
Integrated CRISPR-Cas9 System-Mediated Knockout of IFN-γ and IFN-γ Receptor 1 in the Vero Cell Line Promotes Viral Susceptibility


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
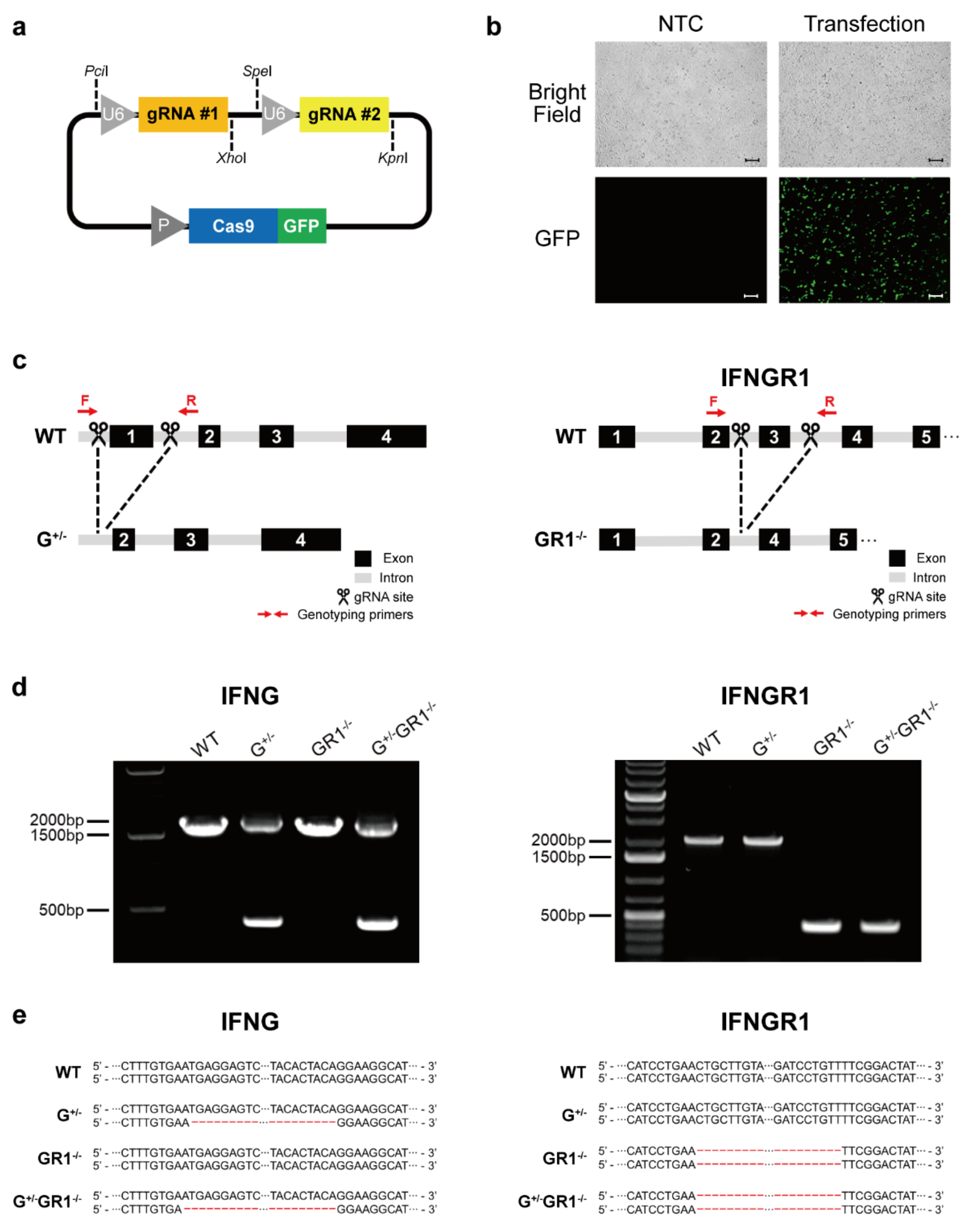
2.1. Generation of IFNG- and IFNGR1-Knockout Cell Lines Using an Integrated CRISPR-Cas9 System
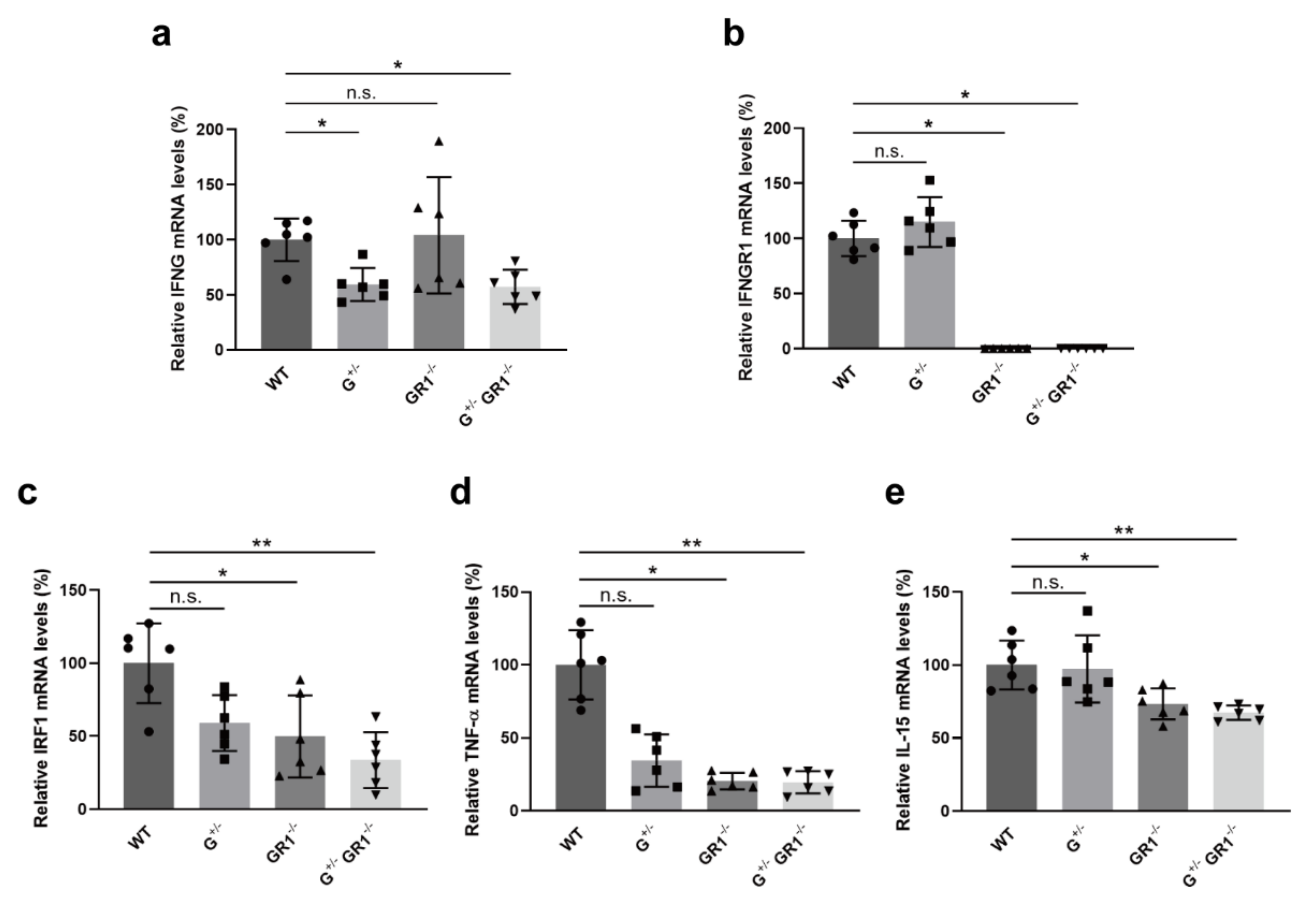
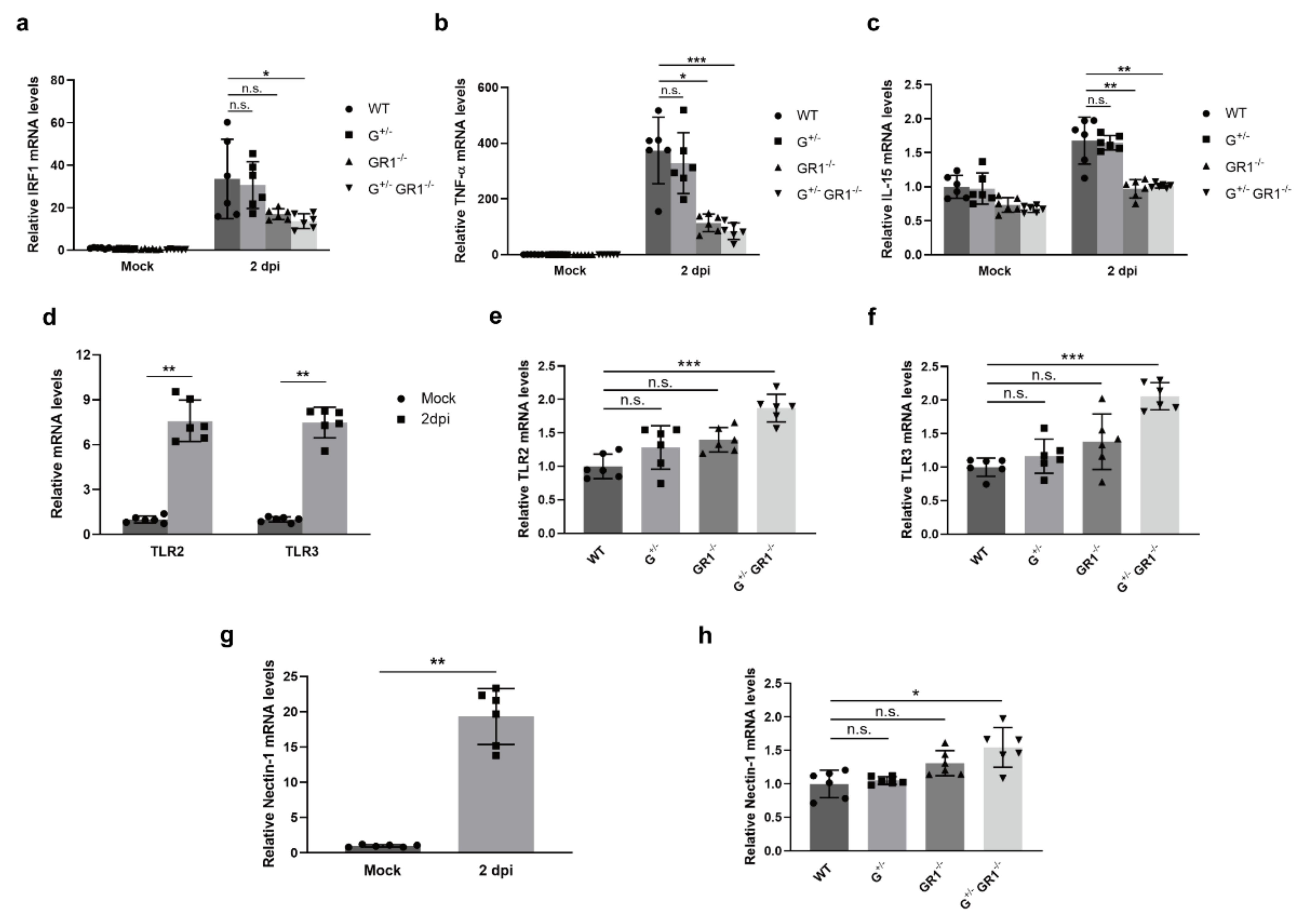
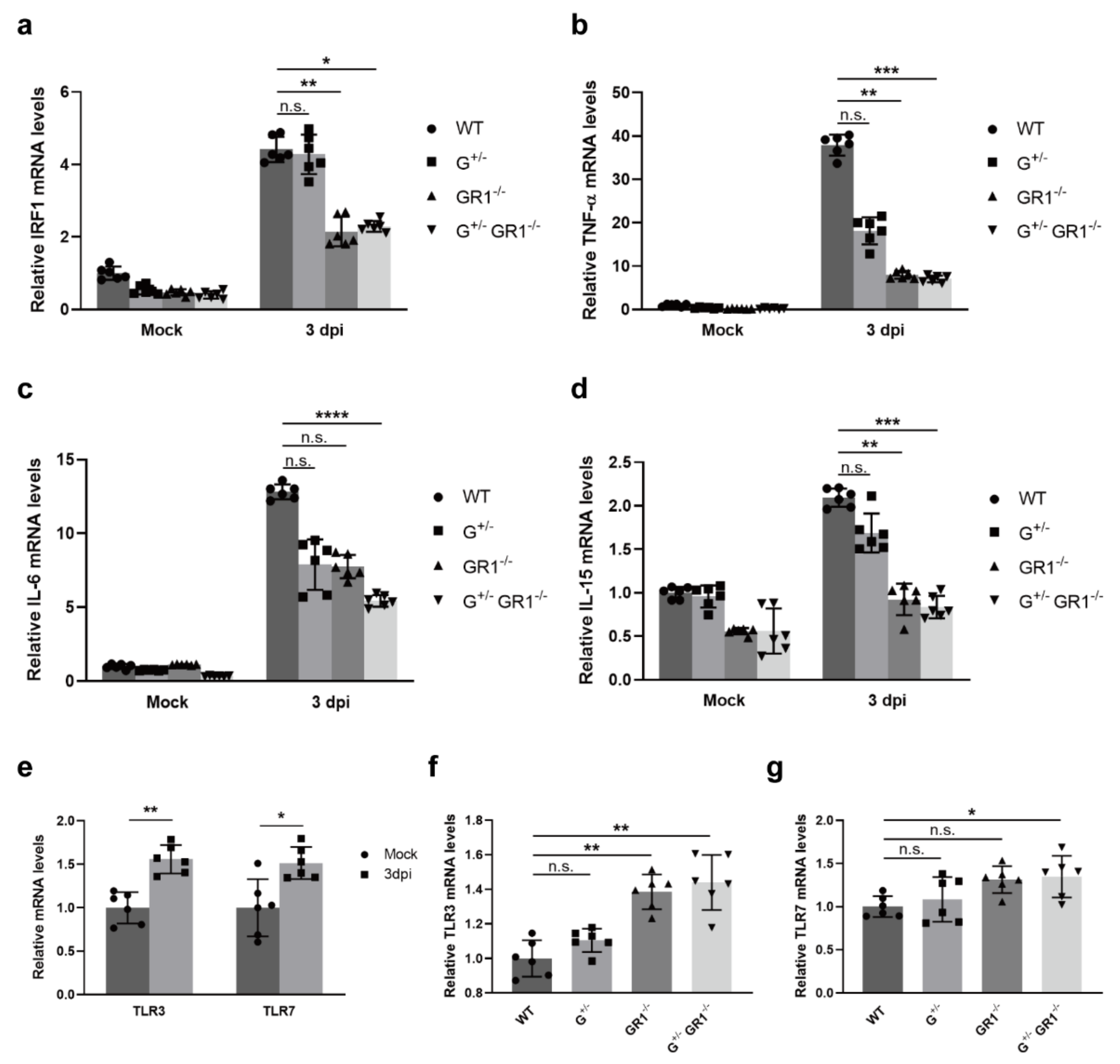
2.2. Downregulation of Immune-Related Genes in IFNG- and IFNGR1-Deficient Cells

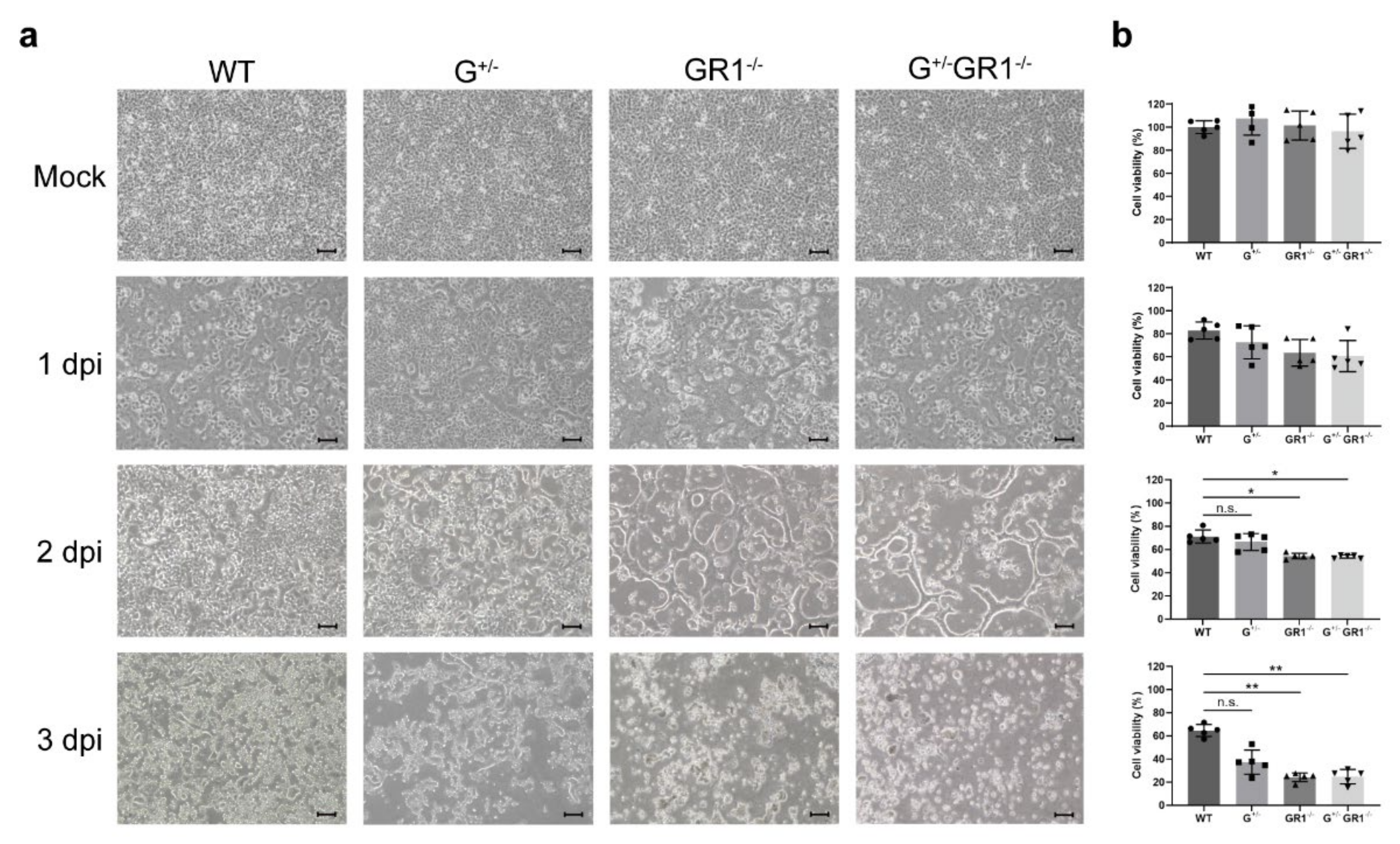
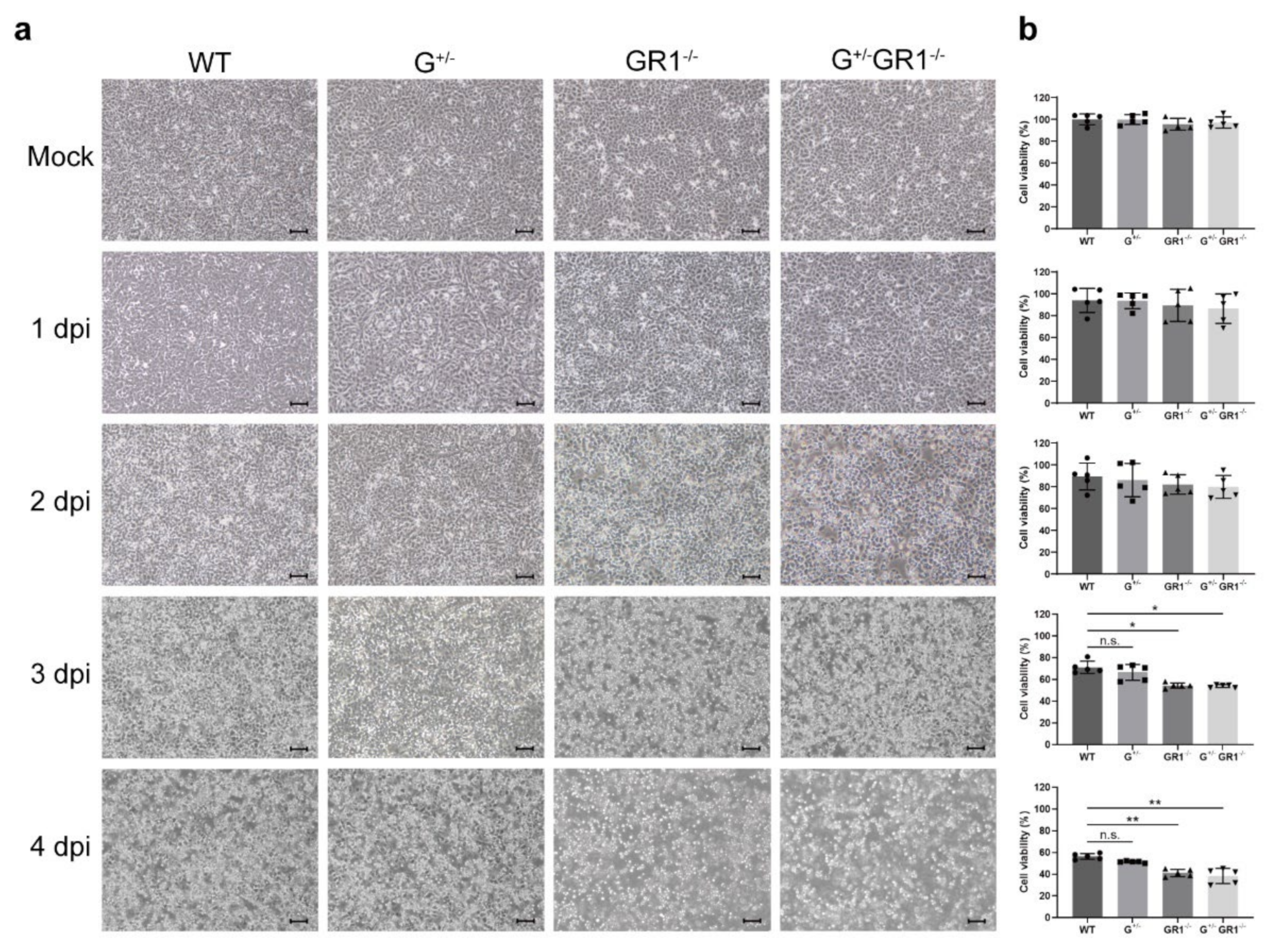
2.3. Enhanced Cytopathic Effect of HSV-1 Infection in IFNG- and IFNGR1-Knockout Vero Cells
2.4. Downregulation of the Immune Response to HSV-1 Infection in IFNG- and IFNGR1-Knockout Vero Cell Lines
2.5. Enhanced Cytopathic Effects in IFNG- and IFNGR1-Knockout Vero Cells after HCoV-OC43 Infection
2.6. Downregulation of the Immune Response to Coronavirus Infection in IFNG- and IFNGR1-Knockout Vero Cell Lines
3. Discussion
4. Materials and Methods
4.1. Integrated CRISPR-Cas9 Vector System
4.2. Design of gRNAs and the Construction of IFNG- and IFNGR1-Targeting Plasmids
4.3. Cell Culture and Transfection
4.4. Single Cell Isolation
4.5. PCR-Based Genotyping of Knockout Cell Lines
4.6. Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-qPCR)
4.7. Viruses
4.8. Virus Infection and Monitoring of Cytopathic Effects
4.9. Virus Quantification by Endpoint Dilution Assay
4.10. MTT Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Govorkova, E.A.; Kaverin, N.V.; Gubareva, L.V.; Meignier, B.; Webster, R.G. Replication of influenza A viruses in a green monkey kidney continuous cell line (Vero). J. Infect. Dis. 1995, 172, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Desmyter, J.; Melnick, J.L.; Rawls, W.E. Defectiveness of interferon production and of rubella virus interference in a line of African green monkey kidney cells (Vero). J. Virol. 1968, 2, 955–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiesslich, S.; Kamen, A.A. Vero cell upstream bioprocess development for the production of viral vectors and vaccines. Biotechnol. Adv. 2020, 44, 107608. [Google Scholar] [CrossRef] [PubMed]
- Loo, S.L.; Wark, P.A.B.; Esneau, C.; Nichol, K.S.; Hsu, A.C.; Bartlett, N.W. Human coronaviruses 229E and OC43 replicate and induce distinct antiviral responses in differentiated primary human bronchial epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L926–L931. [Google Scholar] [CrossRef] [PubMed]
- Ogando, N.S.; Dalebout, T.J.; Zevenhoven-Dobbe, J.C.; Limpens, R.; van der Meer, Y.; Caly, L.; Druce, J.; de Vries, J.J.C.; Kikkert, M.; Barcena, M.; et al. SARS-coronavirus-2 replication in Vero E6 cells: Replication kinetics, rapid adaptation and cytopathology. J. Gen. Virol. 2020, 101, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Hirose, R.; Watanabe, N.; Bandou, R.; Yoshida, T.; Daidoji, T.; Naito, Y.; Itoh, Y.; Nakaya, T. A Cytopathic Effect-Based Tissue Culture Method for HCoV-OC43 Titration Using TMPRSS2-Expressing VeroE6 Cells. mSphere 2021, 6, e00159-21. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, S.; Fernandez-Antunez, C.; Galli, A.; Underwood, A.; Pham, L.V.; Ryberg, L.A.; Feng, S.; Pedersen, M.S.; Mikkelsen, L.S.; Belouzard, S.; et al. Overcoming Culture Restriction for SARS-CoV-2 in Human Cells Facilitates the Screening of Compounds Inhibiting Viral Replication. Antimicrob. Agents Chemother. 2021, 65, e0009721. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Jyothi Prasanna, S.; Chandrasekar, B.; Nandi, D. Gene modulation and immunoregulatory roles of interferon gamma. Cytokine 2010, 50, 1–14. [Google Scholar] [CrossRef]
- Sainz, B., Jr.; Halford, W.P. Alpha/Beta interferon and gamma interferon synergize to inhibit the replication of herpes simplex virus type 1. J. Virol. 2002, 76, 11541–11550. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.Y.; Schultz, K.L.; Griffin, D.E. Mice deficient in interferon-gamma or interferon-gamma receptor 1 have distinct inflammatory responses to acute viral encephalomyelitis. PLoS ONE 2013, 8, e76412. [Google Scholar] [CrossRef] [Green Version]
- Van den Broek, M.F.; Muller, U.; Huang, S.; Aguet, M.; Zinkernagel, R.M. Antiviral defense in mice lacking both alpha/beta and gamma interferon receptors. J. Virol. 1995, 69, 4792–4796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knott, G.J.; Doudna, J.A. CRISPR-Cas guides the future of genetic engineering. Science 2018, 361, 866–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Doudna, J.A. CRISPR-Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adikusuma, F.; Pfitzner, C.; Thomas, P.Q. Versatile single-step-assembly CRISPR/Cas9 vectors for dual gRNA expression. PLoS ONE 2017, 12, e0187236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Herrera-Carrillo, E.; Berkhout, B. A Single H1 Promoter Can Drive Both Guide RNA and Endonuclease Expression in the CRISPR-Cas9 System. Mol. Ther.-Nucleic Acids 2019, 14, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Aguirre, S.; Arancibia, D.; Escorza, A.; Bravo, C.; Andres, M.E.; Zamorano, P.; Martinez, V. Development of a Bicistronic Vector for the Expression of a CRISPR/Cas9-mCherry System in Fish Cell Lines. Cells 2019, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, H.; Xue, B.; Deng, R.; Huang, X.; Xu, Y.; Chen, S.; Tian, R.; Wang, X.; Xun, Z.; et al. IRF1 Promotes the Innate Immune Response to Viral Infection by Enhancing the Activation of IRF3. J. Virol. 2020, 94, e01231-20. [Google Scholar] [CrossRef]
- Li, X.Q.; Li, X.N.; Liang, J.J.; Cai, X.B.; Tao, Q.; Li, Y.X.; Qin, Q.; Xu, S.P.; Luo, T.R. IRF1 up-regulates isg15 gene expression in dsRNA stimulation or CSFV infection by targeting nucleotides -487 to -325 in the 5′ flanking region. Mol. Immunol. 2018, 94, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Vila-del Sol, V.; Punzon, C.; Fresno, M. IFN-gamma-induced TNF-alpha expression is regulated by interferon regulatory factors 1 and 8 in mouse macrophages. J. Immunol. 2008, 181, 4461–4470. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Shin, M.S.; Kang, K.S.; Yoo, S.A.; Mohanty, S.; Montgomery, R.R.; Shaw, A.C.; Kang, I. Human monocytes have increased IFN-gamma-mediated IL-15 production with age alongside altered IFN-gamma receptor signaling. Clin. Immunol. 2014, 152, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; El Bassam, S.; Cordeiro, P.; Menezes, J. Requirement of TLR2-mediated signaling for the induction of IL-15 gene expression in human monocytic cells by HSV-1. Blood 2008, 112, 2360–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motamedifar, M.; Noorafshan, A. Cytopathic effect of the herpes simplex virus type 1 appears stereologically as early as 4 h after infection of Vero cells. Micron 2008, 39, 1331–1334. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Melchjorsen, J.; Matikainen, S.; Paludan, S.R. Activation and evasion of innate antiviral immunity by herpes simplex virus. Viruses 2009, 1, 737–759. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Xu, Q.; Zhang, Y.; Gao, W.; Zhang, M.; Zhai, W.; Rajkumar, R.S.; Liu, Z. Toll-like receptor-mediated innate immunity against herpesviridae infection: A current perspective on viral infection signaling pathways. Virol. J. 2020, 17, 192. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Y.; Gao, S.Q.; Gong, Y.S.; Lin, T.; Tong, S.; Xiong, W.; Shi, C.Y.; Wang, W.Q.; Fang, J.G. Anti-HSV-1 effect of dihydromyricetin from Ampelopsis grossedentata via the TLR9-dependent anti-inflammatory pathway. J. Glob. Antimicrob. Resist. 2020, 23, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Agelidis, A.M.; Shukla, D. Cell entry mechanisms of HSV: What we have learned in recent years. Future Virol. 2015, 10, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Peri, P.; Mattila, R.K.; Kantola, H.; Broberg, E.; Karttunen, H.S.; Waris, M.; Vuorinen, T.; Hukkanen, V. Herpes simplex virus type 1 Us3 gene deletion influences toll-like receptor responses in cultured monocytic cells. Virol. J. 2008, 5, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, D.; Scanlan, P.M.; Tiwari, V.; Sheth, V.; Clement, C.; Guzman-Hartman, G.; Dermody, T.S.; Valyi-Nagy, T. Expression of nectin-1 in normal and herpes simplex virus type 1-infected murine brain. Appl. Immunohistochem. Mol. Morphol. 2006, 14, 341–347. [Google Scholar] [CrossRef]
- Kesheh, M.M.; Hosseini, P.; Soltani, S.; Zandi, M. An overview on the seven pathogenic human coronaviruses. Rev. Med. Virol. 2022, 32, e2282. [Google Scholar] [CrossRef] [PubMed]
- Lambert, F.; Jacomy, H.; Marceau, G.; Talbot, P.J. Titration of human coronaviruses using an immunoperoxidase assay. J. Vis. Exp. 2008, 14, e751. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Min, J.S.; Jang, M.S.; Lee, J.Y.; Shin, Y.S.; Song, J.H.; Kim, H.R.; Kim, S.; Jin, Y.H.; Kwon, S. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells. Biomolecules 2019, 9, 696. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.A.; Denis, F.; Talbot, P.J. Activation of glial cells by human coronavirus OC43 infection. J. Neuroimmunol. 2000, 108, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Channappanavar, R.; Kanneganti, T.D. Coronaviruses: Innate Immunity, Inflammasome Activation, Inflammatory Cell Death, and Cytokines. Trends Immunol. 2020, 41, 1083–1099. [Google Scholar] [CrossRef]
- Chesler, D.A.; Reiss, C.S. The role of IFN-gamma in immune responses to viral infections of the central nervous system. Cytokine Growth Factor Rev. 2002, 13, 441–454. [Google Scholar] [CrossRef]
- Pierce, A.T.; DeSalvo, J.; Foster, T.P.; Kosinski, A.; Weller, S.K.; Halford, W.P. Beta interferon and gamma interferon synergize to block viral DNA and virion synthesis in herpes simplex virus-infected cells. J. Gen. Virol. 2005, 86, 2421–2432. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, C.; Wang, B.; Li, B.; Wang, Q.; Liu, D.; Wang, H.; Zhou, Y.; Shi, L.; Lan, F.; et al. Optimized CRISPR guide RNA design for two high-fidelity Cas9 variants by deep learning. Nat. Commun. 2019, 10, 4284. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Nowakowska, A.; Kim, Y.B.; Shin, H.Y. Integrated CRISPR-Cas9 System-Mediated Knockout of IFN-γ and IFN-γ Receptor 1 in the Vero Cell Line Promotes Viral Susceptibility. Int. J. Mol. Sci. 2022, 23, 8217. https://doi.org/10.3390/ijms23158217
Kim S, Nowakowska A, Kim YB, Shin HY. Integrated CRISPR-Cas9 System-Mediated Knockout of IFN-γ and IFN-γ Receptor 1 in the Vero Cell Line Promotes Viral Susceptibility. International Journal of Molecular Sciences. 2022; 23(15):8217. https://doi.org/10.3390/ijms23158217
Chicago/Turabian StyleKim, Suyeon, Aleksandra Nowakowska, Young Bong Kim, and Ha Youn Shin. 2022. "Integrated CRISPR-Cas9 System-Mediated Knockout of IFN-γ and IFN-γ Receptor 1 in the Vero Cell Line Promotes Viral Susceptibility" International Journal of Molecular Sciences 23, no. 15: 8217. https://doi.org/10.3390/ijms23158217