
TTYH3 Modulates Bladder Cancer Proliferation and Metastasis via FGFR1/H-Ras/A-Raf/MEK/ERK Pathway
 ,
, 
 and
and 
Abstract
:1. Introduction
2. Results
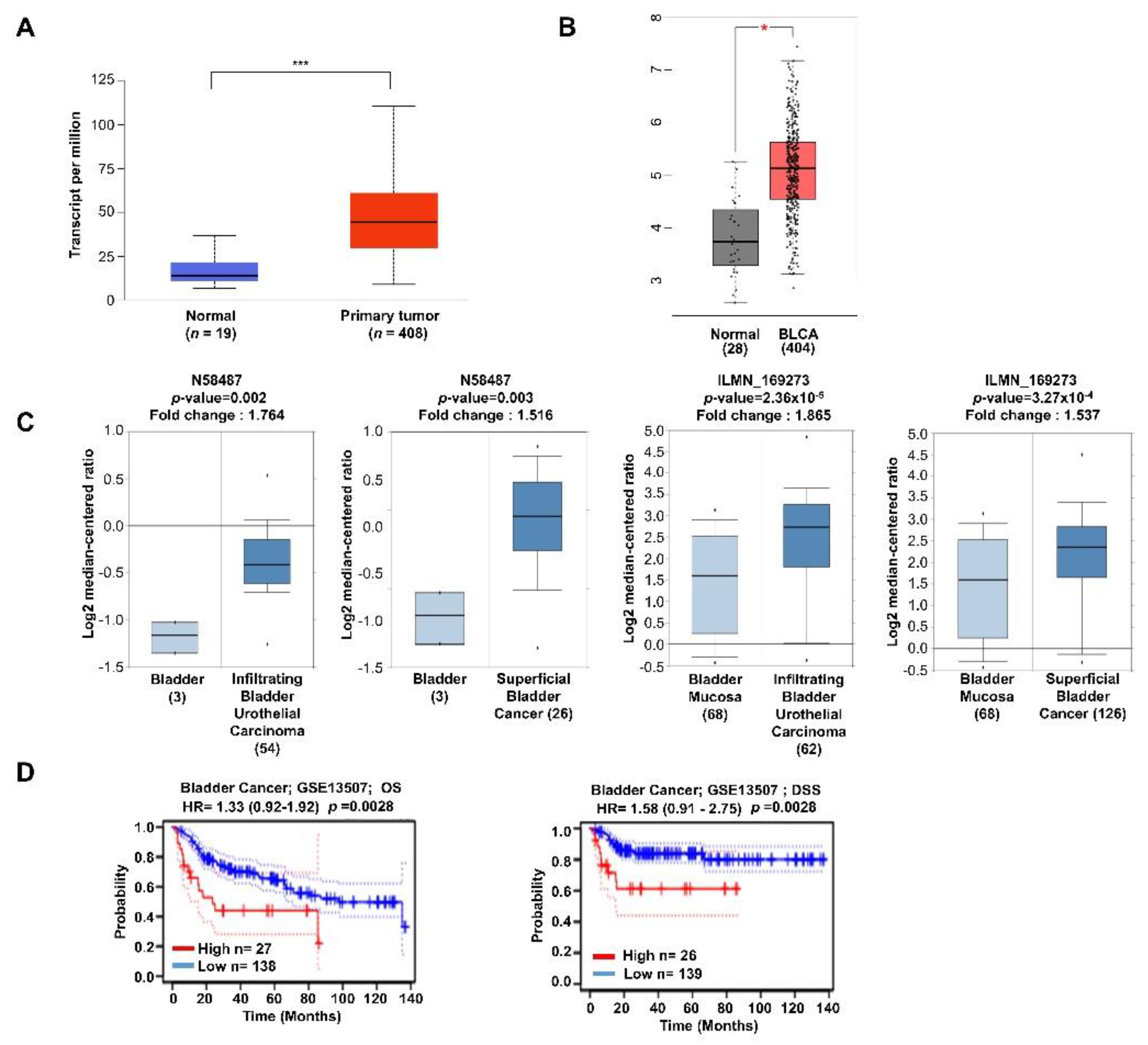
2.1. Expression of TTYH3 Is Associated with Decreased Survival Rates in Bladder Cancer Patients
2.2. TTYH3 Knockdown Suppresses Tumor Growth and Proliferation in Bladder Cancer Cells
2.3. TTYH3 Involvement in a Variety of Signaling Pathways That Regulate Cellular Functions, including MAPK Signaling
2.4. TTYH3 Knockdown Inhibits Bladder Cancer Cell Migration and Invasion
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. TTYH3 Knockdown Using Lentiviral Vector
4.3. RNA Isolation and RT-PCR Analysis
4.4. Western Blotting
4.5. Cell Proliferation
4.6. Sphere-Forming Assay
4.7. Wound Closure Assay
4.8. Transwell Assay
4.9. Statistical Analysis
4.10. Bioinformatics Database Analysis
4.10.1. UALCAN Database Analysis
4.10.2. GEPIA2 Database Analysis
4.10.3. Oncomine Database Analysis
4.10.4. PrognoScan Database Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Bray, F.; Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Westergren, D.-O.; Westergren, D.O.; Gårdmark, T.; Lindhagen, L.; Chau, A.; Malmström, P.U. A nationwide, population based analysis of patients with organ confined, muscle invasive bladder cancer not receiving curative intent therapy in Sweden from 1997 to 2014. J. Urol. 2019, 202, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.; Xiao, J.F.; Agarwal, N.; Duex, J.E.; Theodorescu, D. Advances in bladder cancer biology and therapy. Nat. Rev. Cancer 2021, 21, 104–121. [Google Scholar] [CrossRef] [PubMed]
- Charlton, M.E.; Adamo, M.; Sun, L.; Deorah, S. Bladder cancer collaborative stage variables and their data quality, usage, and clinical implications: A review of SEER data, 2004–2010. Cancer 2014, 120, 3815–3825. [Google Scholar] [CrossRef]
- Dobruch, J.; Oszczudłowski, M. Bladder cancer: Current challenges and future directions. Medicina 2021, 57, 749. [Google Scholar] [CrossRef]
- Mistretta, F.; Lughezzani, G.; Lista, G.; Larcher, A.; Fossati, N.; Abrate, A.; Dell’Oglio, P.; Montorsi, F.; Guazzoni, G.; Lazzeri, M. Bladder cancer and urothelial impairment: The role of TRPV1 as potential drug target. BioMed Res. Int. 2014, 2014, 987149. [Google Scholar] [CrossRef]
- Malysz, J.; Petkov, G.V. Urinary bladder smooth muscle ion channels: Expression, function, and regulation in health and disease. Am. J. Physiol. -Ren. Physiol. 2020, 319, F257–F283. [Google Scholar] [CrossRef]
- Corry, B. Understanding ion channel selectivity and gating and their role in cellular signalling. Mol. Biosyst. 2006, 2, 527–535. [Google Scholar] [CrossRef]
- Brading, A.; Brain, K. Ion. channel modulators and urinary tract function. Urin. Tract 2011, 2011, 375–393. [Google Scholar]
- Litan, A.; Langhans, S.A. Cancer as a channelopathy: Ion channels and pumps in tumor development and progression. Front. Cell. Neurosci. 2015, 9, 86. [Google Scholar] [CrossRef]
- Saberbaghi, T.; Wong, R.; Rutka, J.T.; Wang, G.L.; Feng, Z.P.; Sun, H.S. Role of Cl− channels in primary brain tumour. Cell Calcium 2019, 81, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lu, M.; Liu, B.; Huang, Y.; Wang, K. Inhibition of Ca2+-activated Cl− channel ANO1/TMEM16A expression suppresses tumor growth and invasiveness in human prostate carcinoma. Cancer Lett. 2012, 326, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, S.; Hou, F.; Zhang, C.; Gao, J.; Wang, K. Inhibition of Ca2+-activated chloride channel ANO1 suppresses ovarian cancer through inactivating PI3K/Akt signaling. Int. J. Cancer 2019, 144, 2215–2226. [Google Scholar] [CrossRef]
- Britschgi, A.; Bill, A.; Brinkhaus, H.; Rothwell, C.; Clay, I.; Duss, S.; Rebhan, M.; Raman, P.; Guy, C.T.; Wetzel, K.; et al. Calcium-activated chloride channel ANO1 promotes breast cancer progression by activating EGFR and CAMK signaling. Proc. Natl. Acad. Sci. USA 2013, 110, E1026–E1034. [Google Scholar] [CrossRef]
- Zhang, C.; Li, H.; Gao, J.; Cui, X.; Yang, S.; Liu, Z. Prognostic significance of ANO1 expression in cancers. Medicine 2021, 100, e24525. [Google Scholar] [CrossRef]
- Suzuki, M. The Drosophilatweety family: Molecular candidates for large-conductance Ca2+-activated Cl− channels. Exp. Physiol. 2006, 91, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Mizuno, A. A novel human Cl-channel family related to Drosophila flightless locus. J. Biol. Chem. 2004, 279, 22461–22468. [Google Scholar] [CrossRef]
- Nalamalapu, R.R.; Yue, M.; Stone, A.R.; Murphy, S.; Saha, M.S. The tweety Gene Family: From Embryo to Disease. Front. Mol. Neurosci. 2021, 14, 672511. [Google Scholar] [CrossRef]
- Li, B.; Hoel, C.M.; Brohawn, S.G. Structures of tweety homolog proteins TTYH2 and TTYH3 reveal a Ca2+-dependent switch from intra-to intermembrane dimerization. Nat. Commun. 2021, 12, 1–9. [Google Scholar]
- Saha, S.K.; Biswas, P.K.; Gil, M.; Cho, S.G. High expression of TTYH3 is related to poor clinical outcomes in human gastric cancer. J. Clin. Med. 2019, 8, 1762. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Y.; Dong, B.; Xue, W.; Chen, S.; Mitsuo, S.; Zou, H.; Feng, Y.; Ma, K.; Dong, Q.; et al. The TTYH3/MK5 Positive Feedback Loop regulates Tumor Progression via GSK3-β/β-catenin signaling in HCC. Int. J. Biol. Sci. 2022, 18, 4053. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lian, S.; Khoi, P.N.; Yoon, H.J.; Joo, Y.E.; Chay, K.O.; Kim, K.K.; Jung, Y.D. Chrysin inhibits tumor promoter-induced MMP-9 expression by blocking AP-1 via suppression of ERK and JNK pathways in gastric cancer cells. PLoS ONE 2015, 10, e0124007. [Google Scholar]
- Brennan, A.; Leech, J.T.; Kad, N.M.; Mason, J.M. Selective antagonism of c-Jun for cancer therapy. J. Exp. Clin. Cancer Res. 2020, 39, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Huhe, M.; Liu, S.; Zhang, Y.; Zhang, Z.; Chen, Z. Expression levels of transcription factors c-Fos and c-Jun and transmembrane protein HAb18G/CD147 in urothelial carcinoma of the bladder. Mol. Med. Rep. 2017, 15, 2991–3000. [Google Scholar] [CrossRef]
- Habela, C.W.; Ernest, N.J.; Swindall, A.F.; Sontheimer, H. Chloride accumulation drives volume dynamics underlying cell proliferation and migration. J. Neurophysiol. 2009, 101, 750–757. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Wong, E.W.T.; Chang, F.; Lehmann, B.; Terrian, D.M.; Milella, M.; Tafuri, A.; et al. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim. Biophys. Acta BBA -Mol. Cell Res. 2007, 1773, 1263–1284. [Google Scholar] [CrossRef]
- Rosseland, C.M.; Rosseland, C.M.; Wierød, L.; Flinder, L.I.; Oksvold, M.P.; Skarpen, E.; Huitfeldt, H.S. Distinct functions of H-Ras and K-Ras in proliferation and survival of primary hepatocytes due to selective activation of ERK and PI3K. J. Cell. Physiol. 2008, 215, 818–826. [Google Scholar] [CrossRef]
- Michael, J.; Wurtzel, J.; Goldfinger, L. Regulation of H-Ras-driven MAPK signaling, transformation and tumorigenesis, but not PI3K signaling and tumor progression, by plasma membrane microdomains. Oncogenesis 2016, 5, e228. [Google Scholar] [CrossRef]
- Cheng, Q.; Ma, Z.; Shi, Y.; Parris, A.B.; Kong, L.; Yang, X. FGFR1 Overexpression Induces Cancer Cell Stemness and Enhanced Akt/Erk-ER Signaling to Promote Palbociclib Resistance in Luminal A Breast Cancer Cells. Cells 2021, 10, 3008. [Google Scholar] [CrossRef]
- Sheng, Z.; Evans, S.K.; Green, M.R. An activating transcription factor 5-mediated survival pathway as a target for cancer therapy. Oncotarget 2010, 1, 457. [Google Scholar] [CrossRef]
- Chen, B.; Liu, S.; Gan, L.; Wang, J.; Hu, B.; Xu, H.; Tong, R.; Yang, H.; Cristina, I.; Xue, J.; et al. FGFR1 signaling potentiates tumor growth and predicts poor prognosis in esophageal squamous cell carcinoma patients. Cancer Biol. Ther. 2018, 19, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pander, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Target | Forward Primer | Reverse Primer |
|---|---|---|---|
| Human | TTYH3 | 5′-GACGCCTACGTGACCAAAAT-3′ | 5′-GACGGTCCTCAGAAGCTCAG-3′ |
| GAPDH | 5′-AATCCCATCACCATCTTCCAG-3′ | 5′-CACGATACCAAAGTTGTCATG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, P.K.; Kwak, Y.; Kim, A.; Seok, J.; Kwak, H.J.; Lee, M.; Dayem, A.A.; Song, K.; Park, J.-Y.; Park, K.S.; et al. TTYH3 Modulates Bladder Cancer Proliferation and Metastasis via FGFR1/H-Ras/A-Raf/MEK/ERK Pathway. Int. J. Mol. Sci. 2022, 23, 10496. https://doi.org/10.3390/ijms231810496
Biswas PK, Kwak Y, Kim A, Seok J, Kwak HJ, Lee M, Dayem AA, Song K, Park J-Y, Park KS, et al. TTYH3 Modulates Bladder Cancer Proliferation and Metastasis via FGFR1/H-Ras/A-Raf/MEK/ERK Pathway. International Journal of Molecular Sciences. 2022; 23(18):10496. https://doi.org/10.3390/ijms231810496
Chicago/Turabian StyleBiswas, Polash Kumar, Yeonjoo Kwak, Aram Kim, Jaekwon Seok, Hee Jeong Kwak, Moonjung Lee, Ahmed Abdal Dayem, Kwonwoo Song, Jae-Yong Park, Kyoung Sik Park, and et al. 2022. "TTYH3 Modulates Bladder Cancer Proliferation and Metastasis via FGFR1/H-Ras/A-Raf/MEK/ERK Pathway" International Journal of Molecular Sciences 23, no. 18: 10496. https://doi.org/10.3390/ijms231810496