
Effects and Mechanism of Particulate Matter on Tendon Healing Based on Integrated Analysis of DNA Methylation and RNA Sequencing Data in a Rat Model

 ,
,  , , , , , ,
, , , , , ,  ,
,  and add
Show full author list
and add
Show full author list
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
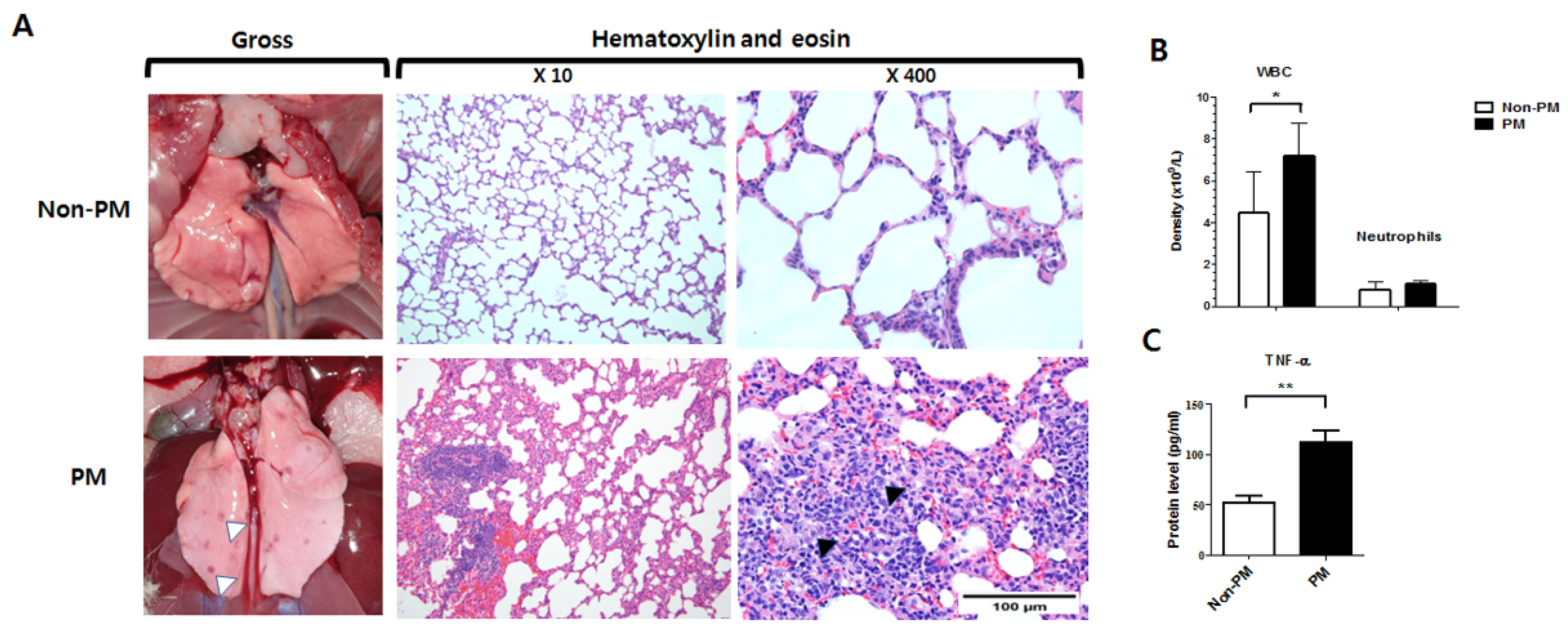
2.1. PM Exposure Induced Lung Inflammation
2.2. PM Exposure Induced Systemic Inflammation Demonstrated by Laboratory Blood Test and ELISA (Enzyme-Linked Immunosorbent Assay)
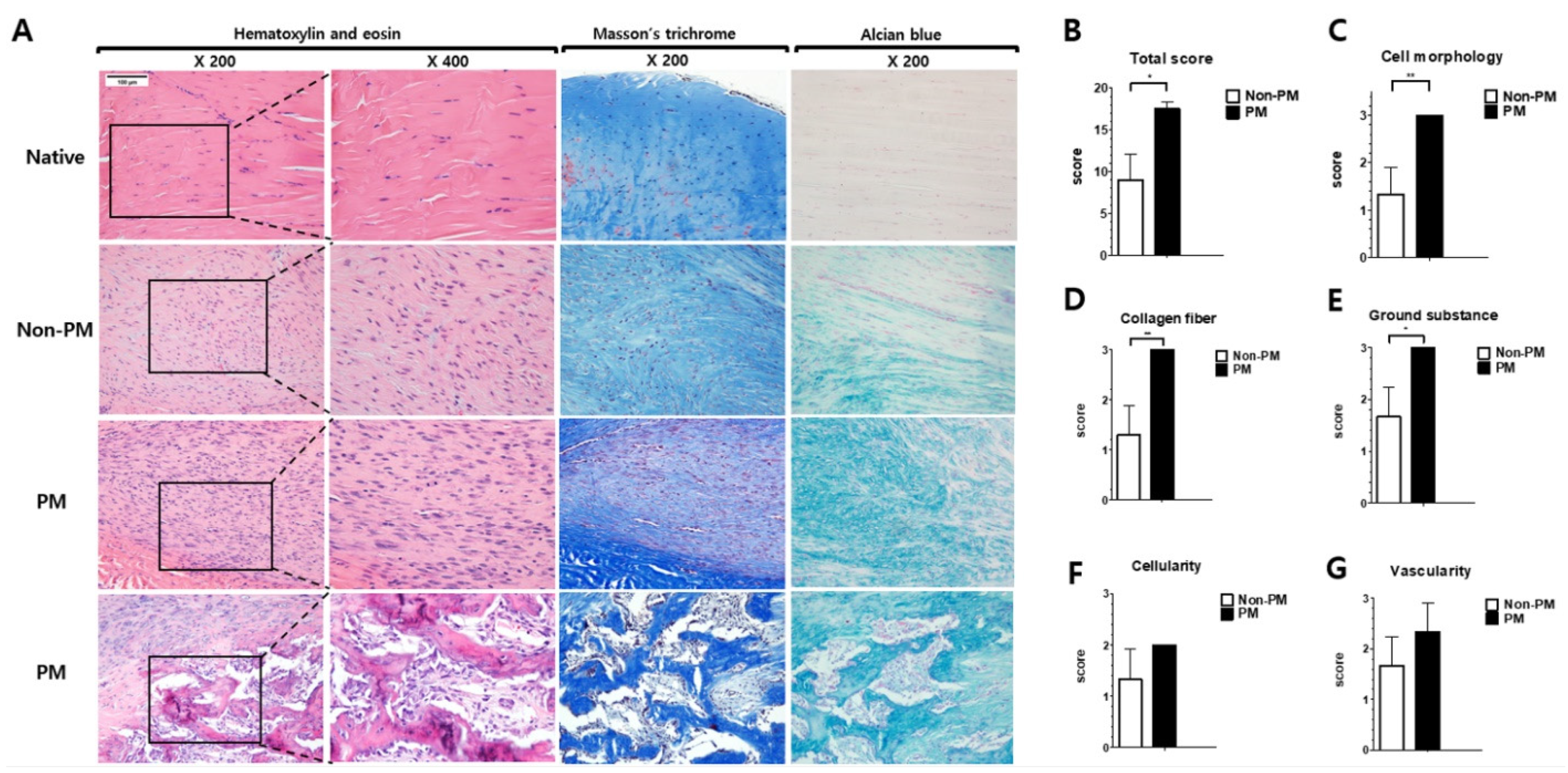
2.3. PM Exposure Had Detrimental Histological Effects on Tendon Healing in Rats with ATR
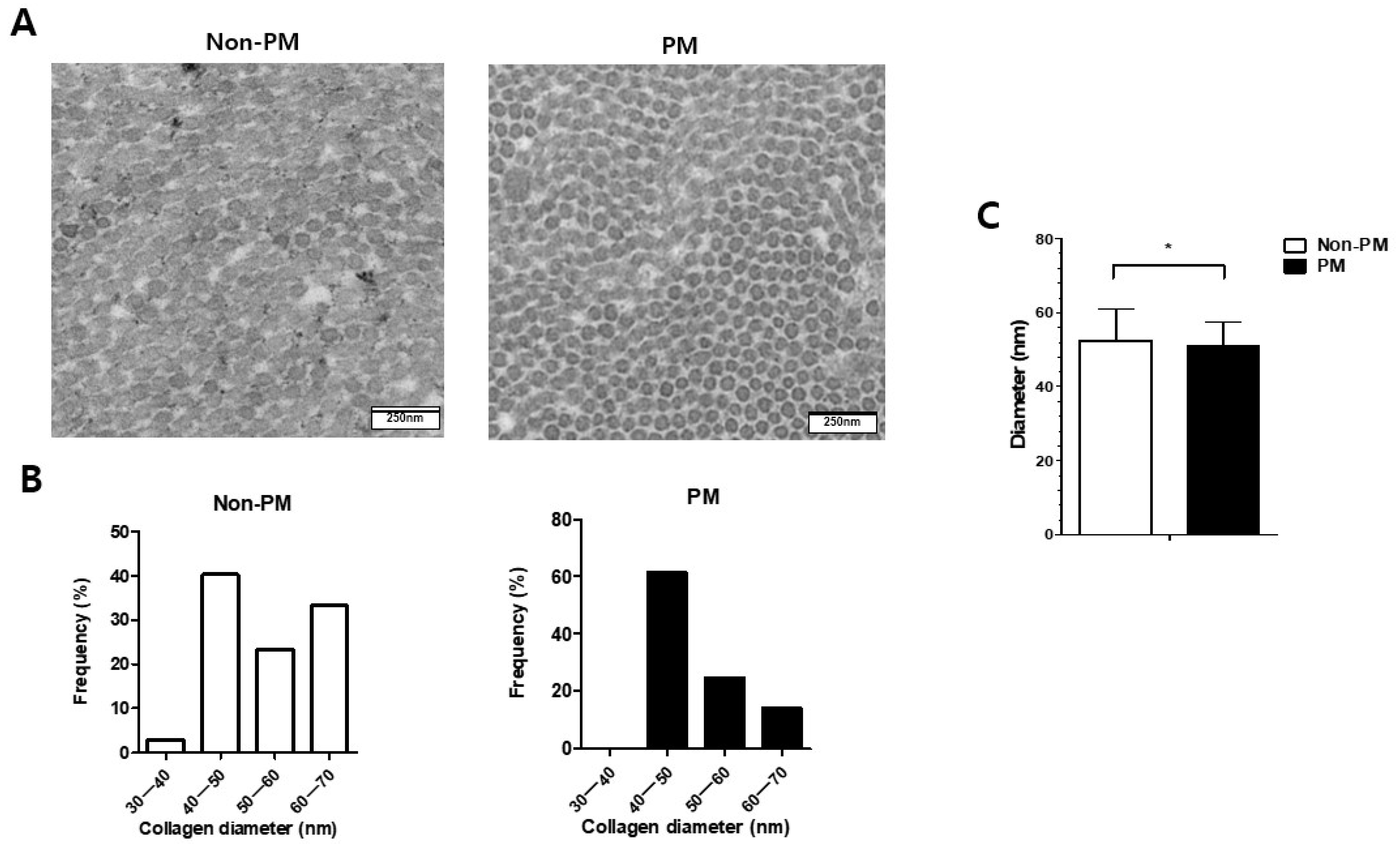
2.4. Transmission Electron Microscopy (TEM) Analysis Revealed Decreased Mean Diameter of Collagen Fibrils in PM Group
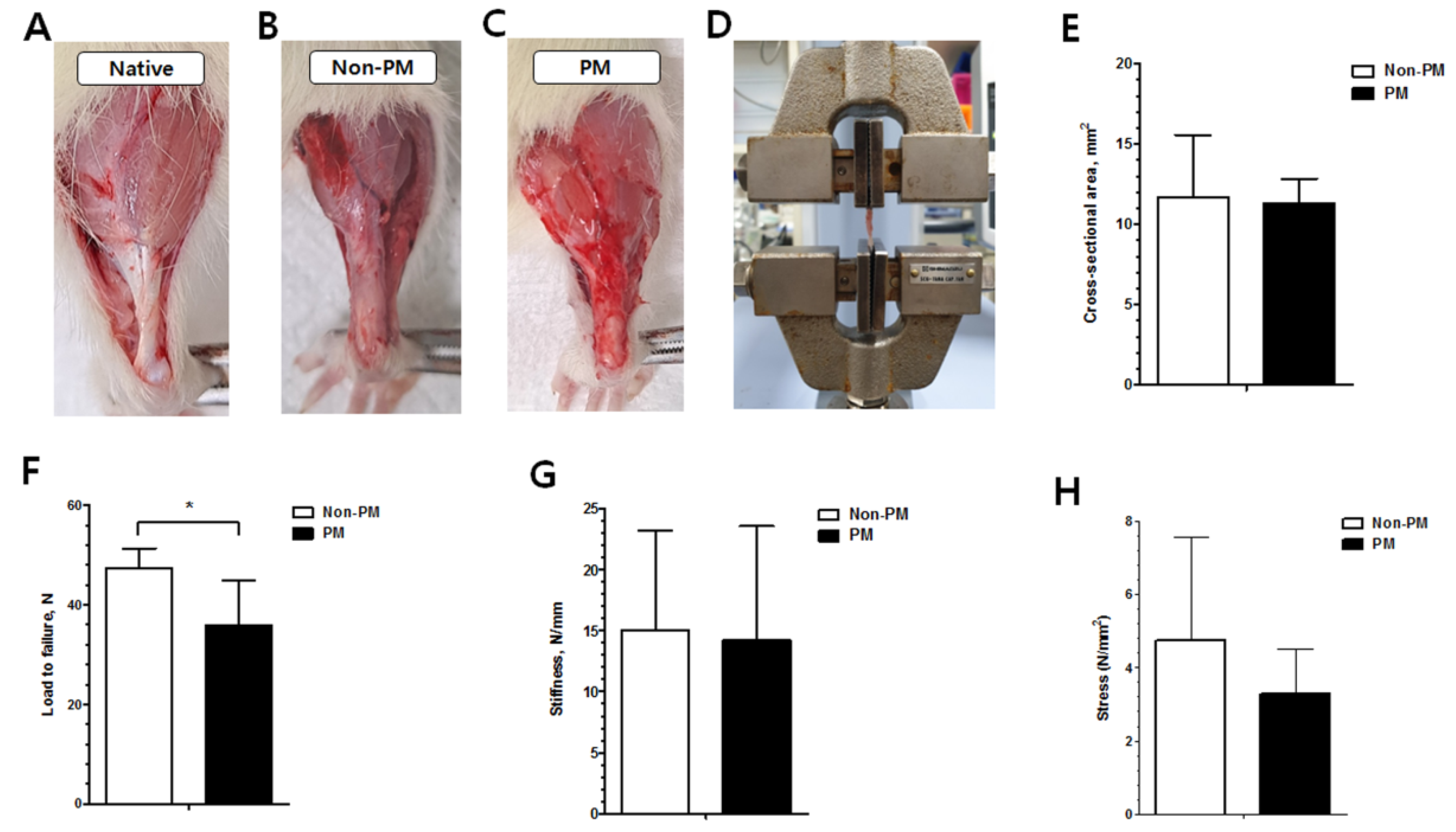
2.5. The Load to Failure of the PM Group Was Significantly Decreased Compared to the Non-PM Group
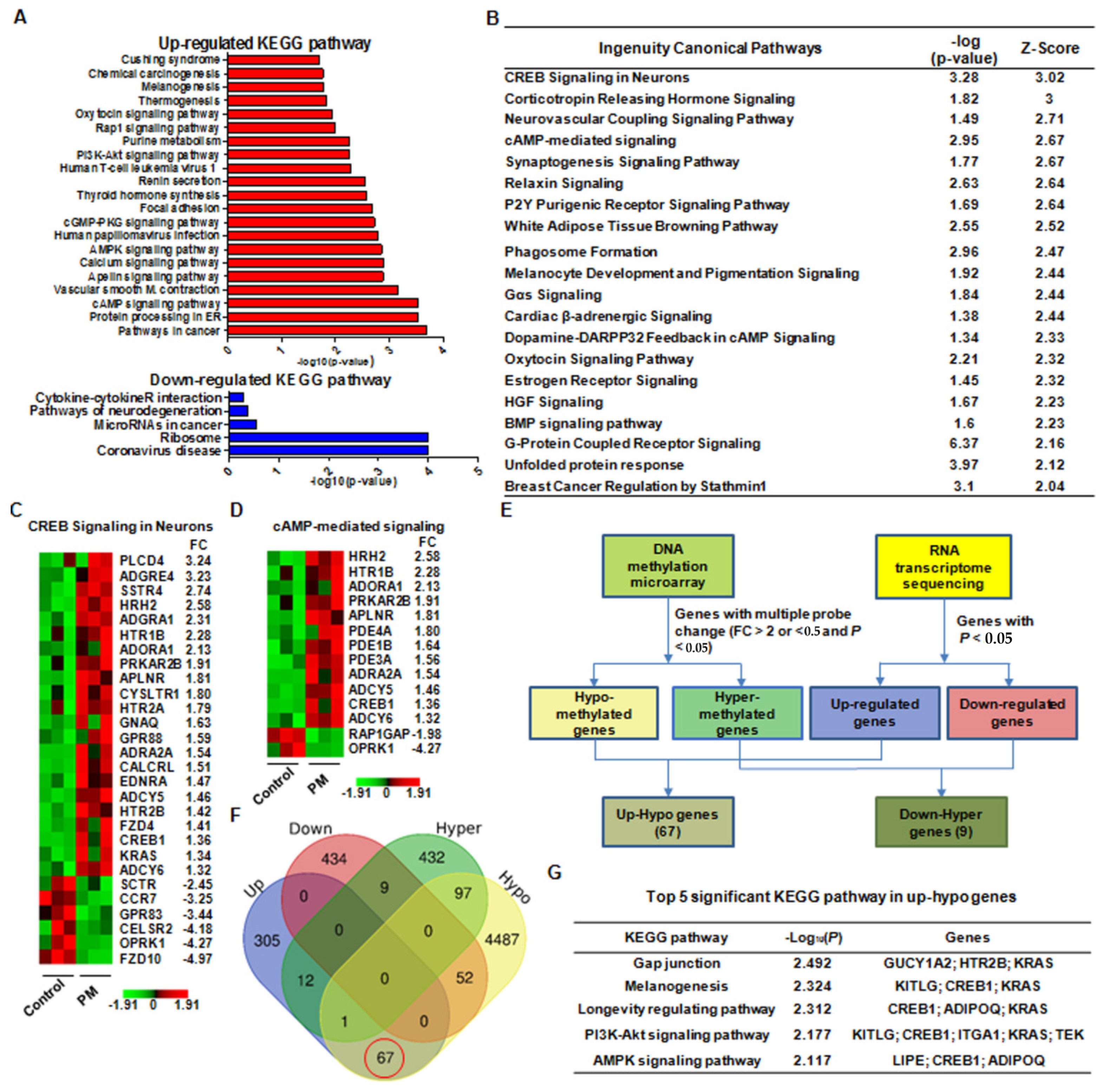
2.6. Integrated Analysis of DNA Methylation and RNA-Seq Data Implied That PM May Have a Considerable Effect on Tendon Healing through cAMP Response Element-Binding Protein 1 (CREB1) Upregulation
2.7. p-CREB Expression Was Significantly Elevated in the PM Group Compared to the Non-PM Group in the Western Blot Analysis
3. Discussion
4. Materials and Methods
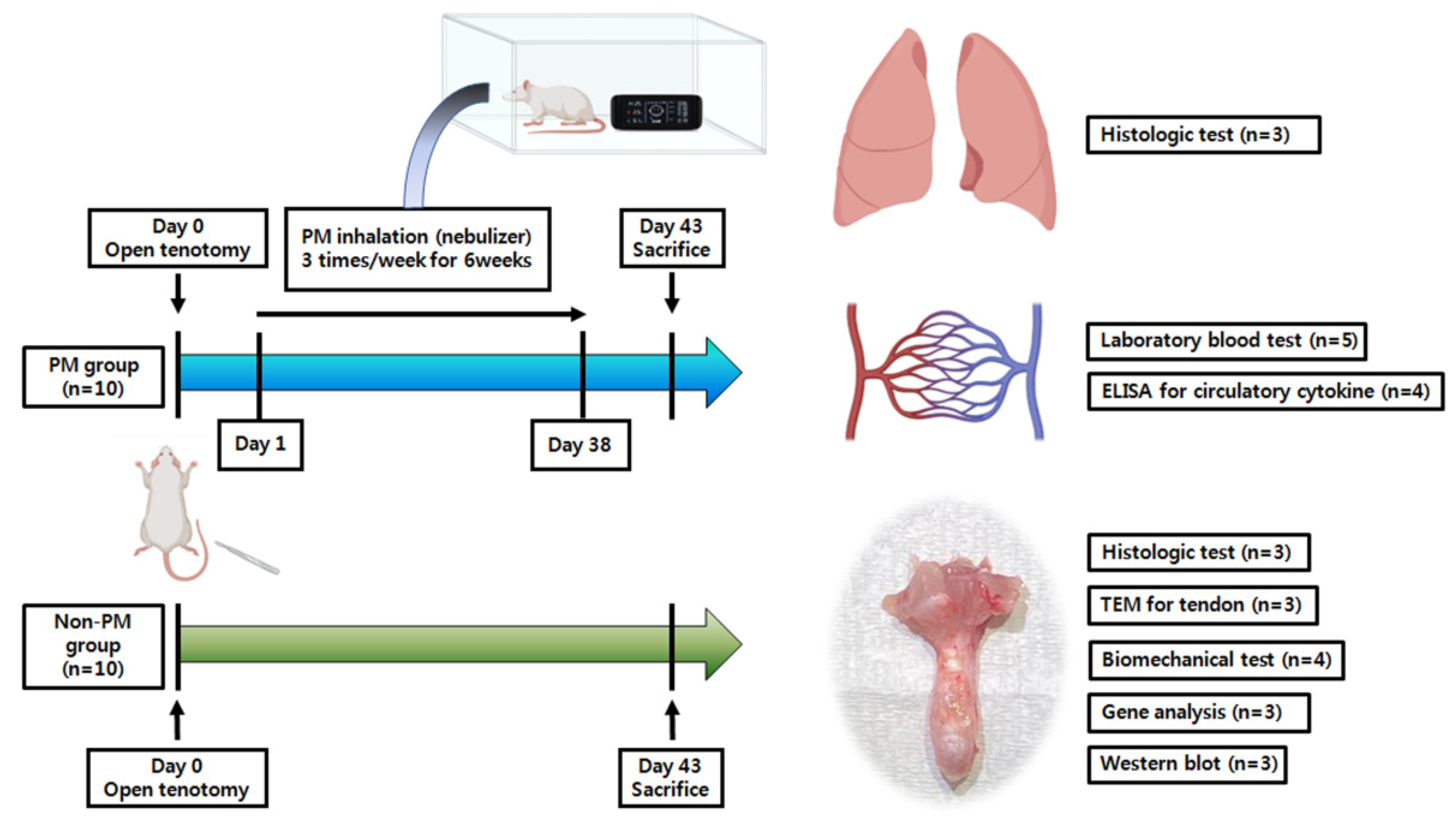
4.1. Animals and Experimental Outline
4.2. Open Achilles Tenotomy
4.3. PM Collection and Exposure
4.4. Flow Simulation in the Chamber
4.5. Laboratory Blood Test and ELISA
4.6. Histological Analysis
4.7. TEM Analysis
4.8. Biomechanical Assessment
4.9. DNA Methylation Microarray
4.10. RNA-Seq Analysis
4.11. Combined Analysis of DNA Methylation and RNA-Seq Data
4.12. Western Blot Analysis
4.13. Statistical Analysis
5. Conclusions
Limitations and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kushta, J.; Paisi, N.; Van Der Gon, H.D.; Lelieveld, J. Disease burden and excess mortality from coal-fired power plant emissions in Europe. Environ. Res. Lett. 2021, 16, 045010. [Google Scholar] [CrossRef]
- Kim, B.-U.; Kim, O.; Kim, H.C.; Kim, S. Influence of fossil-fuel power plant emissions on the surface fine particulate matter in the Seoul Capital Area, South Korea. J. Air Waste Manag. Assoc. 2016, 66, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Agrawal, M. A global perspective of fine particulate matter pollution and its health effects. Rev. Environ. Contam. Toxicol. 2017, 244, 5–51. [Google Scholar]
- Bernstein, J.; Alexis, N.; Barnes, C.; Bernstein, I.; Nel, A.; Peden, D. Health effects of air pollution. J. Allergy.Clin. Immunol. 2004, 114, 1116–1123. [Google Scholar] [CrossRef]
- Lehner, C.; Spitzer, G.; Langthaler, P.; Jakubecova, D.; Klein, B.; Weissenbacher, N.; Wagner, A.; Gehwolf, R.; Trinka, E.; Iglseder, B. Allergy-induced systemic inflammation impairs tendon quality. bioRxiv 2021, 75, 103778. [Google Scholar] [CrossRef]
- Park, S.J.; Ku, G.W.; Lee, S.Y.; Kang, D.; Hwang, W.J.; Jeong, I.B.; Kwon, S.J.; Kang, J.; Son, J.W. Analysis of Single Nucleotide Variants (SNVs) Induced by Exposure to PM10 in Lung Epithelial Cells Using Whole Genome Sequencing. Int. J. Environ. Res. Public Health 2021, 18, 1046. [Google Scholar] [CrossRef]
- Kang, D.; Jung, I.B.; Lee, S.Y.; Park, S.J.; Kwon, S.J.; Park, D.H.; Son, J.W. Particulate matter less than 10 μm (PM10) activates cancer related genes in lung epithelial cells. Inhal. Toxicol. 2020, 32, 487–493. [Google Scholar] [CrossRef]
- Rider, C.F.; Carlsten, C. Air pollution and DNA methylation: Effects of exposure in humans. Clin. Epigenetics 2019, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Peng, K.-T.; Liu, J.F.; Chiang, Y.-C.; Chen, P.-C.; Chiang, M.-H.; Shih, H.-N.; Chang, P.-J.; Lee, C.-W. Particulate matter exposure aggravates osteoarthritis severity. Clin. Sci. 2019, 133, 2171–2187. [Google Scholar] [CrossRef] [Green Version]
- Duygulu, F.; Karaoğlu, S.; Zeybek, N.D.; Kaymaz, F.F.; Güneş, T. The effect of subcutaneously injected nicotine on achilles tendon healing in rabbits. Knee Surg. Sports Traumatol. Arthrosc. 2006, 14, 756–761. [Google Scholar] [CrossRef]
- Abate, M.; Vanni, D.; Pantalone, A.; Salini, V. Cigarette smoking and musculoskeletal disorders. Muscles Ligaments Tendons J. 2013, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Uquillas, C.A.; Guss, M.S.; Ryan, D.J.; Jazrawi, L.M.; Strauss, E.J. Everything Achilles: Knowledge update and current concepts in management: AAOS exhibit selection. JBJS 2015, 97, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Nho, S.J.; Yadav, H.; Shindle, M.K.; Macgillivray, J.D. Rotator cuff degeneration: Etiology and pathogenesis. Am. J. Sports Med. 2008, 36, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Ganestam, A.; Kallemose, T.; Troelsen, A.; Barfod, K.W. Increasing incidence of acute Achilles tendon rupture and a noticeable decline in surgical treatment from 1994 to 2013. A nationwide registry study of 33,160 patients. Knee Surg. Sports Traumatol. Arthrosc. 2016, 24, 3730–3737. [Google Scholar] [CrossRef]
- Fearon, A.; Dahlstrom, J.E.; Twin, J.; Cook, J.; Scott, A. The Bonar score revisited: Region of evaluation significantly influences the standardized assessment of tendon degeneration. J. Sci. Med. Sport 2014, 17, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Liu, J.; Sun, J.; Xu, X.; Hu, Y.; Liu, B. Optical microscopy and electron microscopy for the morphological evaluation of tendons: A mini review. Orthop. Surg. 2020, 12, 366–371. [Google Scholar] [CrossRef]
- Okamoto, N.; Kushida, T.; Oe, K.; Umeda, M.; Ikehara, S.; Iida, H. Treating Achilles tendon rupture in rats with bone-marrow-cell transplantation therapy. JBJS 2010, 92, 2776–2784. [Google Scholar] [CrossRef]
- Möller, M.; Movin, T.; Granhed, H.; Lind, K.; Faxen, E.; Karlsson, J. Acute rupture of tendo Achillis: A prospective, randomised study of comparison between surgical and non-surgical treatment. J. Bone Jt. Surg. 2001, 83, 843–848. [Google Scholar] [CrossRef]
- Müller, S.A.; Dürselen, L.; Heisterbach, P.; Evans, C.; Majewski, M. Effect of a simple collagen type I sponge for Achilles tendon repair in a rat model. Am. J. Sports Med. 2016, 44, 1998–2004. [Google Scholar] [CrossRef]
- Chisari, E.; Rehak, L.; Khan, W.S.; Maffulli, N. Tendon healing in presence of chronic low-level inflammation: A systematic review. Br. Med. Bull. 2019, 132, 97–116. [Google Scholar] [CrossRef]
- Chisari, E.; Rehak, L.; Khan, W.S.; Maffulli, N. The role of the immune system in tendon healing: A systematic review. Br. Med. Bull. 2020, 133, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Kang, C.; Han, S.Y.; Park, W.H.; Kim, M.H.; Moon, J.H.; Tae, J.Y.; Park, H.Y.; Yoo, I.H.; Park, J.H. Comparative analysis of Achilles tendon healing outcomes after open tenotomy versus percutaneous tenotomy: An experimental study in rats. J. Orthop. Res. 2021, 40, 1446–1456. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.-F.; Xu, Y.-H.; Shi, M.-H.; Lian, Y.-X. The impact of PM2. 5 on the human respiratory system. J. Thorac. Dis. 2016, 8, E69. [Google Scholar] [PubMed]
- Kim, C.; Jeong, S.H.; Kim, J.; Kang, J.Y.; Nam, Y.J.; Togloom, A.; Cha, J.; Lee, K.Y.; Lee, C.H.; Park, E.-K. Evaluation of the effect of filtered ultrafine particulate matter on bleomycin-induced lung fibrosis in a rat model using computed tomography, histopathologic analysis, and RNA sequencing. Sci. Rep. 2021, 11, 22672. [Google Scholar] [CrossRef]
- Cui, Y.; Xie, X.; Jia, F.; He, J.; Li, Z.; Fu, M.; Hao, H.; Liu, Y.; Liu, J.Z.; Cowan, P.J. Ambient fine particulate matter induces apoptosis of endothelial progenitor cells through reactive oxygen species formation. Cell. Physiol. Biochem. 2015, 35, 353–363. [Google Scholar] [CrossRef]
- Cook, J.; Feller, J.; Bonar, S.; Khan, K. Abnormal tenocyte morphology is more prevalent than collagen disruption in asymptomatic athletes’ patellar tendons. J. Orthop. Res. 2004, 22, 334–338. [Google Scholar] [CrossRef]
- Maffulli, N.; Barrass, V.; Ewen, S.W. Light microscopic histology of Achilles tendon ruptures: A comparison with unruptured tendons. Am. J. Sports Med. 2000, 28, 857–863. [Google Scholar] [CrossRef]
- Best, T.M.; Collins, A.; Lilly, E.G.; Seaber, A.V.; Goldner, R.; Murrell, G.A. Achilles tendon healing: A correlation between functional and mechanical performance in the rat. J. Orthop. Res. 1993, 11, 897–906. [Google Scholar] [CrossRef]
- Cappelli, E.; Felici, G.; Weitschek, E. Combining DNA methylation and RNA sequencing data of cancer for supervised knowledge extraction. BioData Min. 2018, 11, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xu, J.; Lazarovici, P.; Quirion, R.; Zheng, W. cAMP response element-binding protein (CREB): A possible signaling molecule link in the pathophysiology of schizophrenia. Front. Mol. Neurosci. 2018, 11, 255. [Google Scholar] [CrossRef]
- Hetland, R.B.; Cassee, F.R.; Låg, M.; Refsnes, M.; Dybing, E.; Schwarze, P.E. Cytokine release from alveolar macrophages exposed to ambient particulate matter: Heterogeneity in relation to size, city and season. Part. Fibre Toxicol. 2005, 2, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tintut, Y.; Patel, J.; Parhami, F.; Demer, L.L. Tumor necrosis factor-α promotes in vitro calcification of vascular cells via the cAMP pathway. Circulation 2000, 102, 2636–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Gan, Y.; Xu, Y.; Song, L.; Wang, L.; Ouyang, B.; Zhang, C.; Zhou, Q. The inflammatory cytokine TNF-α promotes the premature senescence of rat nucleus pulposus cells via the PI3K/Akt signaling pathway. Sci. Rep. 2017, 7, 42938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaida, J.E.; Bagge, J.; Purdam, C.; Cook, J.; Alfredson, H.; Forsgren, S. Evidence of the TNF-α system in the human Achilles tendon: Expression of TNF-α and TNF receptor at both protein and mRNA levels in the tenocytes. Cells Tissues Organs 2012, 196, 339–352. [Google Scholar] [CrossRef]
- Thomopoulos, S.; Parks, W.C.; Rifkin, D.B.; Derwin, K.A. Mechanisms of tendon injury and repair. J. Orthop. Res. 2015, 33, 832–839. [Google Scholar] [CrossRef] [Green Version]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The role of the transcription factor CREB in immune function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [Green Version]
- Mayr, B.; Montminy, M. Transcriptional regulation by the phosphorylation-dependent factor CREB. Nat. Rev. Mol. Cell Biol. 2001, 2, 599–609. [Google Scholar] [CrossRef]
- Morita, W.; Dakin, S.; Snelling, S.; Carr, A. Cytokines in tendon disease: A systematic review. Bone Jt. Res. 2017, 6, 656–664. [Google Scholar] [CrossRef]
- John, T.; Lodka, D.; Kohl, B.; Ertel, W.; Jammrath, J.; Conrad, C.; Stoll, C.; Busch, C.; Schulze-Tanzil, G. Effect of pro-inflammatory and immunoregulatory cytokines on human tenocytes. J. Orthop. Res. 2010, 28, 1071–1077. [Google Scholar] [CrossRef]
- Hernandez-Munoz, I.; De la Torre, P.; Sanchez-Alcazar, J.A.; Garcia, I.; Santiago, E.; Munoz-Yague, M.T.; Solis-Herruzo, J.A. Tumor necrosis factor alpha inhibits collagen alpha 1 (I) gene expression in rat hepatic stellate cells through a G protein. Gastroenterology 1997, 113, 625–640. [Google Scholar] [CrossRef]
- Chua, C.C.; Chua, B.H. Tumor necrosis factor-α induces mRNA for collagenase and TIMP in human skin fibroblasts. Connect. Tissue Res. 1990, 25, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Petiet, A.; Delatour, B.; Dhenain, M. Models of neurodegenerative disease–Alzheimer’s anatomical and amyloid plaque imaging. In Vivo NMR Imaging; Springer: Berlin/Heidelberg, Germany, 2011; pp. 293–308. [Google Scholar]
- Han, K.H.; Hyun, J.S.; Choi, W.K.; Lee, J.S. A study of co-combustion characteristics of North Korean anthracite and bituminous coal in 2 MWe CFBC power plant. Korean Chem. Eng. Res. 2009, 47, 580–586. [Google Scholar]
- Pasqua, L.A.; Damasceno, M.V.; Cruz, R.; Matsuda, M.; Garcia Martins, M.; Lima-Silva, A.E.; Marquezini, M.; Saldiva, P.H.N.; Bertuzzi, R. Exercising in air pollution: The cleanest versus dirtiest cities challenge. Int. J. Environ. Res. Public Health 2018, 15, 1502. [Google Scholar] [CrossRef] [Green Version]
- Whiting, C.H.; Jansen, K.E.; Dey, S. Hierarchical basis for stabilized finite element methods for compressible flows. Comput. Methods Appl. Mech. Eng. 2003, 192, 5167–5185. [Google Scholar] [CrossRef]
- Shakib, F.; Hughes, T.J.; Johan, Z. A new finite element formulation for computational fluid dynamics: X. The compressible Euler and Navier-Stokes equations. Comput. Methods Appl. Mech. Eng. 1991, 89, 141–219. [Google Scholar] [CrossRef]
- Hauke, G.; Hughes, T. A unified approach to compressible and incompressible flows. Comput. Methods Appl. Mech. Eng. 1994, 113, 389–395. [Google Scholar] [CrossRef]
- Jansen, K.E.; Whiting, C.H.; Hulbert, G.M. A generalized-α method for integrating the filtered Navier–Stokes equations with a stabilized finite element method. Comput. Methods Appl. Mech. Eng. 2000, 190, 305–319. [Google Scholar] [CrossRef]
- Johan, Z.; Hughes, T.J. A globally convergent matrix-free algorithm for implicit time-marching schemes arising in finite element analysis in fluids. Comput. Methods Appl. Mech. Eng. 1991, 87, 281–304. [Google Scholar] [CrossRef]
- Houghton, E.L.; Carpenter, P.W. Aerodynamics for Engineering Students; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Ran, J.; Hu, Y.; Le, H.; Chen, Y.; Zheng, Z.; Chen, X.; Yin, Z.; Yan, R.; Jin, Z.; Tang, C. Ectopic tissue engineered ligament with silk collagen scaffold for ACL regeneration: A preliminary study. Acta Biomater. 2017, 53, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, M.; Kitaura, T.; Ikoma, K.; Kusaka, Y. A method of in-vitro measurement of the cross-sectional area of soft tissues, using ultrasonography. J. Orthop. Sci. 2002, 7, 247–251. [Google Scholar] [CrossRef]
- Heitman, D.E.; Ng, K.; Crivello, K.M.; Gallina, J. Biomechanical Comparison of the Achillon® Tendon Repair System and the Krackow Locking Loop Technique. Foot Ankle Int. 2011, 32, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Roßbach, B.P.; Gülecyüz, M.F.; Kempfert, L.; Pietschmann, M.F.; Ullamann, T.; Ficklscherer, A.; Niethammer, T.R.; Zhang, A.; Klar, R.M.; Müller, P.E. Rotator cuff repair with autologous tenocytes and biodegradable collagen scaffold: A histological and biomechanical study in sheep. Am. J. Sports Med. 2020, 48, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Caraux, G.; Pinloche, S. PermutMatrix: A graphical environment to arrange gene expression profiles in optimal linear order. Bioinformatics 2005, 21, 1280–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Lee, S.E.; Yi, S.; Jun, S.; Yi, Y.-S.; Nagar, H.; Kim, C.-S.; Shin, C.; Yeo, M.-K.; Kang, Y.E. Tauroursodeoxycholic Acid Decreases Keloid Formation by Reducing Endoplasmic Reticulum Stress as Implicated in the Pathogenesis of Keloid. Int. J. Mol. Sci. 2021, 22, 10765. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Ahn, J.; Kim, S.A.; Go, E.J.; Lee, D.H.; Park, S.C.; Shetty, A.A.; Kim, S.J. Improved Healing of Rabbit Patellar Tendon Defects After an Atelocollagen Injection. Am. J. Sports Med. 2021, 49, 2924–2932. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-Y.; Lee, M.-H.; Jo, S.-K.; Yoo, I.-H.; Sarankhuu, B.-E.; Kim, H.-J.; Kang, Y.-E.; Lee, S.-E.; Kim, T.-Y.; Park, M.-H.; et al. Effects and Mechanism of Particulate Matter on Tendon Healing Based on Integrated Analysis of DNA Methylation and RNA Sequencing Data in a Rat Model. Int. J. Mol. Sci. 2022, 23, 8170. https://doi.org/10.3390/ijms23158170
Lee S-Y, Lee M-H, Jo S-K, Yoo I-H, Sarankhuu B-E, Kim H-J, Kang Y-E, Lee S-E, Kim T-Y, Park M-H, et al. Effects and Mechanism of Particulate Matter on Tendon Healing Based on Integrated Analysis of DNA Methylation and RNA Sequencing Data in a Rat Model. International Journal of Molecular Sciences. 2022; 23(15):8170. https://doi.org/10.3390/ijms23158170
Chicago/Turabian StyleLee, Su-Yel, Min-Hyeok Lee, Seong-Kyeong Jo, In-Ha Yoo, Boler-Erdene Sarankhuu, Hyun-Jin Kim, Yea-Eun Kang, Seong-Eun Lee, Tae-Yeon Kim, Moon-Hyang Park, and et al. 2022. "Effects and Mechanism of Particulate Matter on Tendon Healing Based on Integrated Analysis of DNA Methylation and RNA Sequencing Data in a Rat Model" International Journal of Molecular Sciences 23, no. 15: 8170. https://doi.org/10.3390/ijms23158170