First Discovery of Beta-Sitosterol as a Novel Antiviral Agent against White Spot Syndrome Virus

Abstract
:1. Introduction
2. Results
2.1. In Vivo Antiviral Activity Screening of 15 Herbal Extracts
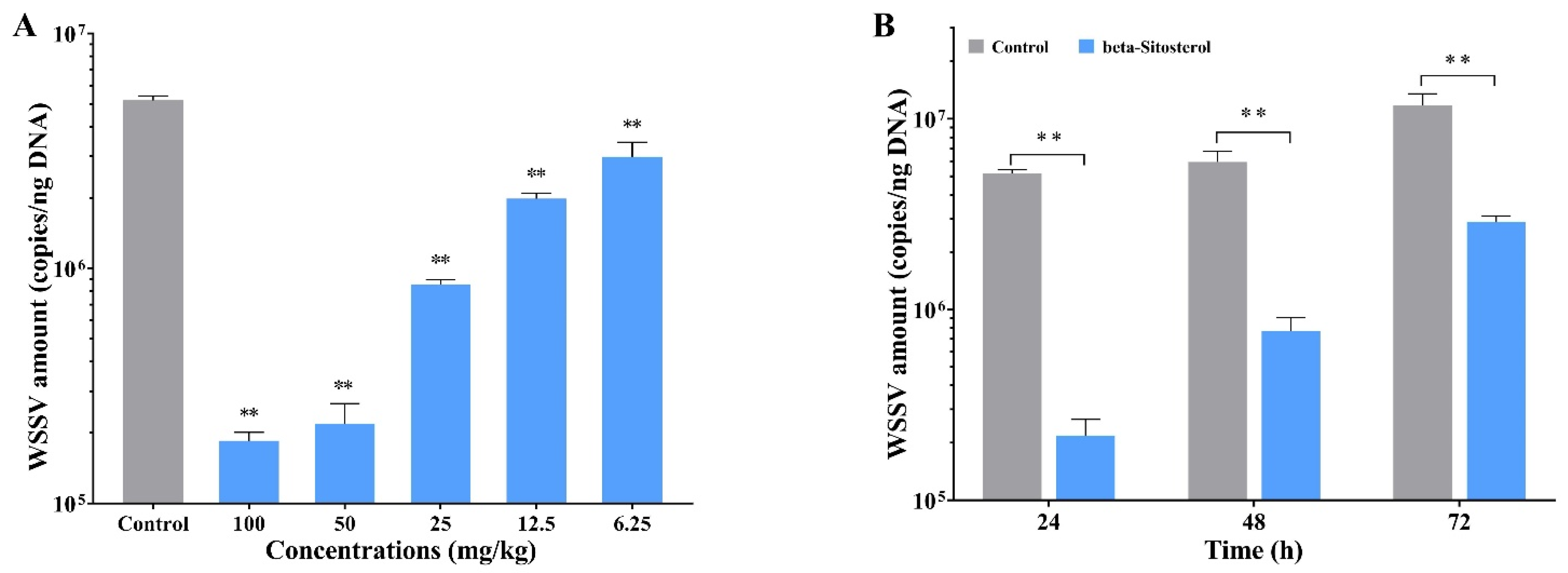
2.2. Beta-Sitosterol Inhibited the Replication of WSSV In Vivo
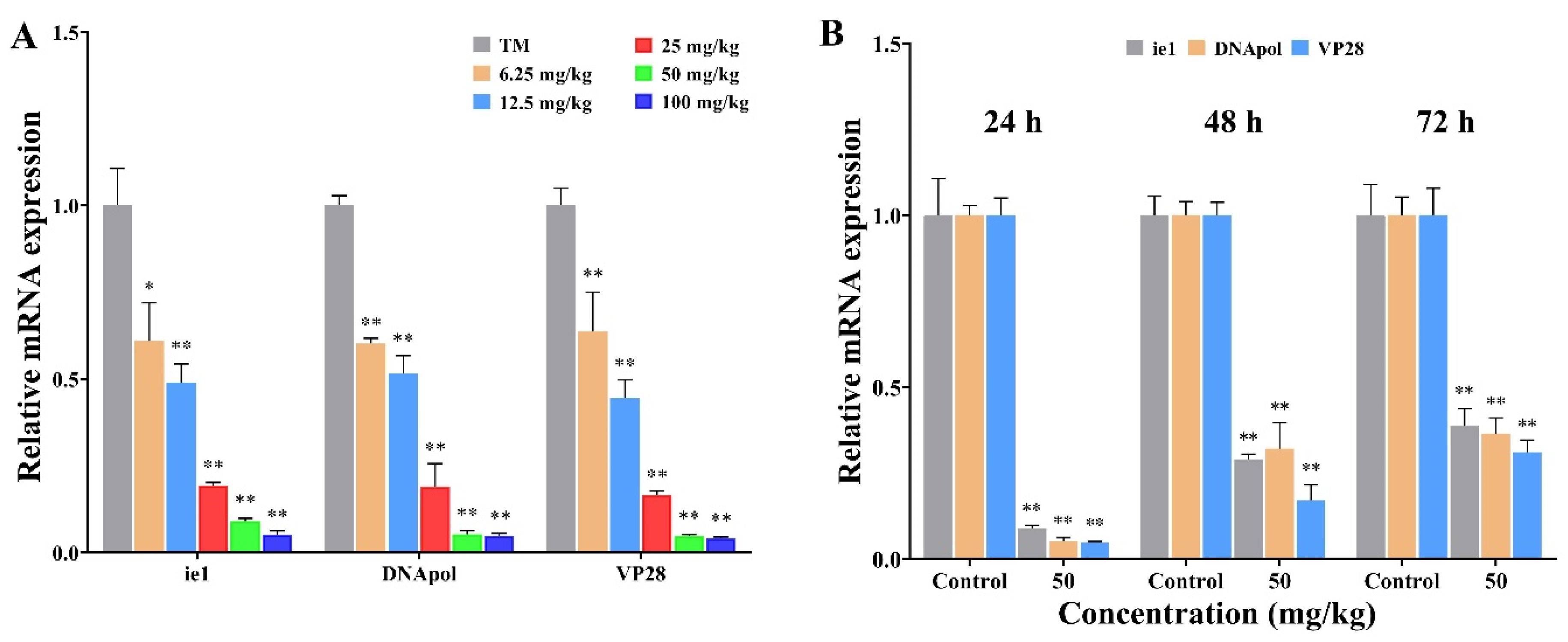
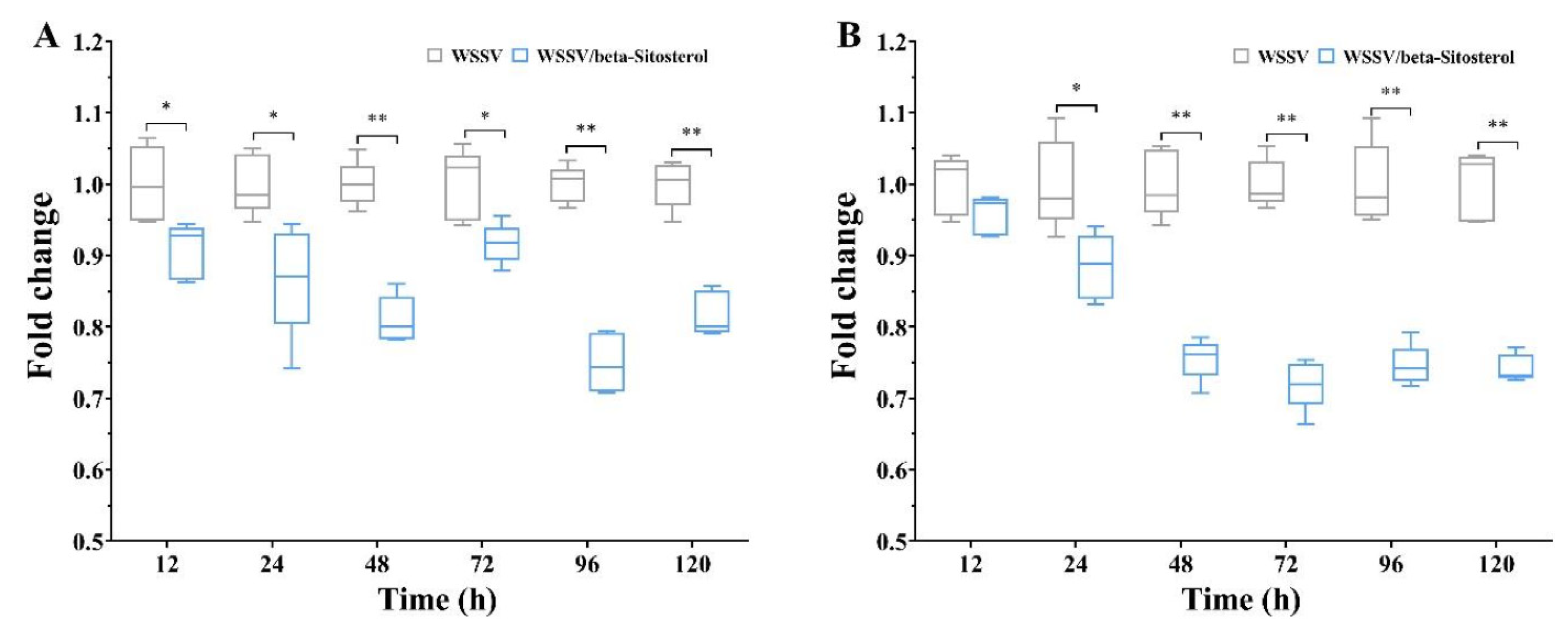
2.3. Beta-Sitosterol Inhibited the Transcription of WSSV Replication-Related Genes
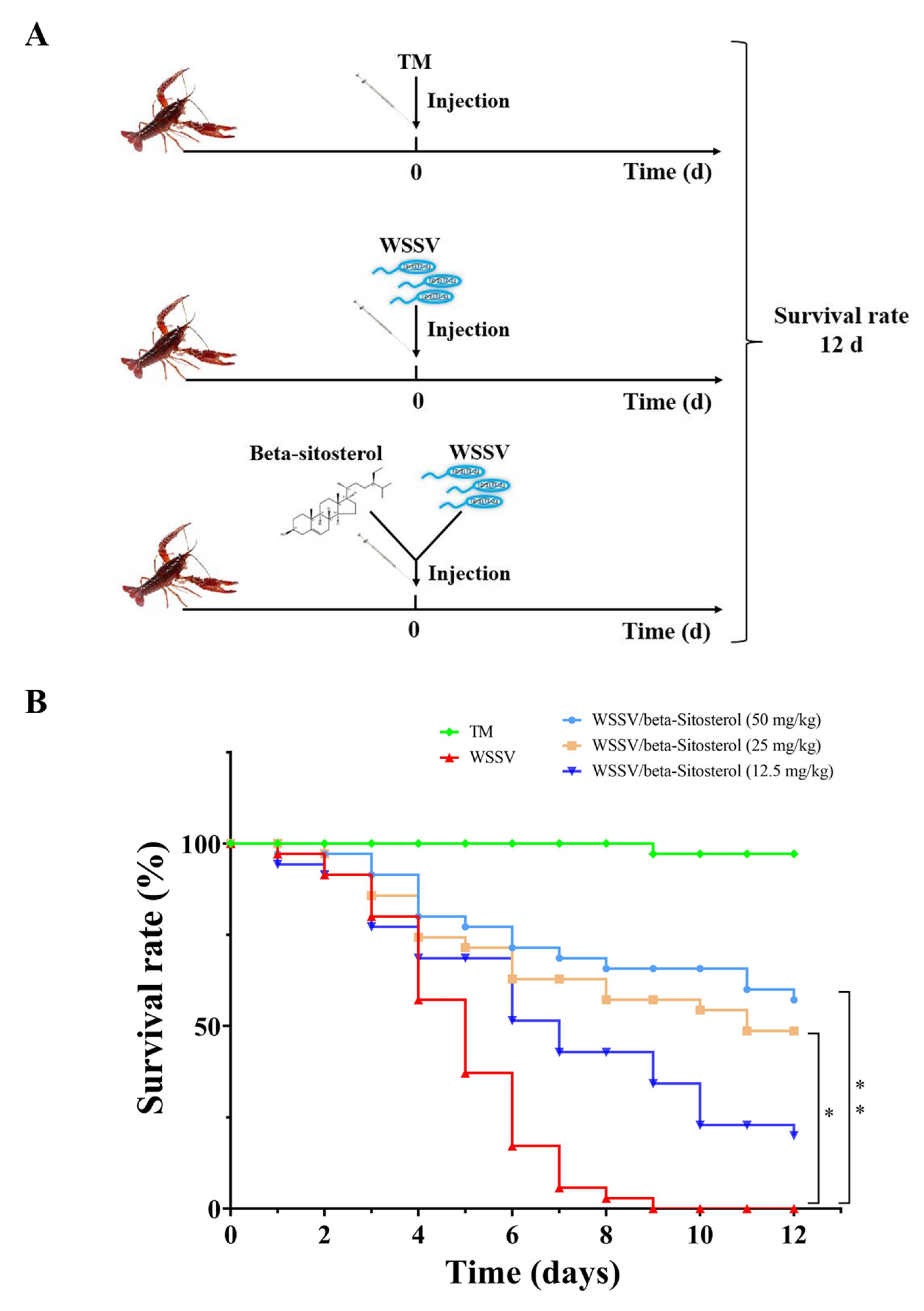
2.4. Beta-Sitosterol Provided Protection against WSSV-Infected Crayfish
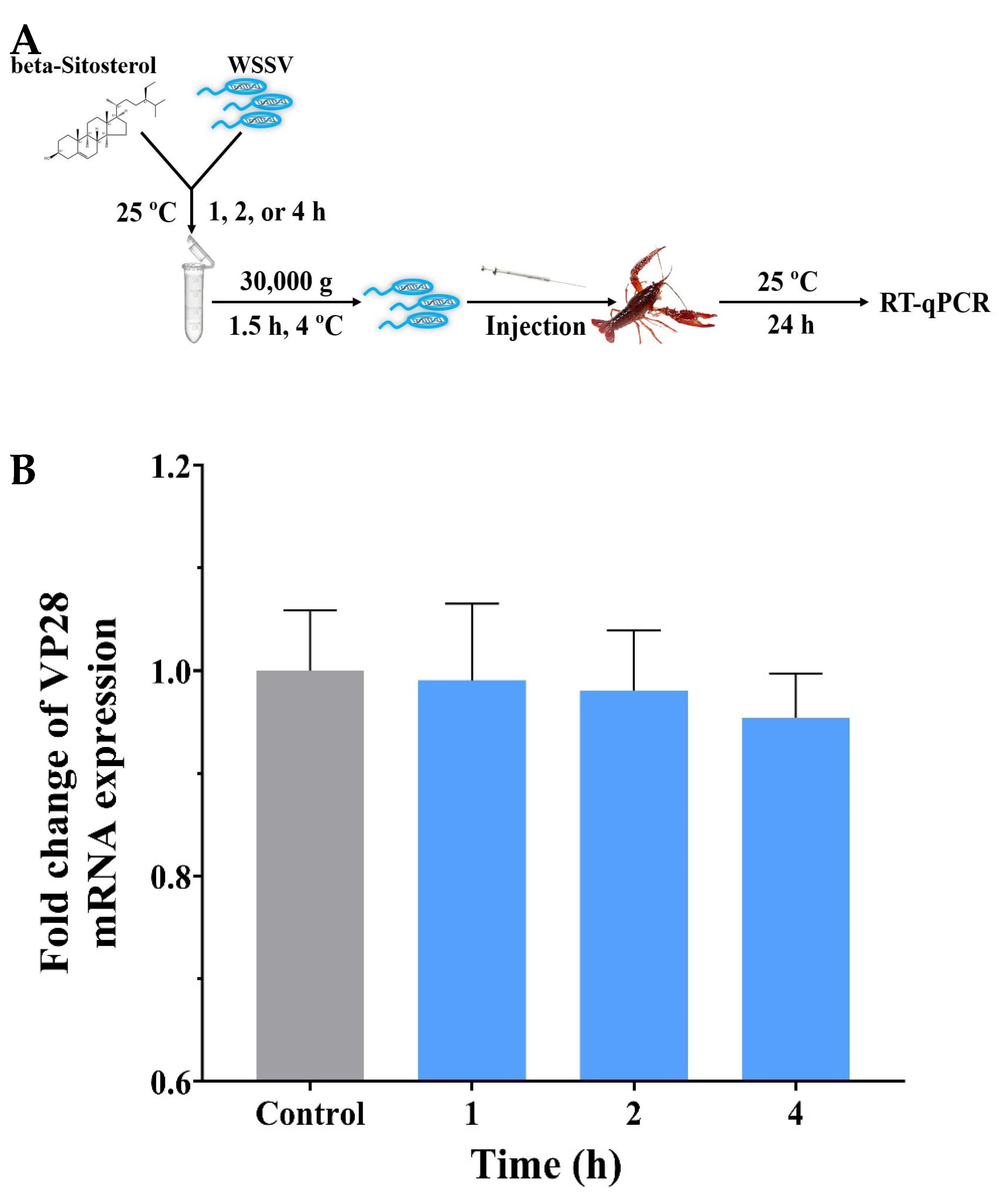
2.5. Beta-Sitosterol Displayed No Discernible Effect on Viral Infectivity
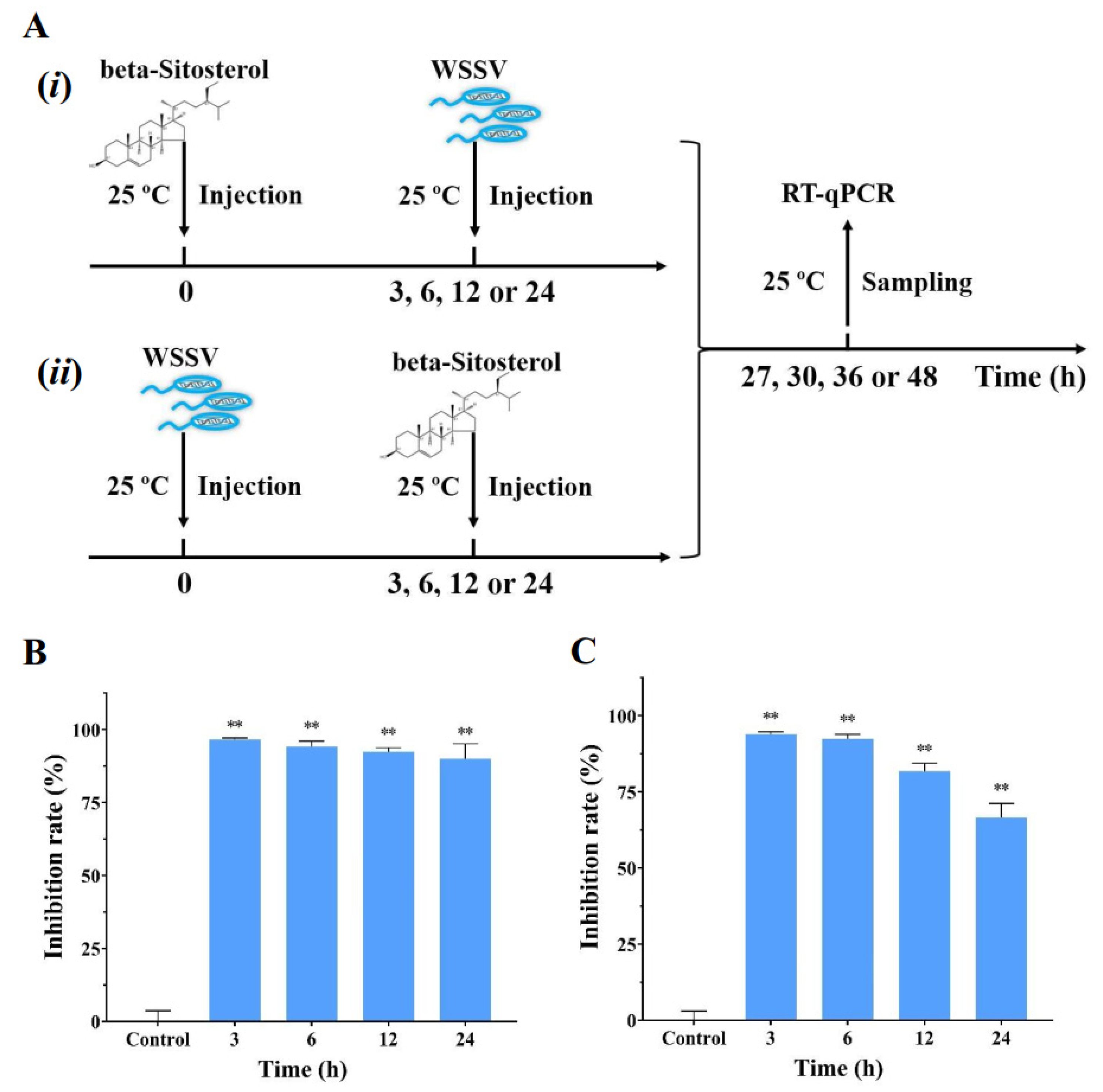
2.6. Beta-Sitosterol Conferred the Prophylactic and Therapeutic Effects against WSSV Infection In Vivo
2.7. Beta-Sitosterol Reduced Total Protein Levels in hEMOLymph and Gill Tissues of Crayfish after WSSV Infection
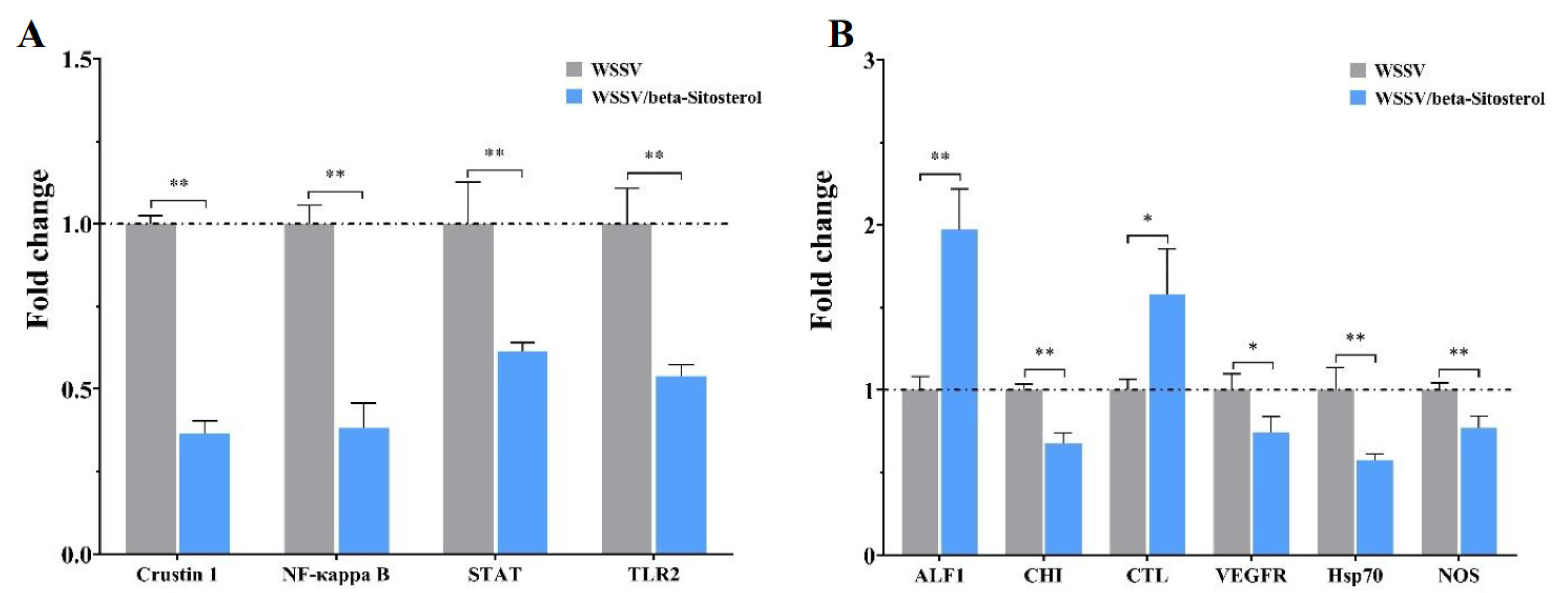
2.8. Beta-Sitosterol Treatment Modulated Innate Immune Signaling Pathways and Immune Factors
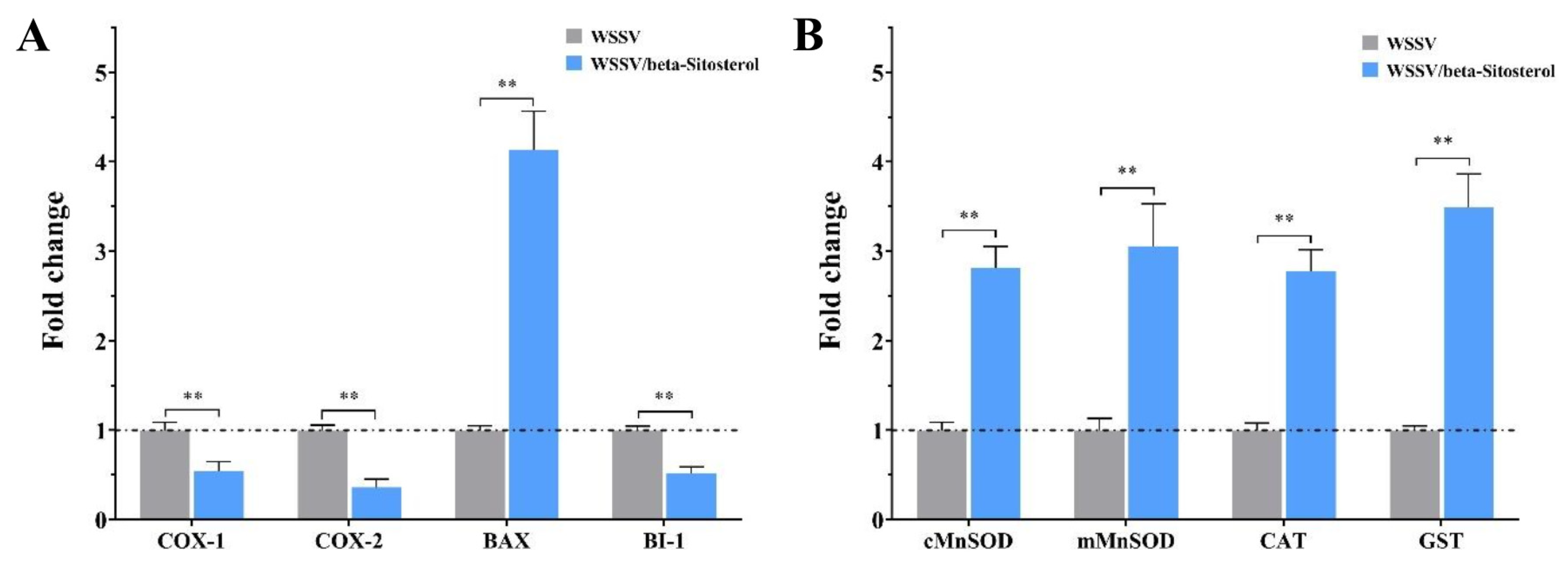
2.9. Beta-Sitosterol Regulated the Expression of Inflammatory-, Apoptosis-, and Antioxidant-Related Factors
3. Discussion
4. Materials and Methods
4.1. Preliminary Screening of Antiviral Medicinal Plants
4.2. Experimental Animals, Viral Inoculum, and Antiviral Compound
4.3. Safety Assessment
4.4. Antiviral Effect of Plant Extracts on WSSV In Vivo
4.5. Antiviral Effect of Beta-Sitosterol on WSSV In Vivo
4.6. Protective Effect of Beta-Sitosterol against WSSV Infection
4.7. The Effect of Beta-Sitosterol on Viral Infectivity In Vitro
4.8. Antiviral Effect of Pre- or Post-Treatment of Beta-Sitosterol against WSSV
4.9. Total Protein Content Determination
4.10. Expression Analysis of Crayfish and Viral Genes
4.11. DNA/RNA Extraction, cDNA Amplification and qRT–PCR
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, X.P.; Li, J.R.; Wang, Y.B.; Fu, L.L.; Fu, Y.Y.; Li, B.Q.; Jiao, B.H. Aquaculture Industry in China: Current State, Challenges, and Outlook. Rev. Fish. Sci. 2011, 19, 187–200. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018; FAO: Rome, Italy, 2018. [Google Scholar]
- Shan, L.-P.; Zhou, Y.; Liu, G.-L.; Yan, M.-C.; Lu, J.-F.; Liu, L.; Chen, J. Evaluation on the antiviral activity of a nitrile phenylpropanoid against white spot syndrome virus infection in Litopenaeus vannamei. Aquaculture 2022, 547, 737384. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Neil, D.M.; Peeler, E.J.; Shields, J.D.; Small, H.J.; Flegel, T.W.; Vlak, J.M.; Jones, B.; Morado, F.; Moss, S.; et al. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J. Invertebr. Pathol. 2012, 110, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Lightner, D.V. Global transboundry disease politics: The OIE perspective. J. Invertebr. Pathol. 2012, 110, 184–187. [Google Scholar] [CrossRef]
- Zhang, H.J.; Yao, X.M.; Ding, Y.F.; Xu, Z.; Liang, R.N.; Zhang, Y.; Wu, Y.L.; Li, B.Q.; Guan, B. PI3K signaling pathways modulated white spot syndrome virus (WSSV) replication in Procambarus clarkii. Fish Shellfish Immunol. 2018, 76, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; He, J.; Lin, X.H.; Li, Q.; Pan, D.; Zhang, X.B.; Xu, X. Complete genome sequence of the shrimp white spot bacilliform virus. J. Virol. 2001, 75, 11811–11820. [Google Scholar] [CrossRef]
- 46th ESAO Congress 3–7 September 2019 Hannover, Germany. Available online: https://journals.sagepub.com/doi/abs/10.1177/0391398819860985?journalCode=jaoa (accessed on 25 March 2021).
- Momoyama, K.; Hiraoka, M.; Nakano, H.; Koube, H.; Inouye, K.; Oseko, N. Mass mortalities of cultured kuruma shrimp, penaeus-japonicus, in Japan in 1993—Histopathological study. Fish Pathol. 1994, 29, 141–148. [Google Scholar] [CrossRef]
- Jiang, H.F.; Chen, C.; Jiang, X.Y.; Shen, J.L.; Ling, F.; Li, P.F.; Wang, G.X. Luteolin in Lonicera japonica inhibits the proliferation of white spot syndrome virus in the crayfish Procambarus clarkii. Aquaculture 2022, 550, 737852. [Google Scholar] [CrossRef]
- Oakey, H.J.; Smith, C.S. Complete genome sequence of a white spot syndrome virus associated with a disease incursion in Australia. Aquaculture 2018, 484, 152–159. [Google Scholar] [CrossRef]
- Peeler, E.J. Costs and benefits of freedom from shrimp diseases in the European Union. J. Invertebr. Pathol. 2012, 110, 188–195. [Google Scholar] [CrossRef]
- Low, C.F.; Chong, C.M.; Loh, J.Y. Feasibility of vaccination against Macrobrachium rosenbergii nodavirus infection in giant freshwater prawn. Fish Shellfish Immunol. 2020, 104, 431–438. [Google Scholar]
- Kannan, S.; Kolandaivel, P. Antiviral potential of natural compounds against influenza virus hemagglutinin. Comput. Biol. Chem. 2017, 71, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, L.; Li, B.Y.; Shen, Y.F.; Wang, G.X.; Zhu, B. Synthesis of arctigenin derivatives against infectious hematopoietic necrosis virus. Eur. J. Med. Chem. 2019, 163, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Mathew, D.; Hsu, W.L. Antiviral potential of curcumin. J. Funct. Foods 2018, 40, 692–699. [Google Scholar] [CrossRef]
- Shen, Y.F.; Liu, L.; Chen, W.C.; Hu, Y.; Zhu, B.; Wang, G.X. Evaluation on the antiviral activity of arctigenin against spring viraemia of carp virus. Aquaculture 2018, 483, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Y.; Hu, Y.; Li, J.; Shi, K.; Shen, Y.F.; Zhu, B.; Wang, G.X. Ursolic acid from Prunella vulgaris L. efficiently inhibits IHNV infection in vitro and in vivo. Virus Res. 2019, 273, 197741. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.G.; Tan, X.P.; Qu, S.Y.; Wang, G.X.; Zhu, B. Evaluation on the antiviral activity of genipin against white spot syndrome virus in crayfish. Fish Shellfish Immunol. 2019, 93, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Toujani, M.M.; Ritta, M.; Civra, A.; Genovese, S.; Epifano, F.; Ghram, A.; Lembo, D.; Donalisio, M. Inhibition of HSV-2 infection by pure compounds from Thymus capitatus extract in vitro. Phytother. Res. 2018, 32, 1555–1563. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Lee, C.L.; Yen, H.R.; Chang, Y.S.; Lin, Y.P.; Huang, S.H.; Lin, C.W. Antiviral Action of Tryptanthrin Isolated from Strobilanthes cusia Leaf against Human Coronavirus NL63. Biomolecules 2020, 10, 366. [Google Scholar] [CrossRef]
- Kim, K.A.; Lee, I.A.; Gu, W.; Hyam, S.R.; Kim, D.H. beta-Sitosterol attenuates high-fat diet-induced intestinal inflammation in mice by inhibiting the binding of lipopolysaccharide to toll-like receptor 4 in the NF-kappa B pathway. Mol. Nutr. Food Res. 2014, 58, 963–972. [Google Scholar] [CrossRef]
- Guo, Q.S.; Duan, J.A.; He, S.A. Comparative study on three types of chemicals extracted from tubers of populations of Pinellia ternata (Thunb.) Breit. China J. Chin. Mater. Med. 2001, 26, 296–299. [Google Scholar]
- Chen, C.; Shen, J.L.; Wang, T.; Yang, B.; Liang, C.S.; Jiang, H.F.; Wang, G.X. Ophiopogon japonicus inhibits white spot syndrome virus proliferation in vivo and enhances immune response in Chinese mitten crab Eriocheir sinensis. Fish Shellfish Immunol. 2021, 119, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.G.; Tu, X.; Qi, X.Z.; Ling, F.; Zhu, B.; Wang, G.X. Gardenia jasminoides Ellis inhibit white spot syndrome virus replication in red swamp crayfish Procambarus clarkii. Aquaculture 2019, 504, 239–247. [Google Scholar] [CrossRef]
- Sun, Z.C.; Chen, C.; Xu, F.F.; Li, B.K.; Shen, J.L.; Wang, T.; Jiang, H.F.; Wang, G.X. Evaluation of the antiviral activity of naringenin, a major constituent of Typha angustifolia, against white spot syndrome virus in crayfish Procambarus clarkii. J. Fish. Dis. 2021, 44, 1503–1513. [Google Scholar] [CrossRef]
- Zhou, B.X.; Li, J.; Liang, X.L.; Pan, X.P.; Hao, Y.B.; Xie, P.F.; Jiang, H.M.; Yang, Z.F.; Zhong, N.S. beta-sitosterol ameliorates influenza A virus-induced proinflammatory response and acute lung injury in mice by disrupting the cross-talk between RIG-I and IFN/STAT signaling. Acta Pharmacol. Sin. 2020, 41, 1178–1196. [Google Scholar] [CrossRef]
- Chen, A.J.; Wang, S.A.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. Enzyme E2 from Chinese White Shrimp Inhibits Replication of White Spot Syndrome Virus and Ubiquitinates Its RING Domain Proteins. J. Virol. 2011, 85, 8069–8079. [Google Scholar] [CrossRef]
- Yao, D.F.; Ruan, L.W.; Lu, H.S.; Shi, H.; Xu, X. Shrimp STAT was hijacked by white spot syndrome virus immediate—Early protein IE1 involved in modulation of viral genes. Fish Shellfish Immunol. 2016, 59, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sanjuktha, M.; Raj, V.S.; Aravindan, K.; Alavandi, S.V.; Poornima, M.; Santiago, T.C. Comparative efficacy of double-stranded RNAs targeting WSSV structural and nonstructural genes in controlling viral multiplication in Penaeus monodon. Arch. Virol. 2012, 157, 993–998. [Google Scholar] [CrossRef]
- Mizushina, Y.; Nakanishi, R.; Kuriyama, I.; Kamiya, K.; Satake, T.; Shimazaki, N.; Koiwai, O.; Uchiyama, Y.; Yonezawa, Y.; Takemura, M.; et al. beta-sitosterol-3-O-beta-D-glucopyranoside: A eukaryotic DNA polymerase lambda inhibitor. J. Steroid Biochem. 2006, 99, 100–107. [Google Scholar] [CrossRef]
- Harwood, S.H.; Beckage, N.E. Purification and characterization of an early-expressed polydnavirus-induced protein from the hemolymph of manduca-sexta larvae parasitized by cotesia-congregata. Insect. Biochem. Molec. 1994, 24, 685–698. [Google Scholar] [CrossRef]
- Lo, C.F.; Ho, C.H.; Chen, C.H.; Liu, K.F.; Chiu, Y.L.; Yeh, P.Y.; Peng, S.E.; Hsu, H.C.; Liu, H.C.; Chang, C.F.; et al. Detection and tissue tropism of white spot syndrome baculovirus (WSBV) in captured brooders of Penaeus monodon with a special emphasis on reproductive organs. Dis. Aquat. Organ. 1997, 30, 53–72. [Google Scholar] [CrossRef] [Green Version]
- Ferrandon, D.; Imler, J.L.; Hetru, C.; Hoffmann, J.A. The Drosophila systemic immune response: Sensing and signalling during bacterial and fungal infections. Nat. Rev. Immunol. 2007, 7, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Li, F.H.; Xiang, J.H. Recent advances in researches on the innate immunity of shrimp in China. Dev. Comp. Immunol. 2013, 39, 11–26. [Google Scholar] [CrossRef]
- Sun, B.Z.; Wang, Z.; Zhu, F. The crustin-like peptide plays opposite role in shrimp immune response to Vibrio alginolyticus and white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2017, 66, 487–496. [Google Scholar] [CrossRef]
- Li, Y.Y.; Chen, X.X.; Lin, F.Y.; Chen, Q.F.; Ma, X.Y.; Liu, H.P. CqToll participates in antiviral response against white spot syndrome virus via induction of anti-lipopolysaccharide factor in red claw crayfish Cherax quadricarinatus. Dev. Comp. Immunol. 2017, 74, 217–226. [Google Scholar] [CrossRef]
- Huang, X.D.; Zhao, L.; Zhang, H.Q.; Xu, X.P.; Jia, X.T.; Chen, Y.H.; Wang, P.H.; Weng, S.P.; Yu, X.Q.; Yin, Z.X.; et al. Shrimp NF-kappa B binds to the immediate-early gene ie1 promoter of white spot syndrome virus and upregulates its activity. Virology 2010, 406, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Gu, Z.H.; Wan, D.H.; Zhang, M.Y.; Weng, S.P.; Yu, X.Q.; He, J.G. The Shrimp NF-kappa B Pathway Is Activated by White Spot Syndrome Virus (WSSV) 449 to Facilitate the Expression of WSSV069 (ie1), WSSV303 and WSSV371. PLoS ONE 2011, 6, e24773. [Google Scholar]
- Liu, L.K.; Liu, M.J.; Li, D.L.; Liu, H.P. Recent insights into anti-WSSV immunity in crayfish. Dev. Comp. Immunol. 2021, 116, 103947. [Google Scholar] [CrossRef]
- Song, L.; Lv, J.J.; Wang, L.; Sun, D.F.; Gao, B.Q.; Liu, P. Characterization of a chitinase-1 gene (PtCht-1) from a marine crab Portunus trituberculatus and its response to immune stress. Gene 2020, 741, 144523. [Google Scholar] [CrossRef]
- Niu, S.W.; Yang, L.W.; Geng, R.; Zuo, H.L.; Guo, Z.X.; Weng, S.P.; He, J.G.; Xu, X.P. A double chitin catalytic domain-containing chitinase targeted by c-Jun is involved in immune responses in shrimp. Dev. Comp. Immunol. 2020, 113, 103808. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, C.Z.; Ma, C.X.; Li, H.Y.; Zuo, H.L.; Weng, S.P.; Chen, X.H.; Zeng, D.G.; He, J.G.; Xu, X.P. Identification of a C-type lectin with antiviral and antibacterial activity from pacific white shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2014, 46, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zheng, Q.; Yu, C.; Pan, C.K.; Luo, P.; Chen, J.M. WSSV056 inhibits shrimp nitric oxide synthase activity by downregulating Litopenaeus vannamei sepiapterin reductase to promote white spot syndrome virus replication. Front. Microbiol. 2021, 12, 796049. [Google Scholar] [CrossRef] [PubMed]
- Li, S.H.; Wang, Z.W.; Li, F.H.; Yu, K.J.; Xiang, J.H. A Novel Vascular Endothelial Growth Factor Receptor Participates in White Spot Syndrome Virus Infection in Litopenaeus vannamei. Front. Immunol. 2017, 8, 1457. [Google Scholar] [CrossRef]
- Wang, Z.W.; Li, S.H.; Li, F.H.; Xie, S.J.; Xiang, J.H. Identification and function analysis of a novel vascular endothelial growth factor, LvVEGF3, in the Pacific whiteleg shrimp Litopenaeus vannamei. Dev. Comp. Immunol. 2016, 63, 111–120. [Google Scholar] [CrossRef]
- Taguwa, S.; Maringer, K.; Li, X.K.; Bernal-Rubio, D.; Rauch, J.N.; Gestwicki, J.E.; Andino, R.; Fernandez-Sesma, A.; Frydman, J. Defining Hsp70 Subnetworks in Dengue Virus Replication Reveals Key Vulnerability in Flavivirus Infection. Cell 2015, 163, 1108–1123. [Google Scholar] [CrossRef]
- Lin, Y.R.; Hung, H.C.; Leu, J.H.; Wang, H.C.; Kou, G.H.; Lo, C.F. The Role of Aldehyde Dehydrogenase and Hsp70 in Suppression of White Spot Syndrome Virus Replication at High Temperature. J. Virol. 2011, 85, 3517–3525. [Google Scholar] [CrossRef]
- Liu, L.; Hu, Y.; Shen, Y.F.; Wang, G.X.; Zhu, B. Evaluation on antiviral activity of coumarin derivatives against spring viraemia of carp virus in epithelioma Papulosum cyprini cells. Antivir. Res. 2017, 144, 173–185. [Google Scholar] [CrossRef]
- Duan, Y.F.; Liu, P.; Li, J.T.; Li, J.; Chen, P. Expression profiles of selenium dependent glutathione peroxidase and glutathione S-transferase from Exopalaemon carinicauda in response to Vibrio anguillarum and WSSV challenge. Fish Shellfish Immunol. 2013, 35, 661–670. [Google Scholar] [CrossRef]
- Miranda-Cruz, M.M.; Poom-Llamas, J.J.; Godoy-Lugo, J.A.; Ortiz, R.M.; Gomez-Jimenez, S.; Rosas-Rodriguez, J.A.; Moran-Palacio, E.F.; Sonanez-Organis, J.G. Silencing of HIF-1 in WSSV-infected white shrimp: Effect on viral load and antioxidant enzymes. Comp. Biochem. Phys. C 2018, 213, 19–26. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.Q. BAX, a novel cell pro-apoptotic protein, involved in hemocytes early antiviral immune response in fresh water crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2016, 55, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.Q.; Lan, J.F.; Weng, Y.D.; Zhao, X.F.; Wang, J.X. BAX inhibitor-1 silencing suppresses white spot syndrome virus replication in red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2013, 35, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Sharmila, R.; Sindhu, G. Modulation of Angiogenesis, Proliferative Response and Apoptosis by beta-Sitosterol in Rat Model of Renal Carcinogenesis. Indian J. Clin. Biochem. IJCB 2017, 32, 142–152. [Google Scholar] [CrossRef]
- Liu, W.J.; Chang, Y.S.; Wang, A.H.J.; Kou, G.H.; Lo, C.F. White spot syndrome virus annexes a shrimp STAT to enhance expression of the immediate-early gene ie1. J. Virol. 2007, 81, 1461–1471. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Accession No. | Primer Sequences (from 5′ to 3′) | Size (bp) |
|---|---|---|---|
| VP28-F | KT995472.1 | AAACCTCCGCATTCCTGTGA | 141 |
| VP28-R | TCCGCATCTTCTTCCTTCAT | ||
| ie1-F | KT995472.1 | GACTCTACAAATCTCTTTGCCA | 283 |
| ie1-R | TGCTGATAAACTCTTGAAGGAA | ||
| DNApol-F | KT995472.1 | CTCGCCAAAGTGAGTAGTGT | 178 |
| DNApol-R | CCTTGTTGATGGAGGTAGAA | ||
| Crustin 1-F | GQ301201.1 | CCACAGATGGCAATCGGAGTC | 131 |
| Crustin 1-R | AGGGAACGAACGCTGGAAAGT | ||
| NF-κappa B-F | KF662471.1 | TAGTGCGTGATGATGGGTCTT | 136 |
| NF-κappa B-R | GCTGATTATGGAGGCAGAAAA | ||
| STAT-F | TGGTAGTGAAGAGAGGTTGAG | 97 | |
| STAT-R | CATTGTTTCCCATCTGTCC | ||
| TLR2-F | KP259728.1 | AAGTCACTACGCAAACCA | 102 |
| TLR2-R | TACCACCATTTAGAGTAGACC | ||
| ALF1-F | KU680792.1 | CGGTTGGCGCCTCTACTACA | 102 |
| ALF1-R | GCGTGCTCGATGGCTCCTG | ||
| CHI-F | FR990062.1 | AATGGTGCTCAACCTCCT | 151 |
| CHI-R | CTGCGCTAAAGAAACAGAA | ||
| CTL-F | KC857544.1 | ACTTTGCTAACGCCAATCCAC | 204 |
| CTL-R | CTACGCTGTCATCGACGAACC | ||
| VEGFR-F | AGTCGCCAGGAACCAGTG | ||
| VEGFR-R | TGCCGAACCTAATGAAGATA | ||
| Hsp70-F | DQ301506.1 | GTTGACCAAGATGAAGGAGAC | 100 |
| Hsp70-R | CTGACGCTGAGAGTCGTTG | ||
| NOS-F | TAATCCTTGACGGTGGTG | ||
| NOS-R | TTGGCATCTTTCTTCTTCTC | ||
| COX-1-F | KX268742.1 | ATGGGATACCTCGACGTTATTC | 202 |
| COX-1-R | GCAGGAGGATAAGAATGCTGT | ||
| COX-2-F | AF437613.1 | GGTCATCAGTGATATTGAAGG | 110 |
| COX-2-R | TCTAATAAACGGAACCCAGAC | ||
| BAX-F | TATAGTTGGCTCATTAGCAG | 196 | |
| BAX-R | ATACTAAGTGAAGATGACTG | ||
| BI-1-F | TGCCATTACATCTTGGGTTCT | 157 | |
| BI-1-R | CGACCTAATCCCATCTCAAGC | ||
| cMnSOD-F | EU254488.3 | GCCACCACTAAAATACGAGTA | 192 |
| cMnSOD-R | CCATTGAACTTTATAGCTGGTA | ||
| mMnSOD-F | KC333178.1 | CATCACTCCAAGCACCACC | 109 |
| mMnSOD-R | GAGCAAGGGATATAACAGTAC | ||
| CAT-F | KM068092.1 | CGACCATACACCGCTTCAC | 250 |
| CAT-R | TTTCAGGAATGCGTTCTCTATC | ||
| GST-F | HQ414581.1 | ACTTAGAGACGGACTTCCAG | 96 |
| GST-R | CGAGGGCGAACTTCACGG | ||
| 18S-F | KX444578.1 | ACCGATTGAATGATTTAGTGAG | 153 |
| 18S-R | TACGGAAACCTTGTTACGAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Shen, J.-L.; Liang, C.-S.; Sun, Z.-C.; Jiang, H.-F. First Discovery of Beta-Sitosterol as a Novel Antiviral Agent against White Spot Syndrome Virus. Int. J. Mol. Sci. 2022, 23, 10448. https://doi.org/10.3390/ijms231810448
Chen C, Shen J-L, Liang C-S, Sun Z-C, Jiang H-F. First Discovery of Beta-Sitosterol as a Novel Antiviral Agent against White Spot Syndrome Virus. International Journal of Molecular Sciences. 2022; 23(18):10448. https://doi.org/10.3390/ijms231810448
Chicago/Turabian StyleChen, Cheng, Jing-Lei Shen, Chang-Shuai Liang, Zhong-Chen Sun, and Hai-Feng Jiang. 2022. "First Discovery of Beta-Sitosterol as a Novel Antiviral Agent against White Spot Syndrome Virus" International Journal of Molecular Sciences 23, no. 18: 10448. https://doi.org/10.3390/ijms231810448