
Microscopic and Transcriptomic Comparison of Powdery Mildew Resistance in the Progenies of Brassica carinata × B. napus
 ,
, 
Abstract
:1. Introduction
2. Results
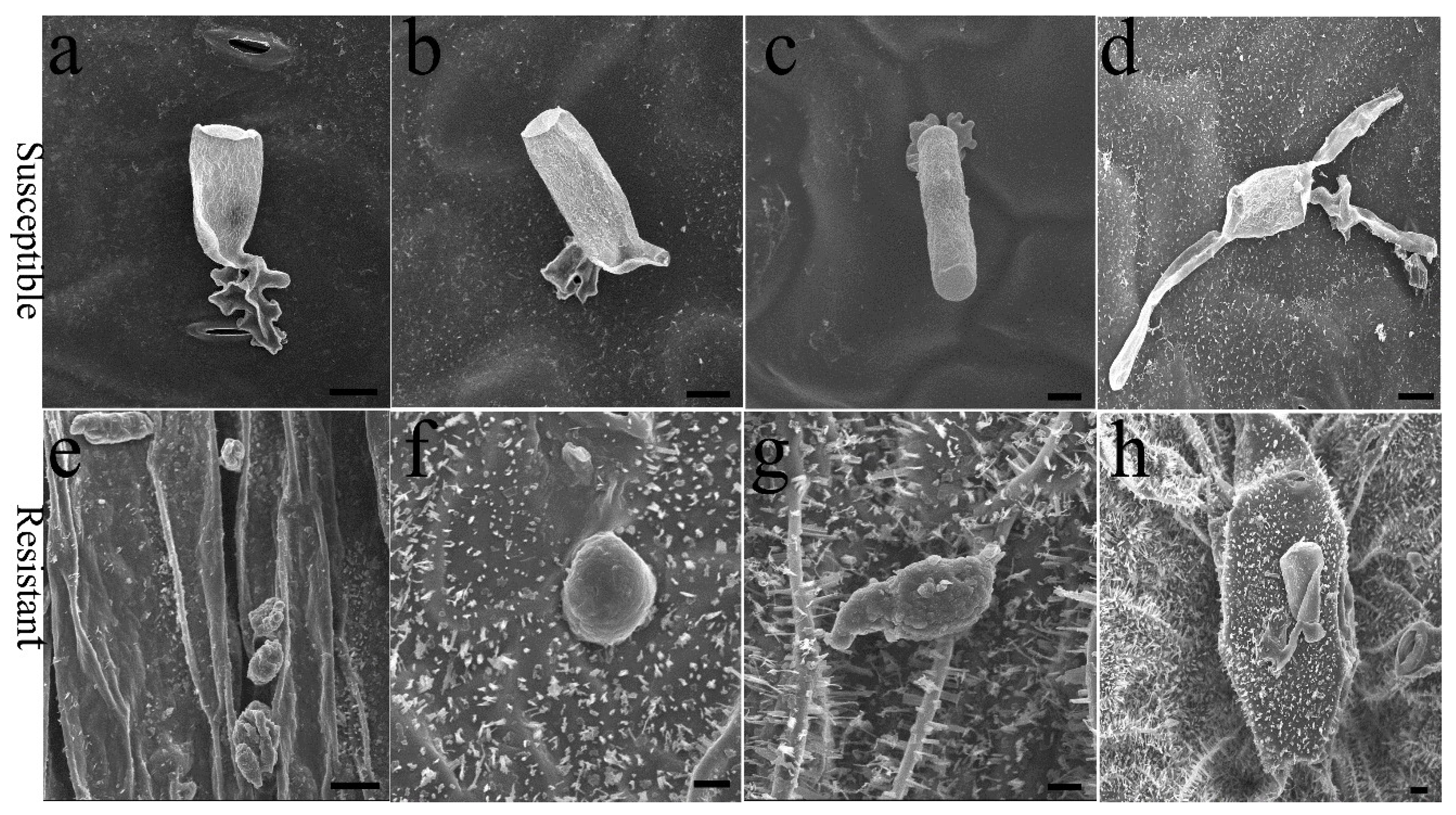
2.1. E. cruciferarum Pathogenesis on the Plant Accessions
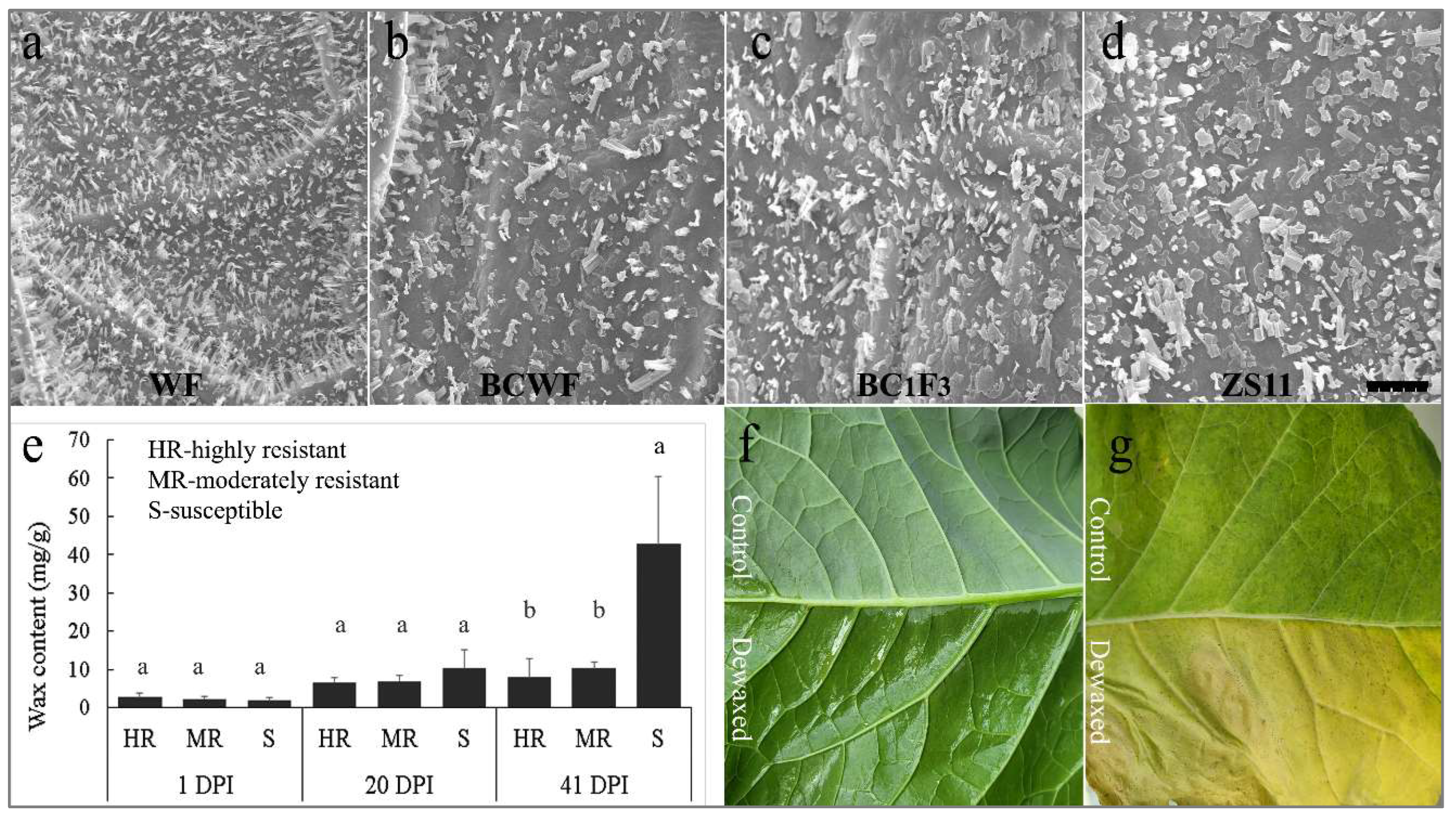
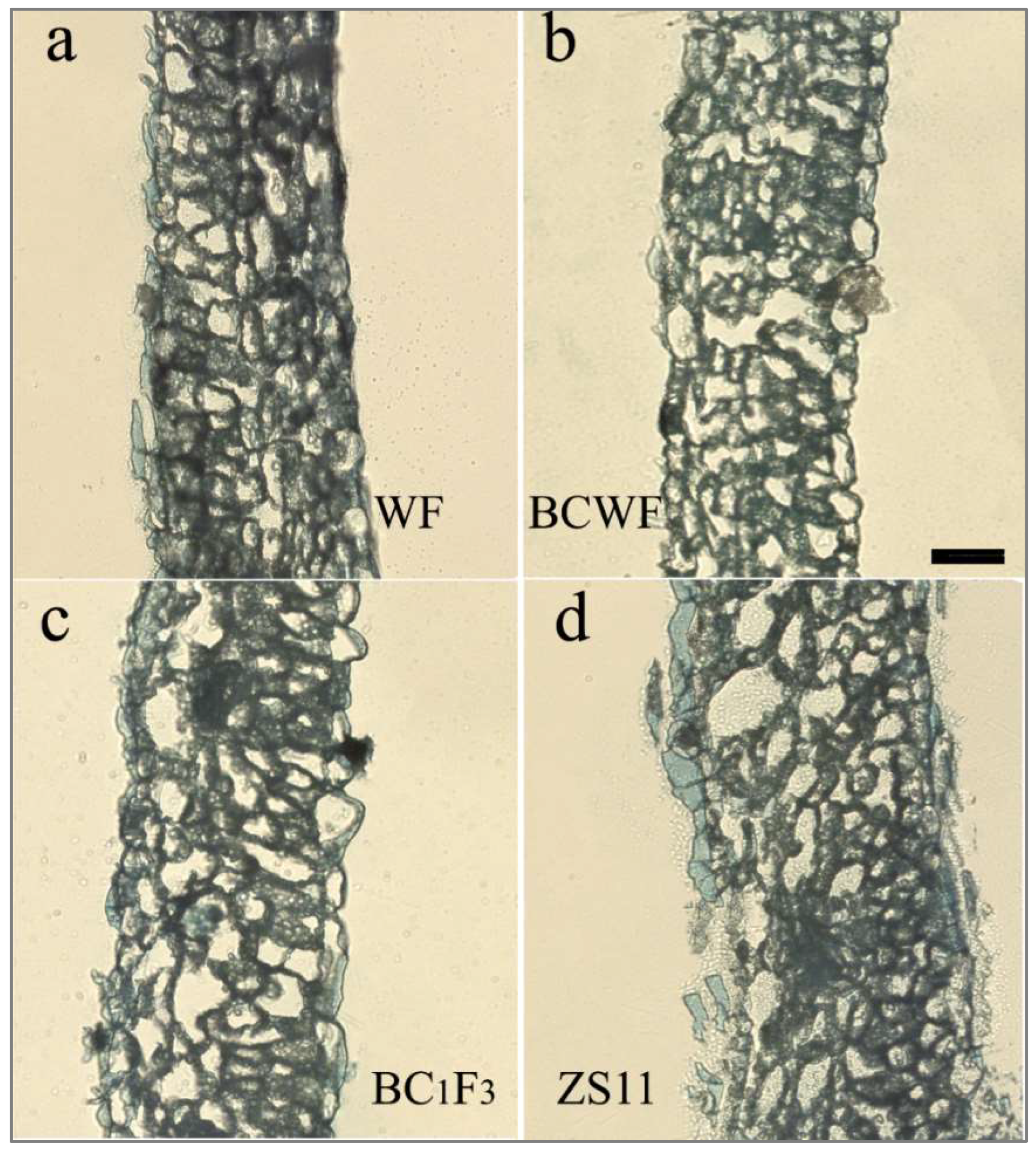
2.2. Difference in Foliar Cuticular Wax of the Plant Accessions
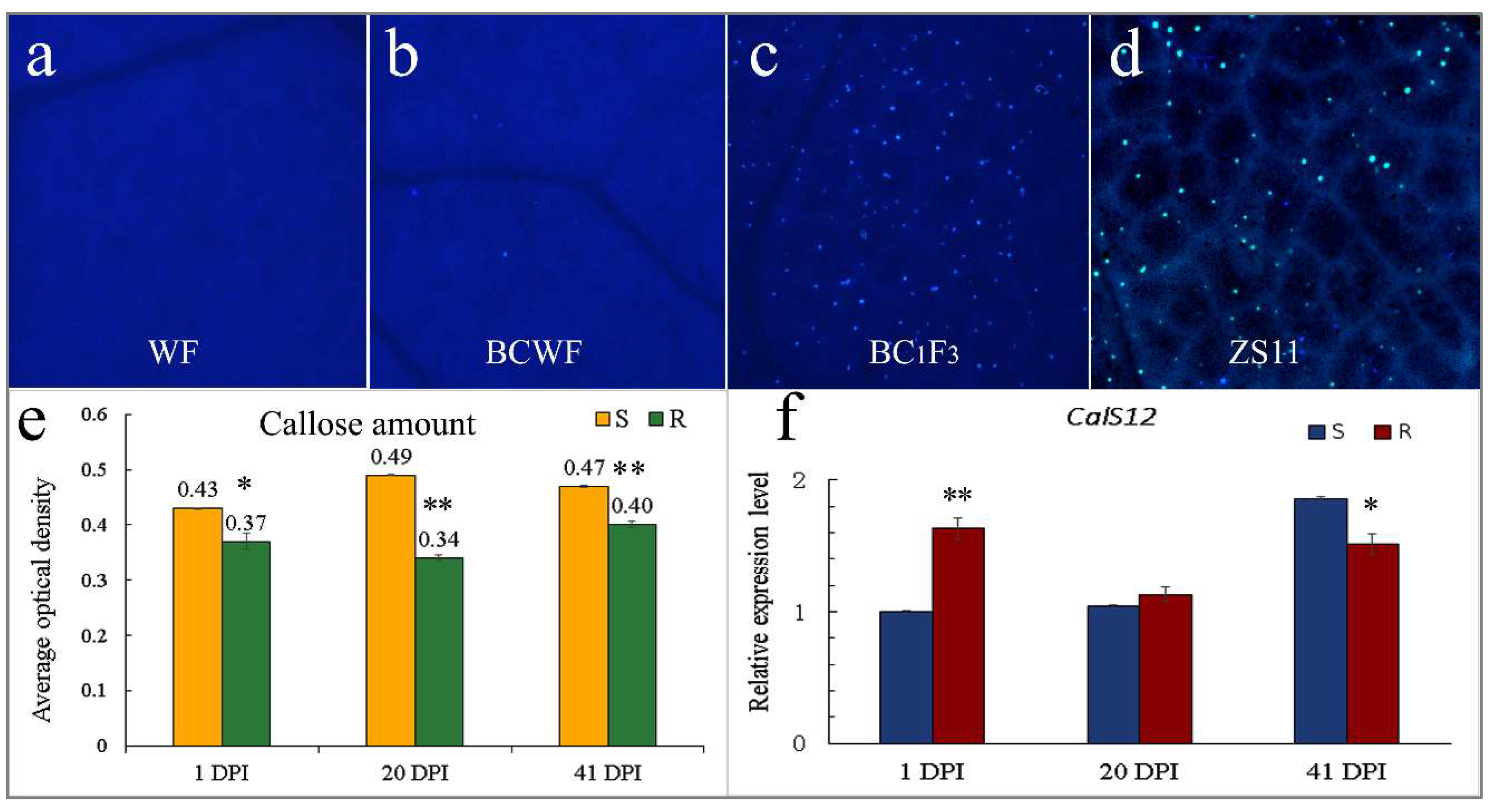
2.3. Callose Deposition on the E. cruciferarum Penetration Site
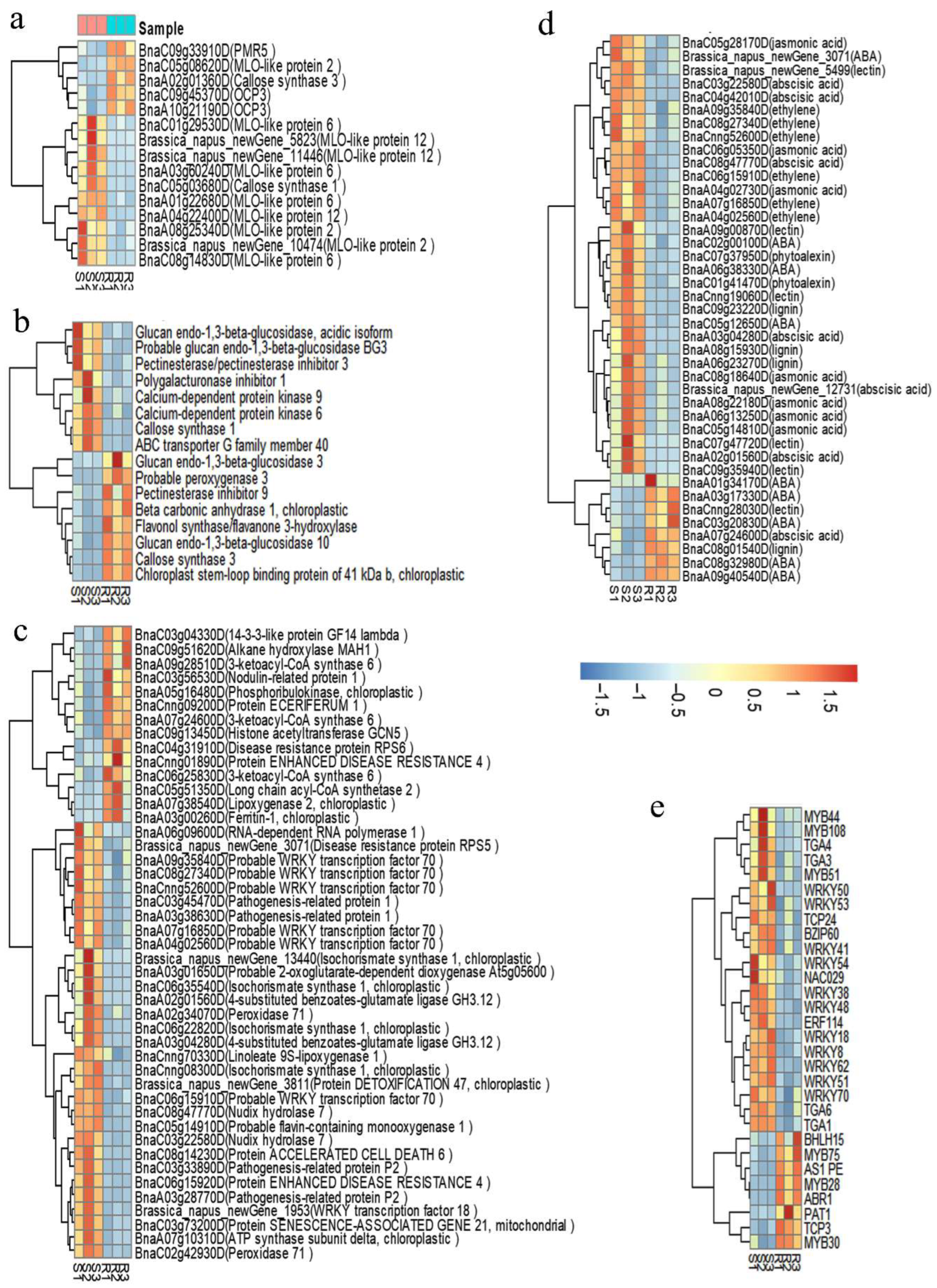
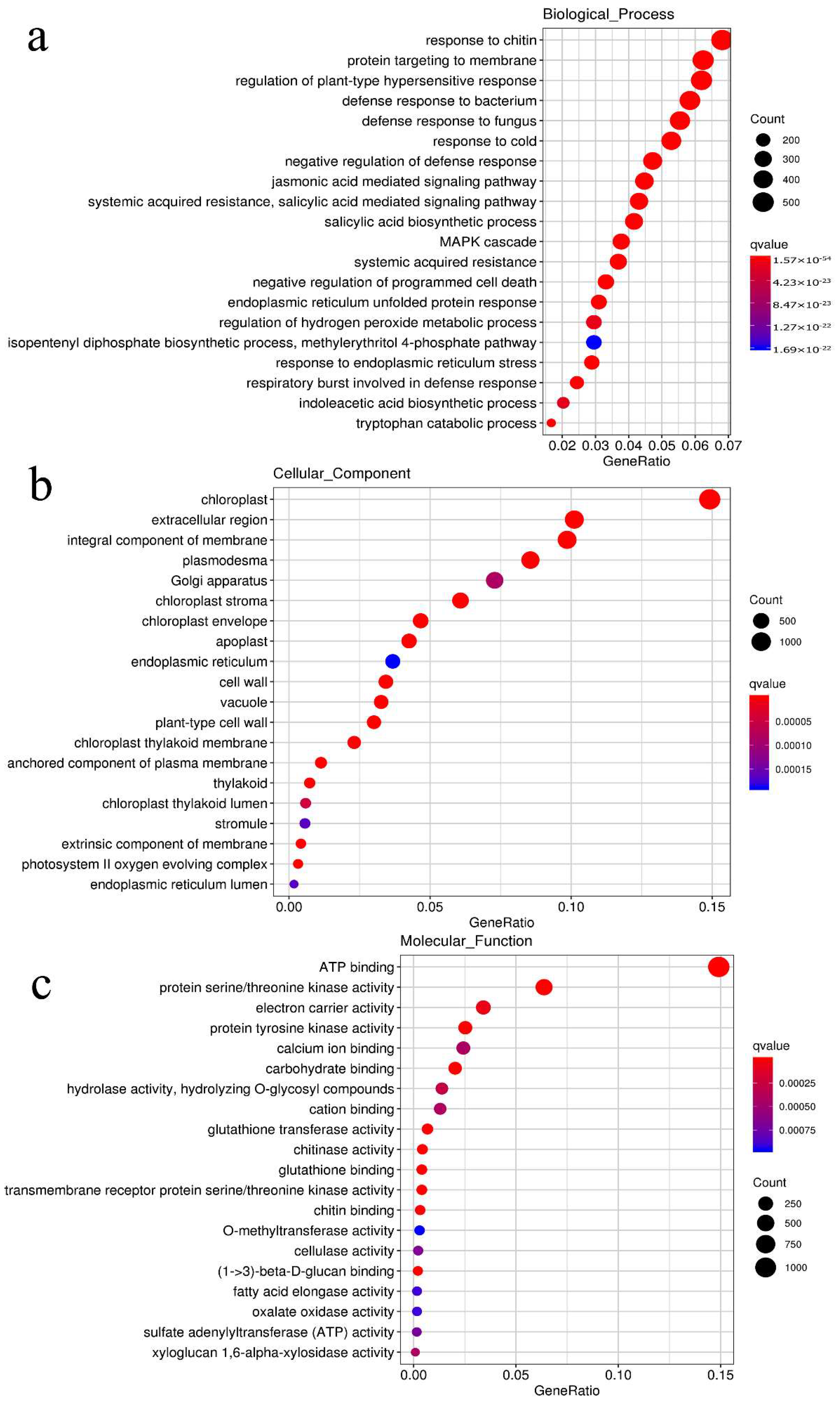
2.4. Transcriptomic Comparison of the S/R Plants in the Segregated Population
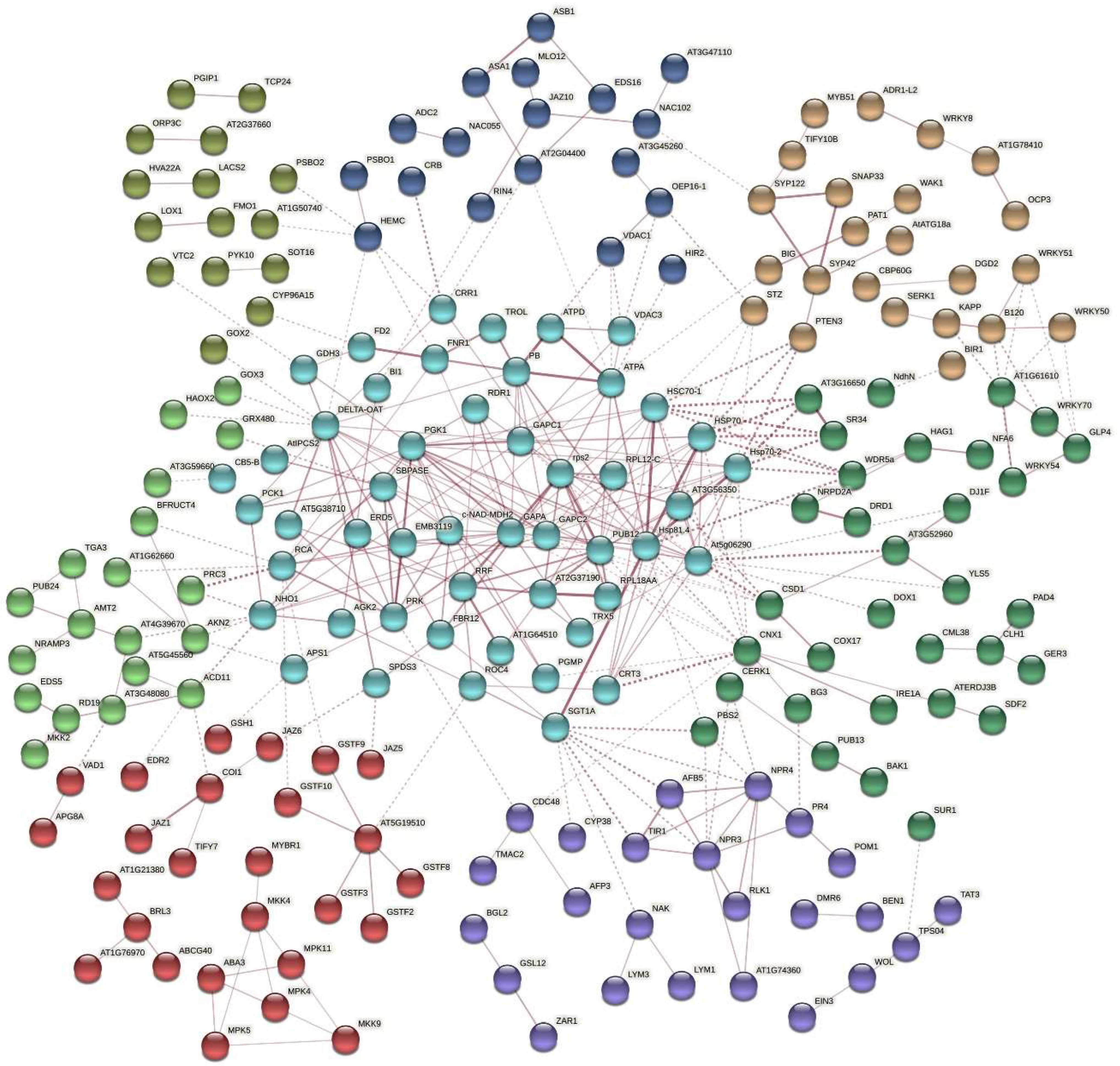
2.5. Protein Interaction Network Estimated by the Differentially Expressed Genes
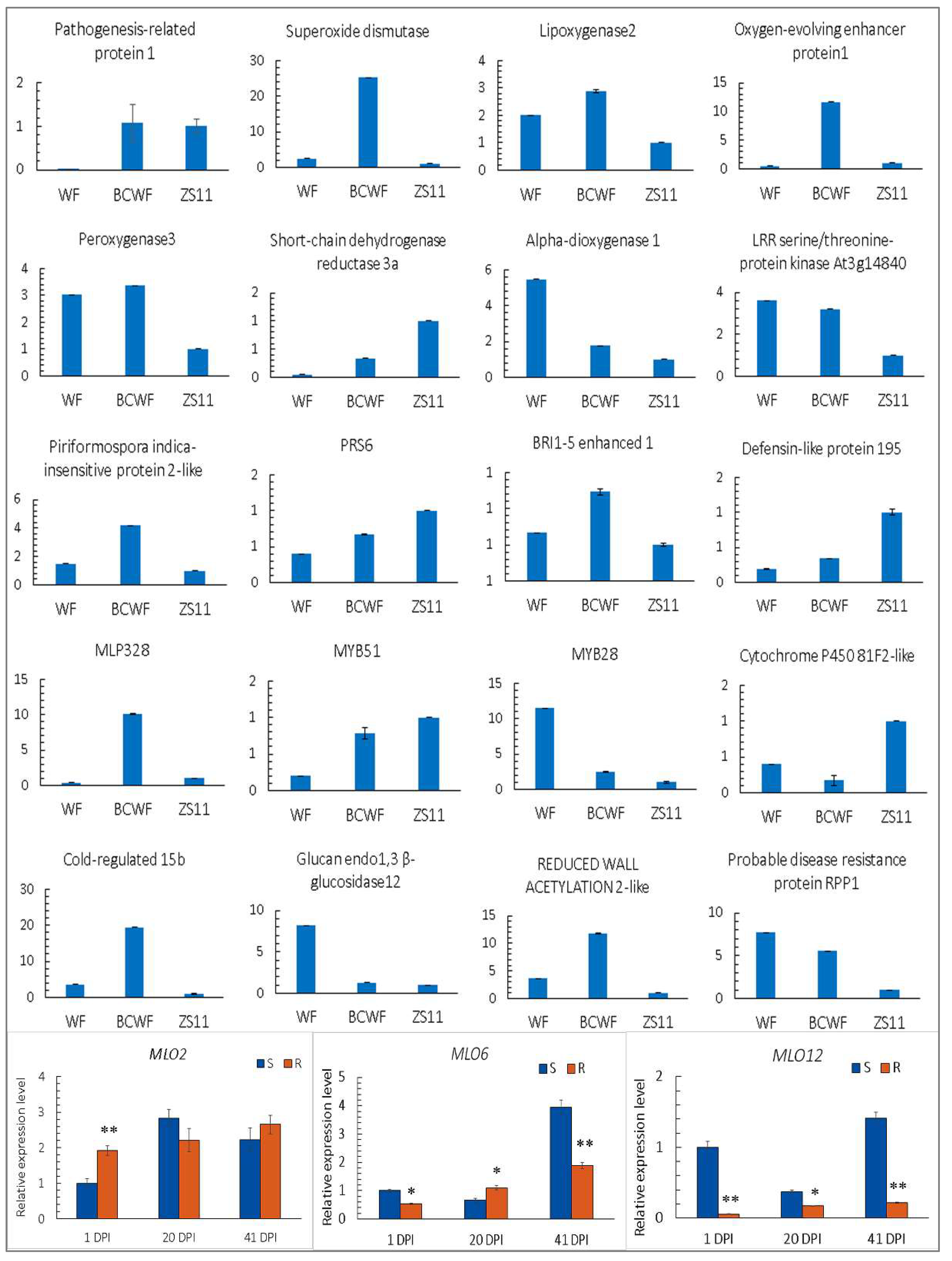
2.6. The Expression Level of the Selected Genes Detected by qPCR
3. Discussion
3.1. The Possible Contribution of Cuticular Wax and Callose to PM Resistance
3.2. Influence of the Composition and Architecture of Plant Cell Wall on PM Infection
3.3. HR and Systemic Acquired Resistance Are Important for the Establishment of Moderate Resistance in Rapeseed Plants
4. Materials and Methods
4.1. Plant Materials
4.2. Microscopic Observation
4.3. Assay of The Leaf Cuticular Wax
4.4. Determination of Callose Content in Leaves
4.5. Real-Time Quantitative PCR (qPCR)
4.6. Transcriptomic Comparison between the Susceptible and Resistant Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix B

Appendix C

References
- Saharan, G.S.; Mehta, N.K.; Meena, P.D. The disease: Powdery mildew. In Powdery Mildew Disease of Crucifers: Biology, Ecology and Disease Management; Springer Nature: Singapore, 2019; pp. 17–51. [Google Scholar] [CrossRef]
- Gong, Q.; Dai, C.Y.; Zhang, X.H.; Wang, X.L.; Huang, Z.; Xu, A.X.; Dong, J.G.; Yu, C.Y. Towards breeding of rapeseed (Brassica napus) with alien cytoplasm and powdery mildew resistance from Ethiopian mustard (Brassica carinata). Breed. Sci. 2020, 70, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Blümke, A.; Voigt, C.A. Secreted fungal effector lipase releases free fatty acids to inhibit innate immunity-related callose formation during wheat head infection. Plant Physiol. 2014, 165, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Forderer, A.; Yu, D.; Chai, J. Structural biology of plant defence. New Phytol. 2021, 229, 692–711. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Nguyen, C.T.; Liang, Y.; Cao, Y.; Stacey, G. Role of LysM receptors in chitin- triggered plant innate immunity. Plant Signal. Behav. 2013, 8, e22598. [Google Scholar] [CrossRef]
- Cao, Y.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. eLife 2014, 3, e03766. [Google Scholar] [CrossRef]
- Chowdhury, J.; Henderson, M.; Schweizer, P.; Burton, R.A.; Fincher, G.B.; Little, A. Differential accumulation of callose, arabinoxylan and cellulose in nonpenetrated versus penetrated papillae on leaves of barley infected with Blumeria graminis f. sp. hordei. New Phytol. 2014, 204, 650–660. [Google Scholar] [CrossRef]
- Blümke, A.; Somerville, S.C.; Voigt, C.A. Transient expression of the Arabidopsis thaliana callose synthase PMR4 increases penetration resistance to powdery mildew in barley. Adv. Biosci. Biotechnol. 2013, 4, 810–813. [Google Scholar] [CrossRef]
- Ellinger, D.; Naumann, M.; Falter, C.; Zwikowics, C.; Jamrow, T.; Manisseri, C.; Somerville, S.C.; Voigt, C.A. Elevated early callose deposition results in complete penetration resistance to powdery mildew in Arabidopsis. Plant Physiol. 2013, 161, 1433–1444. [Google Scholar] [CrossRef]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.M.; He, S.Y.; Xin, X.F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Kusch, S.; Panstruga, R. mlo-based resistance: An apparently universal “weapon” to defeat powdery mildew disease. Mol. Plant Microbe Interact. 2017, 30, 179–189. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, Y.; Wu, G.; Zou, S.; Chen, Y.; Gao, C.; Tang, D. Simultaneous modification of three homoeologs of TaEDR1 by genome editing enhances powdery mildew resistance in wheat. Plant J. 2017, 91, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.; Kwaaitaal, M.; Kusch, S.; Acevedo-Garcia, J.; Wu, H.; Panstruga, R. Biotrophy at its best: Novel findings and unsolved mysteries of the Arabidopsis-powdery mildew pathosystem. Arabidopsis Book 2016, 14, e0184. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P.; Raab, T.K.; Somerville, C.R.; Somerville, S.C. Mutations in PMR5 result in powdery mildew resistance and altered cell wall composition. Plant J. 2004, 40, 968–978. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Zhang, L.L.; Ma, X.F.; Zhao, Z.X.; Zhao, J.H.; Zhao, J.Q.; Zhao, J.Q.; Fan, J.; Li, Y.; He, P.; et al. Multiple intramolecular trafficking signals in Resistance to Powdery Mildew 8.2 are engaged in activation of cell death and defense. Plant J. 2019, 2019 98, 55–70. [Google Scholar] [CrossRef]
- Moore, D.; Robson, G.D.; Trinci, A.P.J. Where to find more information about crop diseases, crop losses and plant pathogens. In 21st Century Guidebook to Fungi, 2nd ed.; Cambridge University Press: London, UK, 2020; p. 600. ISBN 9781108745680. [Google Scholar]
- Göllner, K.; Schweizer, P.; Bai, Y.; Panstruga, R. Natural genetic resources of Arabidopsis thaliana reveal a high prevalence and unexpected phenotypic plasticity of RPW8-mediated powdery mildew resistance. New Phytol. 2008, 177, 725–742. [Google Scholar] [CrossRef] [PubMed]
- Shinya, T.; Nakagawa, T.; Kaku, H.; Shibuya, N. Chitin-mediated plant-fungal interactions: Catching, hiding and handshaking. Curr. Opin. Plant Biol. 2015, 26, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, M.; Lironi, D.; Marti, L.; Paparella, C.; Vecchi, V.; Gust, A.A.; De Lorenzo, G.; Nürnberger, T.; Ferrari, S. The Arabidopsis thaliana LysM-containing Receptor-Like Kinase 2 is required for elicitor-induced resistance to pathogens. Plant Cell Environ. 2021, 44, 3545–3562. [Google Scholar] [CrossRef]
- Gao, M.; Liu, J.; Bi, D.; Zhang, Z.; Cheng, F.; Chen, S.; Zhang, Y. MEKK1, MKK1/MKK2 and MPK4 function together in a mitogen-activated protein kinase cascade to regulate innate immunity in plants. Cell Res. 2008, 18, 1190–1198. [Google Scholar] [CrossRef]
- Yang, X.; Wang, W.; Coleman, M.; Orgil, U.; Feng, J.; Ma, X.; Ferl, R.; Turner, J.G.; Xiao, S. Arabidopsis 14-3-3 lambda is a positive regulator of RPW8-mediated disease resistance. Plant J. 2009, 60, 539–550. [Google Scholar] [CrossRef]
- Wirthmueller, L.; Zhang, Y.; Jones, J.D.; Parker, J.E. Nuclear accumulation of the Arabidopsis immune receptor RPS4 is necessary for triggering EDS1-dependent defense. Curr. Biol. 2007, 17, 2023–2029. [Google Scholar] [CrossRef]
- Ray, S.K.; Macoy, D.M.; Kim, W.Y.; Lee, S.Y.; Kim, M.G. Role of RIN4 in regulating PAMP-triggered immunity and effector-triggered immunity: Current status and future perspectives. Mol. Cells 2019, 42, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.M.; Trifa, Y.; Silva, H.; Pontier, D.; Lam, E.; Shah, J.; Klessig, D.F. NPR1 differentially interacts with members of the TGA/OBF family of transcription factors that bind an element of the PR-1 gene required for induction by salicylic acid. Mol. Plant Microbe Interact. 2000, 13, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Suh, M.H. Advances in the understanding of cuticular waxes in Arabidopsis thaliana and crop species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Ensikat, H.J. The hydrophobic coatings of plant surfaces: Epicuticular wax crystals and their morphologies, crystallinity and molecular self-assembly. Micron 2008, 39, 759–772. [Google Scholar] [CrossRef]
- Greer, S.; Wen, M.; Bird, D.; Wu, X.; Samuels, L.; Kunst, L.; Jetter, R. The cytochrome P450 enzyme CYP96A15 is the midchain alkane hydroxylase responsible for formation of secondary alcohols and ketones in stem cuticular wax of Arabidopsis. Plant Physiol. 2007, 145, 653–667. [Google Scholar] [CrossRef]
- Manabe, Y.; Verhertbruggen, Y.; Gille, S.; Harholt, J.; Chong, S.L.; Pawar, P.M.; Mellerowicz, E.J.; Tenkanen, M.; Cheng, K.; Pauly, M.; et al. RWA proteins play vital and distinct roles in cell wall O-Acetylation in Arabidopsis Thaliana. Plant Physiol. 2013, 163, 1107–1117. [Google Scholar] [CrossRef]
- Pavet, V.; Olmos, E.; Kiddle, G.; Mowla, S.; Kumar, S.; Antoniw, J.; Alvarez, M.E.; Foyer, C.H. Ascorbic acid deficiency activates cell death and disease resistance responses in Arabidopsis. Plant Physiol. 2005, 139, 1291–1303. [Google Scholar] [CrossRef]
- Li, L.; Li, K.; Ali, A.; Guo, Y. AtWAKL10, a cell wall associated receptor-like kinase, negatively regulates leaf senescence in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 4885. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, Y.; Zhan, Y.; Kui, H.; Liu, H.; Yan, L.; Kemmerling, B.; Zhou, J.; He, K.; Li, J. Loss of the common immune coreceptor BAK1 leads to NLR-dependent cell death. Proc. Natl. Acad. Sci. USA 2020, 117, 27044–27053. [Google Scholar] [CrossRef]
- Zeng, H.Y.; Liu, Y.; Chen, D.K.; Bao, H.N.; Huang, L.Q.; Yin, J.; Chen, Y.L.; Xiao, S.; Yao, N. The immune components enhanced disease susceptibility 1 and phytoalexin deficient 4 are required for cell death caused by overaccumulation of ceramides in Arabidopsis. Plant J. 2021, 107, 1447–1465. [Google Scholar] [CrossRef]
- Brodersen, P.; Petersen, M.; Bjorn Nielsen, H.; Zhu, S.; Newman, M.A.; Shokat, K.M.; Rietz, S.; Parker, J.; Mundy, J. Arabidopsis MAP kinase 4 regulates salicylic acid- and jasmonic acid/ethylene-dependent responses via EDS1 and PAD4. Plant J. 2006, 47, 532–546. [Google Scholar] [CrossRef]
- Ge, X.; Li, G.J.; Wang, S.B.; Zhu, H.; Zhu, T.; Wang, X.; Xia, Y. AtNUDT7, a negative regulator of basal immunity in Arabidopsis, modulates two distinct defense response pathways and is involved in maintaining redox homeostasis. Plant Physiol. 2007, 145, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Weigel, R.R.; Pfitzner, U.M.; Gatz, C. Interaction of NIMIN1 with NPR1 modulates PR gene expression in Arabidopsis. Plant Cell 2005, 17, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.; Greene, G.H.; Yuan, M.; Xu, G.; Burton, D.; Liu, L.; Marques, J.; Dong, X. Translational regulation of metabolic dynamics during effector-triggered immunity. Mol. Plant 2020, 13, 88–98. [Google Scholar] [CrossRef]
- Zhao, Z.X.; Feng, Q.; Liu, P.Q.; He, X.R.; Zhao, J.H.; Xu, Y.J.; Zhang, L.L.; Huang, Y.Y.; Zhao, J.Q.; Fan, J.; et al. RPW8.1 enhances the ethylene-signaling pathway to feedback-attenuate its mediated cell death and disease resistance in Arabidopsis. New Phytol 2020, 229, 516–531. [Google Scholar] [CrossRef]
- Solomon, M.; Belenghi, B.; Delledonne, M.; Menachem, E.; Levine, A. The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 1999, 11, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Ponce De León, I.; Sanz, A.; Hamberg, M.; Castresana, C. Involvement of the Arabidopsis alpha-DOX1 fatty acid dioxygenase in protection against oxidative stress and cell death. Plant J. 2002, 29, 61–62. [Google Scholar] [CrossRef]
- Wu, G.; Liu, S.; Zhao, Y.; Wang, W.; Kong, Z.; Tang, D. Enhanced disease Resistance4 associates with clathrin heavy chain2 and modulates plant immunity by regulating relocation of EDR1 in Arabidopsis. Plant Cell 2015, 27, 857–873. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Annotation | Primers Pair (5′→3′) |
|---|---|---|
| ACTIN7 | Beta Actin 7 |
CATCCCTCAGCACCTTCC CCAACCTTAGCACTTCTCC |
| CalS12 | Callose synthase 12 | GTGGCATCAGTTCTTTGGCG AGCTTCAATCTATGAATGGCGTC |
| MLO2 | Mildew Resistance Locus 2 |
AAGAAGCACAAGCAGGCTCT TGCAAGGGTGCATTGTTGAC |
| MLO6 | Mildew Resistance Locus 6 |
CTTCTGGTTCGGTCGTCCAC TTAGACCGAATTCATACGTACTCC |
| MLO12 | Mildew Resistance Locus 12 |
GAACCGGGCGATGATCTCTT GCCAGTTGAAATGCATTCGTG |
| BnaC03g45470D | PR1, pathogenesis-related protein 1 |
TCACAACCAAGCACGACAGG GTTCTCTCCATAAGGCCCACC |
| BnaC08g42970D | Superoxide dismutase [Cu-Zn] |
CACCCATGAAGGAAACGGTG CGATTAGCATCCTCCGGTGC |
| BnaA02g11430D | Lox2, lipoxygenase 2, chloroplastic-like |
CCGTCTGTGAACAAAGTGAGAG ACAATGAGTCCTCAGCCAGT |
| BnaC07g46350D | OEE1 |
TCACCGTCAAAGCAGAAGGT CTGTCACTGCAGCGTAATCT |
| BnaA03g40190D | MYB28-like |
CAATGCCTTCCCTGTCTCGT CCAATCTGCTCAGAGAAGCCA |
| BnaA05g10200D | Probable peroxygenase 3 |
GTACGTAGCGACTTGGAGGAG GGGTAGACAATACCGTCGCC |
| BnaA05g24690D | Probable LRR-RLK At3g14840 |
GGTCCAATTCCTCCCGAATG TGAAATCAAGGCCAAGGAACT |
| BnaA01g31910D | MLP-like 328 |
ACATCTTCCCTGACGCCATC ACGTGACCCTCAAGTCCTCT |
| BnaA07g27410D | PII-2, Piriformospora indica-insensitive protein 2-like |
ATGGAGAAAACAGAGAAAGCTGC AGATCACAAGTTACACCCTGGA |
| BnaA03g56740D | Cold-regulated 15b |
ATTCTTCTTTCCCCAGCGGC CGCGTAATCCGAAGCTCTCT |
| BnaC04g53100D | BEN1, BRI1-5 ENHANCED 1-like |
CCCAGTTTCAGCTACCTTCAGT ATTTTTAACCCCCAACATAAAGAGT |
| BnaC06g11870D | Probable disease resistance protein RPP1 |
CGCCAACAAGAGATTTCGAGG ACATATTTCGCCGTGGGAGA |
| BnaCnng77750D | Alpha-dioxygenase 1 |
TGTGACGCACTTTGAATGACT CACGAAAGCAAAGCAAATAGCA |
| BnaA05g30820D | REDUCED WALL ACETYLATION 2-like |
CACACTTCACTGTTCGTGTAACT ACGAAGAACCAAAGCGCGATA |
| BnaA03g11520D | Glucan endo-1,3-beta-glucosidase 12 |
CCGTGTTCGCGGCCAT GACGTTAAGCGGCTCATTCG |
| BnaC02g11750D | Cytochrome P450 81F2-like |
TATAGAATCAAACCCACCCACC CGATGGGAAACGGAGTTGGT |
| BnaC03g72480D | Short-chain dehydrogenase reductase 3a LOC106438316 |
TGTCGGGAAGCAGACTAGATG GGAAAGAGCAACGCTTAGACC |
| BnaC04g52990D | Defensin-like protein 195 (loc106394639) |
GAGGGAATACGGTGGCGAT TAGTCGCATAAGCACCTGAC |
| BnaA09g44500D | Transcription factor MYB51 |
TACCTCCTACGTTAACCGTCAC
AGAGAGATGATTGAAAGTTCCTCGT |
| BnaC04g12970D | Disease resistance protein RPS6-like |
CGTATGGCTGGACAGGTTGA GAGGCACCGTTCAGCTAGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Gong, Q.; Su, X.; Cheng, Y.; Wu, H.; Huang, Z.; Xu, A.; Dong, J.; Yu, C. Microscopic and Transcriptomic Comparison of Powdery Mildew Resistance in the Progenies of Brassica carinata × B. napus. Int. J. Mol. Sci. 2022, 23, 9961. https://doi.org/10.3390/ijms23179961
Zhang M, Gong Q, Su X, Cheng Y, Wu H, Huang Z, Xu A, Dong J, Yu C. Microscopic and Transcriptomic Comparison of Powdery Mildew Resistance in the Progenies of Brassica carinata × B. napus. International Journal of Molecular Sciences. 2022; 23(17):9961. https://doi.org/10.3390/ijms23179961
Chicago/Turabian StyleZhang, Mingzheng, Qiong Gong, Xing Su, Yaohua Cheng, Haoxue Wu, Zhen Huang, Aixia Xu, Jungang Dong, and Chengyu Yu. 2022. "Microscopic and Transcriptomic Comparison of Powdery Mildew Resistance in the Progenies of Brassica carinata × B. napus" International Journal of Molecular Sciences 23, no. 17: 9961. https://doi.org/10.3390/ijms23179961