
Maternal Iron Deficiency Programs Rat Offspring Hypertension in Relation to Renin—Angiotensin System and Oxidative Stress


 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effect of Low-Iron Diet on Mother Rats
2.2. Body Weight, Serum Iron, and Hemoglobin Concentration of Adult Male Offspring
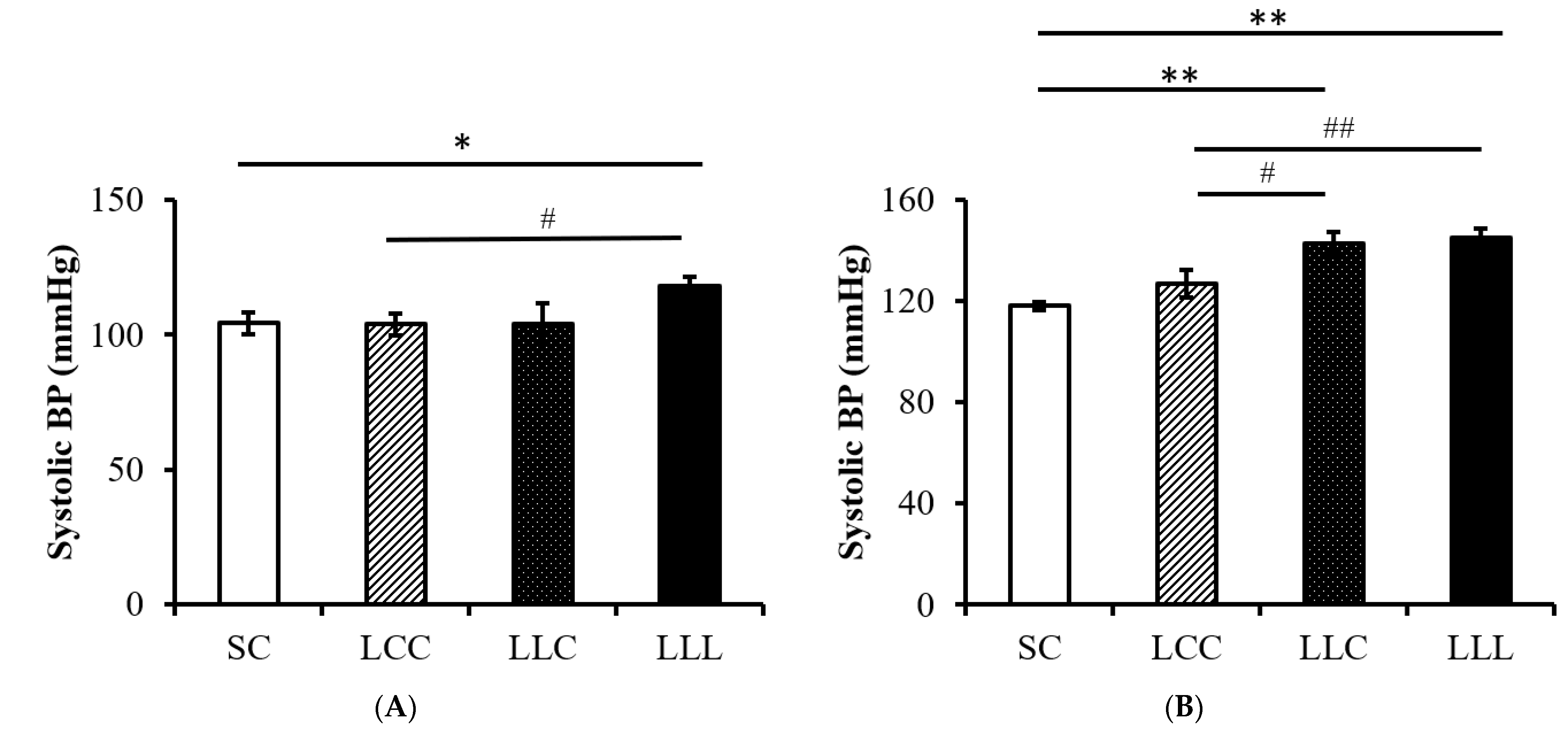
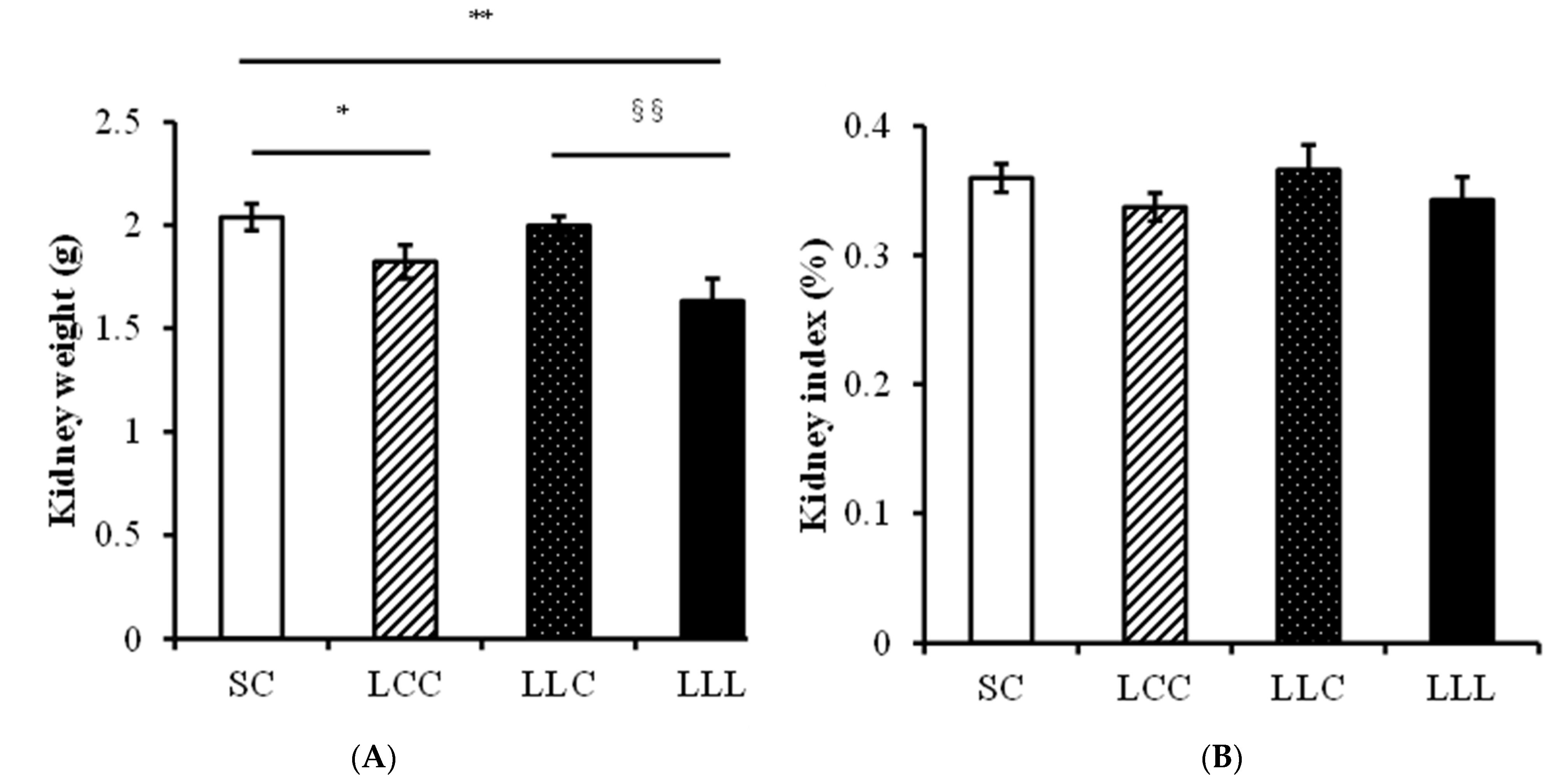
2.3. Blood Pressure and Kidney Index
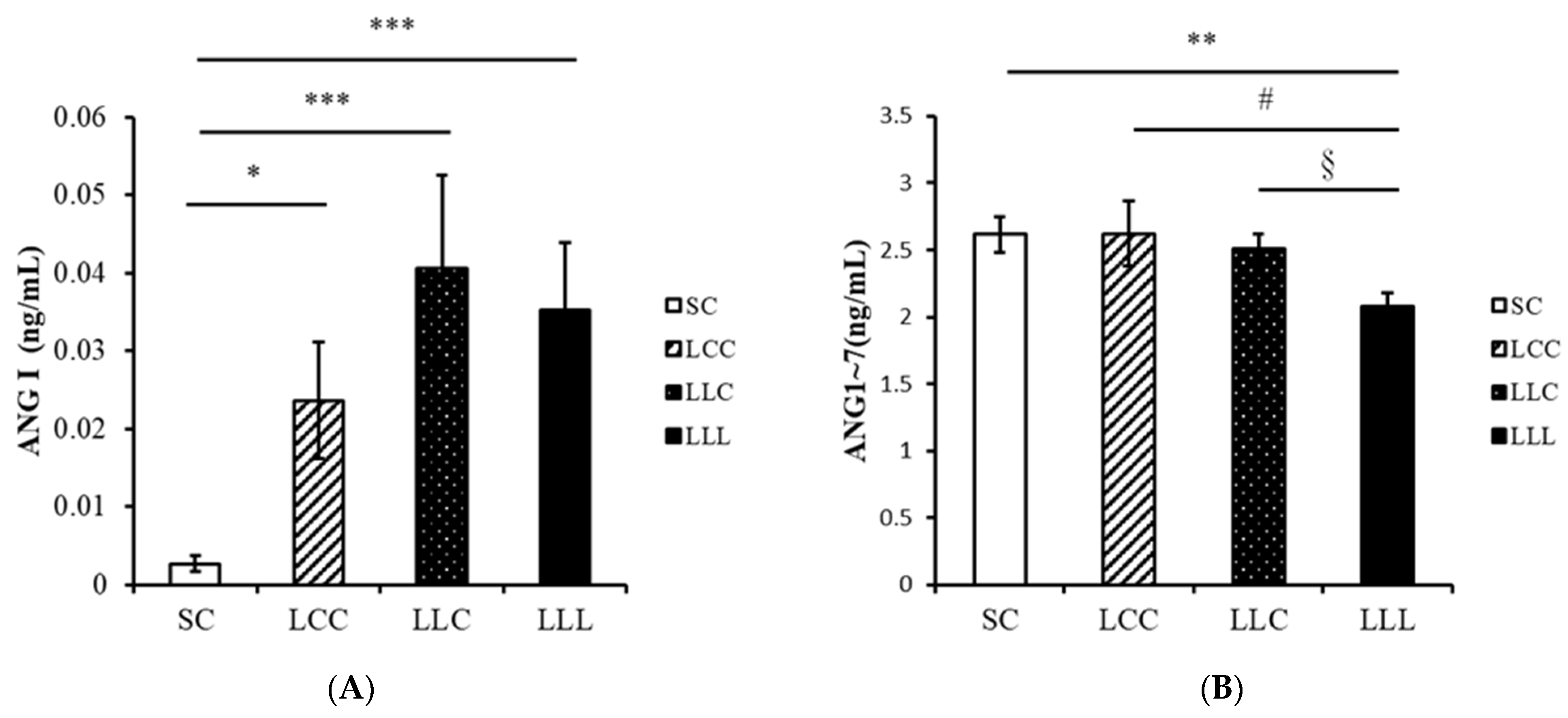
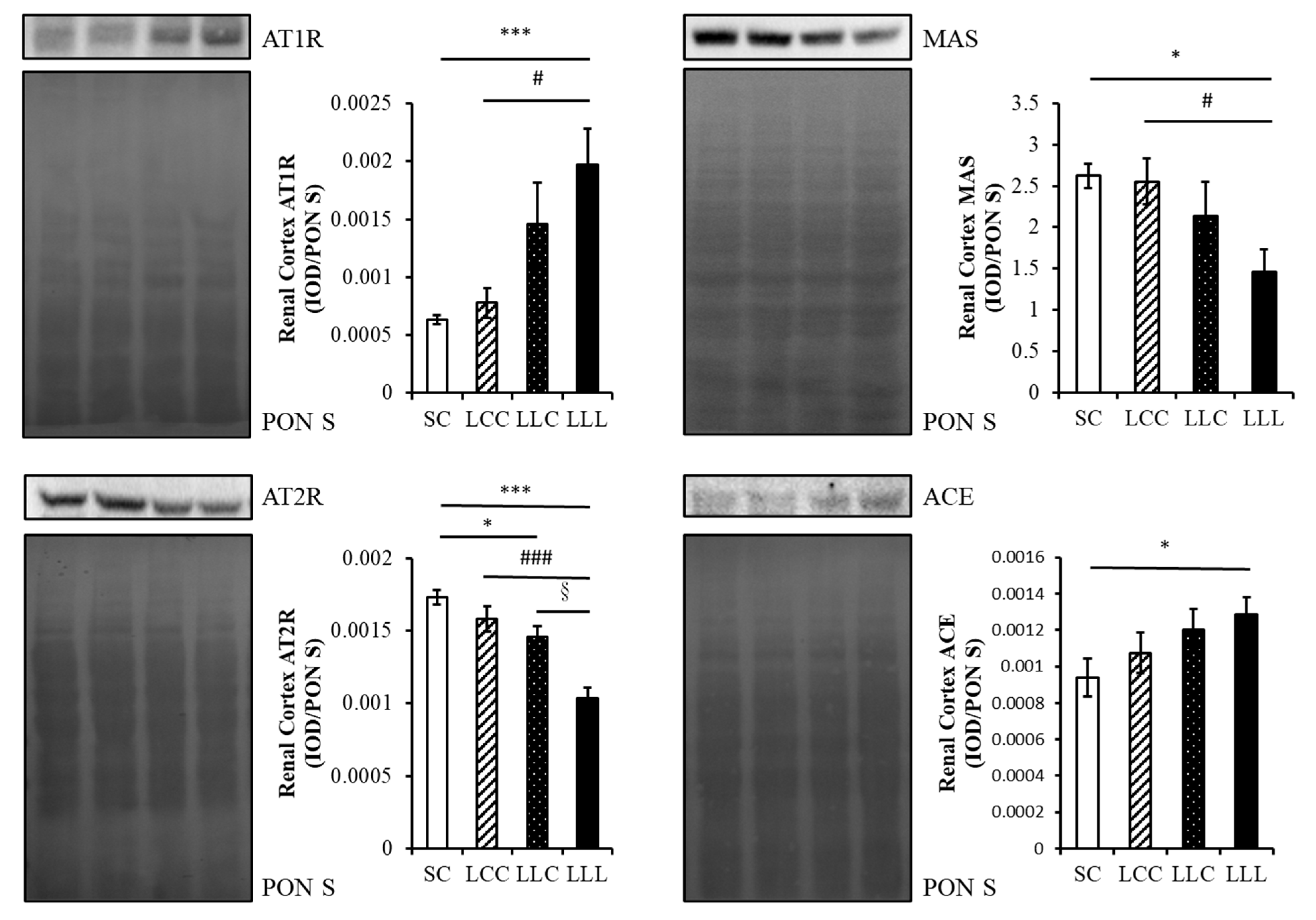
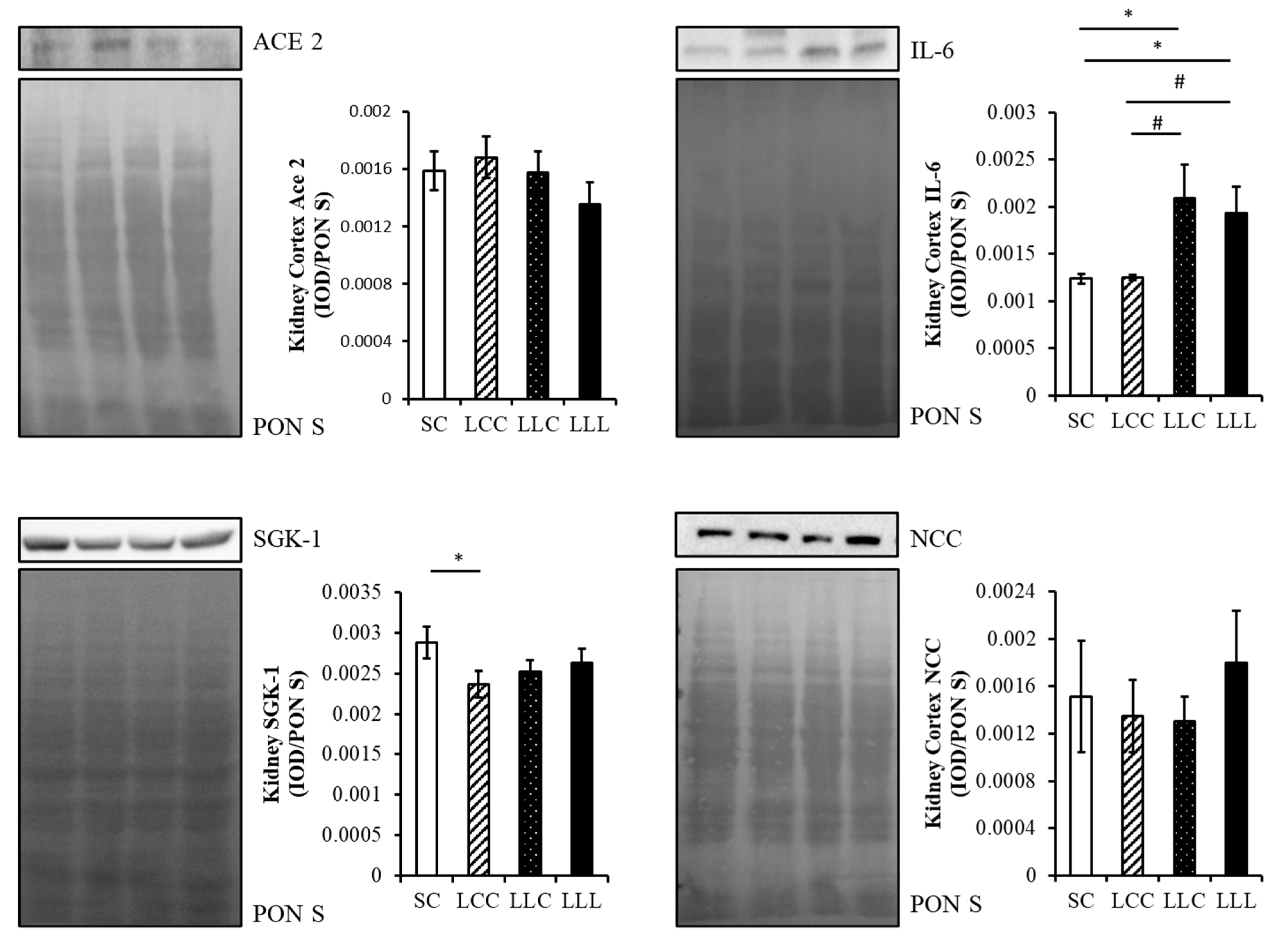
2.4. Renin Angiotensin System
2.5. Renal Cortex Sodium Transporter
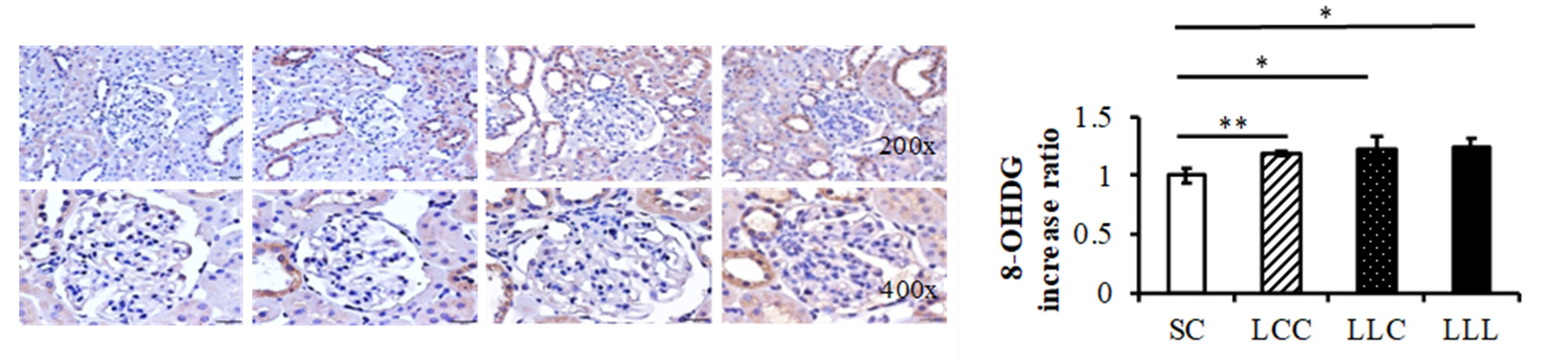
2.6. Renal Cortex IL-6 and 8-OHdG
3. Discussion
3.1. Programming Hypertension and IDA
3.2. Programming Hypertension and RAS and IDA
3.3. Programming Hypertension and Oxidative Stress and IDA
3.4. Programming Hypertension and Inflammation and IDA
3.5. Programming Hypertension and Sodium Transporter and IDA
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Blood Pressure Measurement
4.3. Immunohistochemistry
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Western Blot Assay
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Thrift, A.G.; McNeil, J.J.; Forbes, A.; Donnan, G.A. Risk factors for cerebral hemorrhage in the era of well-controlled hypertension. Melbourne Risk Factor Study (MERFS) Group. Stroke 1996, 27, 2020–2025. [Google Scholar] [CrossRef]
- Wilson, P.W. Established risk factors and coronary artery disease: The Framingham Study. Am. J. Hypertens. 1994, 7, 7S–12S. [Google Scholar] [CrossRef] [PubMed]
- Coresh, J.; Wei, G.L.; McQuillan, G.; Brancati, F.L.; Levey, A.S.; Jones, C.; Klag, M.J. Prevalence of high blood pressure and elevated serum creatinine level in the United States: Findings from the third National Health and Nutrition Examination Survey (1988–1994). Arch. Intern. Med. 2001, 161, 1207. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.Y.; McCulloch, C.E.; Darbinian, J.; Go, A.S.; Iribarren, C. Elevated blood pressure and risk of end-stage renal disease in subjects without baseline kidney disease. Arch. Intern. Med. 2005, 165, 923–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolívar, J.J. Essential hypertension: An approach to its etiology and neurogenic pathophysiology. Int. J. Hypertens. 2013, 2013, 547809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, F.; Monuteaux, M.C.; van Elburg, R.M.; Gillman, M.W.; Belfort, M.B. Systematic review and meta-analysis of preterm birth and later systolic blood pressure. Hypertension 2012, 59, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Mu, M.; Wang, S.F.; Sheng, J.; Zhao, Y.; Li, H.Z.; Hu, C.L.; Tao, F.B. Birth weight and subsequent blood pressure: A meta-analysis. Arch. Cardiovasc. Dis. 2012, 105, 99–113. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Liu, S.J.; Fu, G.J.; Zhao, Y.; Xie, Y.J.; Zhang, Y.; Wang, Y.X. The associations of high birth weight with blood pressure and hypertension in later life: A systematic review and meta-analysis. Hypertens. Res. 2013, 36, 725–735. [Google Scholar] [CrossRef]
- Arima, Y.; Nishiyama, K.; Izumiya, Y.; Kaikita, K.; Hokimoto, S.; Tsujita, K. Fetal Origins of Hypertension. Adv. Exp. Med. Biol. 2018, 1012, 41–48. [Google Scholar] [PubMed]
- Paauw, N.D.; van Rijn, B.B.; Lely, A.T.; Joles, J.A. Pregnancy as a critical window for blood pressure regulation in mother and child: Programming and reprogramming. Acta Physiol. 2017, 219, 241–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, M.B.; Hurrell, R.F. Nutritional iron deficiency. Lancet 2007, 370, 511–520. [Google Scholar] [CrossRef]
- Barker, D.J.; Winter, P.D.; Osmond, C.; Margetts, B.; Simmonds, S.J. Weight in infancy and death from ischaemic heart disease. Lancet 1989, 2, 577–580. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillman, M.W. Developmental origins of health and disease. N. Engl. J. Med. 2005, 353, 1848–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, J.S.; Cooke, C.L.; Davidge, S.T. In utero origins of hypertension: Mechanisms and targets for therapy. Physiol. Rev. 2016, 96, 549–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasinger, J.H.; Alexander, B.T. Gender differences in the developmental programming of cardiovascular disease. Clin. Sci. 2016, 130, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Painter, R.C.; Roseboom, T.J.; De Rooij, S.R. Long-term effects of prenatal stress and glucocorticoid exposure. Birth. Defects Res. C Embryo. Today 2012, 96, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, N.B.; Grigore, D.; Alexander, B.T. Developmental programming of hypertension: Insight from animal models of nutritional manipulation. Hypertension 2008, 52, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Bagby, S.P. Maternal nutrition, low nephron number, and hypertension in later life: Pathways of nutritional programming. J. Nutr. 2007, 137, 1066–1072. [Google Scholar] [CrossRef] [Green Version]
- Chong, E.; Yosypiv, I.V. Developmental programming of hypertension and kidney disease. Int. J. Nephrol. 2012, 2012, 760580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Tain, Y.L. The Double-Edged Sword Effects of Maternal Nutrition in the Developmental Programming of Hypertension. Nutrients 2018, 10, E1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yip, R.; Parvanta, I.; Cogswell, M.E.; McDonnell, S.M.; Bowman, B.A.; Grummer-Strawn, L.M.; Trowbridge, F. Recommendations to prevent and control iron deficiency in the United States. Centers for Disease Control and Prevention. MMWR Recomm. Rep. 1998, 47, 1–29. [Google Scholar]
- Brannon, P.M.; Taylor, C.L. Iron Supplementation during Pregnancy and Infancy: Uncertainties and Implications for Research and Policy. Nutrients 2017, 9, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavord, S.; Myers, B.; Robinson, S.; Allard, S.; Strong, J.; Oppenheimer C; British Committee for Standards in Haematology. UK guidelines on the management of iron deficiency in pregnancy. Br. J. Haematol. 2012, 156, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.Y.; Chen, Y.C.; Hsu, M.H.; Yu, H.R.; Su, C.H.; Tain, Y.L.; Huang, L.T.; Sheen, J.M. Maternal Iron Deficiency Programs Offspring Cognition and Its Relationship with Gastrointestinal Microbiota and Metabolites. Int. J. Environ. Res. Public Health 2020, 17, 6070. [Google Scholar] [CrossRef]
- Lewis, R.M.; Petry, C.J.; Ozanne, S.E.; Hales, C.N. Effects of maternal iron restriction in the rat on blood pressure, glucose tolerance, and serum lipids in the 3-month-old offspring. Metabolism 2001, 50, 562–567. [Google Scholar] [CrossRef]
- Lewis, R.M.; Forhead, A.J.; Petry, C.J.; Ozanne, S.E.; Hales, C.N. Long-term programming of blood pressure by maternal dietary iron restriction in the rat. Br. J. Nutr. 2002, 88, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Gambling, L.; Dunford, S.; Wallace, D.I.; Zuur, G.; Solanky, N.; Srai, S.K.; McArdle, H.J. Iron deficiency during pregnancy affects postnatal blood pressure in the rat. J. Physiol. 2003, 552, 603–610. [Google Scholar] [CrossRef]
- Alwan, N.A.; Hamamy, H. Maternal Iron Status in Pregnancy and Long-Term Health Outcomes in the Offspring. J. Pediatr. Genet. 2015, 4, 111–123. [Google Scholar]
- Lindberg, J.; Norman, M.; Westrup, B.; Domellöf, M.; Berglund, S.K. Lower systolic blood pressure at age 7 y in low-birth-weight children who received iron supplements in infancy: Results from a randomized controlled trial. Am. J. Clin. Nutr. 2017, 106, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley-Evans, S.C.; Jackson, A.A. Captopril normalises systolic blood pressure in rats with hypertension induced by fetal exposure to maternal low protein diets. Comp. Biochem. Physiol. A Physiol. 1995, 110, 223–228. [Google Scholar] [CrossRef]
- Vehaskari, V.M.; Aviles, D.H.; Manning, J. Prenatal programming of adult hypertension in the rat. Kidney Int. 2001, 59, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Vehaskari, V.M.; Stewart, T.; Lafont, D.; Soyez, C.; Seth, D.; Manning, J. Kidney angiotensin and angiotensin receptor expression in prenatally programmed hypertension. Am. J. Physiol. Ren. Physiol. 2004, 287, F262–F267. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.; Wilcox, C.S. Oxidative stress in hypertension: Role of the kidney. Antioxid. Redox. Signal 2014, 20, 74–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira-Filho, L.D.; Farias, J.S.; Cabral, E.V.; Silva, P.A.; Paixao, A.D.; Vieyra, A. Early changes on proximal tubule Na+-ATPase activity precede blood pressure elevation and renal dysfunction induced by intrauterine undernutrition: Reprogramming by alpha-tocopherol. FASEB J. 2013, 27, 907.3. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, Y.J.; Yu, H.R.; Lin, I.C.; Sheen, J.M.; Huang, L.T.; Tain, Y.L. Protection of Male Rat Offspring against Hypertension Programmed by Prenatal Dexamethasone Administration and Postnatal High-Fat Diet with the Nrf2 Activator Dimethyl Fumarate during Pregnancy. Int. J. Mol. Sci. 2019, 20, 3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira-Filho, L.D.; Lara, L.S.; Silva, P.A.; Luzardo, R.; Einicker-Lamas, M.; Cardoso, H.D.; Paixão, A.D.; Vieyra, A. Placental oxidative stress in malnourished rats and changes in kidney proximal tubule sodium ATPases in offspring. Clin. Exp. Pharmacol. Physiol. 2009, 36, 1157–1163. [Google Scholar] [CrossRef]
- Woodman, A.G.; Care, A.S.; Mansour, Y.; Cherak, S.J.; Panahi, S.; Gragasin, F.S.; Bourque, S.L. Modest and severe maternal iron deficiency in pregnancy are associated with fetal anaemia and organ-specific hypoxia in rats. Sci. Rep. 2017, 7, 46573. [Google Scholar] [CrossRef] [Green Version]
- Woodman, A.G.; Mah, R.; Keddie, D.; Noble, R.M.N.; Panahi, S.; Gragasin, F.S.; Lemieux, H.; Bourque, S.L. Prenatal iron deficiency causes sex-dependent mitochondrial dysfunction and oxidative stress in fetal rat kidneys and liver. FASEB J. 2018, 32, 3254–4263. [Google Scholar] [CrossRef] [Green Version]
- Aly, S.S.; Fayed, H.M.; Ahmed, S.S.; Abdella, A.H.; Tamam, A.A.E.; Mohmmed, N.A. Effects of oral iron (ferrous versus ferric) supplementation on oxidative stress and antioxidant status in pregnant women with iron deficiency: Controlled trial. Egypt J. Haematol. 2016, 41, 31–41. [Google Scholar]
- Ross, R. Atherosclerosis--an inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Radaelli, T.; Uvena-Celebrezze, J.; Minium, J.; Huston-Presley, L.; Catalano, P.; Hauguel-de Mouzon, S. Maternal interleukin-6: Marker of fetal growth and adiposity. J. Soc. Gynecol. Investig. 2006, 13, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Jellyman, J.K.; Han, G.; Beall, M.; Lane, R.H.; Ross, M.G. Maternal obesity and high-fat diet program offspring metabolic syndrome. Am. J. Obstet. Gynecol. 2014, 211, e1–e237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.L.; Li, X.H.; Zhou, J.Z. Prenatal exposure to lipopolysaccharide results in increases in blood pressure and body weight in rats. Acta Pharmacol. Sin. 2007, 28, 651–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertram, C.; Trowern, A.R.; Copin, N.; Jackson, A.A.; Whorwood, C.B. The maternal diet during pregnancy programs altered expression of the glucocorticoid receptor and type 2 11beta -hydroxysteroid dehydrogenase: Potential molecular mechanisms underlying the programming of hypertension in utero. Endocrinology 2001, 142, 2841–2853. [Google Scholar] [CrossRef]
- Manning, J.; Beutler, K.; Knepper, M.A.; Vehaskari, V.M. Upregulation of renal BSC1 and TSC in prenatally programmed hypertension. Am. J. Physiol. Renal Physiol. 2002, 283, F202–F206. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Sheen, J.M.; Yu, H.R.; Chen, C.C.; Tiao, M.M.; Hsu, C.N.; Lin, Y.J.; Kuo, K.C.; Huang, L.T. Maternal Melatonin Therapy Rescues Prenatal Dexamethasone and Postnatal High-Fat Diet Induced Programmed Hypertension in Male Rat Offspring. Front. Physiol. 2015, 6, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esen Ağar, B.; Akarsu, S.; Aydin, S. The Effect of Iron Deficiency Anemia and Different Treatment Methods on DNA Damage: 8-hydroxy-2-deoxyguanosine Level. Glob. Pediatr. Health 2021, 8, 2333794X211041337. [Google Scholar] [CrossRef]
- Mian, M.O.; Paradis, P.; Schiffrin, E.L. Innate immunity in hypertension. Curr. Hypertens. Rep. 2014, 16, 413. [Google Scholar] [CrossRef]
- Harrison, D.G.; Guzik, T.J.; Lob, H.E.; Madhur, M.S.; Marvar, P.J.; Thabet, S.R.; Vinh, A.; Weyand, C.M. Inflammation, immunity, and hypertension. Hypertension 2011, 57, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stumpf, C.; John, S.; Jukic, J.; Yilmaz, A.; Raaz, D.; Schmieder, R.E.; Daniel, W.G.; Garlichs, C.D. Enhanced levels of platelet P-selectin and circulating cytokines in young patients with mild arterial hypertension. J. Hypertens. 2005, 23, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Vallon, V.; Schroth, J.; Lang, F.; Kuhl, D.; Uchida, S. Expression and phosphorylation of the Na+-Cl- cotransporter NCC in vivo is regulated by dietary salt, potassium, and SGK1. Am. J. Physiol. Renal Physiol. 2009, 297, F704–F712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheen, J.M.; Yu, H.R.; Tain, Y.L.; Tsai, W.L.; Tiao, M.M.; Lin, I.C.; Tsai, C.C.; Lin, Y.J.; Huang, L.T. Combined maternal and postnatal high-fat diet leads to metabolic syndrome and is effectively reversed by resveratrol: A multiple-organ study. Sci. Rep. 2018, 8, 5607. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day-Old | 42 | 49 | 56 | 63 | |
|---|---|---|---|---|---|
| Body weight (g) | Control diet | 180.5 ± 16.9 | 200.5 ± 13.5 | 221.9 ± 16.9 | 239.8 ± 14.1 |
| Low-iron diet | 184 ± 10 | 204 ± 7.2 | 228.5 ± 10 | 243.7 ± 13.2 | |
| Serum Iron (μM) | Control diet | 58 ± 7.4 | 58.2 ± 2.7 | 70.1 ± 9.7 | 42.6 ± 5.9 |
| Low-iron diet | 38.3 ± 1.2 | 40.5 ± 0.6 | 23.9 ± 4.2 ** | 16.4 ± 4.5 ** | |
| Hemoglobin (g/dL) | Control diet | 14.5 ± 0.1 | 14.7 ± 0.4 | 14.4 ± 0.5 | 14.8 ± 0.1 |
| Low-iron diet | 14.2 ± 0.1 | 14.6 ± 0.3 | 13.3 ± 0.5 | 12± 0.6 * |
| Group | SC | LCC | LLC | LLL |
|---|---|---|---|---|
| Mean ± SD | ||||
| Body weight (g) | 567.1 ± 6.2 | 542.1 ± 24.8 | 552.4 ± 24.7 | 463.75 ± 13.3 ** # § |
| Serum Iron (μM) | 33.5 ± 1.5 | 28.5 ± 5.8 | 22.6 ± 5.2 | 19.6 ± 4.2 * |
| Hemoglobin (g/dL) | 15.9 ± 0.3 | 16 ± 0.2 | 15.9 ± 0.5 | 15.5 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-H.; Chen, W.-H.; Su, C.-H.; Yu, H.-R.; Tain, Y.-L.; Huang, L.-T.; Sheen, J.-M. Maternal Iron Deficiency Programs Rat Offspring Hypertension in Relation to Renin—Angiotensin System and Oxidative Stress. Int. J. Mol. Sci. 2022, 23, 8294. https://doi.org/10.3390/ijms23158294
Chang Y-H, Chen W-H, Su C-H, Yu H-R, Tain Y-L, Huang L-T, Sheen J-M. Maternal Iron Deficiency Programs Rat Offspring Hypertension in Relation to Renin—Angiotensin System and Oxidative Stress. International Journal of Molecular Sciences. 2022; 23(15):8294. https://doi.org/10.3390/ijms23158294
Chicago/Turabian StyleChang, Ya-Hui, Wan-Hsuan Chen, Chung-Hao Su, Hong-Ren Yu, You-Lin Tain, Li-Tung Huang, and Jiunn-Ming Sheen. 2022. "Maternal Iron Deficiency Programs Rat Offspring Hypertension in Relation to Renin—Angiotensin System and Oxidative Stress" International Journal of Molecular Sciences 23, no. 15: 8294. https://doi.org/10.3390/ijms23158294