
LjAMT2;2 Promotes Ammonium Nitrogen Transport during Arbuscular Mycorrhizal Fungi Symbiosis in Lotus japonicus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
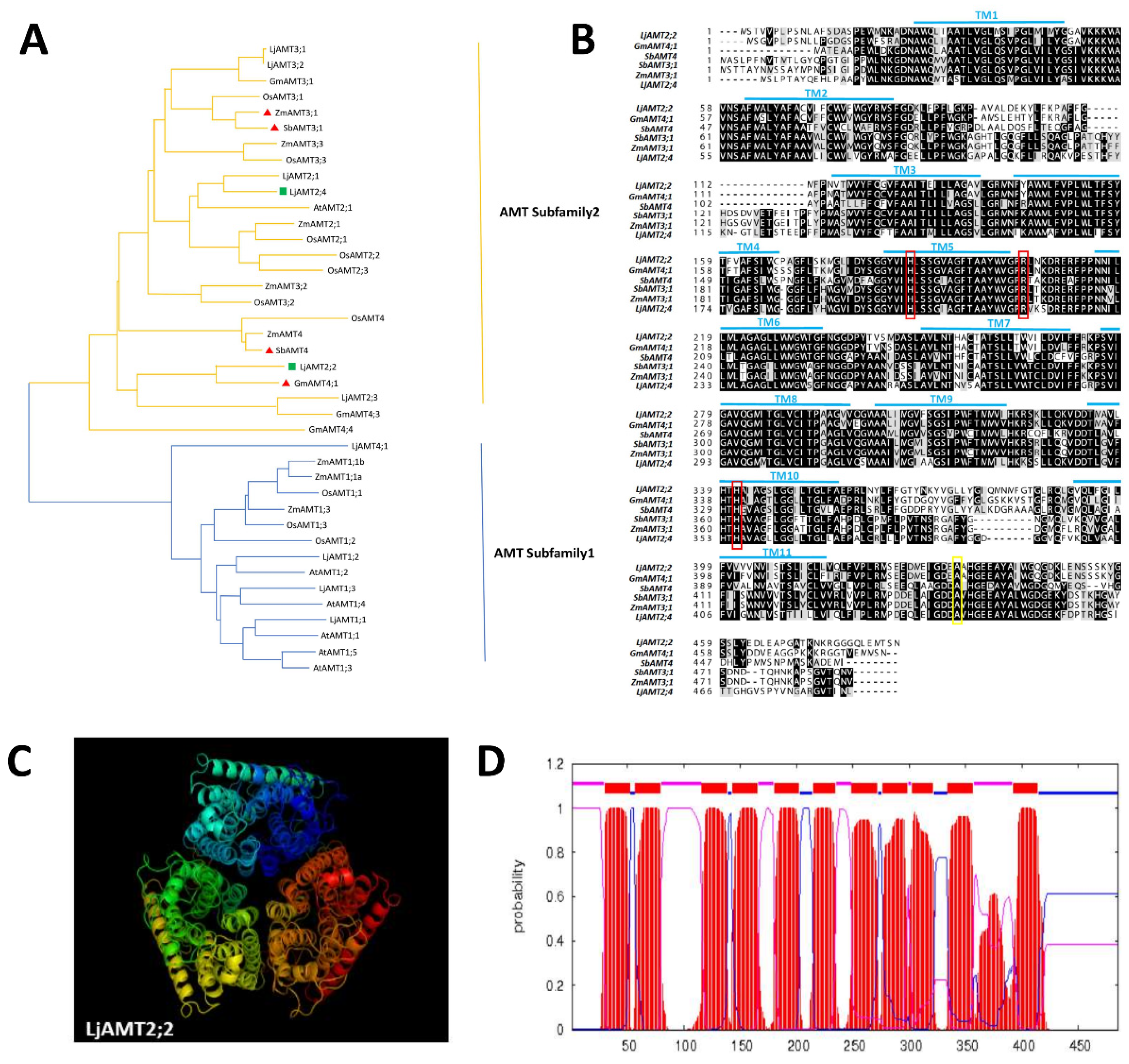
2.1. Bioinformatics Analysis of the LjAMT2;2 Gene
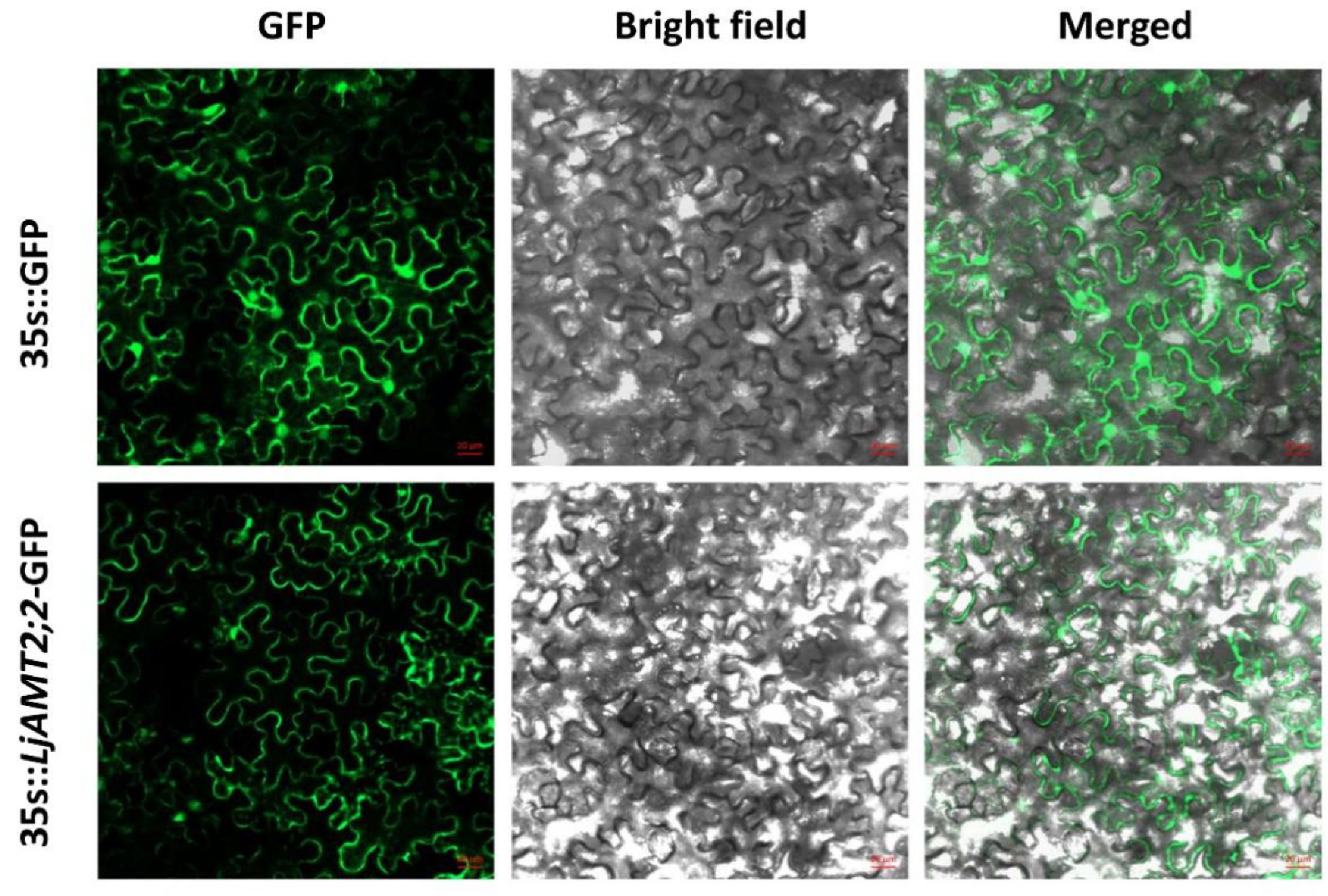
2.2. LjAMT2;2 Subcellular Localisation
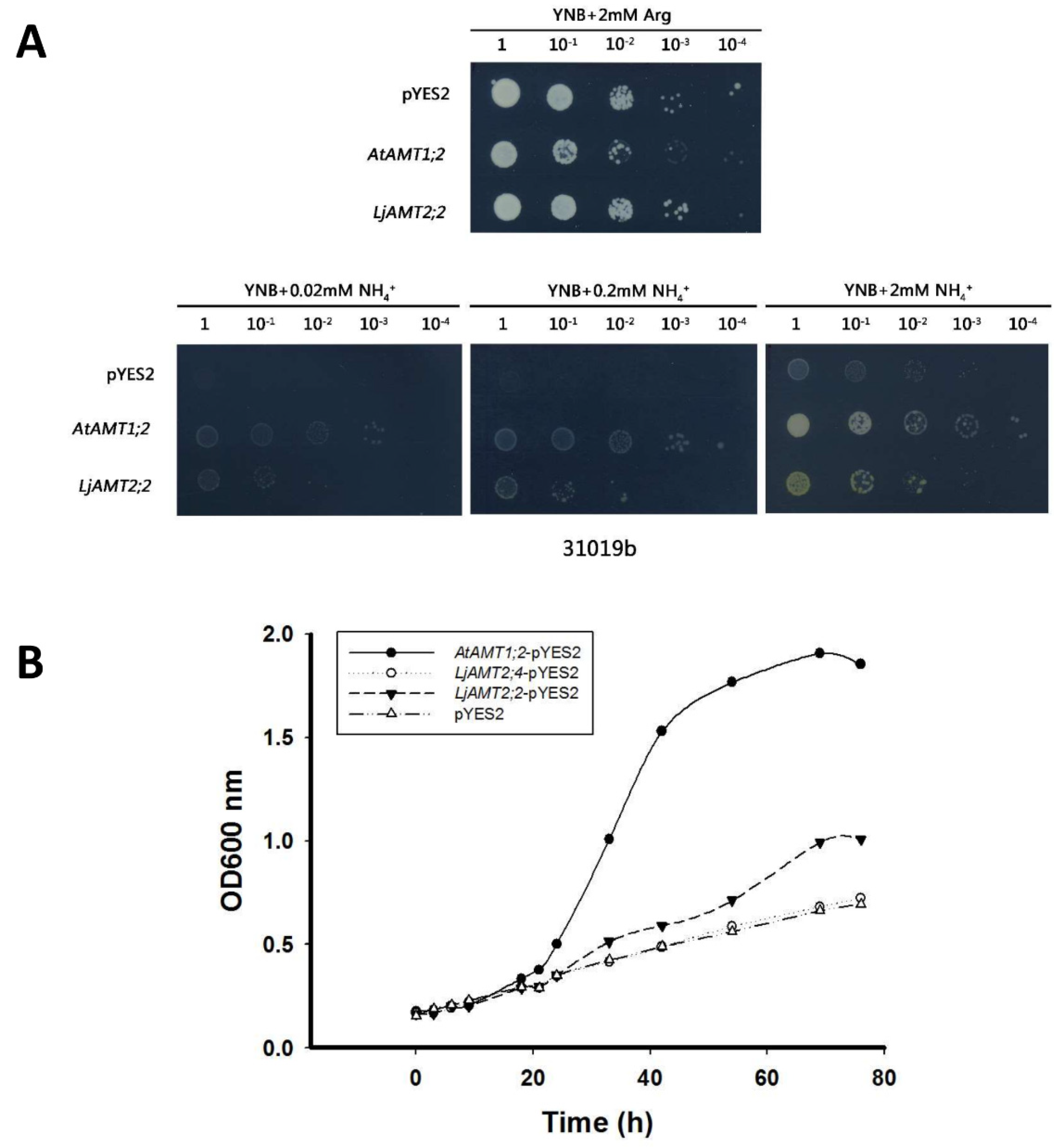
2.3. Functional Analysis of LjAMT2;2 in Ammonium-Deficient Yeast
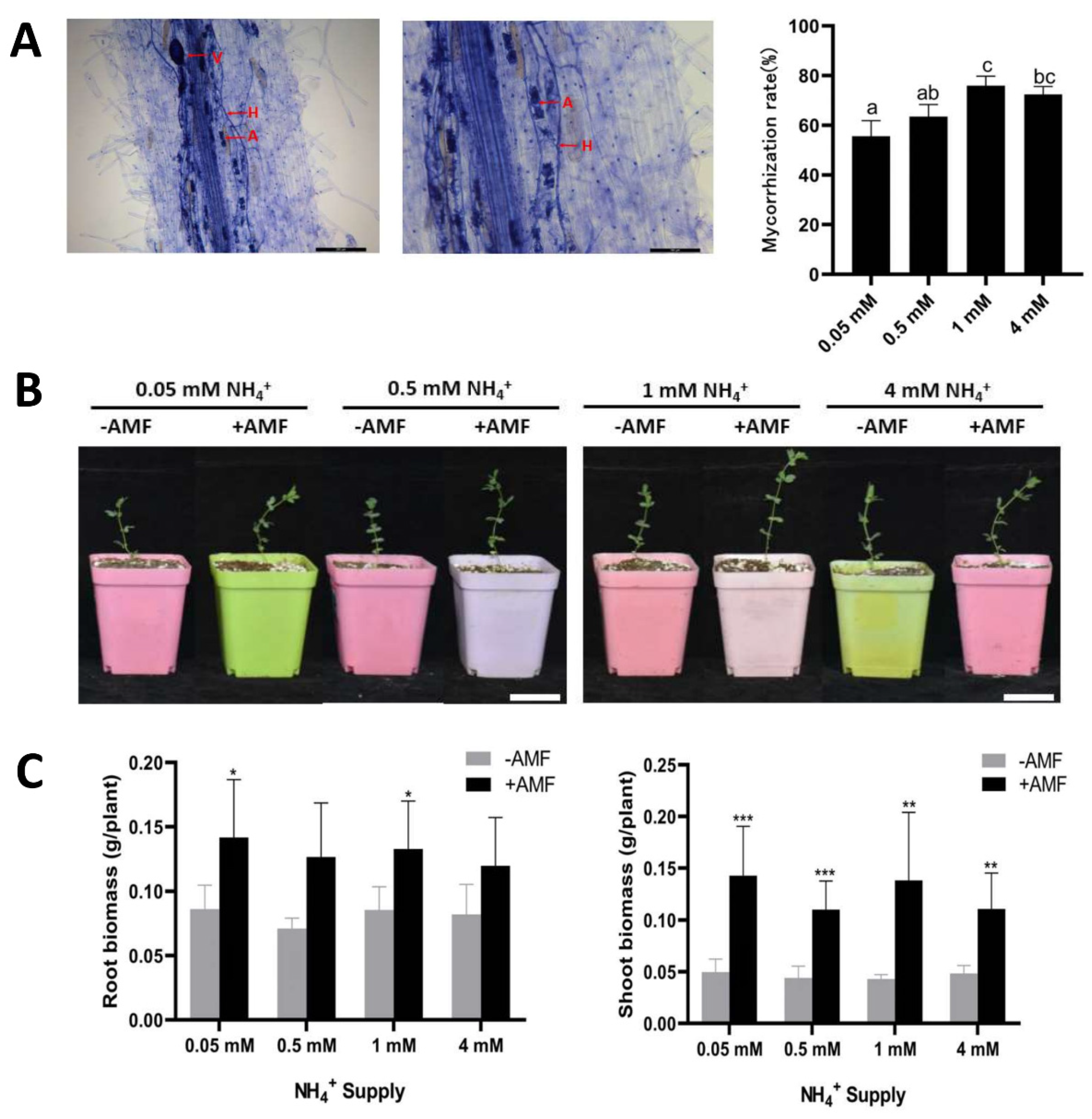
2.4. Effects of AMF Colonisation on Lotus japonicus Growth under Different Ammonium Concentrations
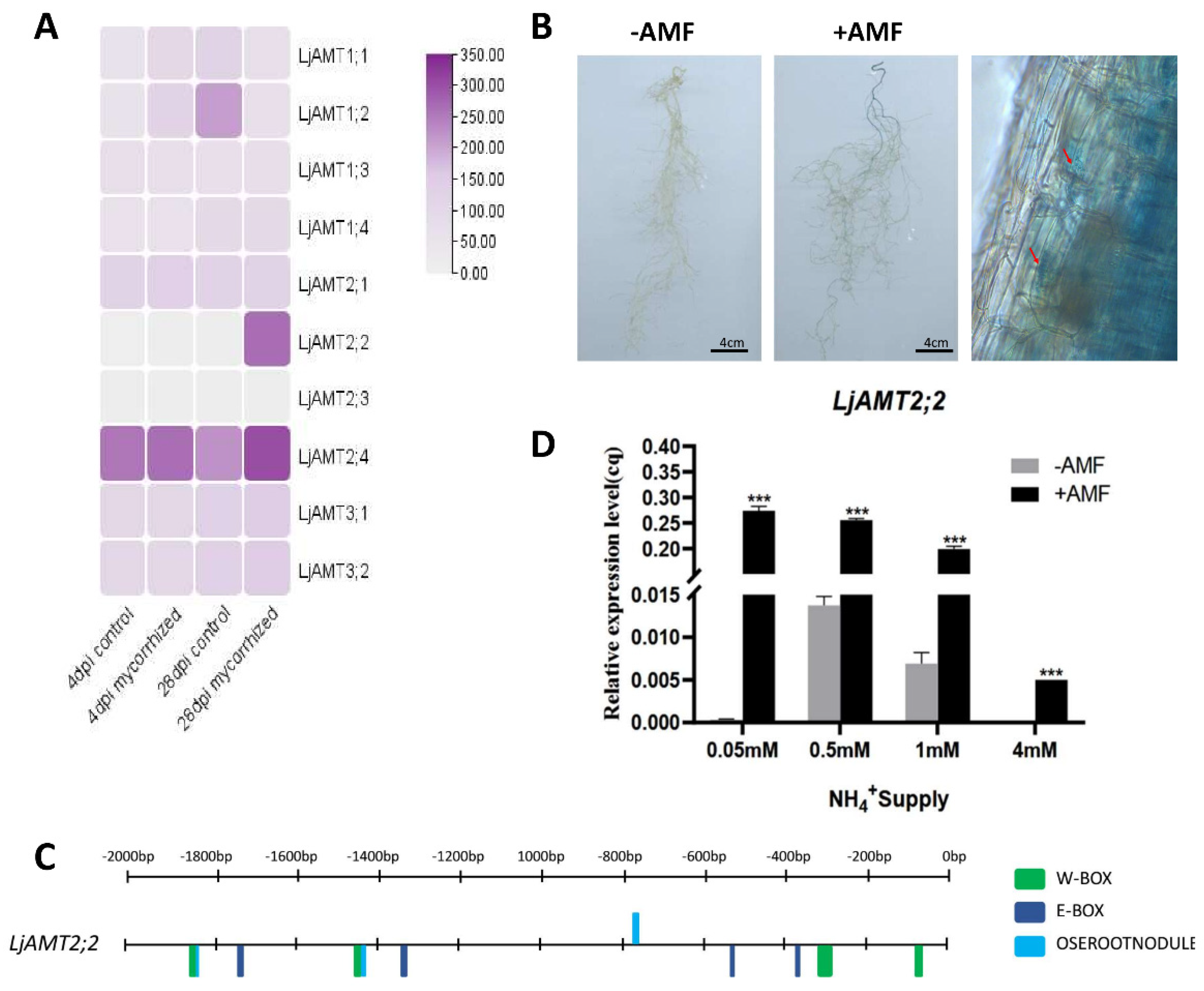
2.5. Mycorrhizal-Inducible Expression of the LjAMT2;2 Gene
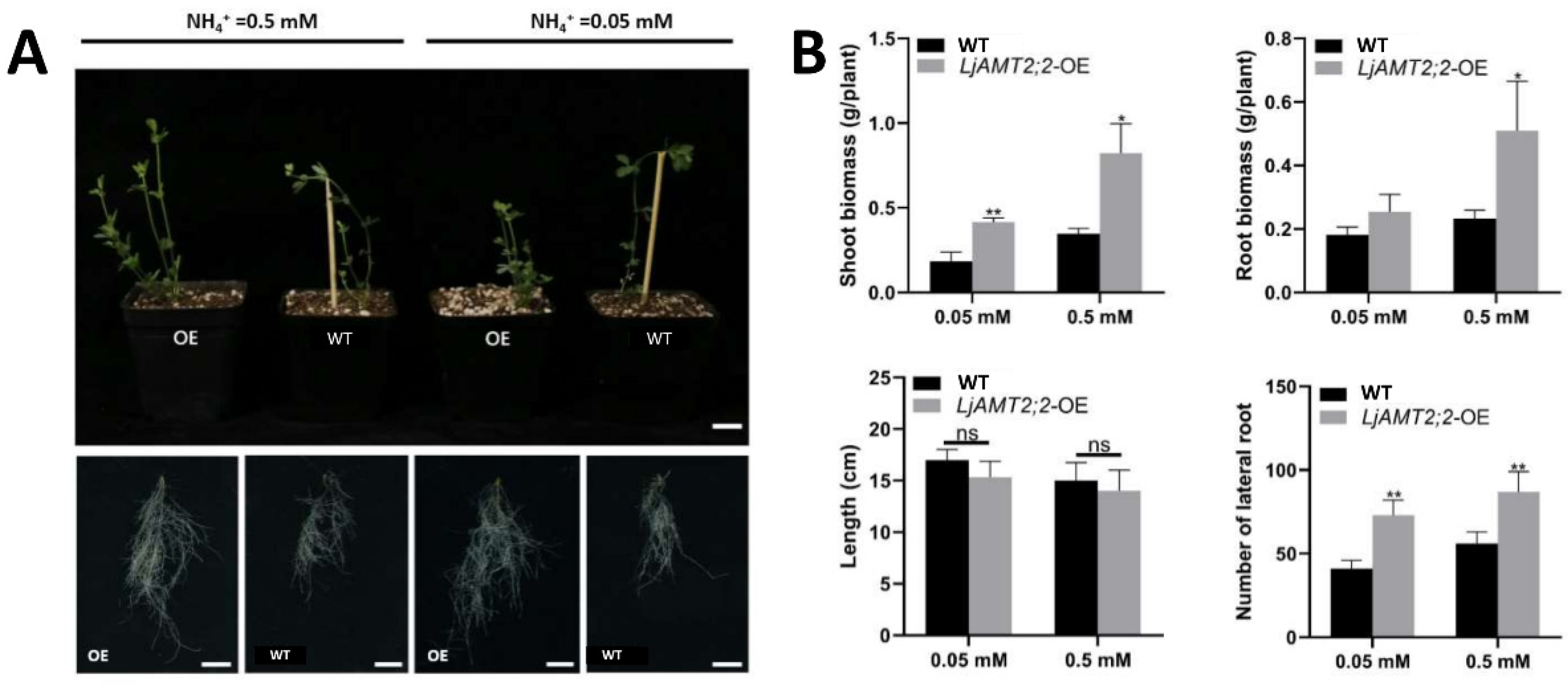
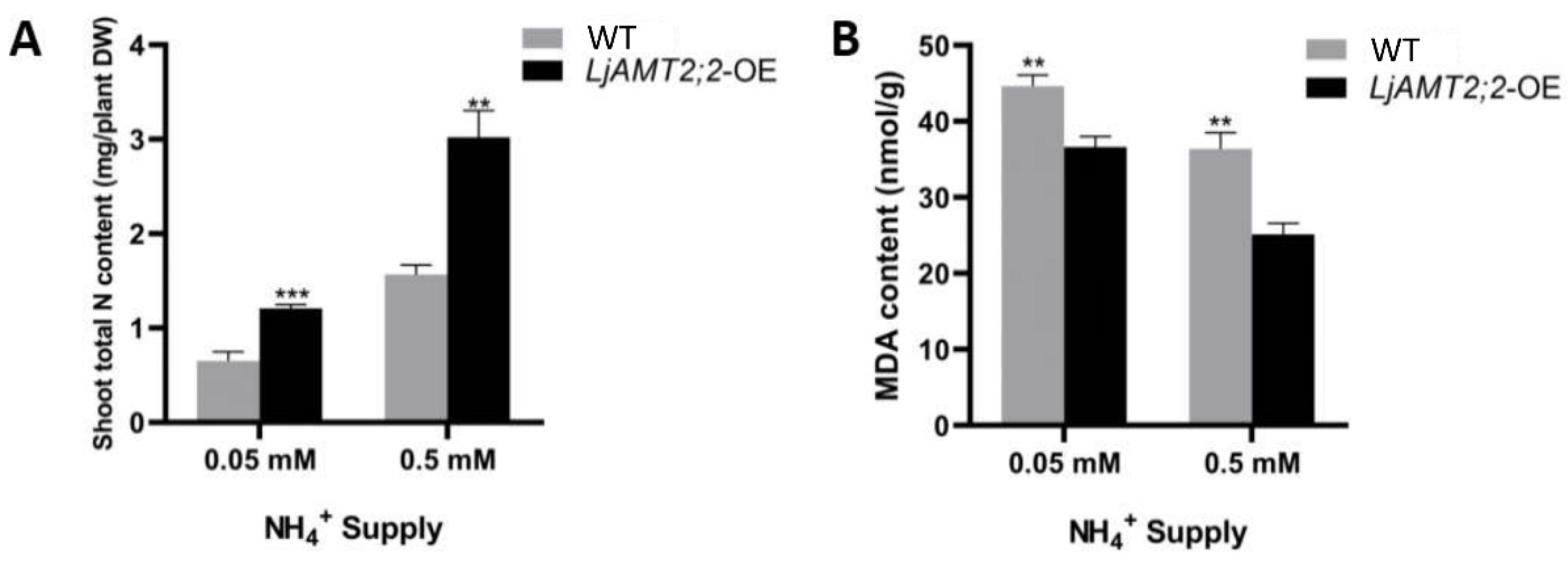
2.6. Phenotypic Identification and Physiological Index Analysis of LjAMT2;2-OE Plants
3. Discussion
4. Materials and Methods
4.1. Bioinformatics Analysis
4.2. Vector Construction
4.3. Subcellular Localisation
4.4. Experimental Materials and Planting Treatments
4.5. Transformation of Hairy Roots and Expression Induction by AMF
4.6. Supplementation of Ammonium-Deficient Yeast
4.7. RNA Isolation and Quantitative Real-Time RT-PCR (qPCR)
4.8. Determination of Nitrogen and Malondialdehyde (MDA) Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, E.S.; Read, D.J. Mycorrhizal Symbiosis. Q. Rev. Biol. 2008, 137, 273–281. [Google Scholar]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfante, P.; Genre, A. Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäder, P.; Vierheilig, H.; Streitwolf-Engel, R.; Boller, T.; Frey, B.; Christie, P.; Wiemken, A. Transport of 15N from a soil compartment separated by a polytetrafluorethylene membrane to plant roots via the hyphae of arbuscular mycorrhizal fungi. New Phytol. 2000, 146, 155–161. [Google Scholar] [CrossRef]
- Dechorgnat, J.; Nguyen, C.T.; Armengaud, P.; Jossier, M.; Diatloff, E.; Filleur, S.; Daniel-Vedele, F. From the soil to the seeds: The long journey of nitrate in plants. J. Exp. Bot. 2011, 62, 1349–1359. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Tienda, J.; Valderas, A.; Camañes, G.; Agustín, P.G.; Ferrol, N. Kinetics of NH+4 uptake by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mycorrhiza 2012, 22, 485–491. [Google Scholar] [CrossRef]
- Koegel, S.; Mieulet, D.; Baday, S.; Chatagnier, O.; Lehmann, M.F.; Wiemken, A.; Boller, T.; Wipf, D.; Bernèche, S.; Guiderdoni, E.; et al. Phylogenetic, structural, and functional characterization of AMT3;1, an ammonium transporter induced by mycorrhization among model grasses. Mycorrhiza 2017, 27, 695–708. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, B.N.; Rawat, S.R.; Siddiqi, M.Y.; Masle, J.; Glass, A.D.M. Functional analysis of an Arabidopsis T-DNA “knockout” of the high-affinity NH4(+) transporter AtAMT1;1. Plant Physiol. 2002, 130, 1263–1275. [Google Scholar] [CrossRef]
- Ludewig, U.; von Wirén, N.; Frommer, W.B. Uniport of NH4+ by the root hair plasma membrane ammonium transporter LeAMT1;1. J. Biol. Chem. 2002, 277, 13548–13555. [Google Scholar] [CrossRef] [Green Version]
- Loqué, D.; von Wirén, N. Regulatory levels for the transport of ammonium in plant roots. J. Exp. Bot. 2004, 55, 1293–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Silim, S.N.; Okamoto, M.; Siddiqi, M.Y.; Glass, A.D.M. Differential expression of three members of the AMT1 gene family encoding putative high-affinity NH4+ transporters in roots of Oryza sativa subspecies indica. Plant Cell Environ. 2003, 26, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Loqué, D.; Yuan, L.; Kojima, S.; Gojon, A.; Wirth, J.; Gazzarrini, S.; Ishiyama, K.; Takahashi, H.; Von Wirén, N. Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J. 2006, 48, 522–534. [Google Scholar] [CrossRef]
- Kakinuma, M.; Nakamoto, C.; Kishi, K.; Coury, D.A.; Amano, H. Isolation and functional characterization of an ammonium transporter gene, PyAMT1, related to nitrogen assimilation in the marine macroalga Pyropia yezoensis (Rhodophyta). Mar. Environ. Res. 2016, 128, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Z.; Duan, F.; An, X.; Liu, X.; Hao, D.; Gu, R.; Wang, Z.; Chen, F.; Yuan, L. Overexpression of the maize ZmAMT1;1a gene enhances root ammonium uptake efficiency under low ammonium nutrition. Plant Biotechnol. Rep. 2018, 12, 47–56. [Google Scholar] [CrossRef]
- Gomez, S.K.; Javot, H.; Deewatthanawong, P.; Torres-Jerez, I.; Tang, Y.; Blancaflor, E.B.; Udvardi, M.K.; Harrison, M.J. Medicago truncatula and Glomus intraradices gene expression in cortical cells harboring arbuscules in the arbuscular mycorrhizal symbiosis. BMC Plant Biol. 2009, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koegel, S.; Lahmidi, N.A.; Arnould, C.; Chatagnier, O.; Walder, F.; Ineichen, K.; Boller, T.; Wipf, D.; Wiemken, A.; Courty, P.-E. The family of ammonium transporters (AMT) in Sorghum bicolor: Two AMT members are induced locally, but not systemically in roots colonized by arbuscular mycorrhizal fungi. New Phytol. 2013, 198, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Kobae, Y.; Tamura, Y.; Takai, S.; Banba, M.; Hata, S. Localized Expression of Arbuscular Mycorrhiza-Inducible Ammonium Transporters in Soybean. Plant Cell Physiol. 2010, 51, 1411–1415. [Google Scholar] [CrossRef]
- Ruzicka, D.R.; Hausmann, N.T.; Barrios-Masias, F.H.; Jackson, L.E.; Schachtman, D.P. Transcriptomic and metabolic responses of mycorrhizal roots to nitrogen patches under field conditions. Plant Soil 2012, 350, 145–162. [Google Scholar] [CrossRef]
- Guether, M.; Neuhäuser, B.; Balestrini, R.; Dynowski, M.; Ludewig, U.; Bonfante, P. A Mycorrhizal-Specific Ammonium Transporter from Lotus japonicus Acquires Nitrogen Released by Arbuscular Mycorrhizal Fungi. Plant Physiol. 2009, 150, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytol. 2007, 174, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, F.; Rehman, S.; Dong, H.; Hu, Q.; Tu, Z.; Li, X. Genome-Wide Analysis of AMT Gene Family and its Response to Mycorrhizal Symbiosis in Maize. J. Plant Growth Regul. 2022, 1–10. [Google Scholar] [CrossRef]
- Qi, J.-L.; Zhang, W.-J.; Liu, S.-H.; Wang, H.; Sun, D.-Y.; Xu, G.-H.; Shi, M.-W.; Liu, Z.; Zhang, M.-S.; Zhang, H.-M.; et al. Expression analysis of light-regulated genes isolated from a full-length-enriched cDNA library of Onosma paniculatum cell cultures. J. Plant Physiol. 2008, 165, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Roth, R.; Paszkowski, U. Plant carbon nourishment of arbuscular mycorrhizal fungi. Curr. Opin. Plant Biol. 2017, 39, 50–56. [Google Scholar] [CrossRef]
- Rich, M.K.; Nouri, E.; Courty, P.-E.; Reinhardt, D. Diet of Arbuscular Mycorrhizal Fungi: Bread and Butter? Trends Plant Sci. 2017, 22, 652–660. [Google Scholar] [CrossRef] [Green Version]
- Bravo, A.; Brands, M.; Wewer, V.; Dörmann, P.; Harrison, M.J. Arbuscular mycorrhiza-specific enzymes FatM and RAM2 fine-tune lipid biosynthesis to promote development of arbuscular mycorrhiza. New Phytol. 2017, 214, 1631–1645. [Google Scholar] [CrossRef] [Green Version]
- Keymer, A.; Pimprikar, P.; Wewer, V.; Huber, C.; Brands, M.; Bucerius, S.L.; Delaux, P.-M.; Klingl, V.; von Röpenack-Lahaye, E.; Wang, T.L.; et al. Lipid transfer from plants to arbuscular mycorrhiza fungi. Elife 2017, 6, e29107. [Google Scholar] [CrossRef]
- Luginbuehl, L.H.; Menard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.H.; Mullins, J.G.; Merrick, M. Membrane topology of the Mep/Amt family of ammonium transporters. Mol. Microbiol. 2000, 37, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Ludewig, U.; Neuhäuser, B.; Dynowski, M. Molecular mechanisms of ammonium transport and accumulation in plants. Febs Lett. 2007, 581, 2301–2308. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Chen, F.; Yuan, L.; Zhang, F.; Mi, G. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.M.; Soussi-Boudekou, S.; Vissers, S.; Andre, B. A family of ammonium transporters in Saccharomyces cerevisiae. Mol Cell Biol. 1997, 17, 4282–4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelden, M.C.; Dong, B.; De Bruxelles, G.L.; Trevaskis, B.; Whelan, J.; Ryan, P.R.; Howitt, S.M.; Udvardi, M.K. Arabidopsis ammonium transporters, AtAMT1;1 and AtAMT1;2, have different biochemical properties and functional roles. Plant Soil 2001, 231, 151–160. [Google Scholar] [CrossRef]
- Hastings, R.P.; Remington, B. Genome-wide reprogramming of regulatory networks, transport, cell wall and membrane biogenesis during arbuscular mycorrhizal symbiosis in Lotus japonicus. Res. Dev. Disabil. 1994, 182, 200–212. [Google Scholar]
- Liao, D.; Chen, X.; Chen, A.; Wang, H.; Liu, J.; Liu, J.; Gu, M.; Sun, S.; Xu, G. The Characterization of Six Auxin-Induced Tomato GH3 Genes Uncovers a Member, SlGH3.4, Strongly Responsive to Arbuscular Mycorrhizal Symbiosis. Plant Cell Physiol. 2014, 56, 674–687. [Google Scholar] [CrossRef] [Green Version]
- Fontana, P. Nitrogen determination of Kjekdahl’s method. Mem. Inst. Oswaldo. Cruz. 1953, 51, 277–288. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhou, W.; Wu, J.; Xie, K.; Li, X. LjAMT2;2 Promotes Ammonium Nitrogen Transport during Arbuscular Mycorrhizal Fungi Symbiosis in Lotus japonicus. Int. J. Mol. Sci. 2022, 23, 9522. https://doi.org/10.3390/ijms23179522
Wang Y, Zhou W, Wu J, Xie K, Li X. LjAMT2;2 Promotes Ammonium Nitrogen Transport during Arbuscular Mycorrhizal Fungi Symbiosis in Lotus japonicus. International Journal of Molecular Sciences. 2022; 23(17):9522. https://doi.org/10.3390/ijms23179522
Chicago/Turabian StyleWang, Yanping, Wenqing Zhou, Jiandong Wu, Kailing Xie, and Xiaoyu Li. 2022. "LjAMT2;2 Promotes Ammonium Nitrogen Transport during Arbuscular Mycorrhizal Fungi Symbiosis in Lotus japonicus" International Journal of Molecular Sciences 23, no. 17: 9522. https://doi.org/10.3390/ijms23179522