | Positive | [50] |
| Kiwifruit | MYB10 MYB110 | Positive | [51] |
| Wolfberry | LrAN2 LbAN2 | Positive | [52] |
| Penoy | PsMYB114L PsMYB12L | Positive | [53] |
| Poplar | PdMYB118 | Positive | [54] |
| Poplar | MYB6 | Positive | [55] |
| Apricot | PaMYB10 | Positive | [56] |
| Radish | RsMYB1 | Positive | [57] |
| Chrysanthemums | CmMYB6 | Positive | [59] |
| Purple kale | GmMYB10 | Positive | [60] |
| Lily | LrMYB15 | Positive | [61] |
| Lily | LcMYB5 | Positive | [62] |
| Anthurium andraeanu | AaMYB2 | Positive | [63] |
| Purple kale | BoPAP1 | Positive | [64] |
| Epimedium sagittatum | EsAN2 | Positive | [65] |
| Celery | AgMYB2 | Positive | [66] |
| Medicago | LAP1 | Positive | [67] |
| Tobacco Arabidopsis | VlmybA2 | Positive | [68] |
| Tobacco Arabidopsis | PcMYB10 | Positive | [69] |
| Mariguana | CsMYB33 CsMYB78 | Positive | [70] |
| Kale | BoMYB | Positive | [71] |
| Kiwifruit | AcMYB123 | Positive | [72] |
| Nitraria sibirica Pall | NsMYB1 | Positive | [73] |
| Dendrobium bigibbum | MYB2 | Positive | [74] |
| Arabidopsis | AtMYBL2 | Negative | [75] |
| Arabidopsis | CPC | Negative | [76] |
| Poplar | MYB182 | Negative | [77] |
| Petunia | MYBx1 | Negative | [78] |
| Tartary buckwheat | FtMYB8 | Negative | [79] |
| Apple | MdMYBL2 | Negative | [80] |
| Chrysanthemum | CmMYB#7 | Negative | [81] |
| Lily | LhR3MYB1 LhR3MYB2 | Negative | [82] |
| Lily | LcMYB1 | Negative | [62] |
| Grapevine | VvMYBC2L2 | Negative | [83] |
| Arabidopsis | MdMYB6 | Negative | [84] |
| Celery | OjMYB1 | Negative | [85] |
| Lettuce | AtMYB60 | Negative | [86] |
| Tobacco | GtMYB1R1 GtMYB1R9 | Negative | [87] |
| Tobacco | AtCPC | Negative | [88] |
| Gerbera | GhMYB1a | Negative | [89] |
| Grape hyacinth | MaMYBx | Negative | [90] |
| Chinese cabbage | BrMYBL2.1 | Negative | [91] |
2.2. MYB TFs Negatively Regulate Anthocyanin Biosynthesis
In Arabidopsis, the identified R3-MYB transcription factor AtMYBL2 acted as a transcriptional repressor and its repressive activity was attributable to the carboxy-terminal region of six amino acids, TLLLFR [75] (Table 1). They also found that AtMYBL2 negatively regulated the biosynthesis of anthocyanin in Arabidopsis [75]. CPC was an R3-MYB transcription factor and found that anthocyanin synthesis genes were significantly downregulated in the 35S:CPC overexpression of Arabidopsis, implying that Arabidopsis was identified as a negative regulator in anthocyanin biosynthesis of Arabidopsis [76]. Overexpression of MYB182 in poplar resulted in reducing anthocyanin levels, indicating that MYB182 as a negative regulator inhibited anthocyanin biosynthesis [77] (Table 1). MYBx1 can negatively regulate anthocyanin production and influence floral pigmentation by repressing the activity of the MBW complex in petunia [78]. Overexpression of FtMYB8 reduced the accumulation of anthocyanin in Arabidopsis, implying that FtMYB8 negatively regulated anthocyanin biosynthesis in Tartary buckwheat [79]. The MdMYBL2-overexpressing callus exhibited lower anthocyanin contents than of the control, suggesting that MdMYBL2 functioned as a negative regulator of anthocyanin biosynthesis in apple [80]. Xiang et al. [81] illustrated that CmMYB#7, an R3 MYB transcription factor, was a negative regulator of anthocyanin biosynthesis in chrysanthemum. Two R3-MYBs, LhR3MYB1 and LhR3MYB2, which were identified and found that they had a C2 suppressor motif downstream of a single MYB repeat. Meanwhile, both stable and transient overexpressed LhR3MYB1 and LhR3MYB2 showed inhibition of anthocyanin biosynthesis in tobacco plants. Thus, LhR3MYB1 and LhR3MYB2 suppressed anthocyanin biosynthesis in lily as a passive factor [82] (Table 1). LvMYB1, an R2R3-MYB transcription factor, inhibited anthocyanin biosynthesis in lily flowers [62]. The isolated VvMYBC2L2 from Rose, an R2R3-MYB transcription factor, was involved in the regulation of anthocyanin biosynthesis in grapevine as a transcriptional repressor [83]. MdMYB6 over-expression inhibited the activities of an anthocyanin biosynthesis-related enzyme and the expression levels of some bHLH genes, indicating that the MdMYB6 transcription factor is an important repressor of anthocyanin biosynthesis in Arabidopsis [84] (Table 1). OjMYB1, an encoding R2R3-MYB transcription factor, could enhance the anthocyanins content in Arabidopsis [85]. The identified AtMYB60 suppressed anthocyanin biosynthesis in the lettuce plant [86]. GtMYB1R1 and GtMYB1R9 acted as antagonistic transcription factors of anthocyanin biosynthesis in gentian flowers [87] (Table 1). MYB-type transcription factor AtCPC acted as a repressor of anthocyanin production [88] (Table 1). In gerbera and tobacco, overexpressed GhMYB1a significantly decreased anthocyanin content [89]. MaMYBx, an R3 MYB transcription factor, acted as a repressor of anthocyanin accumulation in grape hyacinth [90]. R3 MYB transcription factor BrMYBL2.1 suppressed anthocyanin biosynthesis in Chinese cabbage by blocking MBW complex activity [91].
As mentioned above, many MYB TFs positively regulated anthocyanin biosynthesis in plants including Apple, Grape berry, Arabidopsis, Potato, Tomato, etc, which belong to R2R3 and R3 MYB TFs. MYB TFs that negatively regulated anthocyanin synthesis in plants including Arabidopsis, Poplar, Apple, and so on have also been identified. They usually belong to R3 MYB TFs, but some of them belong to R2R3 MYB TFs. To explore the roles of MYB TFs in plants, some methods are implemented, such as overexpression, RNAi, CRISPR/Cas9, and VIGS. So far, it has been found that the MYB TFs regulating apple anthocyanins accumulation was more than those regulating other plants. Further research is required to investigate the role of MYB TFs in regulating anthocyanin biosynthesis in more plants.
3. The Signaling Pathway Regulated by MYB TFs during Anthocyanin Biosynthesis
3.1. Jasmonic Acid (JA) Signaling Pathway
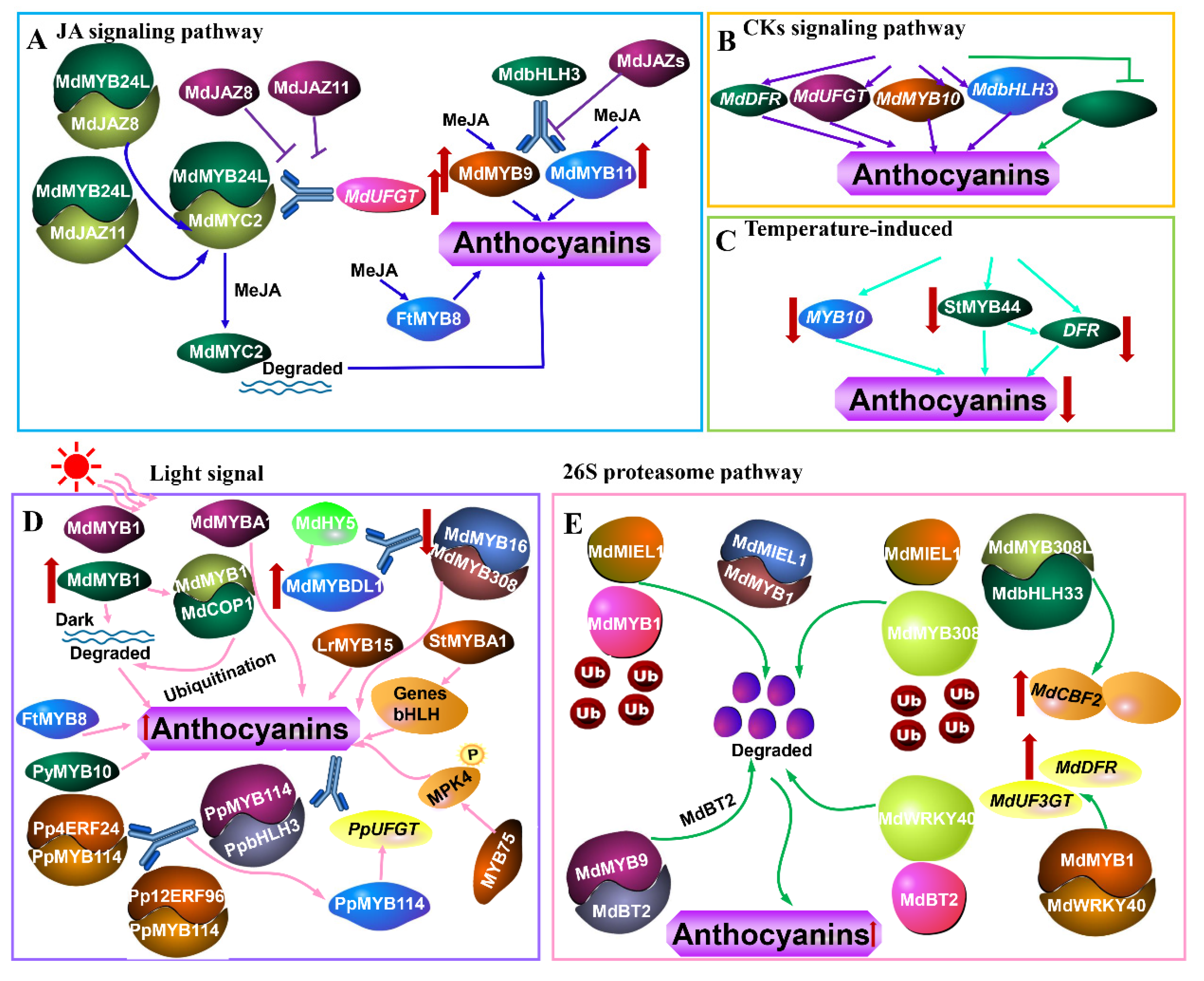
The encoded nuclear protein, MdMYB24L, interacted with JA signaling factors including MdJAZ8, MdJAZ11, and MdMYC2 that were observed [40] (Figure 1A). The MdJAZ8 and MdJAZ11 directly targeted MdMYC2 protein, which are rapidly degraded in the jasmonic acid methyl ester (MeJA) treatment. The interaction between MdMYB24L and MdMYC2 further increased the transcriptional level of MdUFGT; nevertheless, this effect was weakened by MdJAZ8 and MdJAZ11 [40]. Overexpression of MdMYB9 or MdMYB11 promoted anthocyanin accumulation in apple calluses, and the accumulation was further enhanced by MeJA. MdbHLH3 was recruited to the promoters of MdMYB9 and MdMYB11 and regulated their transcription, but MdJAZs inhibited the recruitment of MdbHLH3 to the promoters of MdMYB9 and MdMYB11 [36] (Figure 1A). The expression of FtMYB8 was markedly induced by JA during anthocyanin biosynthesis [79] (Figure 1A).
3.2. Cytokinins (CKs) Signaling Pathway
CKs are one of the major plant hormones that is widely distributed in various tissues of plants [92]. There are two types of CKs: adenine-type CKs represented by kinetin, zeatin, and 6-benzylaminopurine (6-BA), and phenylurea-type CKs such as diphenylurea and thidiazuron (TDZ) [92]. It was well known that CKs induced anthocyanin biosynthesis, and anthocyanin is also regulated by MYB transcription factors [80,92]. CKs treatments (6-BA) upregulated the relative expression of MdDFR, MdUFGT, MdMYB10, and MdbHLH3 genes [80] (Figure 1B). Wang et al. [80] also observed that MdMYBL2 was a negative factor in anthocyanin biosynthesis and was inhibited by CKs treatments, suggesting that MdMYBL2 influenced the CKs-regulated anthocyanin biosynthesis in red-fleshed apples.
3.3. Temperature-Induced
Lin-wang et al. [32] showed that the high temperature significantly down-regulated the expression of MYB10, which was responsible for reductions in anthocyanin accumulation. StMYB44 inhibited anthocyanin accumulation as a negative regulator by suppressing the activity of the DFR promoter [93] (Figure 1C). The high temperature reduced anthocyanin accumulation in potato flesh by enhancing the expression of StMYB44 and down-regulating the activation of the DFR promoter activity [93].
3.4. Light Signal
MdMYB1 and MdMYBA were important regulators of light-induced anthocyanin biosynthesis in apple fruits [19,20]. Under the light condition, MdMYB1 was accumulated, whereas it was degraded via a ubiquitin-dependent pathway in the dark. MdMYB1 interacted with nuclear MdCOP1proteins and MdCOP1 proteins were responsive to the ubiquitination and degradation of MdMYB1 protein in the dark, and they were involved in the light-controlled stability of the MdMYB1 protein [94] (Figure 1D). Liu et al. [39] indicated that MdHY5 bound to the G-box element of the MdMYBDL1 promoter to activate its expression. MdMYB16 can form a dimer with MdMYB308, which negatively regulated anthocyanin biosynthesis. Furthermore, MdMYBDL1 and MdHY5 suppressed the promoter activities of MdMYB16 and MdMYB308. These results implied that MdHY5 responded to light signals and was upstream of various kinds of MYB transcription factors in the regulation of anthocyanin accumulation in apples [39]. LrMYB15 transcription ceased completely when plants were kept in shaded conditions and the colors of the flower buds faded, indicating the regulated light-induced anthocyanin accumulation [61]. Applying the dark conditions to the tobacco plants overexpressing StMYBA1 transiently can down-regulate the expression levels of biosynthetic pathway genes and bHLH transcription factors, and subsequently reduced anthocyanin accumulation. The results suggested that StMYBA1 can positively regulate anthocyanin biosynthesis in tobacco, and light was required for its function on anthocyanin accumulation [95] (Figure 1D). PyMYB10 had core sequences of cis-acting regulatory elements involved in light responsiveness and showed that the expression of PyMYB10 was induced by light during anthocyanin biosynthesis [96]. Dark treatment repressed the expression of FtMYB8 during anthocyanin accumulation [79]. The transcript levels of PaMYB10, PaPAL, PaCHS, PaCHI, PaF3H, PaDFR, PaLDOX, and PaUFGT were inhibited by bagging treatment, resulting in the decrease in anthocyanin contents [56]. Interaction of Pp4ERF24 and Pp12ERF96 with PpMYB114 regulated light-induced anthocyanin biosynthesis by promoting the interaction between PpMYB114 and PpbHLH3 and enhancing the expression of PpMYB114-induced PpUFGT [97]. Li et al. [98] revealed that MPK4 phosphorylation of MYB75 increased its stability and was essential for light-induced anthocyanin accumulation.
3.5. 26S Proteasome Pathway
MdMIEL1 functioned as a ubiquitin 3 ligase to ubiquitinate MdMYB1 protein, followed by degradation through a 26S proteasome pathway, demonstrated that MdMIEL1 negatively regulated anthocyanin accumulation [99]. The interaction between MdMIEL1 and MdMYB1 proteins was verified during this process [99] (Figure 1E). MdMYB308L as a positive regulator of anthocyanin biosynthesis interacted with MdbHLH33 and enhanced its binding to the promoters of MdCBF2 and MdDFR. However, MdMIEL1 as a negative regulator was identified to be a MdMYB308L-interacting protein and promoted the ubiquitination degradation of MdMYB308L via a 26S proteasome pathway in the apple [100]. MdBT2 interacted with MdMYB9 and negatively regulated the abundance of MdMYB9 protein through the 26S proteasome pathway. The degradation of MdMYB9 by MdBT2 down-regulated the expression levels of MdMYB9-mediated anthocyanin-related genes and reduced the accumulation of anthocyanin, which functioned in an MdCUL3-independent pathway [101]. MdMYB1, a positive regulator of anthocyanin biosynthesis, interacted with MdWRKY40 and enhanced its binding to the promoters of MdDFR and MdUF3GT. However, MdBT2 interacted with MdWRKY40 and degraded its protein abundance by the 26S proteasome pathway [102] (Figure 1E).
3.6. NAC TFs
LcR1MYB1, as one R1-MYB type MYB, was identified to physically interact with LcNAC13 and reversed the effect of LcNAC13. LcNAC13 could bound anthocyanin structural genes LcCHS1/2, LcCHI, LcF3H, LcF3′H, LcDFR, and LcMYB1 and negatively regulated the expression of these genes. To sum up, LcNAC13 and LcR1MYB1 may act together to antagonistically regulate anthocyanin biosynthesis during litchi fruit ripening, which helps to provide new insights into the regulatory networks of anthocyanin biosynthesis [103].
3.7. bHLH TFs
Endogenous bHLH partners can be stimulated by overexpressing exogenous gene StMYBA1 in tobacco, and the elevated expression levels of bHLH partners were essential for anthocyanin production in plant tissues [95]. LcbHLH1 and LcbHLH3 were essential partners of LcMYB1 in regulating the anthocyanin production in tobacco and also probably in litchi [104].
4. Regulatory Mechanisms of MYB TFs in Anthocyanin Biosynthesis
4.1. Modulation of Gene Expression by MYB TFs
In transgenic tobaccos, overpressed-MdMYB3 had resulted in transcriptional activation of CHS, CHI, UFGT, and FLS genes, which exhibited increased anthocyanins accumulation in flowers [35] (Table 2). The expression levels of OsPAL, CHS, ANS, and MYB55 genes were up-regulated in the purple leaf (pl) mutant compared to the wild type, which showed the enhancement of anthocyanins contents in leaves of rice [105]. VvMYBC2L2 as a nuclear protein could remarkably down-regulate structural genes including CHS, DFR, LAR and UFGT and regulatory genes including AN1a and AN1b, resulting in the increase in anthocyanin content [83]. The expression of PyMYB10 in the pear skin was positively correlated with CHS, PAL, CHI, DFR, ANS, and F3H genes [96]. The expression levels of anthocyanin biosynthetic including CHS, CHI, F3H, F3′H, DFR, LDOX, and UGT78D2 in the transgenic Arabidopsis carrying AgMYB2 were significantly up-regulated [66] (Table 2). Feng et al. [85] found that OjMYB1 could up-regulate the expression levels of the structural genes-related anthocyanins biosynthesis including CHS, CHI, F3H, F3′H, DFR, LDOX, and UGT78D2 in Arabidopsis to enhance the anthocyanins biosynthesis. Overexpressing EsAN2 significantly upregulated the relative expression of CHS, CHI, F3H, F3′H, FLS, DFR, ANS, An1a, and An1b genes in tobacco [65]. RsMYB1 could up-regulate the relative expression of anthocyanin biosynthetic-related genes F3H, DFR, and ANS involved in anthocyanin production [57]. AaMYB2 appeared to regulate the expression of F3H, ANS, CHS genes and is possibly involved in anthocyanins biosynthesis [98]. Wang et al. [40] reported that the expression levels of the anthocyanin biosynthesis structural genes UFGT and DFR were up-regulated in the transgenic calli of over-pressed MdMYB24L (Table 2). The expression levels of DFR, UFGT, MYB10, and bHLH3 genes were strongly suppressed in the MdMYBL2-overexpressing callus, suggesting that MdMYBL2 negatively regulated anthocyanin accumulation in apple [80]. PsMYB114L and PsMYB12L both enhance anthocyanin accumulation by down-regulating the relative expression of FLS and ANR genes and up-regulating the relative expression of DFR and ANS genes [53]. DcMYB7 could activate expression of bHLH3 and structural genes in the anthocyanin biosynthetic pathway including CHS1, CHI1, F3H1, F3′H1, FR1, LDOX1, and UCGalT1 [49]. Overexpression of DcMYB6 in Arabidopsis upregulated the transcriptional level of CHS1, CHI1, F3H1, F3′H1, DFR, LDOX1, and UGT78D2 genes [48] (Table 2). Expression of CHS1, CHI1, F3H, F3′H, F3′5′H, DFR2, and ANS1 was improved in poplar during the transient over-expressed expression of PdMYB118 [54]. NsMYB1 could promote the transcript of PAL, C4H, 4CL, F3H, F3′H, DFR, and ANS of black fruit in Nitraria sibirica Pall, and induce the anthocyanin accumulation in tobacco [73]. LvMYB5-silencing significantly up-regulated the expression levels of CHS, DFR, and ANS genes [62] (Table 2). Anthocyanin biosynthesis genes DFR, ANS, and CHS were significantly up-regulated in the transient MYB2-overexpressing [74]. Transient overexpression of GhMYB1a resulted in the increase in MYB1, CHS, F3H, F3′H, DFR, ANS, FLS, UFGT, MYB10, and MYC1 genes [89]. SlAN2 acted as an activator of the expression of structural genes including PAL, C4H, 4CL, CHS1, CHS2, CHI, CHI-like, F3H, F3′5′H, DFR, ANS, 3GT, RT, AAC, PAT, GST, and regulator genes including AN1, AN11, MYBATV, MYBATV-like, TRY, MYB76, and positively regulated the anthocyanins accumulation [46]. Overexpression of MaMYBx significantly repressed the relative expression of C4H, 4CL, F3′5′H, FLS, DFR, ANS, AN2, AN1a, and AN1b in tobacco [90] (Table 2).
4.2. DNA Binding of MYB TFs
MdMYBA bound specifically to the MdANS promoter region, which increased anthocyanin accumulation in the apple [20] (Table 3). AtMYBL2 bound directly to the TT8 protein, and this complex suppressed the expression of DFR and TT8 during anthocyanin pathway [75] (Table 3). MdMYB24L positively regulated anthocyanin biosynthesis via directly binding to the MYB-binding site motifs in the promoters of MdDFR and MdUFGT and activating its transcriptional [40]. SlMYB75 as a positive regulator of anthocyanin biosynthesis was able to directly bind to the MYBPLANT and MYBPZM cis-regulatory elements and to activate the promoters of the LOXC, AADC2, and TPS genes [45] (Table 3). CmMYB#7 competed with CmMYB6, which together with CmbHLH2 was an essential component of the anthocyanin activation complex, for interaction with CmbHLH2 through the bHLH binding site in the R3 MYB domain. This reduced binding of the CmMYB6–CmbHLH2 complex inhibited its ability to activate CmDFR and CmUFGT promoters [81]. DcMYB7 could activate directly DcUCGXT1 and DcSAT1 to regulate the glycosylation and acylation of anthocyanins [48]. MdMYB16 and LESMdMYB16 interacted the promoters of MdANS and MdUFGT yeast, respectively [38] (Table 3). PdMYB118 can directly activate the promoters of CHS1, DRF2, and ANS1 genes which functioned as an essential transcription factor regulating anthocyanin biosynthesis in poplar [54] (Table 3). AcMYB123 and AcbHLH42 were involved in the regulation of anthocyanin biosynthesis by activating promoters of AcANS and AcF3GT1 genes [72]. Nakatsuka et al. [106] reported that co-expression of GtMYB3 and GtbHLH1 in transient expression assay could enhance the promoter activities of anthocyanin biosynthetic genes including CHS, F3′5′H, and 5AT in tobacco BY2 cells. GtDFR promoter was activated by the GtMYB3-GtbHLH1 complex [87] (Table 3). MYBA1, MYBA6.1, and MYBA7 promoted anthocyanin accumulation in grapevine hairy roots by activating the promoters of UFGT and 3AT but only MYBA1 inducing F3′5′H promoter [107]. PavMYB10.1 bound to the promoter of PavANS and PavUFGT, which was involved in the regulation of anthocyanin accumulation [47] (Table 3). LcMYB5 enhanced anthocyanin biosynthesis by directly activating the expression of DFR or by indirectly up-regulating the expressions of bHLH1 [50] (Table 3). CmMYB6 significantly activated the CmDFR promoter. The activity of CmDFR was further enhanced by the combination of CmMYB6 and MrbHLH1, leading to anthocyanin accumulation [59] (Table 3). LvMYB1 bound to the promoter of the LvANS gene, and enhanced its expression, thereby promoting anthocyanin synthesis [62]. GhMYB1 significantly activated the promoters of NtCHS and NtFLS genes, which are required for anthocyanin biosynthesis in gerbera [89]. MaMYBx bound MaMybA and MabHLH1 promoters and suppressed their expression, thereby reducing anthocyanin accumulation in grape hyacinth [90].
4.3. Interactions between MYB TFs and Other Proteins
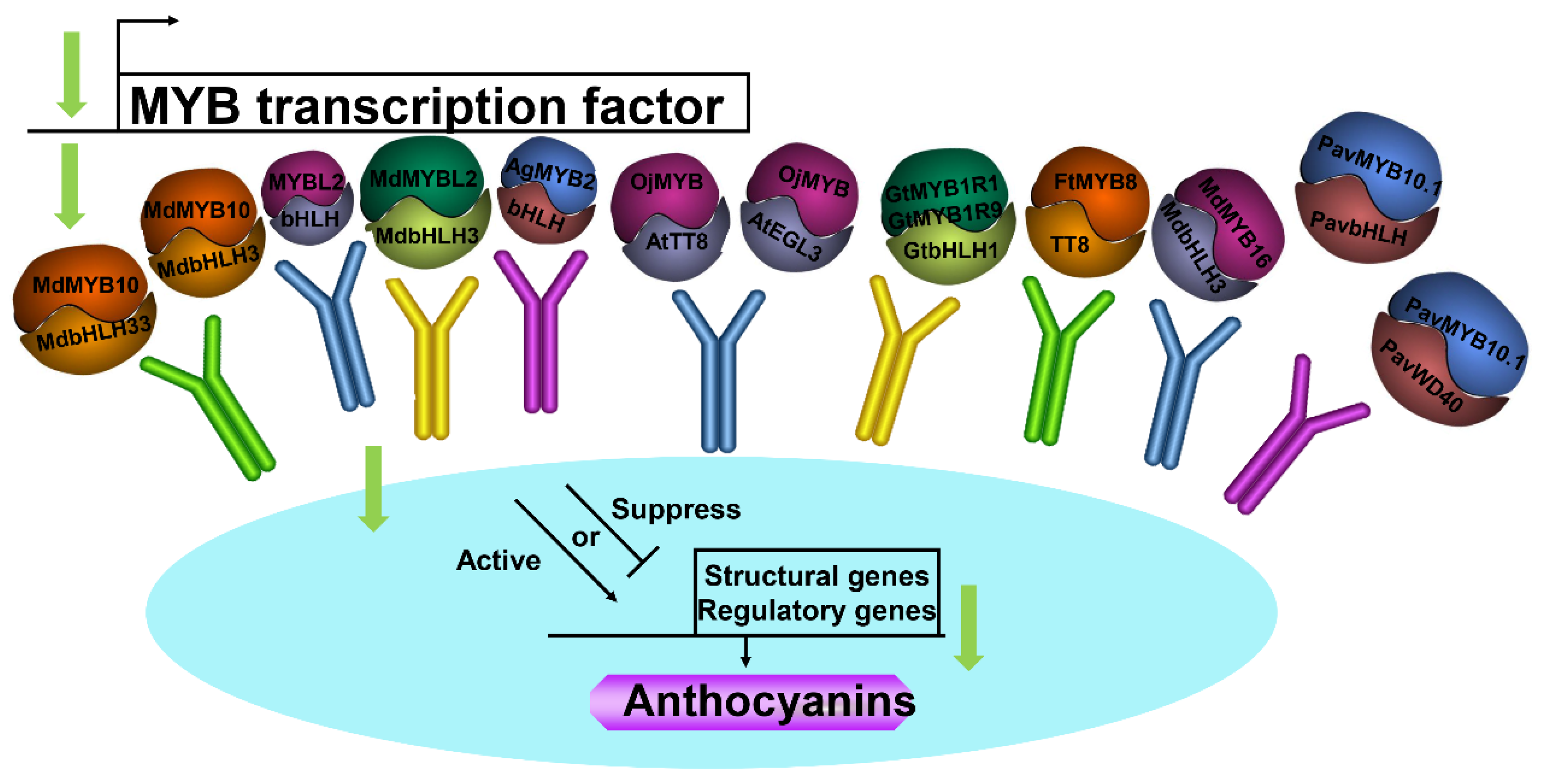
Efficient induction of anthocyanin biosynthesis in transient assays by MdMYB10 was dependent on the co-expression of two distinct bHLH proteins from the apple: MdbHLH3 and MdbHLH33 [21] (Figure 2). MYBL2 interacted with bHLH proteins and modulated the expression of anthocyanin biosynthesis-related genes, which influenced anthocyanin accumulation [8]. Wang et al. [80] indicated that MdMYBL2 interacted with MdbHLH3 in the apple and the interaction enhanced anthocyanin accumulation by regulating the expression of anthocyanin biosynthesis-related genes (Figure 2). Yeast two-hybrid assay confirmed that EsAN2 was capable of interacting with NtAn1a, NtAn1b, EsTT8, and AtTT8 regulators of the anthocyanin biosynthetic pathway [65] (Figure 2). The interaction between AgMYB2 and bHLH proteins was shown by yeast two-hybrid assay [66]. OjMYB1 could interact with AtTT8 and AtEGL3 proteins in yeast [85]. Nakatsuka et al. [87] demonstrated that both GtMYB1R1 and GtMYB1R9 proteins interacted with the GtbHLH1 protein, previously identified as an anthocyanin biosynthesis regulator in gentian flowers. FtMYB8 inhibited anthocyanin accumulation by inhibiting TT12 expression and interacting with TT8 [79]. MdMYB16 interacted with MdbHLH3, which may control the anthocyanin biosynthetic pathway [38]. PavMYB10.1 interacted with proteins PavbHLH and PavWD40 using yeast two-hybrid assays and chromatin immunoprecipitation assays during anthocyanin biosynthesis [47] (Figure 2).
5. Conclusions and Future Perspective
In conclusion, in plants, some MYB TFs could regulate, positively or negatively, anthocyanins accumulation. JA signaling pathway, CK signaling pathway, temperature-induced, light signal, 26S proteasome pathway, NAC TFs, and bHLH TFs are essential pathways for regulating the anthocyanin biosynthesis of plants. Additionally, we have also concluded that MYB TFs induced the expression of structural genes and regulator genes, bound and activated or suppressed anthocyanin biosynthesis-related target genes, and interacted with other proteins, which improved or inhibited anthocyanins accumulation in plants. These insights might be advantageous in identifying new varieties and breeding new cultivars.
Nowadays, although it is a well-known fact that MYB TFs-medicated signaling pathways regulate anthocyanins accumulation, the signaling pathways induced by MYB TFs involved in this process still needs to be further elucidated. Additionally, MYB TFs regulated structural genes and regulator genes, bound and activated or suppressed target genes, and interacted with other proteins during anthocyanin biosynthesis. Clearly, protein modifications induced by MYB TFs including S-nitrosylation, methylation, phosphorylation, and so on are lacking. More research works will improve knowledge concerning the regulator mechanism of MYB TFs in the anthocyanin biosynthesis pathway with the aim of identifying new varieties and breeding new cultivars.
Author Contributions
Conceptualization, C.L.; methodology, C.L. and W.Y.; formal analysis, C.L., W.Y., J.X., X.L. and Y.L.; writing—original draft preparation, C.L.; writing—review and editing, C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Nature Science Foundation of China (31660568), Guangxi science and technology major project (GuikeAA22068088) and start-up funding for introduced talents in Guangxi University (to C.L.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, B.C.; Tennessen, D.J.; Last, R.L. UVB induced photomorphogenesis in Arabidopsis thaliana. Plant J. 2002, 15, 667–674. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Giusti, M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The case for anthocyanin consumption to promote human health: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Dooner, H.K.; Robbins, T.P.; Jorgensen, R.A. Genetic and developmental control of anthocyanin biosynthesis. Annu. Rev. Genet. 1991, 25, 173–199. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Dubos, C.; Gourrierec, L.J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.M.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 2008, 55, 940–953. [Google Scholar] [CrossRef]
- Schaberg, P.G.; Van den Berg, A.K.; Murakami, P.F.; Shane, J.B.; Donnelly, J.R. Factors influencing red expression in autumn foliage of sugar maple trees. Tree Physiol. 2003, 23, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Kadir, S.A. Fruit quality at harvest of ″Jonathan″ apple treated with foliarly-applied calcium chloride. J. Plant Nutr. 2005, 27, 1991–2006. [Google Scholar] [CrossRef]
- Rubin, G.; Tohge, T.; Matsuda, F.; Saito, K.; Scheible, W.R. Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis. Plant Cell 2009, 21, 3567–3584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinilal, B.; Ovadia, R.; Nissim-Levi, A.; Perl, A.; Carmeli-Weissberg, M.; Oren-Shamir, M. Increased accumulation and decreased catabolism of anthocyanins in red grape cell suspension culture following magnesium treatment. Planta 2011, 234, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Saito, T.; Honda, C.; Hatsuyama, Y.; Ito, A.; Moriguchi, T. An apple B-box protein, MdCOL11, is involved in UV-B-and temperature-induced anthocyanin biosynthesis. Planta 2014, 240, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Li, H.; Xie, Y.; Wu, W.; Li, M.; Wang, X.; Huang, J. THF1 mutations lead to increased basal and wound-induced levels of oxylipins that stimulate anthocyanin biosynthesis via COI1 signaling in Arabidopsis. J. Integr. Plant Biol. 2014, 56, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Olsen, K.M.; Slimestad, R.; Lea, U.S.; Brede, C.; Lovdal, T.; Ruoff, P.; Verheul, M.; Lillo, C. Temperature and nitrogen effects on regulators and products of the flavonoid pathway: Experimental and kinetic model studies. Plant Cell Environ. 2009, 32, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Azuma, A.; Kobayashi, S.; Mitani, N.; Shiraishi, M.; Yamada, M.; Ueno, T.; Kono, A.; Yakushiji, H.; Koshita, Y. Genomic and genetic analysis of Myb-related genes that regulate anthocyanin biosynthesis in grape berry skin. Theor. Appl. Genet. 2008, 117, 1009–1019. [Google Scholar] [CrossRef]
- Borevitz, J.O.; Xia, Y.; Blount, J.; Dixon, R.A.; Lamb, C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant Cell 2000, 2, 2383–2394. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.; Zhao, M.Z.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Takos, A.M.; Jaffé, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [Green Version]
- Ban, Y.; Honda, C.; Hatsuyama, Y.; Igarashi, M.; Bessho, H.; Moriguchi, T. Isolation and functional analysis of a MYB transcription factor gene that is a key regulator for the development of red coloration in apple skin. Plant Cell Physiol. 2007, 48, 958–970. [Google Scholar] [CrossRef]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Wang, T.; McGie, T.; Voogd, C.; Allan, A.C.; Hellens, R.P.; Varkonyi-Gasic, E. Overexpression of the kiwifruit SVP3 gene affects reproductive development and suppresses anthocyanin biosynthesis in petals, but has no effect on vegetative growth, dormancy, or flowering time. J. Exp. Bot. 2014, 65, 4985–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Lin-Wang, K.; Wang, H.; Gu, C.; Dare, A.P.; Espley, R.V.; He, H.; Allan, A.C.; Han, Y. Molecular genetics of blood-fleshed peach reveals activation of anthocyanin biosynthesis by NAC transcription factors. Plant J. 2015, 82, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, C.; Yan, D.; Wen, X.H.; Liu, Y.L.; Wang, H.J.; Dai, J.Y.; Zhang, Y.J.; Liu, Y.F.; Zhou, B.; et al. MdHB1 down-regulation activates anthocyanin biosynthesis in the white-fleshed apple cultivar ‘Granny Smith’. J. Exp. Bot. 2017, 68, 1055–1069. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xu, H.F.; Wang, N.; Jiang, S.H.; Fang, H.C.; Zhang, Z.Y.; Yang, G.X.; Wang, Y.C.; Su, M.Y.; Xu, L.; et al. The ethylene response factor MdERF1B regulates anthocyanin and proanthocyanidin biosynthesis in apple. Plant Mol. Biol. 2018, 98, 205–218. [Google Scholar] [CrossRef]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Mehrtens, F.; Niehaus, K.; Weisshaar, B. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef] [Green Version]
- Chagné, D.; Lin-Wang, K.; Espley, R.V.; Volz, R.K.; How, N.M.; Rouse, S.; Brendolise, C.; Carlisle, C.M.; Kumar, S.; De Silva, N.; et al. An ancient duplication of apple MYB transcription factors is responsible for novel red fruit-flesh phenotypes. Plant Physiol. 2013, 161, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Ballester, A.R.; Molthoff, J.; de Vos, R.; Hekkert, B.; Orzaez, D.; Fernandez-Moreno, J.; Tripodi, P.; Grandillo, S.; Martin, C.; Heldens, J.; et al. Biochemical and molecular analysis of pink tomatoes: Deregulated expression of the gene encoding transcription factor SlMYB12 leads to pink tomato fruit color. Plant Physiol. 2010, 152, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.S.; Griffiths, H.M.; De Jong, D.M.; Cheng, S.; Bodis, M.; Kim, T.S.; De Jong, W.S. The potato developer (D) locus encodes an R2R3 MYB transcription factor that regulates expression of multiple anthocyanin structural genes in tuber skin. Theor. Appl. Genet. 2009, 120, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kui, L.W.; Espley, R.V.; Wang, L.; Yang, H.; Yu, B.; Andrew, D.; Erika, V.G.; Wang, J.; Zhang, J. Functional diversification of the potato R2R3 MYB anthocyanin activators AN1, MYBA1, and MYB113 and their interaction with basic helix-loop-helix cofactors. J. Exp. Bot. 2016, 67, 2159–2176. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin-Wang, K.; Micheletti, D.; Palmer, J.; Volz, R.; Lozano, L.; Espley, R.; Hellens, R.P.; Chagne, D.; Rowan, D.D.; Troggio, M.; et al. High temperature reduces apple fruit colour via modulation of the anthocyanin regulatory complex. Plant Cell Environ. 2011, 34, 1176–1190. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, A.; De Vos, C.H.; Wein, M.; Sun, Z.; Greco, R.; Kroon, A.; Mol, J.N.; O’Connell, A.P. The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco. Plant J. 2001, 28, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Albert, N.W.; Lewis, D.H.; Zhang, H.; Schwinn, K.E.; Jameson, P.E.; Davies, K.M. Members of an R2R3-MYB transcription factor family in Petunia are developmentally and environmentally regulated to control complex floral and vegetative pigmentation patterning. Plant J. 2011, 65, 771–784. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Han, Y.P.; Wei, G.C.; Korban, S.S. An apple MYB transcription factor, MdMYB3, is involved in regulation of anthocyanin biosynthesis and flower development. BMC Plant Biol. 2013, 13, 176. [Google Scholar] [CrossRef] [Green Version]
- An, X.H.; Tian, Y.; Chen, K.Q.; Liu, X.J.; Liu, D.D.; Xie, X.B.; Cheng, C.G.; Cong, P.H.; Hao, Y.J. MdMYB9 and MdMYB11 are involved in the regulation of the JA-induced biosynthesis of anthocyanin and proanthocyanidin in apples. Plant Cell Physiol. 2015, 56, 650–662. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.G.; Sun, C.H.; Ma, Q.J.; You, C.X.; Cheng, L.L.; Hao, Y.J. MdMYB1 regulates MdMYB1 regulates anthocyanin and malate accumulation by directly facilitating their transport into vacuoles in apples. Plant Physiol. 2016, 170, 1315–1330. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.F.; Wang, N.; Liu, J.X.; Qu, C.Z.; Wang, Y.C.; Jiang, S.H.; Lu, N.L.; Wang, D.Y.; Zhang, Z.Y.; Chen, X.S. The molecular mechanism underlying anthocyanin metabolism in apple using the MdMYB16 and MdbHLH33 genes. Plant Mol. Biol. 2017, 94, 149–165. [Google Scholar] [CrossRef]
- Liu, W.J.; Wang, Y.C.; Sun, J.J.; Jiang, H.Y.; Xu, H.F.; Wang, N.; Jiang, S.H.; Fang, H.C.; Zhang, Z.Y.; Wang, Y.L.; et al. MdMYBDL1 employed by MdHY5 increases anthocyanin accumulation via repression of MdMYB16/308 in apple. Plant Sci. 2019, 283, 32–40. [Google Scholar] [CrossRef]
- Wang, Y.C.; Liu, W.J.; Jiang, H.Y.; Mao, Z.L.; Wang, N.; Jiang, S.H.; Xu, H.F.; Yang, G.X.; Zhang, Z.Y.; Chen, X.S. The R2R3-MYB transcription factor MdMYB24-like is involved in methyl jasmonate-induced anthocyanin biosynthesis in apple. Plant Physiol. Biochem. 2019, 139, 273–282. [Google Scholar] [CrossRef]
- Deluc, L.; Bogs, J.; Walker, A.R.; Ferrier, T.; Decendit, A.; Merillon, J.M.; Robinson, S.P.; Barrieu, F. The transcription factor VvMYB5b contributes to the regulation of anthocyanin and proanthocyanidin biosynthesis in developing grape berries. Plant Physiol. 2008, 147, 2041–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrier, N.; Torregrosa, L.; Ageorges, A.; Vialet, S.; Verriès, C.; Cheynier, V.; Romieu, C. Ectopic expression of VvMybPA2 promotes proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant Physiol. 2009, 149, 1028–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotkowska, M.E.; Tohge, T.; Fernie, A.R.; Xue, G.P.; Balazadeh, S.; Mueller-Robeber, B. The Arabidopsis transcription factor MYB112 promotes anthocyanin formation during salinity and under high light stress. Plant Physiol. 2015, 169, 1862–1880. [Google Scholar] [CrossRef] [Green Version]
- Jian, W.; Cao, H.H.; Yuan, S.; Liu, Y.D.; Lu, J.F.; Lu, W.; Li, N.; Wang, J.H.; Zou, J.; Tang, N.; et al. SlMYB75, an MYB-type transcription factor, promotes anthocyanin accumulation and enhances volatile aroma production in tomato fruits. Hortic. Res. 2019, 1, 22. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.S.; Chen, N.; Huang, Z.J.; Li, D.J.; Zhi, J.J.; Yu, B.W.; Liu, X.X.; Cao, B.H.; Qiu, Z.K. Anthocyanin fruit encodes an R2R3-MYB transcription factor, SlAN2-like, activating the transcription of SlMYBATV to fine-tune anthocyanin content in tomato fruit. New Phytol. 2020, 225, 2048–2063. [Google Scholar] [CrossRef]
- Jin, W.M.; Wang, H.; Li, M.F.; Wang, J.; Yang, Y.; Zhang, X.M.; Yan, G.H.; Zhang, H.; Liu, J.S.; Zhang, K.C. The R2R3 MYB transcription factor PavMYB10.1 involves in anthocyanin biosynthesis and determines fruit skin colour in sweet cherry (Prunus avium L.). Plant Biotechnol. J. 2016, 14, 2120–2133. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.S.; Feng, K.; Que, F.; Wang, F.; Xiong, A.S. A MYB transcription factor, DcMYB6, is involved in regulating anthocyanin biosynthesis in purple carrot taproots. Sci. Rep. 2017, 7, 45324. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.S.; Yang, Q.Q.; Feng, K.; Xiong, A.S. Changing carrot color: Insertions in DcMYB7 alter the regulation of anthocyanin biosynthesis and modification. Plant Physiol. 2019, 181, 195–207. [Google Scholar] [CrossRef]
- Lai, B.; Du, L.N.; Hu, B.; Wang, D.; Huang, X.M.; Zhao, J.T.; Wang, H.C.; Hu, G.B. Characterization of a novel litchi R2R3-MYB transcription factor that involves in anthocyanin biosynthesis and tissue acidification. BMC Plant Biol. 2019, 19, 62. [Google Scholar] [CrossRef]
- Peng, Y.Y.; Lin-Wang, K.; Cooney, J.M.; Wang, T.C.; Espley, R.V.; Allan, A.C. Differential regulation of the anthocyanin profile in purple kiwifruit (Actinidia species). Hortic. Res. 2019, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Y.; Zhu, X.; Liu, Z.; Xi, X.; Liu, B. Functional MYB transcription factor encoding gene AN2 is associated with anthocyanin biosynthesis in Lycium ruthenicum Murray. BMC Plant Biol. 2019, 19, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.P.; Xu, Z.D.; Yu, X.Y.; Zhao, L.Y.; Zhao, M.Y.; Han, X.; Qi, S. Identification of two novel R2R3-MYB transcription factors, PsMYB114L and PsMYB12L, related to anthocyanin biosynthesis in Paeonia suffruticosa. Int. J. Mol. Sci. 2019, 20, 1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Wang, X.Q.; Song, W.M.; Bao, Y.; Jin, Y.L.; Jiang, C.M.; Wang, C.T.; Li, B.; Zhang, H.X. PdMYB118, isolated from a red leaf mutant of Populus deltoids, is a new transcription factor regulating anthocyanin biosynthesis in poplar. Plant Cell Rep. 2019, 38, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Lu, W.X.; Ran, L.Y.; Dou, L.W.; Yao, S.; Hu, J.; Fan, D.; Li, C.F.; Luo, K.M. R2R3-MYB transcription factor MYB6 promotes anthocyanin and proanthocyanidin biosynthesis but inhibits secondary cell wall formation in Populus tomentosa. Plant J. 2019, 99, 733–751. [Google Scholar] [CrossRef]
- Xi, W.P.; Feng, J.; Liu, Y.; Zhang, S.K.; Zhao, G.H. The R2R3-MYB transcription factor PaMYB10 is involved in anthocyanin biosynthesis in apricots and determines red blushed skin. BMC Plant Biol. 2019, 19, 287. [Google Scholar] [CrossRef]
- Lim, S.H.; Song, J.H.; Kim, D.H.; Kim, J.K.; Lee, J.Y.; Kim, Y.M.; Ha, S.H. Activation of anthocyanin biosynthesis by expression of the radish R2R3-MYB transcription factor gene RsMYB1. Plant Cell Rep. 2016, 35, 641–653. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, J.; Rhee, J.; Lee, J.Y.; Lim, S.H. Loss of the R2R3 MYB transcription factor RsMYB1 shapes anthocyanin biosynthesis and accumulation in Raphanus sativus. Int. J. Mol. Sci. 2021, 22, 10927. [Google Scholar] [CrossRef]
- Liu, X.F.; Xiang, L.L.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. The identification of a MYB transcription factor controlling anthocyanin biosynthesis regulation in Chrysanthemum flowers. Sci. Hortic. 2015, 194, 278–285. [Google Scholar] [CrossRef]
- Palapol, Y.; Ketsa, S.; Lin-Wang, K.; Ferguson, I.B.; Allan, A.C. A MYB transcription factor regulates anthocyanin biosynthesis in mangosteen (Garcinia mangostana L.) fruit during ripening. Planta 2009, 229, 1323–1334. [Google Scholar] [CrossRef]
- Yamagishi, M. A novel R2R3-MYB transcription factor regulates light-mediated floral and vegetative anthocyanin pigmentation patterns in Lilium regale. Mol. Breed. 2016, 36, 3. [Google Scholar] [CrossRef]
- Yin, X.J.; Zhang, Y.B.; Zhang, L.; Wang, B.H.; Zhao, Y.D.; Irfan, M.; Chen, L.J.; Feng, Y.L. Regulation of MYB transcription factors of anthocyanin synthesis in Lily flowers. Front. Plant Sci. 2021, 12, 761668. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Qiu, J.; Yang, G.S.; Huang, S.Y.; Yin, J.M. Isolation and characterization of a R2R3-MYB transcription factor gene related to anthocyanin biosynthesis in the spathes of Anthurium andraeanum (Hort.). Plant Cell Rep. 2016, 35, 2151–2165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Hu, Z.L.; Zhang, Y.J.; Li, Y.L.; Shuang, Z.; Chen, G.P. A putative functional MYB transcription factor induced by low temperature regulates anthocyanin biosynthesis in purple kale (Brassica Oleracea var. acephala f. tricolor). Plant Cell Rep. 2012, 31, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.J.; Khaldun, A.B.M.; Lv, H.Y.; Du, L.W.; Zhang, C.J.; Wang, Y. Isolation and functional characterization of a R2R3-MYB regulator of the anthocyanin biosynthetic pathway from Epimedium sagittatum. Plant Cell Rep. 2016, 35, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Liu, J.X.; Duan, A.Q.; Li, T.; Yang, Q.Q.; Xu, Z.S.; Xiong, A.S. AgMYB2 transcription factor is involved in the regulation of anthocyanin biosynthesis in purple celery (Apium graveolens L.). Planta 2018, 248, 1249–1261. [Google Scholar] [CrossRef]
- Peel, G.J.; Pang, Y.; Modolo, L.V.; Dixon, R.A. The LAP1 MYB transcription factor orchestrates anthocyanidin biosynthesis and glycosylation in Medicago. Plant J. 2009, 59, 136–149. [Google Scholar] [CrossRef]
- Geekiyanage, S.; Takase, T.; Ogura, Y.; Kiyosue, T. Anthocyanin production by over-expression of grape transcription factor gene VlmybA2 in transgenic tobacco and Arabidopsis. Plant Biotechnol. Rep. 2007, 1, 11–18. [Google Scholar] [CrossRef]
- Pierantoni, L.; Dondini, L.; Franceschi, P.D.; Musacci, S.; Winkel, B.S.J.; Sansavini, S. Mapping of an anthocyanin-regulating MYB transcription factor and its expression in red and green pear, Pyrus communis. Plant Physiol. Biochem. 2010, 48, 1020–1026. [Google Scholar] [CrossRef]
- Kundan, M.; Gani, U.; Fayaz, M.; Angmo, T.; Kesari, R.; Rahul, V.P.; Gairola, S.; Misra, P. Two R2R3-MYB transcription factors, CsMYB33 and CsMYB78 are involved in the regulation of anthocyanin biosynthesis in Cannabis sativa L. Ind. Crop. Prod. 2022, 188, 115546. [Google Scholar] [CrossRef]
- Wang, Y.S.; Wang, H.; Gao, M.L.; Fan, Z.Y.; Chen, Y.; Jin, Y.F. Overexpression of kale (Brassica oleracea L. var. acephala) BoMYB increases anthocyanin content in Arabidopsis thaliana. Biotechnol. Biotechnol. Equip. 2019, 33, 902–910. [Google Scholar]
- Wang, L.H.; Tang, W.; Hu, Y.W.; Zhang, Y.B.; Sun, J.Q.; Guo, X.H.; Lu, H.; Yang, Y.; Fang, C.B.; Niu, X.L.; et al. A MYB/bHLH complex regulates tissue-specific anthocyanin biosynthesis in the inner pericarp of red-centered kiwifruit Actinidia chinensis cv. Hongyang. Plant J. 2019, 99, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.M.; Zong, Y.; Hu, N.; Li, S.M.; Liu, B.L.; Wang, H.L. Functional R2R3-MYB transcription factor NsMYB1, regulating anthocyanin biosynthesis, was relative to the fruit color differentiation in Nitraria sibirica Pall. BMC Plant Biol. 2022, 22, 186. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.H.; Kim, S.W.; Ryu, J.; Kang, S.Y.; Kim, J.B.; Kim, S.H. Upregulation of the MYB2 transcription factor is associated with increased accumulation of anthocyanin in the leaves of Dendrobium bigibbum. Int. J. Mol. Sci. 2020, 21, 5653. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 2008, 55, 954–967. [Google Scholar] [CrossRef]
- Zhu, H.F.; Fitzsimmons, K.; Khandelwal, A.; Kranz, R.G. CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Mol. Plant 2009, 2, 790–802. [Google Scholar] [CrossRef]
- Yoshida, K.; Ma, D.; Constabel, C.P. The MYB182 protein down-regulates proanthocyanidin and anthocyanin biosynthesis in poplar by repressing both structural and regulatory flavonoid genes. Plant Physiol. 2015, 167, 693–710. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Wang, L.M.; Shang, H.Q.; Dong, X.Y.; Jiang, H.; Zhang, J.; Wang, H.J.; Li, Y.M.; Yuan, X.; Meng, S.Y.; et al. An R3-MYB gene of Phalaenopsis, MYBx1, represses anthocyanin accumulation. Plant Growth Regul. 2019, 88, 129–138. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, Q.; Wang, S.; Shi, J.; Dong, Q.; Yao, P.; Shi, G.; Xu, S.; Deng, R.; Li, C.; et al. FtMYB8 from Tartary buckwheat inhibits both anthocyanin/proanthocyanidin accumulation and marginal Trichome initiation. BMC Plant Biol. 2019, 19, 263. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Sun, J.; Wang, N.; Xu, H.; Qu, C.; Jiang, S.; Fang, H.; Su, M.; Zhang, Z.; Chen, X. MdMYBL2 helps regulate cytokinin-induced anthocyanin biosynthesis in red-fleshed apple (Malus sieversii f. niedzwetzkyana) callus. Funct. Plant Biol. 2019, 46, 187–196. [Google Scholar] [CrossRef]
- Xiang, L.; Liu, X.; Li, H.; Yin, X.; Grierson, D.; Li, F.; Chen, K. CmMYB# 7, an R3 MYB transcription factor, acts as a negative regulator of anthocyanin biosynthesis in chrysanthemum. J. Exp. Bot. 2019, 70, 3111–3123. [Google Scholar] [PubMed]
- Sakai, M.; Yamagishi, M.; Matsuyama, K. Repression of anthocyanin biosynthesis by R3-MYB transcription factors in lily (Lilium spp.). Plant Cell Rep. 2019, 38, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, G.; Liu, L.; Zhang, Q.T.; Han, Z.H.; Chen, X.S.; Li, B. A R2R3-MYB transcription factor, VvMYBC2L2, functions as a transcriptional repressor of anthocyanin biosynthesis in grapevine (Vitis vinifera L.). Molecules 2019, 24, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.J.; Shen, X.F.; Zhang, Z.; Peng, R.H.; Xiong, A.S.; Xu, J.; Zhu, B.; Yao, Z.Q.H. The MYB transcription factor MdMYB6 suppresses anthocyanin biosynthesis in transgenic Arabidopsis. Plant Cell Tissue Organ Cult. 2011, 106, 235–242. [Google Scholar] [CrossRef]
- Feng, K.; Xu, Z.S.; Que, F.; Liu, J.X.; Wang, F.; Xiong, A.S. An R2R3-MYB transcription factor, OjMYB1, functions in anthocyanin biosynthesis in Oenanthe javanica. Planta 2018, 247, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Kim, J.B.; Cho, K.J.; Cheon, C.I.; Sung, M.K.; Choung, M.G.; Roh, K.H. Arabidopsis R2R3-MYB transcription factor AtMYB60 functions as a transcriptional repressor of anthocyanin biosynthesis in lettuce (Lactuca sativa). Plant Cell Rep. 2008, 27, 985–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsuka, T.; Yamada, E.; Saito, M.; Fujita, K.; Nishihara, M. Heterologous expression of gentian MYB1R transcription factors suppresses anthocyanin pigmentation in tobacco flowers. Plant Cell Rep. 2013, 32, 1925–1937. [Google Scholar] [CrossRef]
- Zhang, W.; Ning, G.G.; Lv, H.Y.; Liao, L.; Bao, M.Z. Single MYB-type transcription factor AtCAPRICE: A new efficient tool to engineer the production of anthocyanin in tobacco. Biochem. Biophys. Res. Commun. 2009, 388, 742–747. [Google Scholar] [CrossRef]
- Zhong, C.M.; Tang, Y.; Pang, B.; Li, X.K.; Yang, Y.P.; Deng, J.; Feng, C.Y.; Li, L.F.; Ren, G.P.; Wang, Y.Q.; et al. The R2R3-MYB transcription factor GhMYB1a regulates flavonol and anthocyanin accumulation in Gerbera hybrida. Hortic. Res. 2020, 7, 78. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, J.X.; Chen, K.L.; Yao, W.K.; Zhang, B.X.; Wang, J.Y.; Tian, S.T.; Liu, H.L.; Wang, Y.Q.; Liu, Y.L.; et al. A novel R3 MYB transcriptional repressor, MaMYBx, finely regulates anthocyanin biosynthesis in grape hyacinth. Plant Sci. 2020, 298, 110588. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, D.H.; Lee, J.Y.; Lim, S.H. The R3-Type MYB transcription factor BrMYBL2.1 negatively regulates anthocyanin biosynthesis in Chinese Cabbage (Brassica rapa L.) by repressing MYB-bHLH-WD40 complex activity. Int. J. Mol. Sci. 2022, 23, 3382. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, N.; Nakamae, H.; Maekawa, L. Effects of light and kinetin on anthocyanin accumulation in the petals of Rosa hybrida Hort cv. Ehigasa. Z. Pflanzenphysiol. 1980, 98, 263–270. [Google Scholar] [CrossRef]
- Liu, Y.H.; Lin-Wang, K.; Espley, R.V.; Wang, L.; Li, Y.M.; Liu, Z.; Zhou, P.; Zeng, L.H.; Zhang, X.J.; Zhang, J.L.; et al. StMYB44 negatively regulates anthocyanin biosynthesis at high temperatures in tuber flesh of potato. J. Exp. Bot. 2019, 70, 3809–3824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhao, X.Y.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. MdCOP1 ubiquitin E3 ligases interact with MdMYB1 to regulate light-induced anthocyanin biosynthesis and red fruit coloration in apple. Plant Physiol. 2012, 160, 1011–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.H.; Wang, L.; Zhang, J.L.; Yu, B.; Wang, J.; Wang, D. The MYB transcription factor StMYBA1 from potato requires light to activate anthocyanin biosynthesis in transgenic tobacco. J. Plant Biol. 2017, 60, 93–101. [Google Scholar] [CrossRef]
- Feng, S.Q.; Wang, Y.L.; Yang, S.; Xu, Y.T.; Shen, X.S. Anthocyanin biosynthesis in pears is regulated by a R2R3-MYB transcription factor PyMYB10. Planta 2010, 232, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.B.; Bai, S.L.; Zhao, Y.; Qian, M.J.; Tao, R.Y.; Yin, L.; Gao, L.; Teng, Y.W. Ethylene response factors Pp4ERF24 and Pp12ERF96 regulate blue light-induced anthocyanin biosynthesis in ‘Red Zaosu’ pear fruits by interacting with MYB114. Plant Mol. Biol. 2019, 99, 67–78. [Google Scholar] [CrossRef]
- Li, S.N.; Wang, W.Y.; Gao, J.L.; Yin, K.Q.; Wang, R.; Wang, C.C.; Petersen, M.; Mundy, J.; Qiu, J.L. MYB75 phosphorylation by MPK4 is required for light-induced anthocyanin accumulation in Arabidopsis. Plant Cell 2016, 28, 2866. [Google Scholar] [CrossRef] [Green Version]
- An, J.P.; Liu, X.; Li, H.H.; You, C.X.; Wang, X.F.; Hao, Y.J. Apple ring finger E3 ubiquitin ligase MdMIEL1 negatively regulates salt and oxidative stresses tolerance. J. Plant Biol. 2017, 60, 137–145. [Google Scholar] [CrossRef]
- An, J.P.; Wang, X.F.; Zhang, X.W.; Xu, H.F.; Bi, S.Q.; You, C.X.; Hao, Y.J. An apple MYB transcription factor regulates cold tolerance and anthocyanin accumulation and undergoes MIEL1-mediated degradation. Plant Biotechnol. J. 2020, 18, 337–353. [Google Scholar] [CrossRef] [Green Version]
- An, J.P.; An, X.H.; Yao, J.F.; Wang, X.N.; You, C.X.; Wang, X.F.; Hao, Y.J. BTB protein MdBT2 inhibits anthocyanin and proanthocyanidin biosynthesis by triggering MdMYB9 degradation in apple. Tree Physiol. 2018, 38, 1578–1587. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Zhang, X.W.; You, C.X.; Bi, S.Q.; Wang, X.F.; Hao, Y.J. MdWRKY40 promotes wounding-induced anthocyanin biosynthesis in association with MdMYB1 and undergoes MdBT1-mediated degradation. New Phytol. 2019, 224, 380–395. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.X.; Li, Z.W.; Song, Y.B.; Zhu, H.; Lin, S.; Huang, R.; Jiang, Y.M.; Duan, X.W. LcNAC13 physically interacts with LcR1MYB1 to coregulate anthocyanin biosynthesis-related genes during litchi fruit ripening. Biomolecules 2019, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Lai, B.; Du, L.N.; Liu, R.; Hu, B.; Su, W.B.; Qin, Y.H.; Zhao, J.T.; Wang, H.C.; Hu, G.B. Two LcbHLH transcription factors interacting with LcMYB1 in regulating late structural genes of anthocyanin biosynthesis in Nicotiana and Litchi chinensis during anthocyanin accumulation. Front. Plant Sci. 2016, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- Akhter, D.; Qin, R.; Nath, U.K.; Eshag, J.; Jin, X.L.; Shi, C.H. A rice gene, OsPL, encoding a MYB family transcription factor confers anthocyanin synthesis, heat stress response and hormonal signaling. Gene 2019, 699, 62–72. [Google Scholar] [CrossRef]
- Nakatsuka, T.; Haruta, K.S.; Pitaksutheepong, C.; Abe, Y.; Kakizaki, Y.; Yamamoto, K.; Shimada, N.; Yamamura, S.; Nishihara, M. Identification and characterization of R2R3-MYB and bHLH transcription factors regulating anthocyanin biosynthesis in gentian flowers. Plant Cell Physiol. 2008, 49, 1818–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matus, J.T.; Cavallini, E.; Loyola, R.; Holl, J.; Finezzo, L.; Santo, S.D.; Vialet, S.; Commisso, M.; Roman, F.; Schubert, A.; et al. A group of grapevine MYBA transcription factors located in chromosome 14 control anthocyanin synthesis in vegetative organs with different specificities compared with the berry color locus. Plant J. 2017, 91, 220–236. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The signaling pathway medicated by MYB TFs during anthocyanin biosynthesis, such as JA signaling pathway (A), cytokinins (CKs) signaling pathway (B), temperature-induced (C), light signal (D), 26S proteasome pathway (E). JA, jasmonic acid; MeJA, jasmonic acid methyl ester.
Figure 1.
The signaling pathway medicated by MYB TFs during anthocyanin biosynthesis, such as JA signaling pathway (A), cytokinins (CKs) signaling pathway (B), temperature-induced (C), light signal (D), 26S proteasome pathway (E). JA, jasmonic acid; MeJA, jasmonic acid methyl ester.

Figure 2.
Schematic model of the interaction between MYB TFs with other proteins. TFs, transcription factors.
Figure 2.
Schematic model of the interaction between MYB TFs with other proteins. TFs, transcription factors.

Table 2.
The genes regulated by MYB TFs during anthocyanin biosynthesis.
| MYB TFs | Genes | Effect | References |
|---|---|---|---|
| MdMYB3 | CHS CHI UFGT FLS | Activate | [35] |
| OsPL | PAL CHS ANS MYB55 | Suppress | [105] |
| VvMYBC2L2 | CHS DFR LAR UFGT AN1a AN1b | Suppress | [83] |
| PyMYB10 | CHS PAL CHI DFR ANS F3H | Activate | [96] |
| AgMYB2 | CHS CHI F3H F3′H DFR LDOX UGT78D2 | Activate | [66] |
| OjMYB1 | CHS CHI F3H F3′H DFR LDOX UGT78D2 | Activate | [85] |
| EsAN2 | CHS CHI F3H F3′H FLS DFR ANS An1a An1b | Activate | [65] |
| RsMYB1 | F3H DFR ANS | Activate | [57] |
| AaMYB2 | F3H ANS CHS | Activate | [98] |
| MdMYB24L | UFGT DFR | Activate | [40] |
| MdMYBL2 | DFR UFGT MYB10 bHLH3 | Suppress | [80] |
| PsMYB114L PsMYB12L | FLS ANR | Suppress | [53] |
| PsMYB114L PsMYB12L | DFR ANS | Activate | [53] |
| DcMYB7 | CHS1 CHI1 F3H1 F3′H1 FR1 LDOX1 UCGalT1 | Activate | [49] |
| DcMYB6 | CHS1 CHI1 F3H1 F3′H1 DFR LDOX1 UGT78D2 | Activate | [48] |
| PdMYB118 | CHS1 CHI1 F3H F3′H F3′5′H DFR2 ANS1 | Activate | [54] |
| NsMYB1 | PAL C4H 4CL F3H F3′H DFR ANA | Activate | [73] |
| LvMYB5 | CHS DFR ANS | Activate | [62] |
| MYB2 | DFR ANS CHS | Activate | [74] |
| GhMYB1a | MYB1 CHS F3H F3′H DFR ANS FLS UFGT MYB10 MYC1 | Activate | [89] |
| SlAN2 | PAL C4H 4CL CHS1 CHS2 CHI CHI-like F3H F3′5′H DFR ANS 3GT RT AAC PAT GST AN1 AN11 MYBATV MYBATV-like TRY MYB76 | Activate | [46] |
| MaMYBx | C4H 4CL F3′5′H FLS DFR ANS AN2 AN1a AN1b | Suppress | [90] |
Table 3.
The target genes bound by MYB TFs during anthocyanin biosynthesis.
| Species | MYB TFs | Target Genes | References |
|---|---|---|---|
| Apple | MdMYBA | MdANS | [20] |
| Arabidopsis | AtMYBL2 | DFR TT8 | [75] |
| Apple | MdMYB24L | MdDFR MdUFGT | [40] |
| Tomato | SlMYB75 | LOXC AADC2 TPS | [45] |
| Chrysanthemum | CmMYB6 | CmDFR CmUFGT | [81] |
| Carrot | DcMYB7 | DcUCGXT1 DcSAT1 | [49] |
| Poplar | PdMYB118 | CHS1 DRF2 ANS1 | [54] |
| Kiwifruit | AcMYB123 | AcANS AcF3GT1 | [72] |
| Tobacco | GtMYB3 | CHS F3′5′H 5AT | [106] |
| Tobacco | GtMYB3-GtbHLH1 | GtDFR | [87] |
| Grapevine | MYBA1, MYBA6.1 MYBA7 | UFGT 3AT | [107] |
| Grapevine | MYBA1 | F3′5′H | [107] |
| Sweet cherry | PavMYB10.1 | PavANS PavUFGT | [47] |
| Litchi | LcMYB5 | Bhlh1 | [50] |
| Chrysanthemum | CmMYB6 | CmDFR | [59] |
| Lily | LvMYB1 | LvANS | [62] |
| Gerbera | GhMYB1 | NtCHS NtFLS | [89] |
| Grape hyacinth | MaMYBx | MaMybA MabHLH1 | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, C.; Yu, W.; Xu, J.; Lu, X.; Liu, Y. Anthocyanin Biosynthesis Induced by MYB Transcription Factors in Plants. Int. J. Mol. Sci. 2022, 23, 11701. https://doi.org/10.3390/ijms231911701
AMA Style
Li C, Yu W, Xu J, Lu X, Liu Y. Anthocyanin Biosynthesis Induced by MYB Transcription Factors in Plants. International Journal of Molecular Sciences. 2022; 23(19):11701. https://doi.org/10.3390/ijms231911701
Chicago/Turabian StyleLi, Changxia, Wenjin Yu, Junrong Xu, Xuefang Lu, and Yunzhi Liu. 2022. "Anthocyanin Biosynthesis Induced by MYB Transcription Factors in Plants" International Journal of Molecular Sciences 23, no. 19: 11701. https://doi.org/10.3390/ijms231911701
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.