
Long-Chain Acyl-CoA Synthetases Promote Poplar Resistance to Abiotic Stress by Regulating Long-Chain Fatty Acid Biosynthesis


 ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Classification of PtLACS Family Genes
2.2. Multiple Alignments and Phylogenetic Analysis of PtLACSs
2.3. Conserved Motif and Gene Structure Analysis of PtLACSs
2.4. PtLACS 3D Structure and Phosphorylation Site Analysis
2.5. Chromosome Localization and Collinearity of PtLACS Genes Analysis
2.6. Cis-Acting Element Analysis of PtLACS Genes
2.7. Interaction Network and GO Enrichment Analysis of PtLACSs
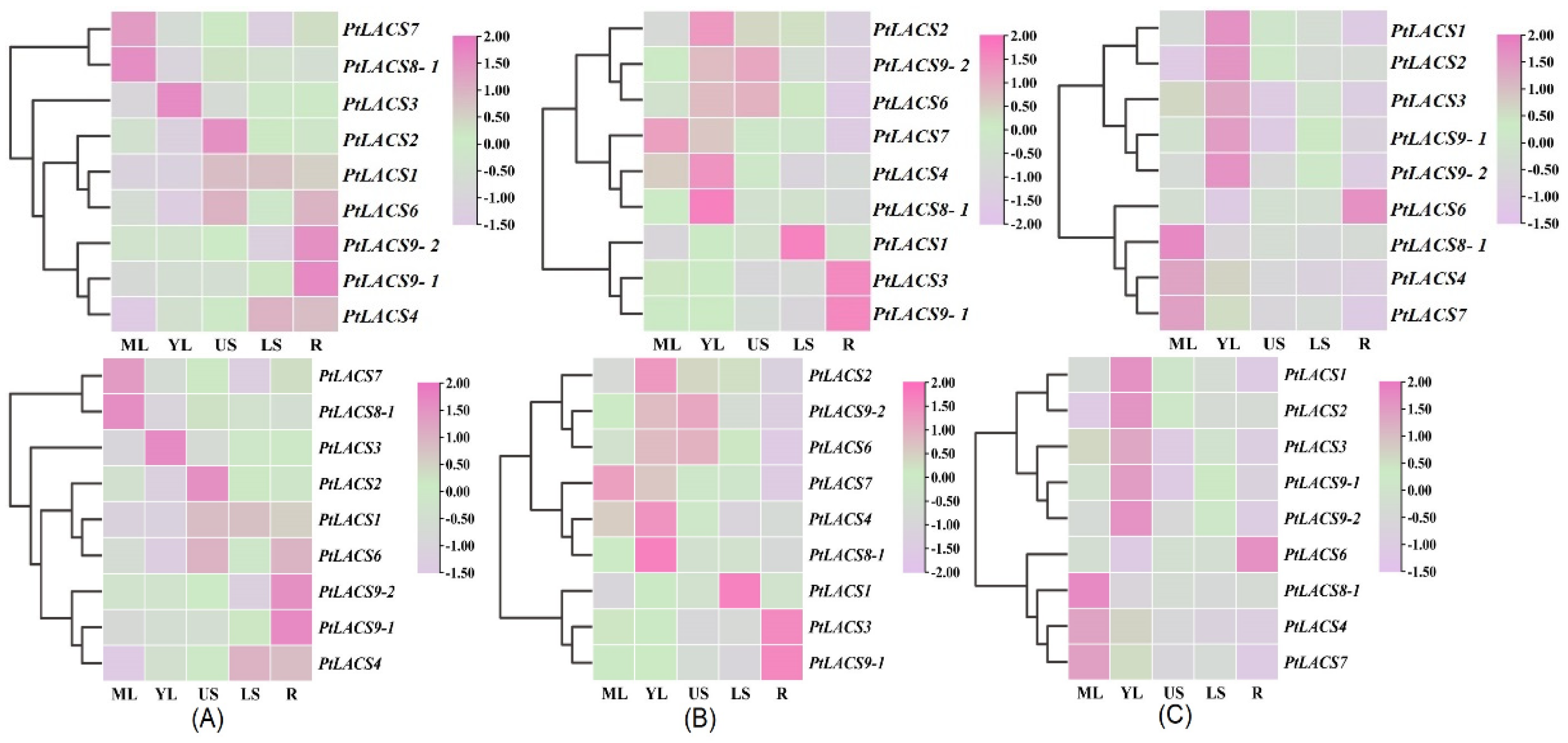
2.8. Expression Patterns of PtLACSs in Various Tissues
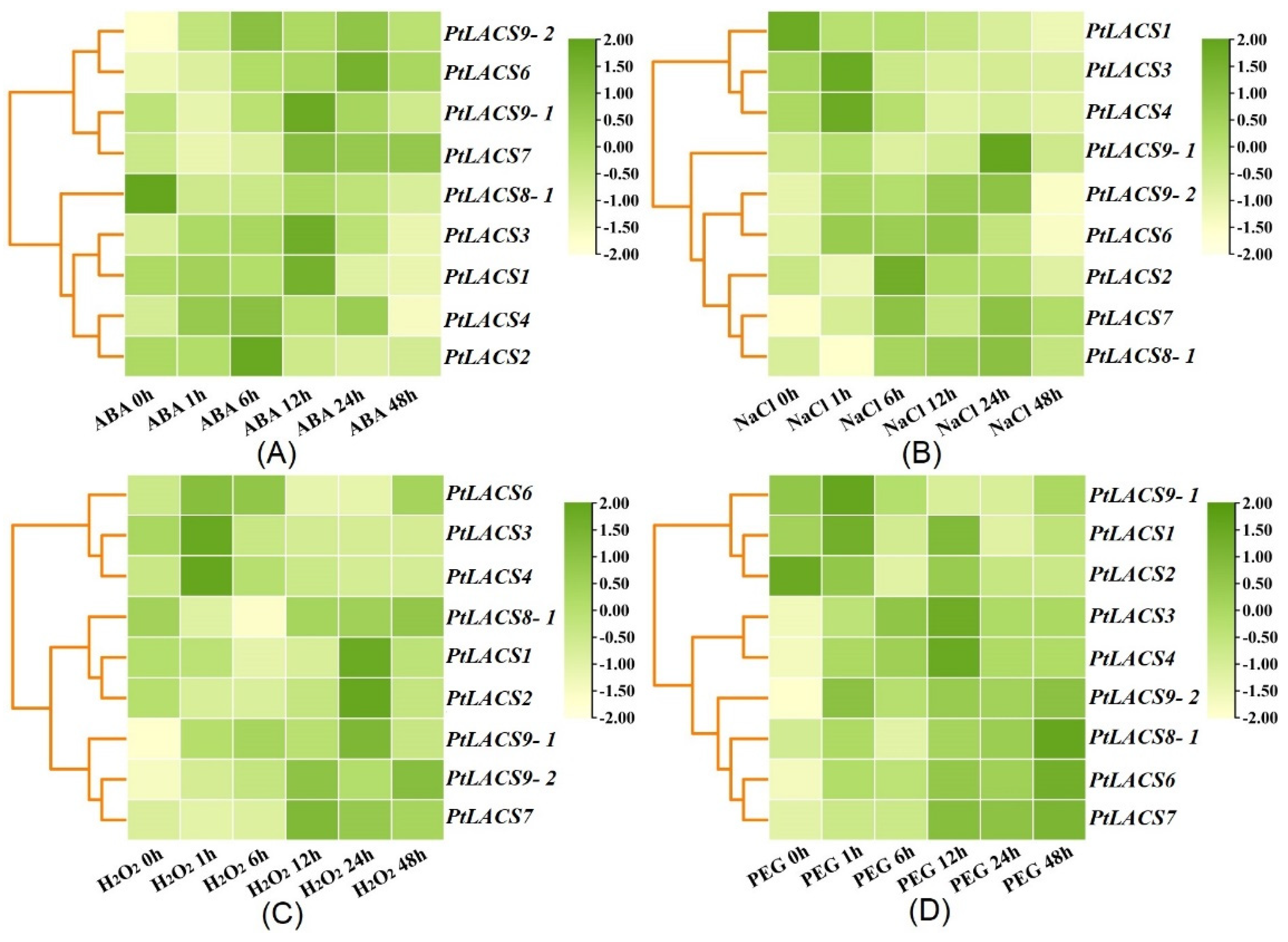
2.9. Expression Patterns of PtLACSs in Response to Different Stress Conditions
2.10. Cluster Network Analysis among PtLACSs and PtLTPGs
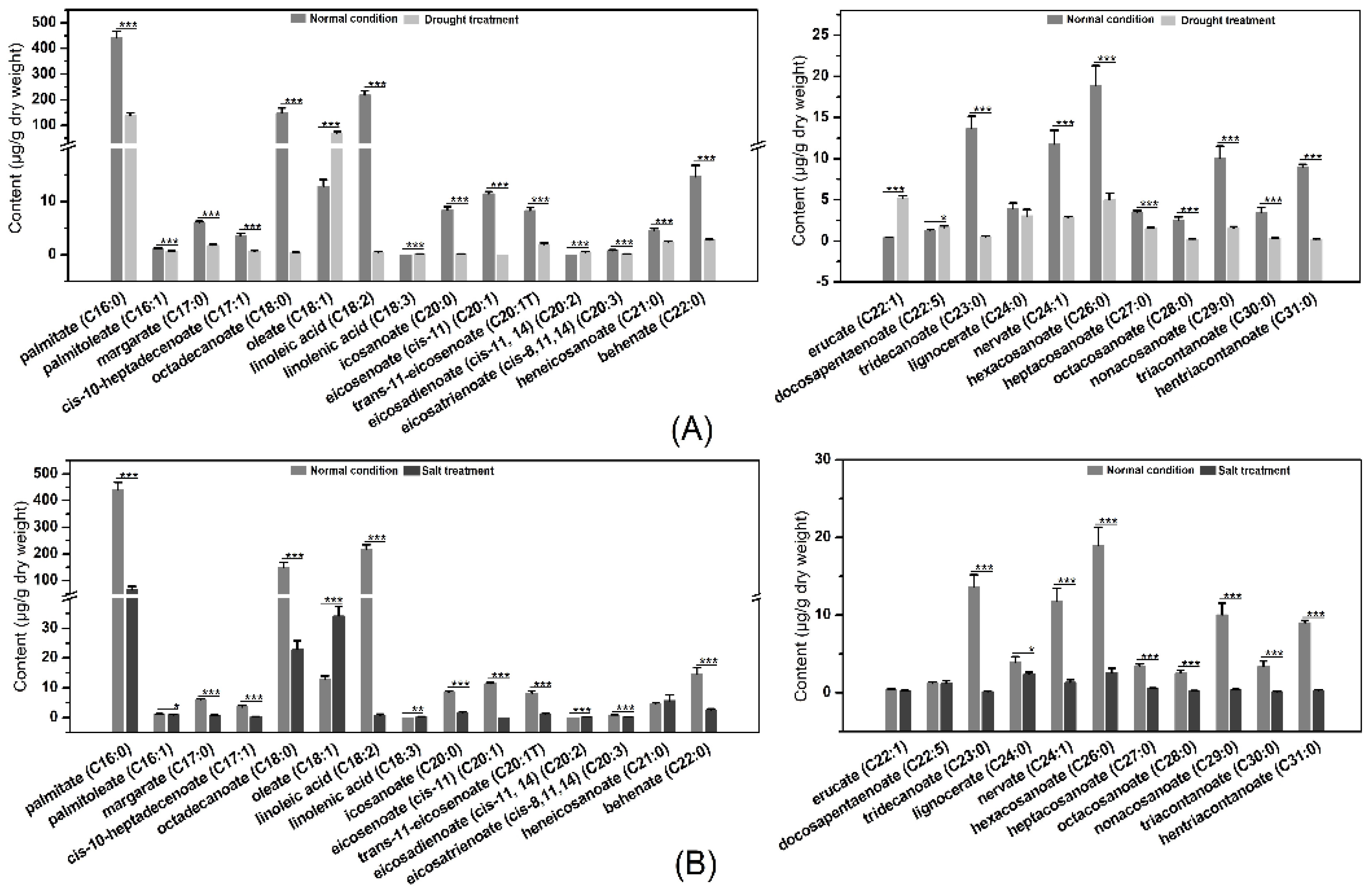
2.11. LCFAs and VLCFAs Analysis before and after Osmotic Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Abiotic Stress Treatments
4.2. Identification and Classification of LACS Proteins in Poplar
4.3. Characteristics and Phylogenetic Tree of LACS Members
4.4. Gene Structure, Conserved Motif, and 3D Structure Analysis of PtLACS Members
4.5. Chromosomal Location and Collinearity Analysis of PtLACS Members
4.6. Cis-Regulatory Element, Interaction Network, and GO Analysis of PtLACS Members
4.7. RNA Isolation and Expression Patterns of PtLACS Genes
4.8. Gene Cluster Network Analysis of PtLACS and PtLTPG Genes
4.9. LCFAs and VLCFAs Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ge, W.; Kong, X.; Zhao, Y.; Wei, B.; Zhou, Q.; Ji, S. Insights into the metabolism of membrane lipid fatty acids associated with chilling injury in post-harvest bell peppers. Food Chem. 2019, 295, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Marsafari, M.; Wang, F.; Deng, L.; Xu, P. Engineering acetyl-CoA metabolic shortcut for eco-friendly production of polyketides triacetic acid lactone in Yarrowia lipolytica. Metab. Eng. 2019, 56, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Kosma, D.; Lu, S. Functional role of long chain acyl-CoA synthetases (LACS) in plant development and stress responses. Front. Plant Sci. 2021, 12, 276. [Google Scholar]
- Zhang, C.L.; Mao, K.; Zhou, L.J.; Wang, G.L.; Zhang, Y.L.; Li, Y.Y.; Hao, Y.J. Genome-wide identification and characterization of apple long-chain acyl-CoA synthetases and expression analysis under different stresses. Plant Physiol. Biochem. 2018, 132, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Haslam, T.M.; Sonntag, A.; Molina, I.; Kunst, L. Functional overlap of long-chain acyl-CoA synthetases in Arabidopsis. Plant Cell Physiol. 2019, 60, 1041–1054. [Google Scholar] [CrossRef]
- Kim, B.H.; Akoh, C.C. Chemical and physical properties of butterfat-vegetable oil blend spread prepared with enzymatically transesterified canola oil and caprylic acid. J. Agric. Food Chem. 2005, 53, 4954–4961. [Google Scholar] [CrossRef]
- Wang, L.; Ding, X.; Gao, Y.; Yang, S. Genome-wide identification and characterization of GRAS genes in soybean (Glycine max). BMC Plant Biol. 2020, 20, 415. [Google Scholar] [CrossRef]
- Smirnova, A.; Leide, J.; Riederer, M. Deficiency in a very-long-chain fatty acid β-ketoacyl-coenzyme a synthase of tomato. impairs microgametogenesis and causes floral organ fusion. Plant Physiol. 2013, 161, 196–209. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Xia, Q.; Xie, W.; Dumonceaux, T.; Zou, J.; Datla, R.; Selvaraj, G. Male gametophyte development in bread wheat (Triticum aestivum L.): Molecular, cellular, and biochemical analyses of a sporophytic contribution to pollen wall ontogeny. Plant J. 2002, 30, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Roudier, F.; Gissot, L.; Beaudoin, F.; Haslam, R.; Michaelson, L.; Marion, J.; Molino, D.; Lima, A.; Bach, L.; Morin, H.; et al. Very-long-chain fatty acids are involved in polar auxin transport and developmental patterning in Arabidopsis. Plant Cell 2010, 22, 364–375. [Google Scholar] [CrossRef] [Green Version]
- Bach, L.; Gissot, L.; Marion, J.; Tellier, F.; Moreau, P.; Satiat-Jeunemaître, B.; Palauqui, J.-C.; Napier, J.A.; Faure, J.-D. Very-long-chain fatty acids are required for cell plate formation during cytokinesis in Arabidopsis thaliana. J. Cell Sci. 2011, 124, 3223–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.; Tian, L.; Chen, M.; Chen, Y.; Wei, A. Low temperature affects fatty acids profiling and key synthesis genes expression patterns in Zanthoxylum bungeanum Maxim. Int. J. Mol. Sci. 2022, 23, 2319. [Google Scholar] [CrossRef] [PubMed]
- Ozolina, N.V.; Gurina, V.V.; Nesterkina, I.S.; Nurminsky, V.N. Variations in the content of tonoplast lipids under abiotic stress. Planta 2020, 251, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pulsifer, I.P.; Kluge, S.; Rowland, O. Arabidopsis long-chain acyl-CoA synthetase 1 (LACS1), LACS2, and LACS3 facilitate fatty acid uptake in yeast. Plant Physiol. Biochem. 2012, 51, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Yeats, T.H.; Rose, J.K. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Jessen, D.; Olbrich, A.; Knüfer, J.; Krüger, A.; Hoppert, M.; Polle, A.; Fulda, M. Combined activity of LACS1 and LACS4 is required for proper pollen coat formation in Arabidopsis. Plant J. 2011, 68, 715–726. [Google Scholar] [CrossRef]
- Xu, C.; Fan, J.; Shanklin, J. Metabolic and functional connections between cytoplasmic and chloroplast triacylglycerol storage. Prog. Lipid Res. 2020, 80, 101069. [Google Scholar] [CrossRef]
- Shockey, J.; Browse, J. Genome-level and biochemical diversity of the acyl-activating enzyme superfamily in plants. Plant J. 2011, 66, 143–160. [Google Scholar] [CrossRef]
- Jessen, D.; Roth, C.; Wiermer, M.; Fulda, M. Two activities of long-chain acyl-coenzyme A synthetase are involved in lipid trafficking between the endoplasmic reticulum and the plastid in Arabidopsi. Plant Physiol. 2015, 167, 351–366. [Google Scholar] [CrossRef] [Green Version]
- Digel, M.; Ehehalt, R.; Stremmel, W.; Füllekrug, J. Acyl-CoA synthetases: Fatty acid uptake and metabolic channeling. Mol. Cell. Biochem. 2009, 326, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.M.; Fulda, M.S.; Browse, J.A. Arabidopsis contains nine long-chain acyl-coenzyme a synthetase genes that participate in fatty acid and glycerolipid metabolism. Plant Physiol. 2002, 129, 1710–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Katavic, V.; Li, F.; Haughn, G.W.; Kunst, L. Insertional mutant analysis reveals that long-chain acyl-CoA synthetase 1 (LACS1), but not LACS8, functionally overlaps with LACS9 in Arabidopsis seed oil biosynthesis. Plant J. 2010, 64, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, A.; Saqib, S.; Huang, H.; Zaman, W.; Lü, S.; Zhao, H. Genome-wide comparative analysis of long-chain acyl-CoA synthetases (LACSs) gene family: A focus on identification, evolution and expression profiling related to lipid synthesis. Plant Physiol. Biochem. 2021, 161, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fulda, M.; Shockey, J.; Werber, M.; Wolter, F.P.; Heinz, E. Two long-chain acyl-CoA synthetases from Arabidopsis thaliana involved in peroxisomal fatty acid β-oxidation. Plant J. 2002, 32, 93–103. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, J.; Chen, H.; Zhang, Q.; Zheng, Y.; Li, D. Molecular cloning and characterization of long-chain acyl-CoA synthetase 9 from the mesocarp of African oil palm (Elaeis guineensis Jacq.). Sci. Hortic. 2021, 276, 109751. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, G.; Mietkiewska, E.; Song, Z.; Caldo, K.M.P.; Singer, S.D.; Dyer, J.; Smith, M.; McKeon, T.; Weselake, R.J. Castor patatin-like phospholipase A IIIβ facilitates removal of hydroxy fatty acids from phosphatidylcholine in transgenic Arabidopsis seeds. Plant Mol. Biol. 2019, 101, 521–536. [Google Scholar] [CrossRef]
- Tonon, T.; Qing, R.; Harvey, D.; Li, Y.; Larson, T.R.; Graham, I.A. Identification of a long-chain polyunsaturated fatty acid acyl-coenzyme A synthetase from the diatom Thalassiosira pseudonana. Plant Physiol. 2005, 138, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.-L.; Zhang, Y.-L.; Hu, X.; Xiao, X.; Wang, G.-L.; You, C.-X.; Li, Y.-Y.; Hao, Y.-J. An apple long-chain acyl-CoA synthetase, MdLACS4, induces early flowering and enhances abiotic stress resistance in Arabidopsis. Plant Sci. 2020, 297, 110529. [Google Scholar] [CrossRef]
- Wang, X.L.; Li, X.B. The GhACS1 gene encodes an acyl-CoA synthetase which is essential for normal microsporogenesis in early anther development of cotton. Plant J. 2009, 57, 473–486. [Google Scholar] [CrossRef]
- Yu, L.; Tan, X.; Jiang, B.; Sun, X.; Gu, S.; Han, T.; Hou, W. A peroxisomal long-chain acyl-CoA synthetase from Glycine max involved in lipid degradation. PLoS ONE 2014, 9, e100144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aznar-Moreno, J.A.; Caleron, M.V.; Martínez-Force, E.; Garcés, R.; Mullen, R.; Gidda, S.K.; Salas, J.J. Sunflower (Helianthus annuus) long-chain acyl-coenzyme A synthetases expressed at high levels in developing seeds. Physiol. Plant. 2014, 150, 363–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.N.; Gu, S.L.; Zhu, F.G.; Ma, Z.Y.; Li, J.; Li, M.; Wang, Z.; Tan, X.L. Long-chain acyl-CoA synthetase 2 is involved in seed oil production in Brassica napus. BMC Plant Biol. 2020, 20, 21. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Holic, R.; Li, D.; Pan, X.; Mietkiewska, E.; Chen, G.; Ozga, J.; Weselake, R.J. Substrate preferences of long-chain acyl-CoA synthetase and diacylglycerol acyltransferase contribute to enrichment of flax seed oil with α-linolenic acid. Biochem J. 2018, 475, 1473–1489. [Google Scholar] [CrossRef] [PubMed]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Cai, X.; Zhou, Z.; Wang, X.; Diouf, L.; Xu, Y.; Hou, Y.; Hu, Y.; et al. Whole genome analysis of cyclin dependent kinase (CDK) gene family in cotton and functional evaluation of the role of CDKF4 gene in drought and salt stress tolerance in plants. Int. J. Mol. Sci. 2018, 19, 2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Yang, H.; Wang, X.; Qiu, Y.; Tian, L.; Qi, X.; Qu, L.Q. Cytochrome P450 family member CYP96B5 hydroxylates alkanes to primary alcohols and is involved in rice leaf cuticular wax synthesis. New Phytol. 2020, 225, 2094–2107. [Google Scholar] [CrossRef] [PubMed]
- Eastmond, P.J. SUGAR-DEPENDENT1 encodes a patatin domain triacylglycerol lipase that initiates storage oil breakdown in germinating Arabidopsis seeds. Plant Cell 2006, 18, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Pinfield-Wells, H.; Rylott, E.L.; Gilday, A.D.; Graham, S.; Job, K.; Larson, T.R.; Graham, I.A. Sucrose rescues seedling establishment but not germination of Arabidopsis mutants disrupted in peroxisomal fatty acid catabolism. Plant J. 2005, 43, 861–872. [Google Scholar] [CrossRef]
- Kataya, A.; Schei, E.; Lillo, C. Towards understanding peroxisomal phosphoregulation in Arabidopsis thaliana. Planta 2016, 243, 699–717. [Google Scholar] [CrossRef]
- Carrie, C.; Murcha, M.W.; Millar, A.H.; Smith, S.M.; Whelan, J. Nine 3-ketoacyl-CoA thiolases (KATs) and acetoacetyl-CoA thiolases (ACATs) encoded by five genes in Arabidopsis thaliana are targeted either to peroxisomes or cytosol but not to mitochondria. Plant Mol. Biol. 2007, 63, 97–108. [Google Scholar] [CrossRef]
- Cameron, K.D.; Teece, M.A.; Smart, L.B. Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiol. 2006, 140, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Pearl, S.A.; Jackson, S.A. Gene networks in plant biology: Approaches in reconstruction and analysis. Trends Plant Sci. 2015, 20, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Fich, E.A.; Segerson, N.A.; Rose, J.K. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, K.I.; Kobayashi, N.; Saito, K. Lipid synthesis and acyl-CoA synthetase in developing rice seeds. Lipids 2003, 38, 881–884. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, S.; Xu, Y.; Li, S.; Zhang, S.; Yuan, Z.; Li, J.; Ni, Y. Overexpression of BnKCS1-1, BnKCS1-2, and BnCER1-2 promotes cuticular wax production and increases drought tolerance in Brassica napus. Crop J. 2020, 8, 26–37. [Google Scholar] [CrossRef]
- Carianopol, C.S.; Chan, A.L.; Dong, S.; Provart, N.J.; Lumba, S.; Gazzarrini, S. An abscisic acid-responsive protein interaction network for sucrose non-fermenting related kinase1 in abiotic stress response. Commun. Biol. 2020, 3, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, A.; Joubès, J. Arabidopsis cuticular waxes: Advances in synthesis, export and regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef]
- Bessire, M.; Chassot, C.; Jacquat, A.-C.; Humphry, M.; Borel, S.; Petétot, J.M.-C.; Métraux, J.-P.; Nawrath, C. A permeable cuticle in Arabidopsis leads to a strong resistance to Botrytis cinerea. EMBO J. 2007, 26, 2158–2168. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Li, N.; Wang, S.; Sun, J.; Zhang, L.; Zhang, C.; Yang, H.; Zhao, H.; Yang, B.; Wei, J.; et al. Genome-wide identification and comparative expression profile analysis of the long-chain Acyl-CoA synthetase (LACS) gene family in two different oil content cultivars of Brassica napus. Biochem. Genet. 2019, 57, 781–800. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.L.; Zheng, X.F.; Zhang, Z.Y.; Zheng, W.A.N.G.; Xia, H.C.; Changming, L.U.; Gu, S.L. Long chain acyl-coenzyme A synthetase 4 (BnLACS4) gene from Brassica napus enhances the yeast lipid contents. J. Integr. Agric. 2014, 13, 54–62. [Google Scholar] [CrossRef]
- Meng, D.; He, M.; Bai, Y.; Xu, H.; Dandekar, A.M.; Fei, Z.; Cheng, L. Decreased sorbitol synthesis leads to abnormal stamen development and reduced pollen tube growth via an MYB transcription factor, MdMYB39L, in apple (Malus domestica). New Phytol. 2018, 217, 641–656. [Google Scholar] [CrossRef] [Green Version]
- Edstam, M.M.; Blomqvist, K.; Eklöf, K.; Wennergren, U.; Edqvist, J. Coexpression patterns indicate that GPI-anchored non-specific lipid transfer proteins are involved in accumulation of cuticular wax, suberin and sporopollenin. Plant Mol. Biol. 2013, 83, 625–649. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Suh, M.C. Disruption of glycosylphosphatidylinositol-anchored lipid transfer protein 15 affects seed coat permeability in Arabidopsis. Plant J. 2018, 96, 1206–1217. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Movahedi, A.; Liu, G.; Zhu, S.; Chen, Y.; Yu, C.; Zhong, F.; Zhang, J. Poplar glycosylphosphatidylinositol-anchored lipid transfer proteins respond to osmotic stress by regulating fatty acid biosynthesis. Ind. Crops Prod. 2022, 179, 114683. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Movahedi, A.; Zhang, Y.; Aghaei-Dargiri, S.; Liu, G.; Zhu, S.; Yu, C.; Chen, Y.; Zhong, F.; Zhang, J. Long-Chain Acyl-CoA Synthetases Promote Poplar Resistance to Abiotic Stress by Regulating Long-Chain Fatty Acid Biosynthesis. Int. J. Mol. Sci. 2022, 23, 8401. https://doi.org/10.3390/ijms23158401
Wei H, Movahedi A, Zhang Y, Aghaei-Dargiri S, Liu G, Zhu S, Yu C, Chen Y, Zhong F, Zhang J. Long-Chain Acyl-CoA Synthetases Promote Poplar Resistance to Abiotic Stress by Regulating Long-Chain Fatty Acid Biosynthesis. International Journal of Molecular Sciences. 2022; 23(15):8401. https://doi.org/10.3390/ijms23158401
Chicago/Turabian StyleWei, Hui, Ali Movahedi, Yanyan Zhang, Soheila Aghaei-Dargiri, Guoyuan Liu, Sheng Zhu, Chunmei Yu, Yanhong Chen, Fei Zhong, and Jian Zhang. 2022. "Long-Chain Acyl-CoA Synthetases Promote Poplar Resistance to Abiotic Stress by Regulating Long-Chain Fatty Acid Biosynthesis" International Journal of Molecular Sciences 23, no. 15: 8401. https://doi.org/10.3390/ijms23158401