
Phylogenetic and Transcriptional Analyses of the HSP20 Gene Family in Peach Revealed That PpHSP20-32 Is Involved in Plant Height and Heat Tolerance
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Whole Genome Identification and Protein Structure of Peach HSP20 Genes
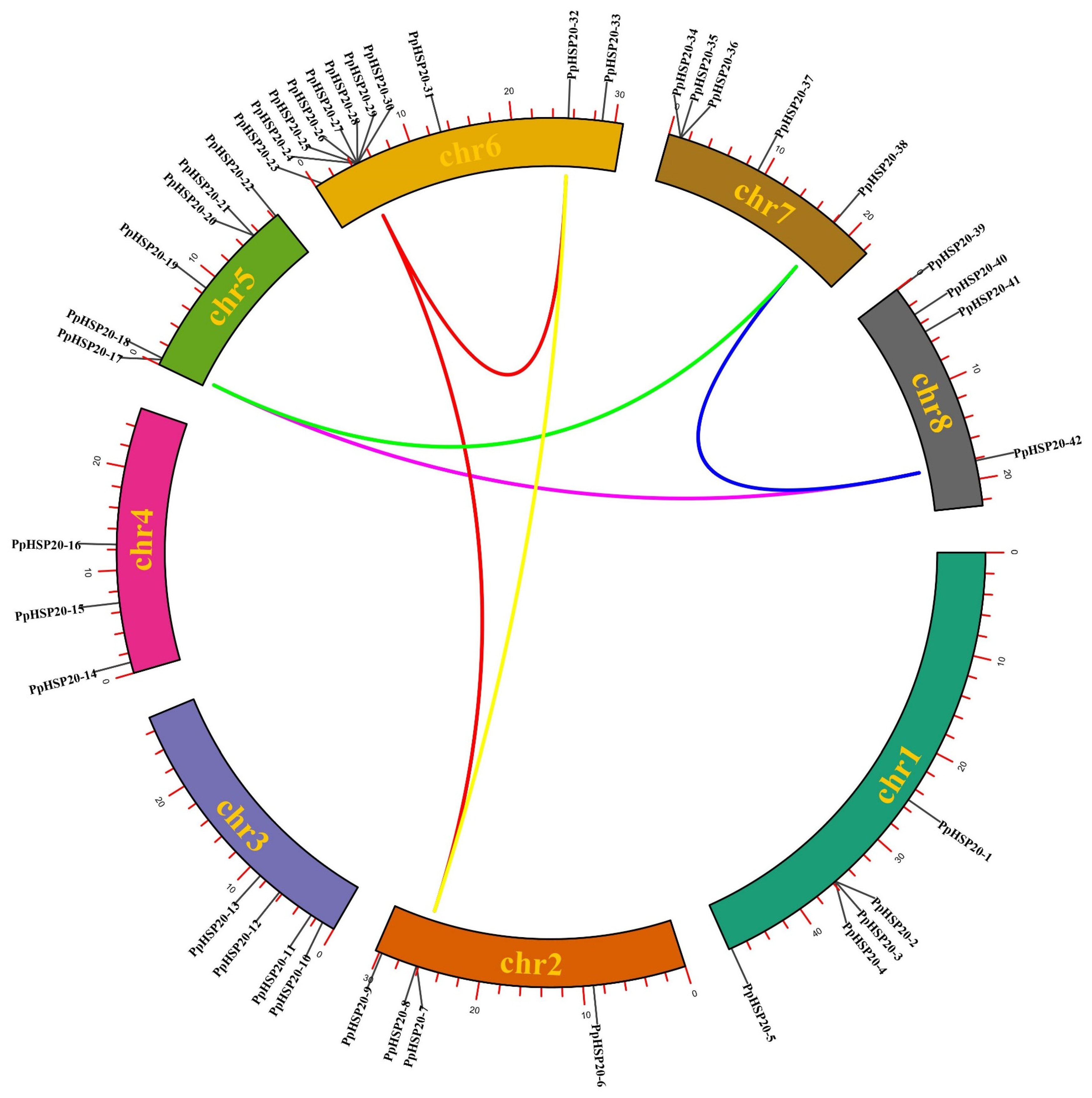
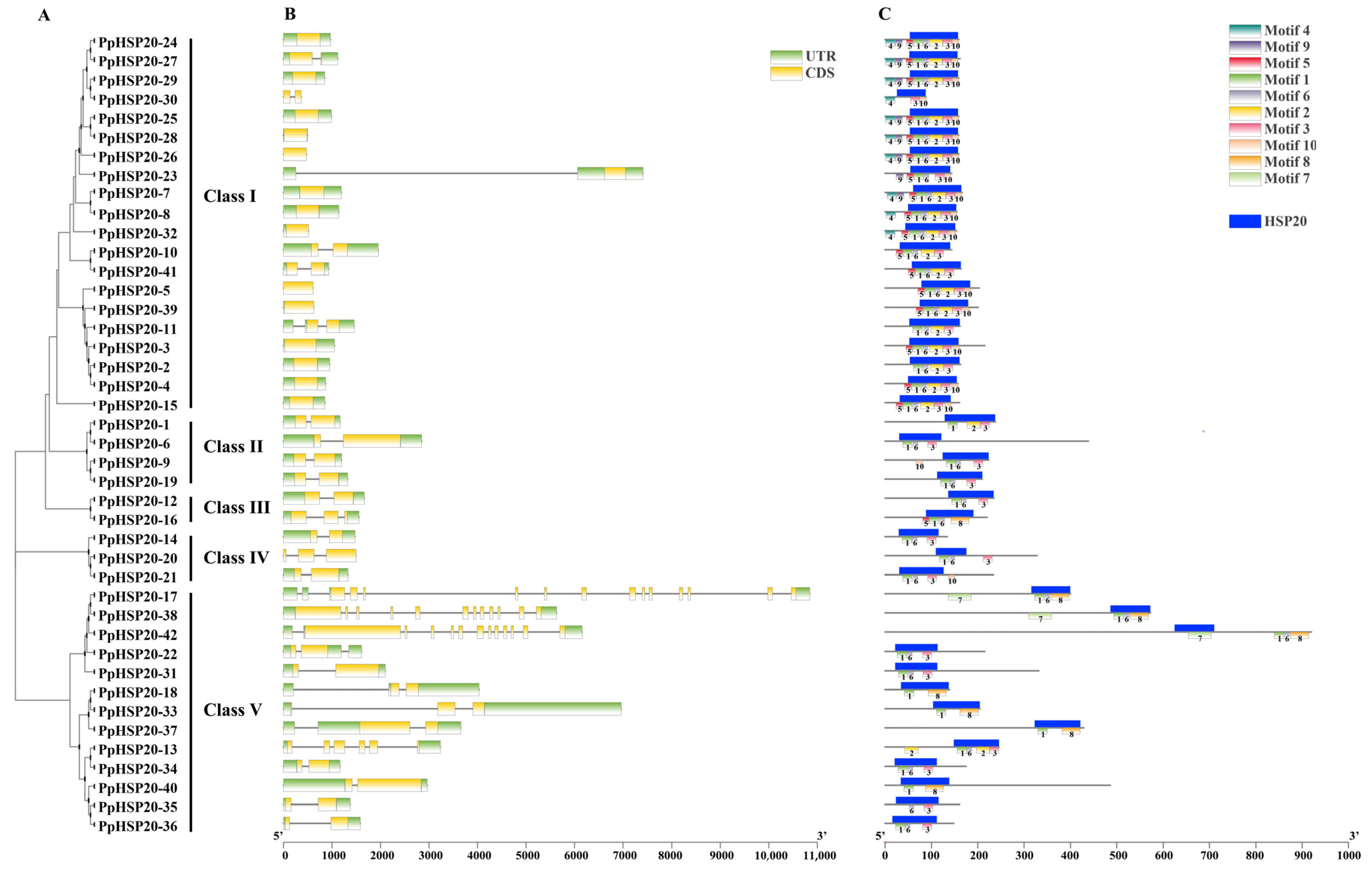
2.2. Chromosome Distribution, Gene Duplication, Gene Structure, and Conserved Motif Analysis of PpHSP20 Genes
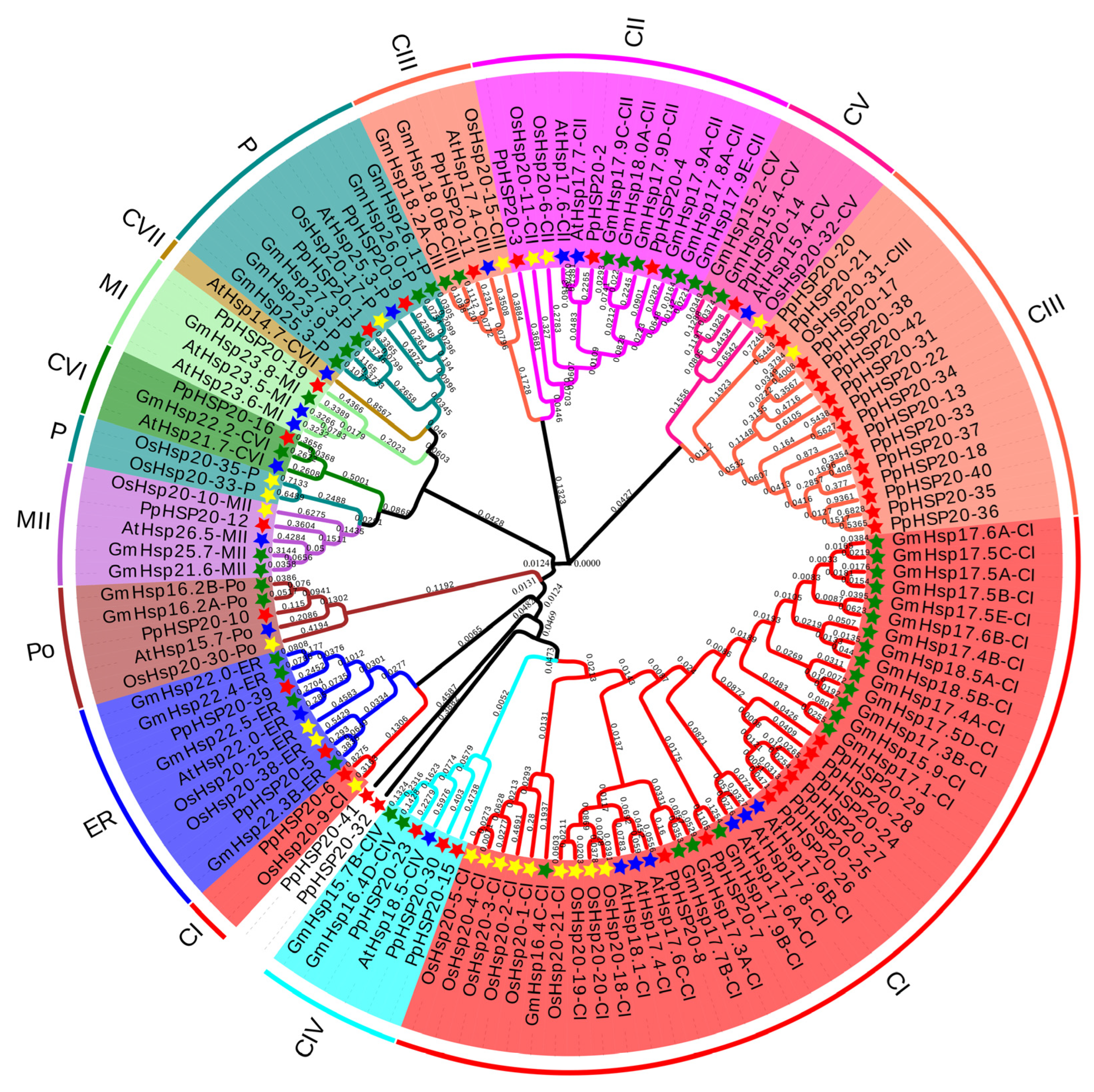
2.3. Phylogenetic Analysis of PpHSP20 Proteins
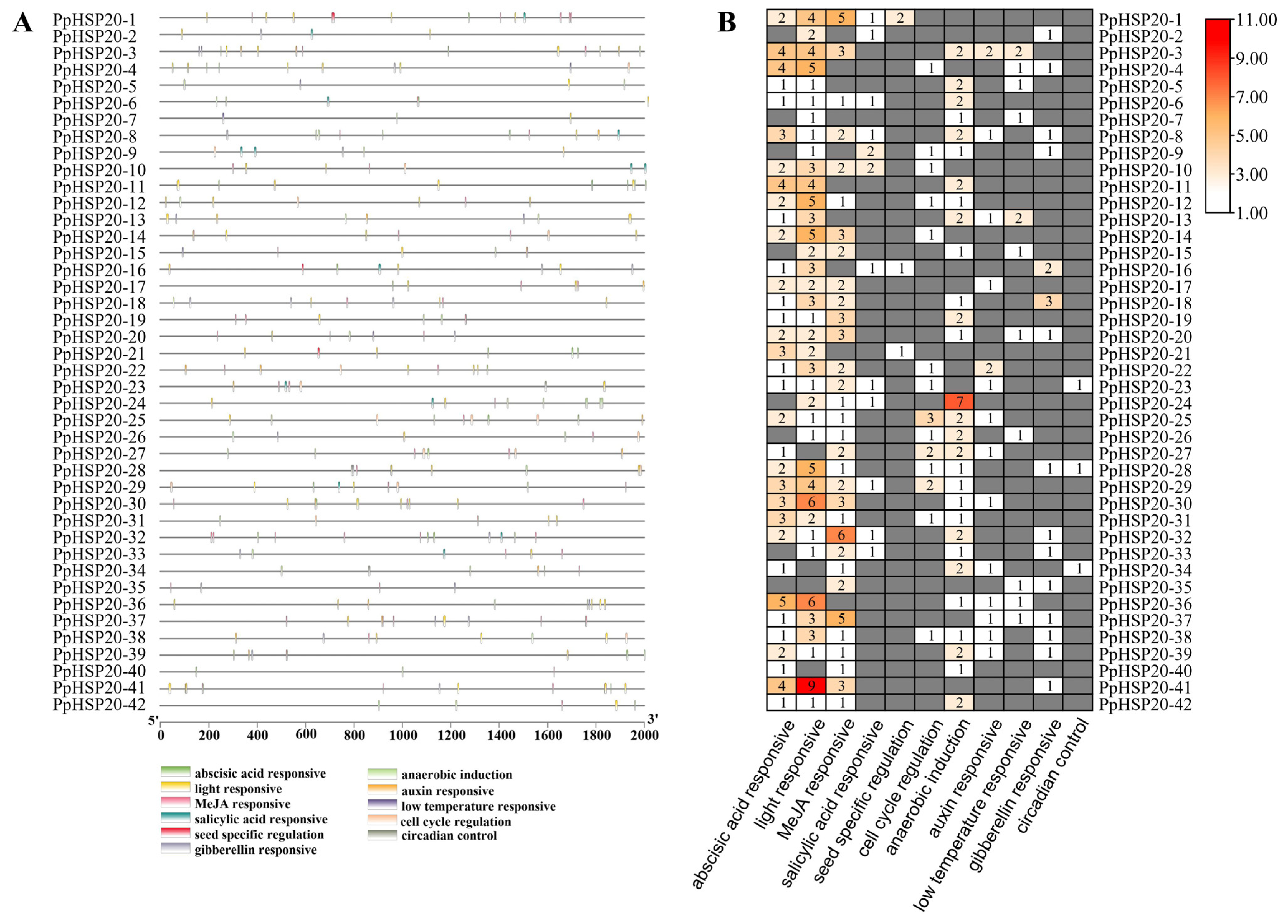
2.4. Analysis of Cis-Acting Elements of PpHSP20s Promoters
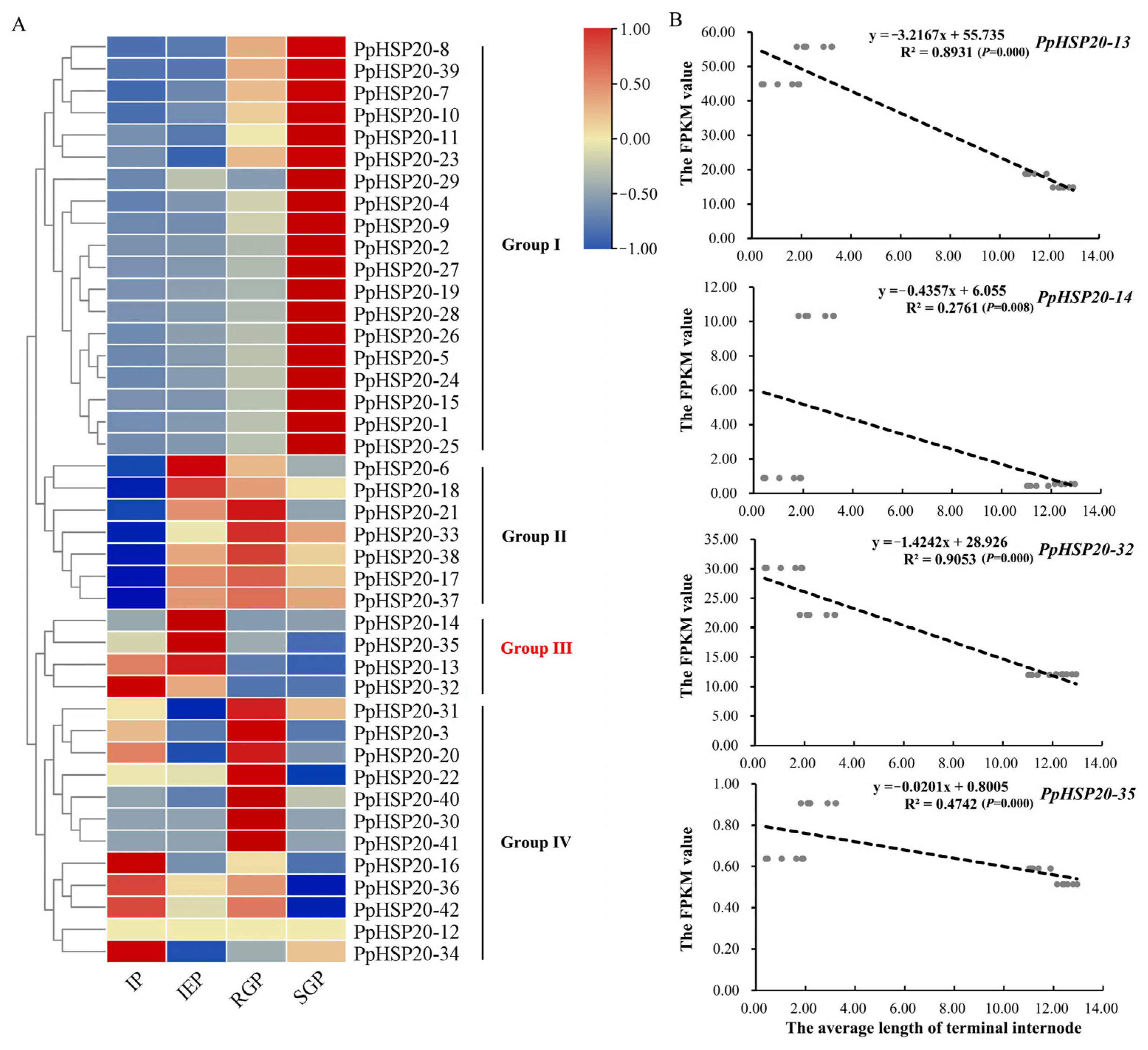
2.5. Expression of PpHSP20s during the Shoot Elongation of ‘Zhongyoutao 14’
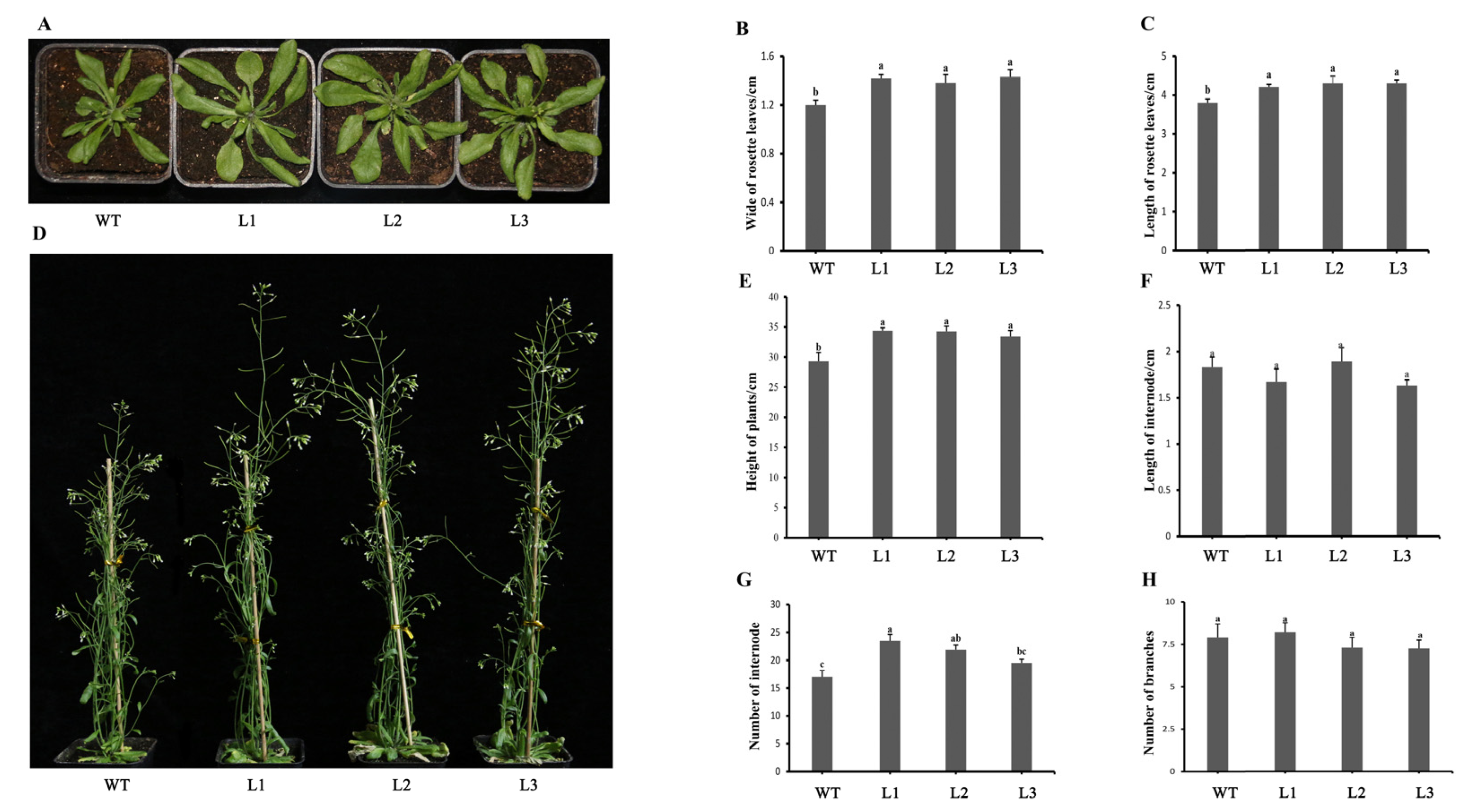
2.6. Overexpression of PpHSP20-32 in Arabidopsis Leads to an Increase in Plant Height
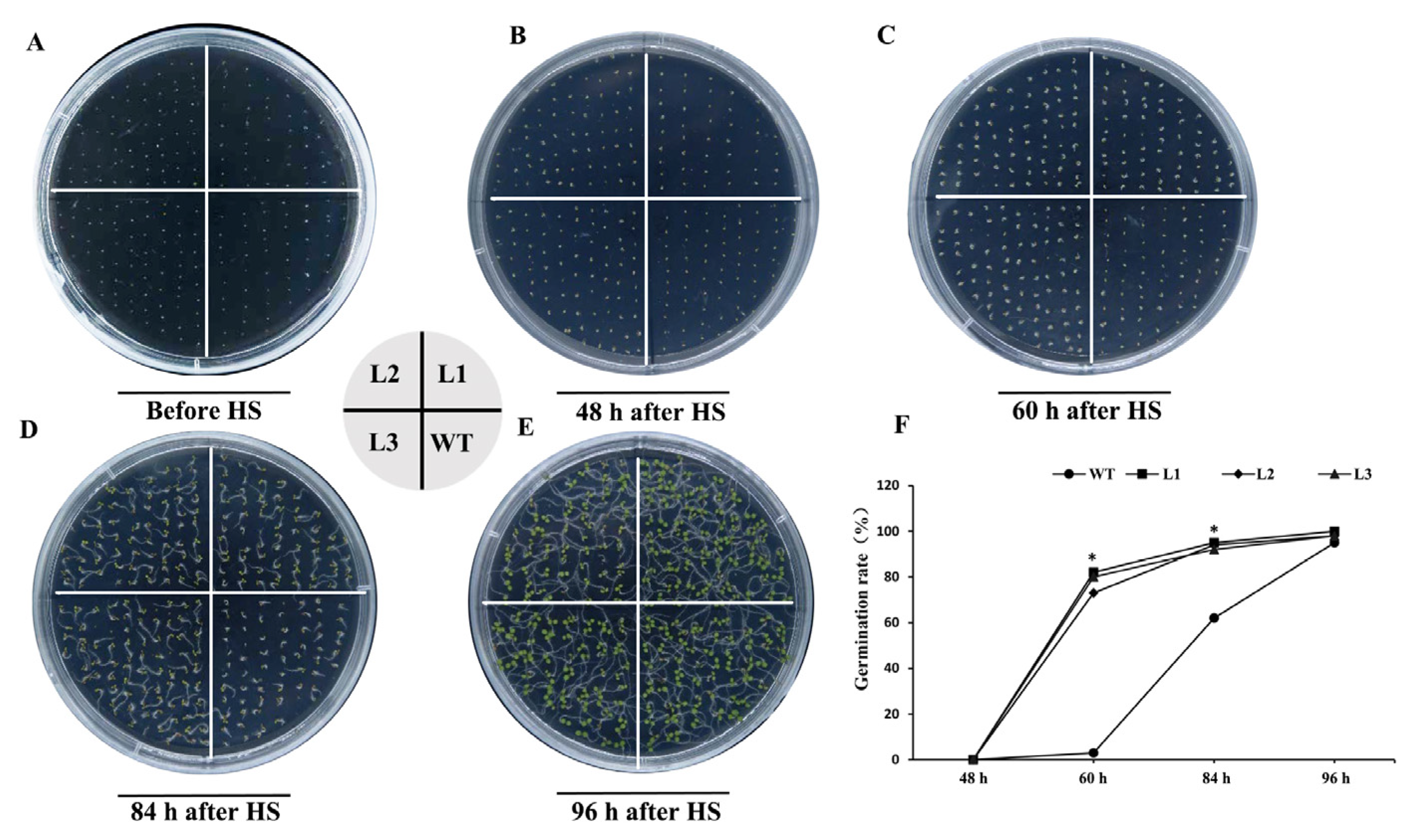
2.7. PpHSP20-32-OE Seeds Exhibit Enhanced Thermotolerance
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Identification and Characteristic Analysis of Peach HSP20s
4.3. Chromosome Location and Gene Structure Analysis of the PpHSP20 Genes
4.4. Phylogeny and Motif Analysis of PpHSP20s
4.5. Analysis of Cis-Acting Elements of PpHSP20s
4.6. PpHSP20 Gene Expression in Different Growth Stages of Peach
4.7. Generation and Phenotypic Observation of PpHSP20-32-Overexpression in Arabidopsis Lines
4.8. Heat Stress Treatment
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Betancourt, J.L.; Travers, S.E.; Pau, S.; Regetz, J.; Davies, T.J.; Kraft, N.J.B.; et al. Warming experiments underpredict plant phenological responses to climate change. Nature 2012, 485, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Argyris, J.; Truco, M.J.; Ochoa, O.; Knapp, S.J.; Still, D.W.; Lenssen, G.M.; Schut, J.W.; Michelmore, R.W.; Bradford, K.J. Quantitative trait loci associated with seed and seedling traits in Lactuca. Appl. Genet. 2005, 111, 1365–1376. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Wang, N.; Guo, T.; Sun, X.; Jia, X.; Wang, P.; Shao, Y.; Liang, B.; Gong, X.; Ma, F. Functions of two Malus hupehensis (Pamp.) Rehd. YTPs (MhYTP1 and MhYTP2) in biotic- and abiotic-stress responses. Plant Sci. Int. J. Exp. Plant Biol. 2017, 261, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N. Temperature Stress and Responses in Plants. Int. J. Mol. Sci. 2019, 20, 2001. [Google Scholar] [CrossRef] [PubMed]
- VanWallendael, A.; Soltani, A.; Emery, N.C.; Peixoto, M.M.; Olsen, J.; Lowry, D.B. A Molecular View of Plant Local Adaptation: Incorporating Stress-Response Networks. Annu. Rev. Plant Biol. 2019, 70, 559–583. [Google Scholar] [CrossRef]
- Quint, M.; Delker, C.; Franklin, K.A.; Wigge, P.A.; Halliday, K.J.; van Zanten, M. Molecular and genetic control of plant thermomorphogenesis. Nat. Plants 2016, 2, 15190. [Google Scholar] [CrossRef]
- Zhou, Y.; Xun, Q.; Zhang, D.; Lv, M.; Ou, Y.; Li, J. TCP Transcription Factors Associate with PHYTOCHROME INTERACTING FACTOR 4 and CRYPTOCHROME 1 to Regulate Thermomorphogenesis in Arabidopsis thaliana. iScience 2019, 15, 600–610. [Google Scholar] [CrossRef]
- Lian, X.; Tan, B.; Yan, L.; Jiang, C.; Cheng, J.; Zheng, X.; Wang, W.; Chen, T.; Ye, X.; Li, J.; et al. Transcript profiling provides insights into molecular processes during shoot elongation in temperature-sensitive peach (Prunus persica). Sci. Rep. 2020, 10, 7801. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Doring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Fragkostefanakis, S.; RÖTh, S.; Schleiff, E.; Scharf, K.-D. Prospects of engineering thermotolerance in crops through modulation of heat stress transcription factor and heat shock protein networks. Plant Cell Environ. 2015, 38, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Song, C.; Wang, H.; Song, S.; Jiao, J.; Wang, M.; Zheng, X.; Bai, T. Genome-Wide Characterization of the HSP20 Gene Family Identifies Potential Members Involved in Temperature Stress Response in Apple. Front. Genet. 2020, 11, 609184. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zheng, Y.; Chen, B.; Zhi, C.; Qiao, L.; Liu, C.; Pan, Y.; Cheng, Z. Genome-wide identification of small heat shock protein (HSP20) homologs in three cucurbit species and the expression profiles of CsHSP20s under several abiotic stresses. Int. J. Biol. Macromol. 2021, 190, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Vierling, E. The Roles of Heat Shock Proteins in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef]
- Scharf, K.D.; Siddique, M.; Vierling, E. The expanding family of Arabidopsis thaliana small heat stress proteins and a new family of proteins containing alpha-crystallin domains (Acd proteins). Cell Stress Chaperones 2001, 6, 225–237. [Google Scholar] [CrossRef]
- Sarkar, N.K.; Kim, Y.K.; Grover, A. Rice sHsp genes: Genomic organization and expression profiling under stress and development. BMC Genom. 2009, 10, 393. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Lu, J.P.; Zhai, Y.F.; Wang, H.; Gong, Z.H.; Wang, S.B.; Lu, M.H. Genome-wide analysis of the CaHsp20 gene family in pepper: Comprehensive sequence and expression profile analysis under heat stress. Front. Plant Sci. 2015, 6, 806. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Cheng, Y.; Feng, K.; Ruan, M.; Ye, Q.; Wang, R.; Li, Z.; Zhou, G.; Yao, Z.; Yang, Y.; et al. Genome-Wide Identification and Expression Profiling of Tomato Hsp20 Gene Family in Response to Biotic and Abiotic Stresses. Front. Plant Sci. 2016, 7, 1215. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Fan, M.; Sun, Y.; Li, L. Genome-Wide Analysis of Watermelon HSP20s and Their Expression Profiles and Subcellular Locations under Stresses. Int. J. Mol. Sci. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.R.; Yu, Y.H.; Ni, P.Y.; Zhang, G.H.; Guo, D.L. Genome-wide identification of small heat-shock protein (HSP20) gene family in grape and expression profile during berry development. BMC Plant Biol. 2019, 19, 433. [Google Scholar] [CrossRef]
- Nagaraju, M.; Reddy, P.S.; Kumar, S.A.; Kumar, A.; Rajasheker, G.; Rao, D.M.; Kavi Kishor, P.B. Genome-wide identification and transcriptional profiling of small heat shock protein gene family under diverse abiotic stress conditions in Sorghum bicolor (L.). Int. J. Biol. Macromol. 2020, 142, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J.; Jia, H.; Li, Y.; Xu, X.; Wang, L.; Lu, M. The Populus trichocarpa PtHSP17.8 involved in heat and salt stress tolerances. Plant Cell Rep. 2016, 35, 1587–1599. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.M.; Li, J.; He, J.; Liu, H.; Zhang, H.M. A class I cytosolic HSP20 of rice enhances heat and salt tolerance in different organisms. Sci. Rep. 2020, 10, 1383. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Liu, Y.; Wu, Y.; Xie, Q. The sHSP22 Heat Shock Protein Requires the ABI1 Protein Phosphatase to Modulate Polar Auxin Transport and Downstream Responses. Plant Physiol. 2018, 176, 2406–2425. [Google Scholar] [CrossRef]
- Huang, L.J.; Cheng, G.X.; Khan, A.; Wei, A.M.; Yu, Q.H.; Yang, S.B.; Luo, D.X.; Gong, Z.H. CaHSP16.4, a small heat shock protein gene in pepper, is involved in heat and drought tolerance. Protoplasma 2019, 256, 39–51. [Google Scholar] [CrossRef]
- Lopes-Caitar, V.S.; de Carvalho, M.C.C.G.; Darben, L.M.; Kuwahara, M.K.; Nepomuceno, A.L.; Dias, W.P.; Abdelnoor, R.V.; Marcelino-Guimarães, F.C. Genome-wide analysis of the Hsp 20 gene family in soybean: Comprehensive sequence, genomic organization and expression profile analysis under abiotic and biotic stresses. BMC Genom. 2013, 14, 577. [Google Scholar] [CrossRef]
- Li, Y.; Cao, K.; Li, N.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Guo, J.; Wang, Q.; Ding, T.; et al. Genomic analyses provide insights into peach local adaptation and responses to climate change. Genome Res. 2021, 31, 592–606. [Google Scholar] [CrossRef]
- Cao, K.; Wang, L.; Zhu, G.; Fang, W.; Chen, C.; Luo, J. Genetic diversity, linkage disequilibrium, and association mapping analyses of peach (Prunus persica) landraces in China. Tree Genet. Genomes 2012, 8, 975–990. [Google Scholar] [CrossRef]
- Lu, Z.H.; Niu, L.; Chagne, D.; Cui, G.C.; Pan, L.; Foster, T.; Zhang, R.P.; Zeng, W.F.; Wang, Z.Q. Fine mapping of the temperature-sensitive semi-dwarf (Tssd) locus regulating the internode length in peach (Prunus persica). Mol. Breed. 2016, 36, 20. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef]
- Martin, H.; Elizabeth, V. A First Line of Stress Defense: Small Heat Shock Proteins and Their Function in Protein Homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar]
- Yan, H.; Zhang, A.; Chen, J.; He, X.; Xu, B.; Xie, G.; Miao, Z.; Zhang, X.; Huang, L. Genome-Wide Analysis of the PvHsp20 Family in Switchgrass: Motif, Genomic Organization, and Identification of Stress or Developmental-Related Hsp20s. Front. Plant Sci. 2017, 8, 1024. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-wide analysis of the potato Hsp20 gene family: Identification, genomic organization and expression profiles in response to heat stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef]
- Tan, B.; Lian, X.; Cheng, J.; Zeng, W.; Zheng, X.; Wang, W.; Ye, X.; Li, J.; Li, Z.; Zhang, L.; et al. Genome-wide identification and transcriptome profiling reveal that E3 ubiquitin ligase genes relevant to ethylene, auxin and abscisic acid are differentially expressed in the fruits of melting flesh and stony hard peach varieties. BMC Genom. 2019, 20, 892. [Google Scholar] [CrossRef]
- Wang, G.M.; Yin, H.; Qiao, X.; Tan, X.; Gu, C.; Wang, B.H.; Cheng, R.; Wang, Y.Z.; Zhang, S.L. F-box genes: Genome-wide expansion, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri). Plant Sci. Int. J. Exp. Plant Biol. 2016, 253, 164–175. [Google Scholar] [CrossRef]
- Tan, B.; Yan, L.; Li, H.; Lian, X.; Cheng, J.; Wang, W.; Zheng, X.; Wang, X.; Li, J.; Ye, X.; et al. Genome-wide identification of HSF family in peach and functional analysis of PpHSF5 involvement in root and aerial organ development. PeerJ 2021, 9, e10961. [Google Scholar] [CrossRef]
- International Peach Genome, I.; Verde, I.; Abbott, A.G.; Scalabrin, S.; Jung, S.; Shu, S.; Marroni, F.; Zhebentyayeva, T.; Dettori, M.T.; Grimwood, J.; et al. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nat. Genet. 2013, 45, 487–494. [Google Scholar]
- Siddique, M.; Gernhard, S.; von Koskull-Doring, P.; Vierling, E.; Scharf, K.D. The plant sHSP superfamily: Five new members in Arabidopsis thaliana with unexpected properties. Cell Stress Chaperones 2008, 13, 183–197. [Google Scholar] [CrossRef]
- Kumar, S.V.; Wigge, P.A. H2A.Z-containing nucleosomes mediate the thermosensory response in Arabidopsis. Cell 2010, 140, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Y.; Kieffer, M.; Yu, H.; Kepinski, S.; Estelle, M. HSP90 regulates temperature-dependent seedling growth in Arabidopsis by stabilizing the auxin co-receptor F-box protein TIR1. Nat. Commun. 2016, 7, 10269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, E.; Mano, S.; Nomoto, M.; Tada, Y.; Hara-Nishimura, I.; Nishimura, M.; Yamada, K. HSP90 Stabilizes Auxin-Responsive Phenotypes by Masking a Mutation in the Auxin Receptor TIR1. Plant Cell Physiol. 2016, 57, 2245–2254. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Chung, M.H.; Chen, M.K.; Pan, S.M. Floral spray transformation can efficiently generate Arabidopsis transgenic plants. Transgenic Res. 2000, 9, 471–476. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Amino Acid Number | Molecular Weight/Da | Isoelectric Point | Instability Index | Aliphatic Index | Average of Hydropathicity |
|---|---|---|---|---|---|---|---|
| Prupe.1G237800 | PpHSP20-1 | 237 | 26,812.57 | 9.13 | 53.71 | 57.59 | −0.802 |
| Prupe.1G407100 | PpHSP20-2 | 162 | 18,223.84 | 6.64 | 53.2 | 85.99 | −0.54 |
| Prupe.1G407200 | PpHSP20-3 | 214 | 22,898.39 | 5.78 | 45.85 | 65.09 | −0.86 |
| Prupe.1G407300 | PpHSP20-4 | 156 | 17,514.17 | 5.58 | 39.38 | 80.51 | −0.439 |
| Prupe.1G586200 | PpHSP20-5 | 202 | 22,714.25 | 9.12 | 34.67 | 90.4 | −0.356 |
| Prupe.2G065600 | PpHSP20-6 | 438 | 47,399.43 | 9.18 | 41.9 | 66.28 | −0.828 |
| Prupe.2G243400 | PpHSP20-7 | 165 | 18,345.7 | 6.21 | 48.88 | 76.12 | −0.621 |
| Prupe.2G243800 | PpHSP20-8 | 154 | 17,381.68 | 5.81 | 55.45 | 81.04 | −0.671 |
| Prupe.2G317700 | PpHSP20-9 | 223 | 24,801.18 | 7.75 | 47.95 | 85.61 | −0.533 |
| Prupe.3G017400 | PpHSP20-10 | 143 | 16,084.37 | 6.84 | 42.01 | 85.17 | −0.45 |
| Prupe.3G034800 | PpHSP20-11 | 161 | 17,783.21 | 6.16 | 42.95 | 82.92 | −0.463 |
| Prupe.3G085200 | PpHSP20-12 | 234 | 26,468.86 | 6.87 | 55.03 | 86.2 | −0.672 |
| Prupe.3G108500 | PpHSP20-13 | 245 | 27,393.14 | 9.26 | 58.68 | 75.27 | −0.469 |
| Prupe.4G023100 | PpHSP20-14 | 133 | 15,541.7 | 5.52 | 49.18 | 89.4 | −0.353 |
| Prupe.4G125800 | PpHSP20-15 | 160 | 18,417.87 | 7.13 | 37.8 | 73.06 | −0.832 |
| Prupe.4G201100 | PpHSP20-16 | 219 | 25,002.23 | 6.91 | 50.18 | 73.42 | −0.624 |
| Prupe.5G004500 | PpHSP20-17 | 399 | 44,140.85 | 6.35 | 31.61 | 75.21 | −0.619 |
| Prupe.5G006000 | PpHSP20-18 | 138 | 14,813.27 | 8.54 | 41.03 | 100.14 | 0.082 |
| Prupe.5G071200 | PpHSP20-19 | 209 | 23,640.61 | 5.46 | 70.11 | 74.07 | −0.626 |
| Prupe.5G186700 | PpHSP20-20 | 327 | 37,390.03 | 5.13 | 55.85 | 81.07 | −0.488 |
| Prupe.5G186800 | PpHSP20-21 | 233 | 25,847.96 | 5.61 | 44.56 | 78.58 | −0.609 |
| Prupe.5G242900 | PpHSP20-22 | 214 | 23,777.01 | 9.26 | 28.11 | 75.09 | −0.575 |
| Prupe.6G008800 | PpHSP20-23 | 143 | 16,155.26 | 4.69 | 67.62 | 83.85 | −0.298 |
| Prupe.6G065900 | PpHSP20-24 | 158 | 18,076.55 | 7.91 | 50.58 | 74.62 | −0.666 |
| Prupe.6G066100 | PpHSP20-25 | 158 | 18,105.5 | 5.83 | 49.72 | 73.99 | −0.683 |
| Prupe.6G066200 | PpHSP20-26 | 158 | 17,961.49 | 6.76 | 48.21 | 80.13 | −0.555 |
| Prupe.6G066300 | PpHSP20-27 | 161 | 18,427.82 | 5.57 | 49.59 | 73.23 | −0.671 |
| Prupe.6G066400 | PpHSP20-28 | 158 | 18,222.61 | 6.34 | 51.41 | 71.52 | −0.697 |
| Prupe.6G066500 | PpHSP20-29 | 158 | 18,109.53 | 5.57 | 55.98 | 77.09 | −0.613 |
| Prupe.6G066600 | PpHSP20-30 | 87 | 9408.79 | 9.1 | 57.83 | 74.02 | −0.299 |
| Prupe.6G151500 | PpHSP20-31 | 331 | 36,970.14 | 9.32 | 43.37 | 63.08 | −0.871 |
| Prupe.6G266500 | PpHSP20-32 | 153 | 17,931.3 | 6.45 | 49.89 | 65.56 | −0.746 |
| Prupe.6G328400 | PpHSP20-33 | 204 | 22,150.2 | 5.1 | 29.97 | 79.75 | −0.317 |
| Prupe.7G008300 | PpHSP20-34 | 174 | 19,246.36 | 9.33 | 39.03 | 86.32 | −0.243 |
| Prupe.7G008400 | PpHSP20-35 | 160 | 18,391.29 | 9.28 | 43.56 | 85.81 | −0.296 |
| Prupe.7G008500 | PpHSP20-36 | 147 | 16,918.68 | 9.44 | 32.95 | 88.78 | −0.276 |
| Prupe.7G053900 | PpHSP20-37 | 428 | 48,041.14 | 5.88 | 43.93 | 77.15 | −0.42 |
| Prupe.7G187900 | PpHSP20-38 | 572 | 62,944.2 | 4.93 | 48.98 | 60.28 | −0.957 |
| Prupe.8G000400 | PpHSP20-39 | 200 | 22,775.12 | 5.91 | 46.96 | 97.4 | −0.442 |
| Prupe.8G031300 | PpHSP20-40 | 485 | 54,400.11 | 5.36 | 50.45 | 72.76 | −0.911 |
| Prupe.8G046200 | PpHSP20-41 | 163 | 18,241.31 | 5.62 | 48.21 | 67.48 | −0.799 |
| Prupe.8G182600 | PpHSP20-42 | 919 | 100,089.87 | 4.59 | 43.82 | 74.11 | −0.633 |
| Protein ID | Alpha Helix | Extended Strand | Beta Turn | Random Coil |
|---|---|---|---|---|
| PpHSP20-1 | 51 | 34 | 6 | 146 |
| PpHSP20-2 | 56 | 34 | 12 | 60 |
| PpHSP20-3 | 60 | 35 | 17 | 102 |
| PpHSP20-4 | 49 | 36 | 10 | 61 |
| PpHSP20-5 | 78 | 29 | 12 | 83 |
| PpHSP20-6 | 154 | 36 | 8 | 240 |
| PpHSP20-7 | 34 | 29 | 9 | 93 |
| PpHSP20-8 | 32 | 32 | 7 | 83 |
| PpHSP20-9 | 32 | 46 | 10 | 135 |
| PpHSP20-10 | 32 | 31 | 10 | 70 |
| PpHSP20-11 | 50 | 33 | 9 | 69 |
| PpHSP20-12 | 84 | 33 | 13 | 104 |
| PpHSP20-13 | 51 | 38 | 8 | 148 |
| PpHSP20-14 | 28 | 31 | 7 | 67 |
| PpHSP20-15 | 33 | 38 | 6 | 83 |
| PpHSP20-16 | 62 | 46 | 18 | 93 |
| PpHSP20-17 | 103 | 48 | 18 | 230 |
| PpHSP20-18 | 14 | 44 | 8 | 72 |
| PpHSP20-19 | 49 | 30 | 11 | 119 |
| PpHSP20-20 | 99 | 53 | 28 | 147 |
| PpHSP20-21 | 57 | 36 | 14 | 126 |
| PpHSP20-22 | 56 | 49 | 12 | 97 |
| PpHSP20-23 | 15 | 38 | 10 | 80 |
| PpHSP20-24 | 24 | 24 | 10 | 91 |
| PpHSP20-25 | 26 | 34 | 8 | 90 |
| PpHSP20-26 | 29 | 31 | 8 | 90 |
| PpHSP20-27 | 28 | 28 | 8 | 97 |
| PpHSP20-28 | 29 | 27 | 7 | 95 |
| PpHSP20-29 | 30 | 30 | 9 | 89 |
| PpHSP20-30 | 13 | 14 | 3 | 57 |
| PpHSP20-31 | 143 | 20 | 8 | 160 |
| PpHSP20-32 | 31 | 35 | 10 | 77 |
| PpHSP20-33 | 21 | 53 | 13 | 117 |
| PpHSP20-34 | 60 | 30 | 30 | 30 |
| PpHSP20-35 | 48 | 34 | 12 | 66 |
| PpHSP20-36 | 56 | 29 | 12 | 50 |
| PpHSP20-37 | 85 | 84 | 27 | 232 |
| PpHSP20-38 | 144 | 67 | 23 | 338 |
| PpHSP20-39 | 76 | 31 | 9 | 84 |
| PpHSP20-40 | 202 | 61 | 30 | 192 |
| PpHSP20-41 | 42 | 29 | 8 | 84 |
| PpHSP20-42 | 312 | 130 | 51 | 426 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, X.; Wang, Q.; Li, T.; Gao, H.; Li, H.; Zheng, X.; Wang, X.; Zhang, H.; Cheng, J.; Wang, W.; et al. Phylogenetic and Transcriptional Analyses of the HSP20 Gene Family in Peach Revealed That PpHSP20-32 Is Involved in Plant Height and Heat Tolerance. Int. J. Mol. Sci. 2022, 23, 10849. https://doi.org/10.3390/ijms231810849
Lian X, Wang Q, Li T, Gao H, Li H, Zheng X, Wang X, Zhang H, Cheng J, Wang W, et al. Phylogenetic and Transcriptional Analyses of the HSP20 Gene Family in Peach Revealed That PpHSP20-32 Is Involved in Plant Height and Heat Tolerance. International Journal of Molecular Sciences. 2022; 23(18):10849. https://doi.org/10.3390/ijms231810849
Chicago/Turabian StyleLian, Xiaodong, Qiuping Wang, Tianhao Li, Hongzhu Gao, Huannan Li, Xianbo Zheng, Xiaobei Wang, Haipeng Zhang, Jun Cheng, Wei Wang, and et al. 2022. "Phylogenetic and Transcriptional Analyses of the HSP20 Gene Family in Peach Revealed That PpHSP20-32 Is Involved in Plant Height and Heat Tolerance" International Journal of Molecular Sciences 23, no. 18: 10849. https://doi.org/10.3390/ijms231810849