
Improving the Therapeutic Index of Smp24, a Venom-Derived Antimicrobial Peptide: Increased Activity against Gram-Negative Bacteria


Abstract
:1. Introduction
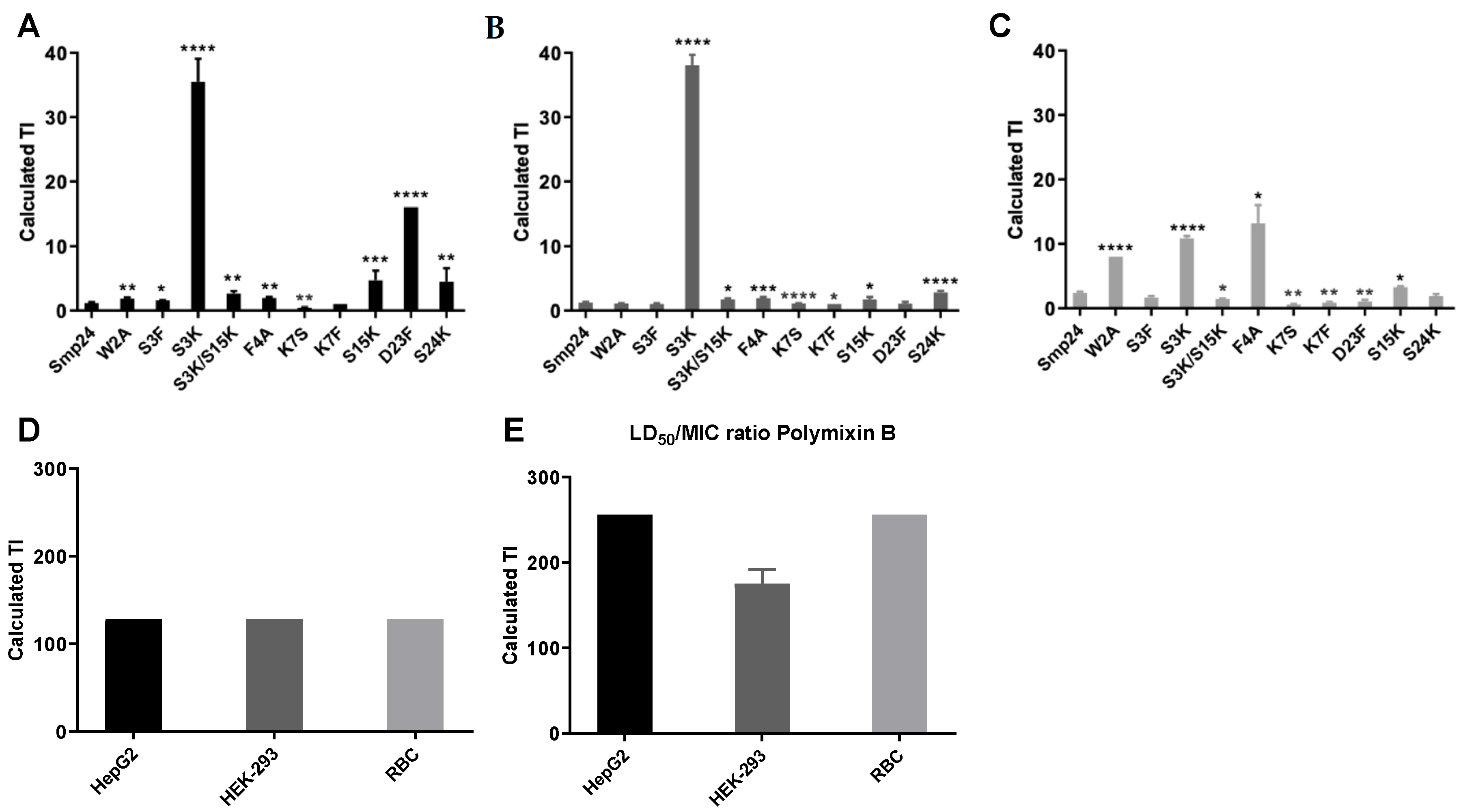
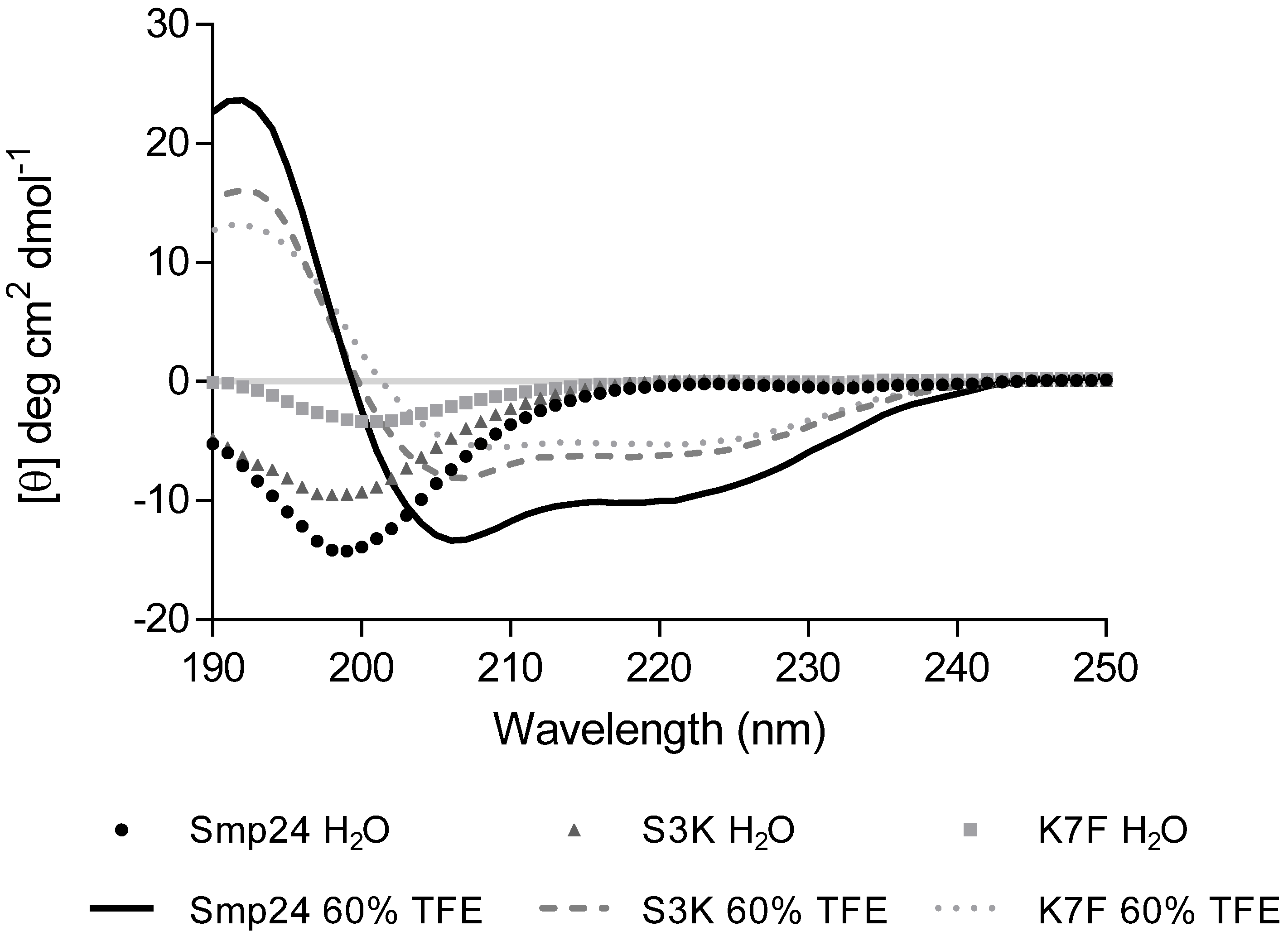
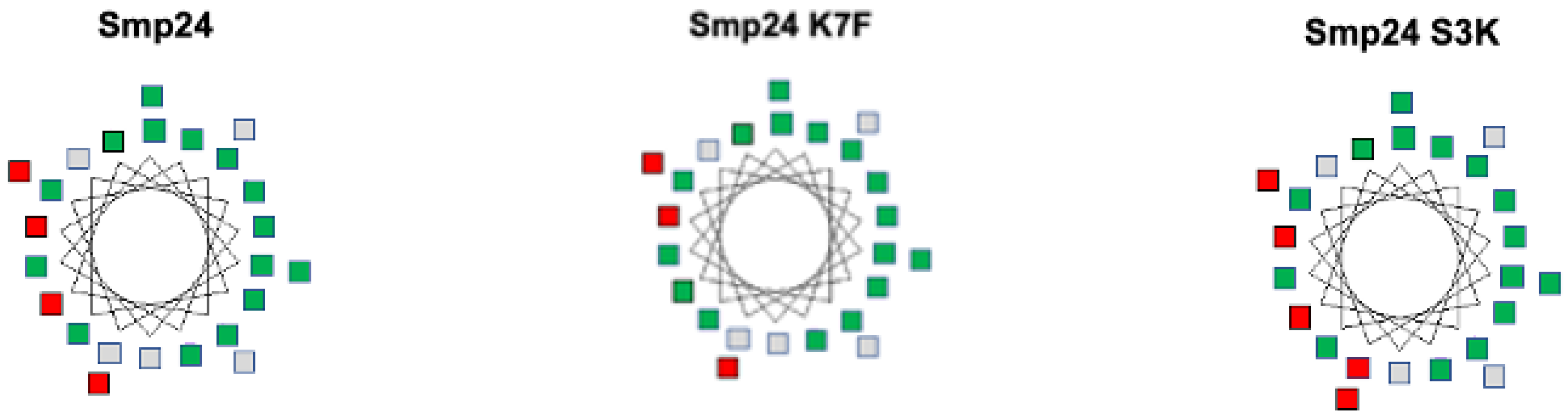
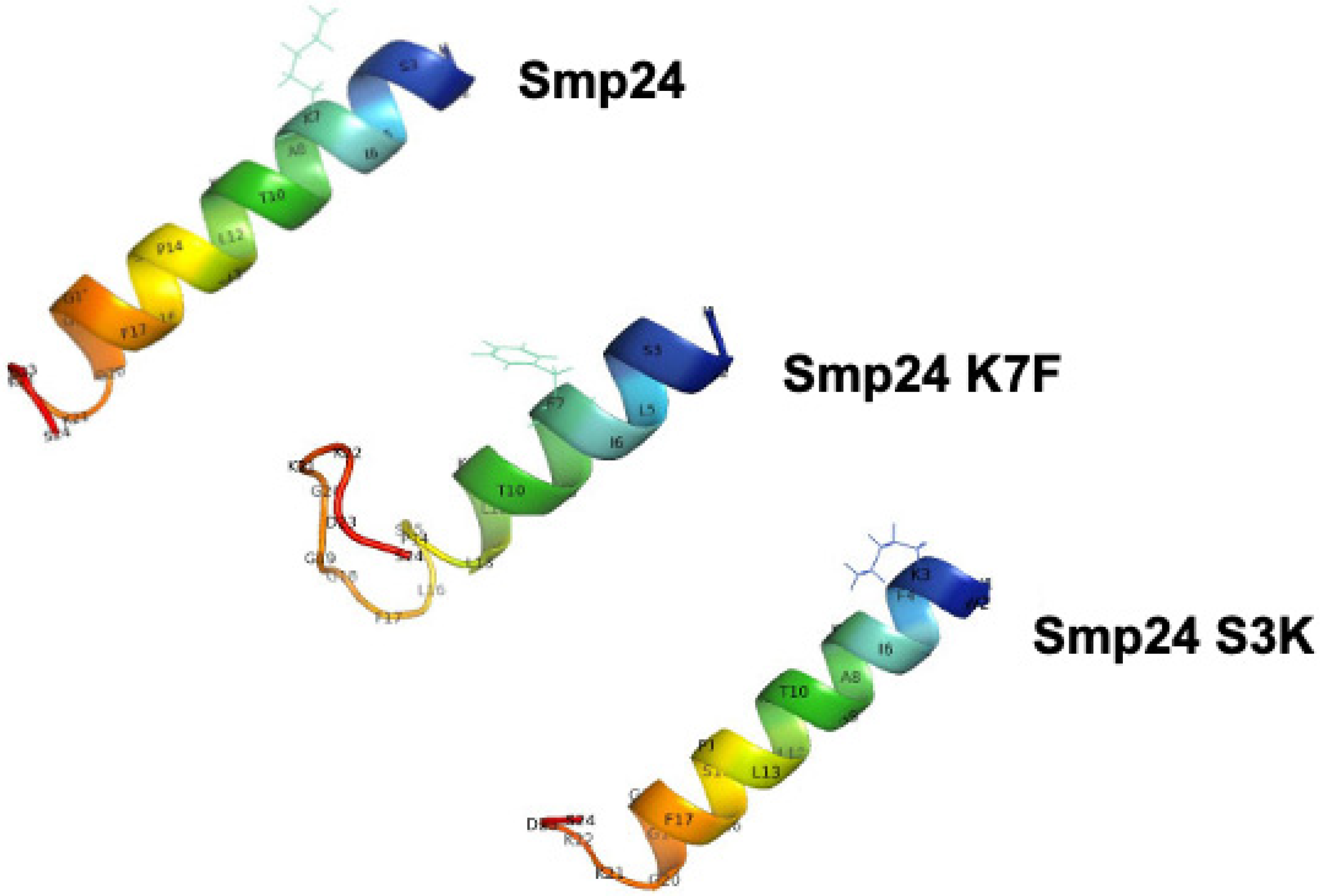
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Bacterial Strains
4.3. Antimicrobial Assays
4.4. Haemolysis Assay
4.5. Cytotoxicity Assays
4.6. Therapeutic Indices
4.7. Circular Dichroism (CD) Spectra
4.8. Modelling Studies
4.8.1. Helical Wheel Projections
4.8.2. Ab-Initio Modelling
4.8.3. 3D Modelling
4.8.4. Hydrophobic Moment
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez, S.; Demain, A.L. The Amazing World of Antibiotics. Biochem. Pharmacol. 2017, 133, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Schäberle, T.F.; Hack, I.M. Overcoming the Current Deadlock in Antibiotic Research. Trends Microbiol. 2014, 22, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Santos-Beneit, F.; Ordóñez-Robles, M.; Martín, J.F. Glycopeptide Resistance: Links with Inorganic Phosphate Metabolism and Cell Envelope Stress. Biochem. Pharmacol. 2017, 133, 74–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, A.; Kundu, P.; Bhunia, A. Designing Potent Antimicrobial Peptides by Disulphide Linked Dimerization and N-Terminal Lipidation to Increase Antimicrobial Activity and Membrane Perturbation: Structural Insights into Lipopolysaccharide Binding. J. Colloid Interface Sci. 2015, 461, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Dobson, A.; Cotter, P.D.; Paul Ross, R.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Singh, B.; van Belkum, M.J.; Diep, D.B.; Chikindas, M.L.; Ermakov, A.M.; Tiwari, S.K. Halocins, Natural Antimicrobials of Archaea: Exotic or Special or Both? Biotechnol. Adv. 2021, 53, 107834. [Google Scholar] [CrossRef]
- Leippe, M. Antimicrobial and Cytolytic Polypeptides of Amoeboid Protozoa—Effector Molecules of Primitive Phagocytes. Dev. Comp. Immunol. 1999, 23, 267–279. [Google Scholar] [CrossRef]
- Lübeck, M.; Lübeck, P.S. Fungal Cell Factories for Efficient and Sustainable Production of Proteins and Peptides. Microorganisms 2022, 10, 753. [Google Scholar] [CrossRef]
- Landi, N.; Clemente, A.; Pedone, P.V.; Ragucci, S.; Di Maro, A. An Updated Review of Bioactive Peptides from Mushrooms in a Well-Defined Molecular Weight Range. Toxins 2022, 14, 84. [Google Scholar] [CrossRef]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant Antimicrobial Peptides: Structures, Functions, and Applications. Bot. Stud. 2021, 62, 5. [Google Scholar] [CrossRef] [PubMed]
- De Smet, K.; Contreras, R. Human Antimicrobial Peptides: Defensins, Cathelicidins and Histatins. Biotechnol. Lett. 2005, 27, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.L.; Abdel-Rahman, M.A.; Miller, K.; Strong, P.N. Antimicrobial Peptides from Scorpion Venoms. Toxicon 2014, 88, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, P.; Niu, Z.-W. A Perspective on General Direction and Challenges Facing Antimicrobial Peptides. Chin. Chem. Lett. 2017, 28, 703–708. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide Antimicrobial Agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.; Alayande, A.B.; Chae, S.; Kim, I.S. Applications of Nisin for Biofouling Mitigation of Reverse Osmosis Membranes. Desalination 2017, 429, 52–59. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Quintero-Hernandez, V.; Possani, L.D. Venom Proteomic and Venomous Glands Transcriptomic Analysis of the Egyptian Scorpion Scorpio Maurus Palmatus (Arachnida: Scorpionidae). Toxicon 2013, 74, 193–207. [Google Scholar] [CrossRef]
- Harrison, P.L.; Abdel-Rahman, M.A.; Strong, P.N.; Tawfik, M.M.; Miller, K. Characterisation of Three Alpha-Helical Antimicrobial Peptides from the Venom of Scorpio Maurus Palmatus. Toxicon 2016, 117, 30–36. [Google Scholar] [CrossRef]
- Heath, G.R.; Harrison, P.L.; Strong, P.N.; Evans, S.D.; Miller, K. Visualization of Diffusion Limited Antimicrobial Peptide Attack on Supported Lipid Membranes. Soft Matter 2018, 14, 6146–6154. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Gao, C.; Zhao, Q.; Li, C. Antimicrobial Characterization of Site-Directed Mutagenesis of Porcine Beta Defensin 2. PLoS ONE 2015, 10, e0118170. [Google Scholar] [CrossRef]
- Bobone, S.; Roversi, D.; Giordano, L.; De Zotti, M.; Formaggio, F.; Toniolo, C.; Park, Y.; Stella, L. The Lipid Dependence of Antimicrobial Peptide Activity Is an Unreliable Experimental Test for Different Pore Models. Biochemistry 2012, 51, 10124–10126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, J.M. Determination of Minimum Inhibitory Concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corzo, G.; Escoubas, P.; Villegas, E.; Barnham, K.J.; He, W.; Norton, R.S.; Nakajima, T. Characterization of Unique Amphipathic Antimicrobial Peptides from Venom of the Scorpion Pandinus Imperator. Biochem. J. 2001, 359, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Al-Qarawi, A.A.; Mousa, H.M. Lipid Concentrations in Erythrocyte Membranes in Normal, Starved, Dehyrated and Rehydrated Camels (Camelus dromedarius), and in Normal Sheep (Ovis aries) and Goats (Capra hircus) ARTICLE IN PRESS. J. Arid Environ. 2004, 59, 675–683. [Google Scholar] [CrossRef]
- Jiang, Z.; Vasil, A.I.; Vasil, M.L.; Hodges, R.S. “Specificity Determinants” Improve Therapeutic Indices of Two Antimicrobial Peptides Piscidin 1 and Dermaseptin S4 Against the Gram-Negative Pathogens Acinetobacter Baumannii and Pseudomonas Aeruginosa. Pharmaceuticals 2014, 7, 366–391. [Google Scholar] [CrossRef] [Green Version]
- Bacalum, M.; Radu, M. Cationic Antimicrobial Peptides Cytotoxicity on Mammalian Cells: An Analysis Using Therapeutic Index Integrative Concept. Int. J. Pept. Res. Ther. 2015, 21, 47–55. [Google Scholar] [CrossRef]
- Li, C.; Arakawa, T. Feasibility of Circular Dichroism to Study Protein Structure at Extreme Concentrations. Int. J. Biol. Macromol. 2019, 132, 1290–1295. [Google Scholar] [CrossRef]
- Reißer, S.; Strandberg, E.; Steinbrecher, T.; Ulrich, A.S. 3D Hydrophobic Moment Vectors as a Tool to Characterize the Surface Polarity of Amphiphilic Peptides. Biophys. J. 2014, 106, 2385–2394. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.L. Antimicrobial Peptides Stage a Comeback. Nat. Biotechnol. 2013, 31, 379–382. [Google Scholar] [CrossRef]
- Ciumac, D.; Gong, H.; Hu, X.; Ren Lu, J. Membrane Targeting Cationic Antimicrobial Peptides. J. Colloid Interface Sci. 2019, 537, 163–185. [Google Scholar] [CrossRef]
- Tossi, A.; Sandri, L.; Giangaspero, A. Amphipathic, α-Helical Antimicrobial Peptides. Biopolymers 2000, 55, 4–30. [Google Scholar] [CrossRef]
- Gleason, N.J.; Vostrikov, V.V.; Greathouse, D.V.; Koeppe, R.E. Buried Lysine, but Not Arginine, Titrates and Alters Transmembrane Helix Tilt. Proc. Natl. Acad. Sci. USA 2013, 110, 1692–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Jia, Z.; Peng, Y.; Chakravorty, A.; Sun, L.; Alexov, E. DelPhiForce Web Server: Electrostatic Forces and Energy Calculations and Visualization. Bioinformatics 2017, 33, 3661–3663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringstad, L.; Schmidtchen, A.; Malmsten, M. Effects of Single Amino Acid Substitutions on Peptide Interaction with Lipid Membranes and Bacteria–Variants of GKE21, an Internal Sequence from Human LL-37. Colloids Surfaces A Physicochem. Eng. Asp. 2010, 354, 65–71. [Google Scholar] [CrossRef]
- Rodríguez, A.; Villegas, E.; Montoya-Rosales, A.; Rivas-Santiago, B.; Corzo, G. Characterization of Antibacterial and Hemolytic Activity of Synthetic Pandinin 2 Variants and Their Inhibition against Mycobacterium Tuberculosis. PLoS ONE 2014, 9, e101742. [Google Scholar] [CrossRef]
- Irazazabal, L.N.; Porto, W.F.; Ribeiro, S.M.; Casale, S.; Humblot, V.; Ladram, A.; Franco, O.L. Selective Amino Acid Substitution Reduces Cytotoxicity of the Antimicrobial Peptide Mastoparan. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 2699–2708. [Google Scholar] [CrossRef]
- Rodríguez, A.; Villegas, E.; Satake, H.; Possani, L.D.; Corzo, G. Amino Acid Substitutions in an Alpha-Helical Antimicrobial Arachnid Peptide Affect Its Chemical Properties and Biological Activity towards Pathogenic Bacteria but Improves Its Therapeutic Index. Amino Acids 2011, 40, 61–68. [Google Scholar] [CrossRef]
- Greco, I.; Molchanova, N.; Holmedal, E.; Jenssen, H.; Hummel, B.D.; Watts, J.L.; Håkansson, J.; Hansen, P.R.; Svenson, J. Correlation between Hemolytic Activity, Cytotoxicity and Systemic in Vivo Toxicity of Synthetic Antimicrobial Peptides. Sci. Rep. 2020, 10, 13206. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial Membrane Lipids: Diversity in Structures and Pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Tam, K. Biomembrane Mimics and Their Roles in Anti-Bacterial Drug Discovery. ADMET DMPK 2017, 5, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Bot, C.T.; Prodan, C. Quantifying the Membrane Potential during E. Coli Growth Stages. Biophys. Chem. 2010, 146, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Hayami, M.; Okabe, A.; Kariyama, R.; Abe, M.; Kanemasa, Y. Lipid Composition of Staphylococcus Aureus and Its Derived L-Forms. Microbiol. Immunol. 1979, 23, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Mates, S.M.; Patel, L.; Kaback, H.R.; Mandel, L.J.; Milazzo, J.J.; Edberg, S.C.; Miller, M.H. Membrane Potential and Gentamicin Uptake in Staphylococcus Aureus. Proc. Natl. Acad. Sci. USA 1982, 79, 6693–6697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, S.A.; Desbois, A.P.; Coote, P.J.; Smith, T.K. Characterisation of Staphylococcus Aureus Lipids by Nanoelectrospray Ionisation Tandem Mass Spectrometry (NESI-MS/MS). bioRxiv 2019, 593483. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Pomorski, T.; Porwoli, S.; Tauber, R.; Herrmann, A. Transverse Movement of Spin-Labeled Phospholipids in the Plasma Membrane of a Hepatocytic Cell Line (HepG2): Implications for Biliary Lipid Secretion. Hepatology 1996, 24, 1497–1503. [Google Scholar] [CrossRef]
- Edwards, I.A.; Elliott, A.G.; Kavanagh, A.M.; Zuegg, J.; Blaskovich, M.A.T.; Cooper, M.A. Contribution of Amphipathicity and Hydrophobicity to the Antimicrobial Activity and Cytotoxicity of β-Hairpin Peptides. ACS Infect. Dis. 2016, 2, 442–450. [Google Scholar] [CrossRef]
- Forbes, S.; Latimer, J.; Bazaid, A.; Mcbain, A.J. Altered Competitive Fitness, Antimicrobial Susceptibility, and Cellular Morphology in a Triclosan-Induced Small-Colony Variant of Staphylococcus Aureus. Antimicrob. Agents Chemother. 2015, 59, 4809–4816. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, J.A.; Cheng, K.H.; Somerharju, P. Phospholipid Composition of the Mammalian Red Cell Membrane Can Be Rationalized by a Superlattice Model. Biophysics 1998, 95, 4964–4969. [Google Scholar] [CrossRef] [Green Version]
- Cole, C.; Barber, J.D.; Barton, G.J. The Jpred 3 Secondary Structure Prediction Server. Nucleic Acids Res. 2008, 36, W197–W201. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence |
|---|---|
| Smp24 | IWSFLIKAATKLLPSLFGGGKKDS |
| Smp24 W2A | IASFLIKAATKLLPSLFGGGKKDS |
| Smp24 S3F | IWFFLIKAATKLLPSLFGGGKKDS |
| Smp24 S3K | IWKFLIKAATKLLPSLFGGGKKDS |
| Smp24 S3K/S15K | IWKFLIKAATKLLPKLFGGGKKDS |
| Smp24 F4A | IWSALIKAATKLLPSLFGGGKKDS |
| Smp24 K7S | IWSFLISAATKLLPSLFGGGKKDS |
| Smp24 K7F | IWSFLIFAATKLLPSLFGGGKKDS |
| Smp24 S15K | IWSFLIKAATKLLPKLFGGGKKDS |
| Smp24 D23F | IWSFLIKAATKLLPSLFGGGKKFS |
| Smp24 S24K | IWSFLIKAATKLLPSLFGGGKKDK |
| Daptomycin | WDNDTGODDADGD |
| Polymyxin B | DabT(Dab)3FL(Dab)2T |
| Peptide | Charge | GRAVY | MIC (mg/L) | MBC (mg/L) | ||||
|---|---|---|---|---|---|---|---|---|
| E. coli | P. aeruginosa | S. aureus | E. coli | P. aeruginosa | S. aureus | |||
| Smp24 | +3 | 0.31 | 32 | 64 | 8 | 32 | 64 | 8 |
| Smp24 W2A | +3 | 0.43 | 64 | 128 | 32 | 64 | 256 | 64 |
| Smp24 S3F | +3 | 0.46 | 32 | 64 | 16 | 64 | 128 | 64 |
| Smp24 S3K | +4 | 0.18 | 1 | 32 | 0.5 | 8 | 128 | 1 |
| Smp24 S3K/S15K | +5 | 0.05 | 16 | 16 | 16 | 16 | 32 | 16 |
| Smp24 F4A | +3 | 0.27 | 32 | 128 | 16 | 64 | 256 | 16 |
| Smp24 K7S | +2 | 0.44 | 64 | 128 | 16 | 128 | 256 | 32 |
| Smp24 K7F | +2 | 0.59 | >512 | >512 | >512 | >512 | >512 | >512 |
| Smp24 S15K | +4 | 0.18 | 8 | 32 | 4 | 8 | 32 | 8 |
| Smp24 D23F | +4 | 0.58 | 32 | 64 | 32 | 32 | 64 | 32 |
| Smp24 S24K | +4 | 0.18 | 8 | 32 | 4 | 8 | 128 | 8 |
| Daptomycin | −6 | −1.93 | >512 | >512 | 4 | >512 | >512 | 8 |
| Polymyxin B | +5 | nc | 2 | 2 | 32 | 4 | 8 | 64 |
| Peptide | Charge | GRAVY | HC50 (μg/mL) |
|---|---|---|---|
| Smp24 | +3 | 0.31 | 76 |
| Smp24 W2A | +3 | 0.43 | >512 * |
| Smp24 S3F | +3 | 0.46 | 52 |
| Smp24 S3K | +4 | 0.18 | 11 * |
| Smp24 S3K/S15K | +5 | 0.05 | 23 * |
| Smp24 F4A | +3 | 0.27 | 423 * |
| Smp24 K7S | +2 | 0.44 | 36 |
| Smp24 K7F | +2 | 0.59 | 442 * |
| Smp24 S15K | +4 | 0.18 | 26 * |
| Smp24 D23F | +4 | 0.58 | 33 * |
| Smp24 S24K | +4 | 0.18 | 15 * |
| Daptomycin | −6 | −1.93 | >512 |
| Polymyxin B | +5 | nc | >512 |
| Peptide | Charge | GRAVY | LD50 (μg/mL) | |
|---|---|---|---|---|
| HepG2 | HEK293 | |||
| Smp24 | +3 | 0.31 | 37 | 39 |
| Smp24 W2A | +3 | 0.43 | 121 * | 72 * |
| Smp24 S3F | +3 | 0.46 | 51 * | 32 |
| Smp24 S3K | +4 | 0.18 | 34 | 38 |
| Smp24 S3K/S15K | +5 | 0.05 | 42 | 29 * |
| Smp24 F4A | +3 | 0.27 | 63 * | 56 * |
| Smp24 K7S | +2 | 0.44 | 30 | 15 * |
| Smp24 K7F | +2 | 0.59 | >512 * | >512 * |
| Smp24 S15K | +4 | 0.18 | 38 | 13 * |
| Smp24 D23F | +4 | 0.58 | >512 * | 38 |
| Smp24 S24K | +4 | 0.18 | 36 | 23 * |
| Daptomycin | −6 | −1.93 | >512 | >512 |
| Polymyxin B | +5 | nc | >512 | 351 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rawson, K.M.; Lacey, M.M.; Strong, P.N.; Miller, K. Improving the Therapeutic Index of Smp24, a Venom-Derived Antimicrobial Peptide: Increased Activity against Gram-Negative Bacteria. Int. J. Mol. Sci. 2022, 23, 7979. https://doi.org/10.3390/ijms23147979
Rawson KM, Lacey MM, Strong PN, Miller K. Improving the Therapeutic Index of Smp24, a Venom-Derived Antimicrobial Peptide: Increased Activity against Gram-Negative Bacteria. International Journal of Molecular Sciences. 2022; 23(14):7979. https://doi.org/10.3390/ijms23147979
Chicago/Turabian StyleRawson, Kirstie M., Melissa M. Lacey, Peter N. Strong, and Keith Miller. 2022. "Improving the Therapeutic Index of Smp24, a Venom-Derived Antimicrobial Peptide: Increased Activity against Gram-Negative Bacteria" International Journal of Molecular Sciences 23, no. 14: 7979. https://doi.org/10.3390/ijms23147979