
3-keto-DON, but Not 3-epi-DON, Retains the in Planta Toxicological Potential after the Enzymatic Biotransformation of Deoxynivalenol


Abstract
:1. Introduction
2. Results
2.1. Lemna minor Studies
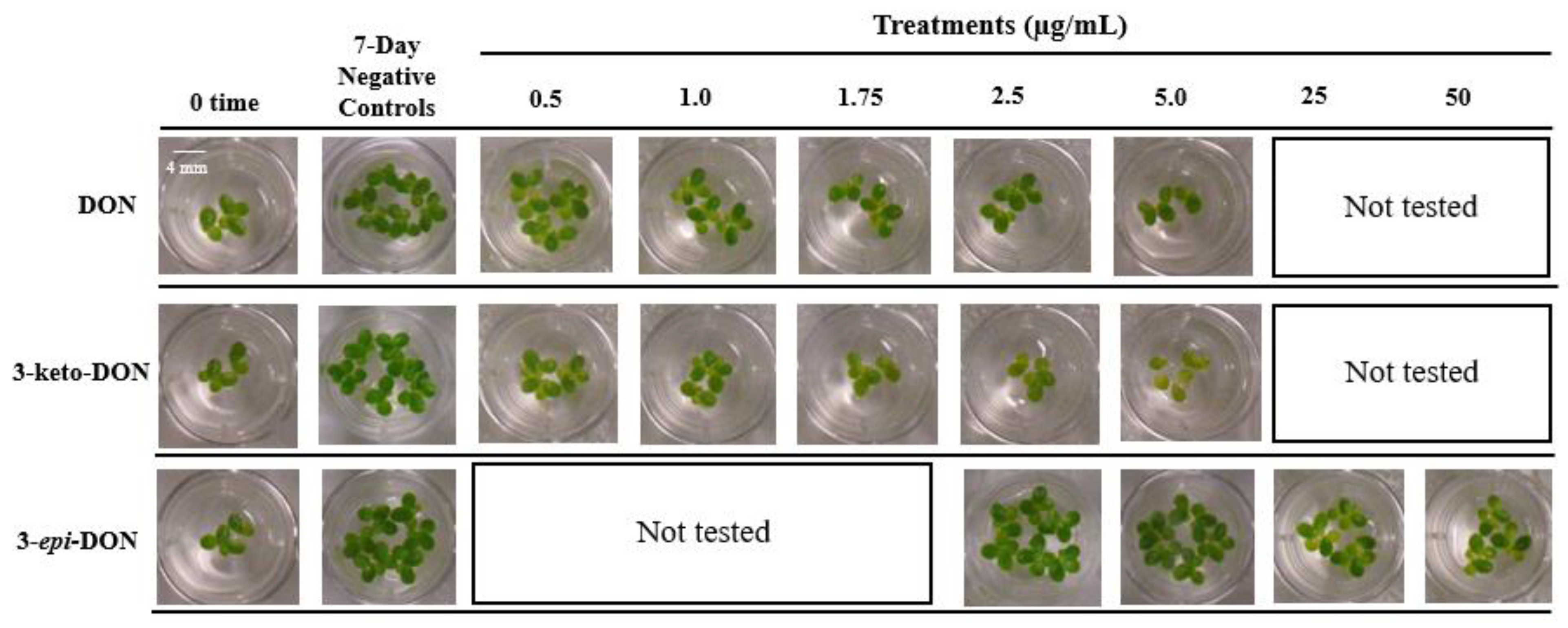
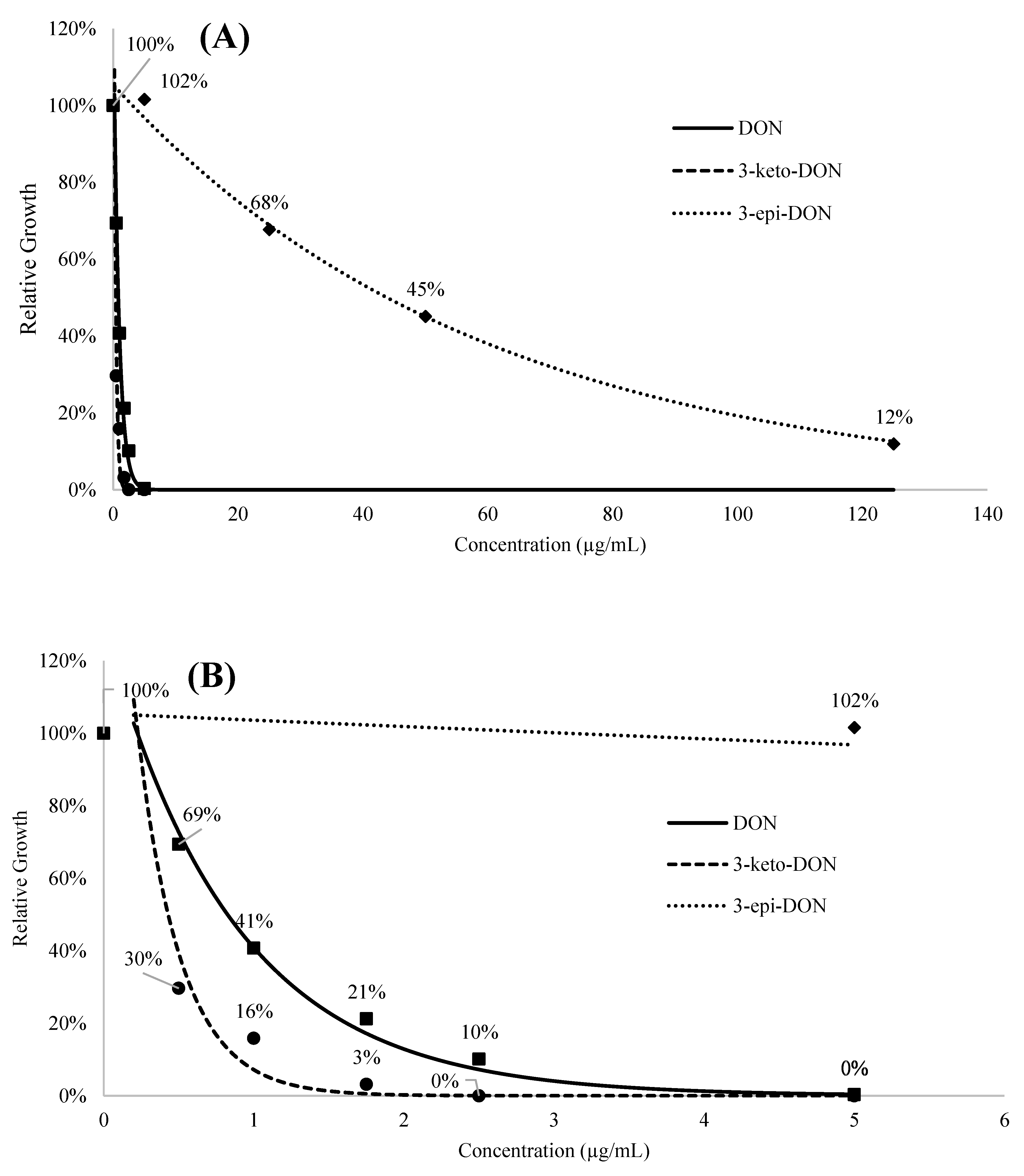
2.1.1. The Lemna minor Bioassay Is a Valid and Sensitive System to Assess DON Toxicity
2.1.2. 3-keto-DON Exhibits in Planta Toxicological Effects against L. minor Comparable to DON
2.1.3. High Concentrations of 3-epi-DON Do Not Negatively Affect L. minor Growth
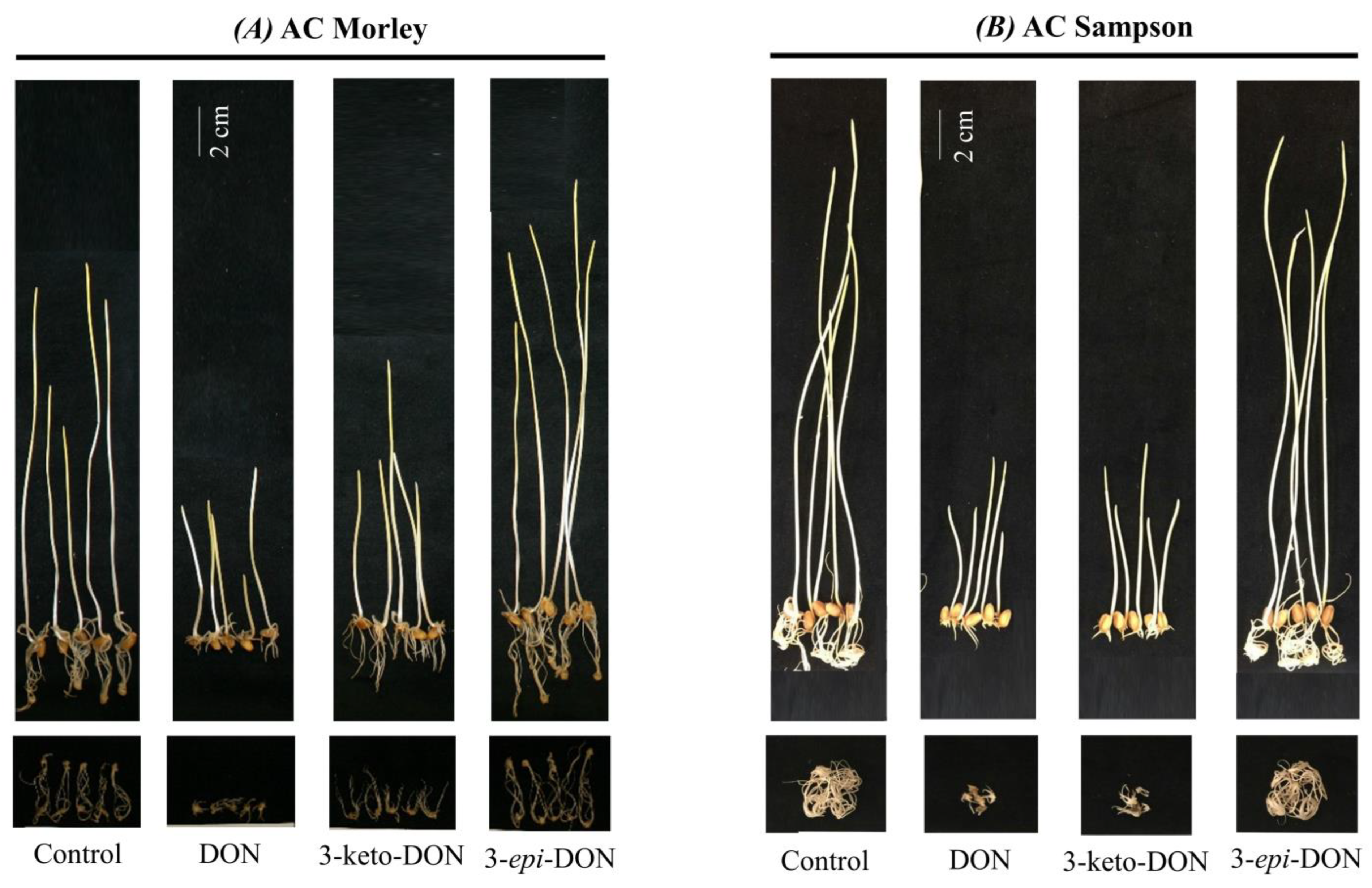
2.2. Phytotoxic Effects of DON and Its Metabolites on Wheat Seedlings Are Similar to Those Observed with L. minor
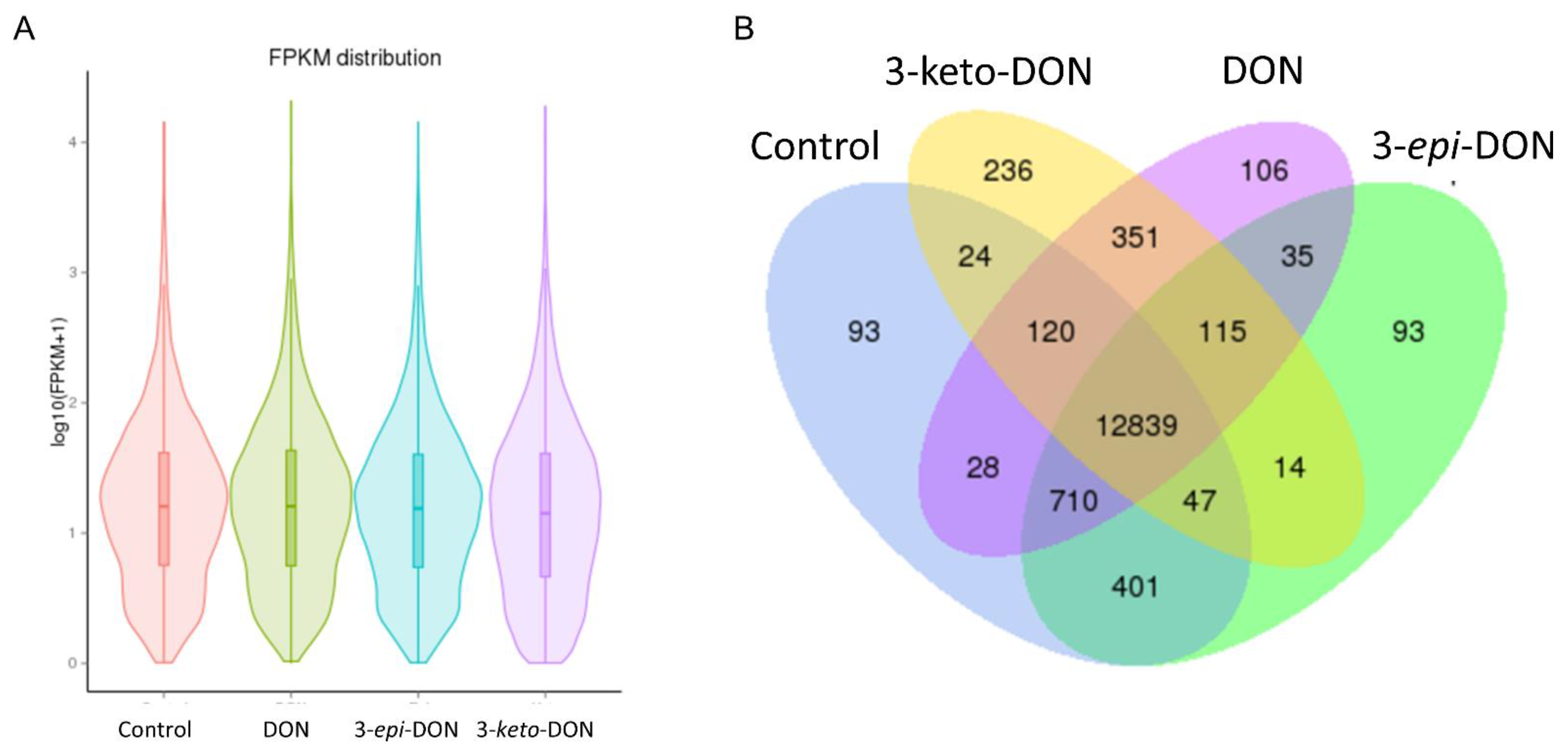
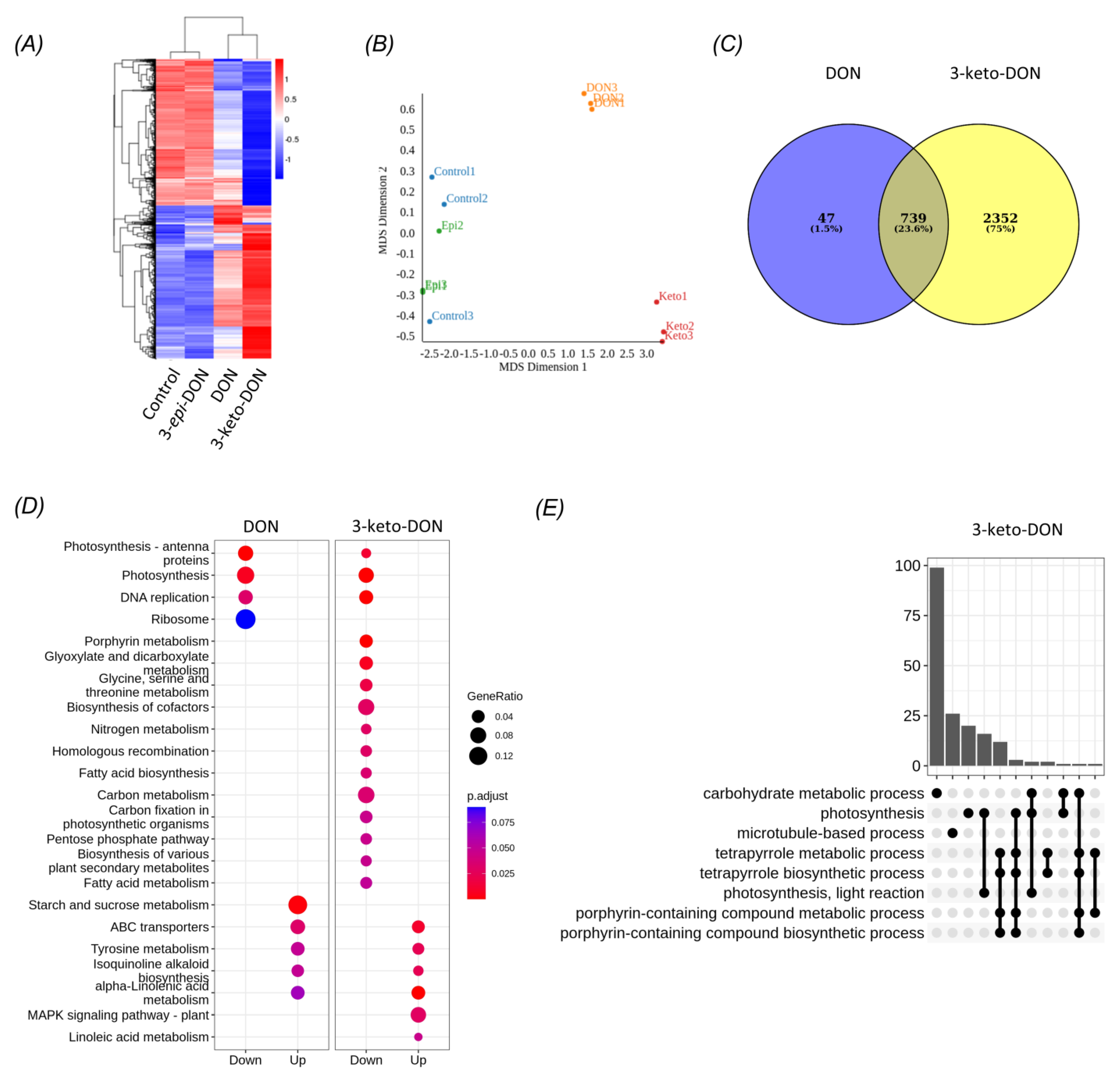
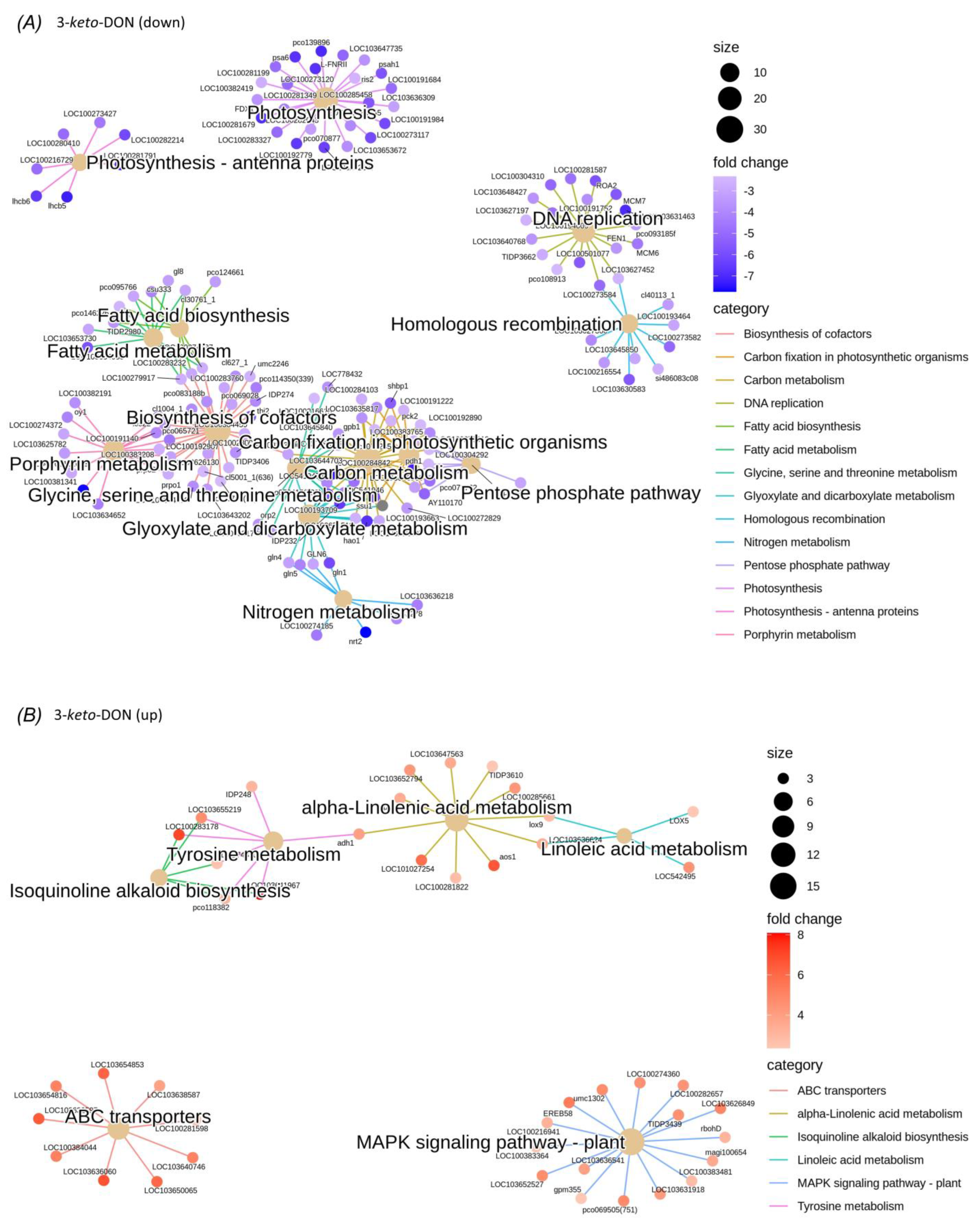
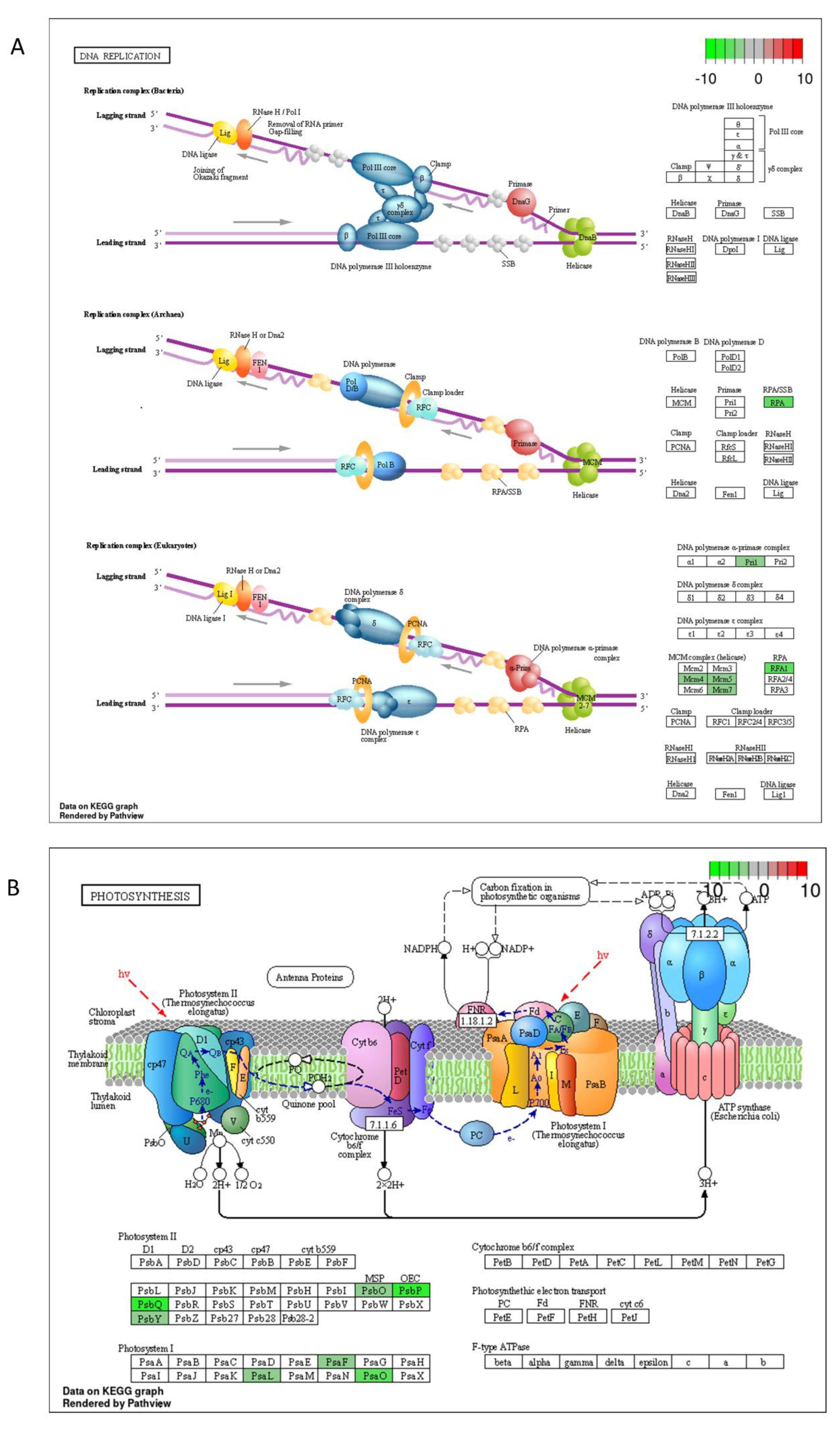
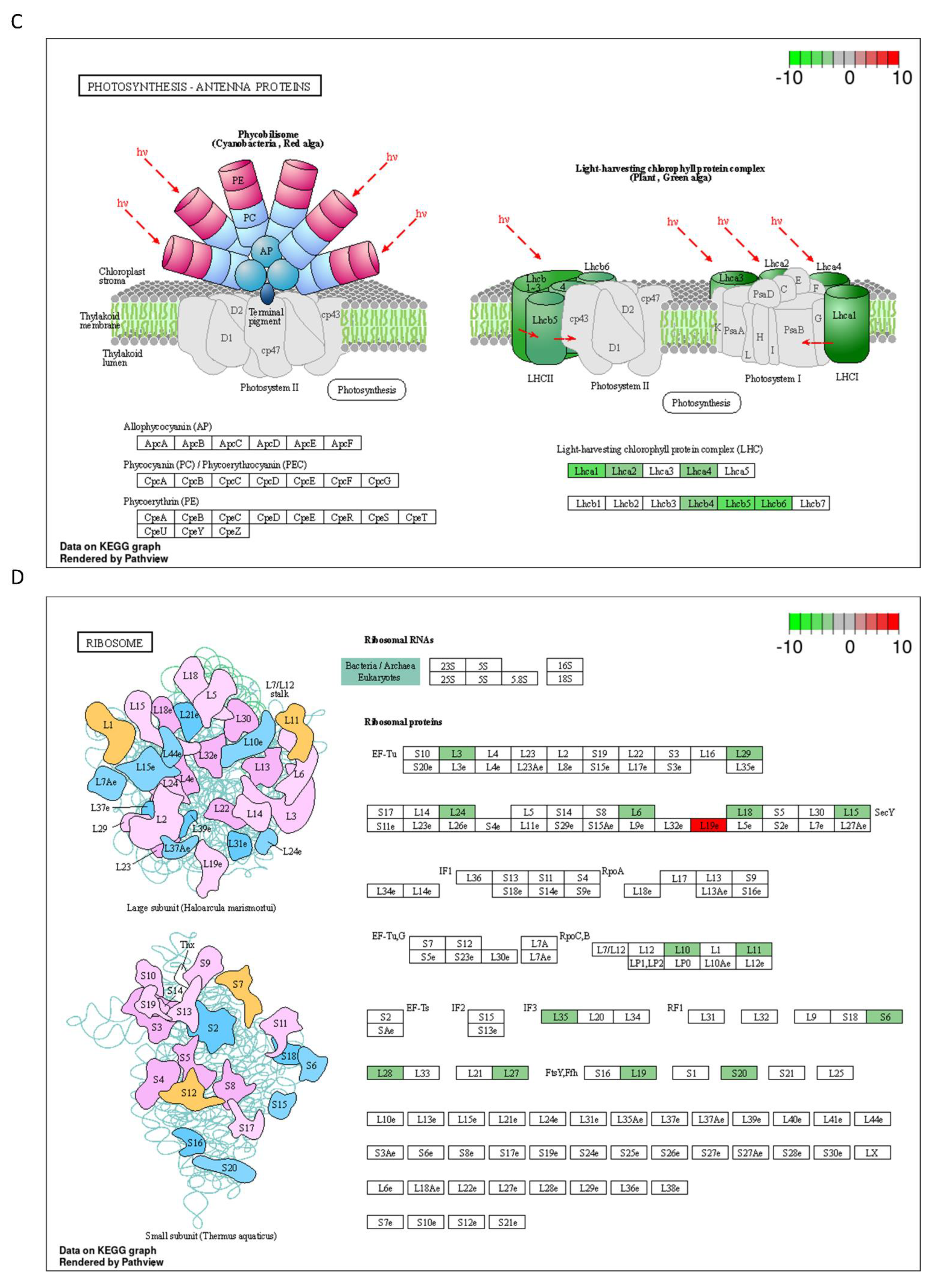
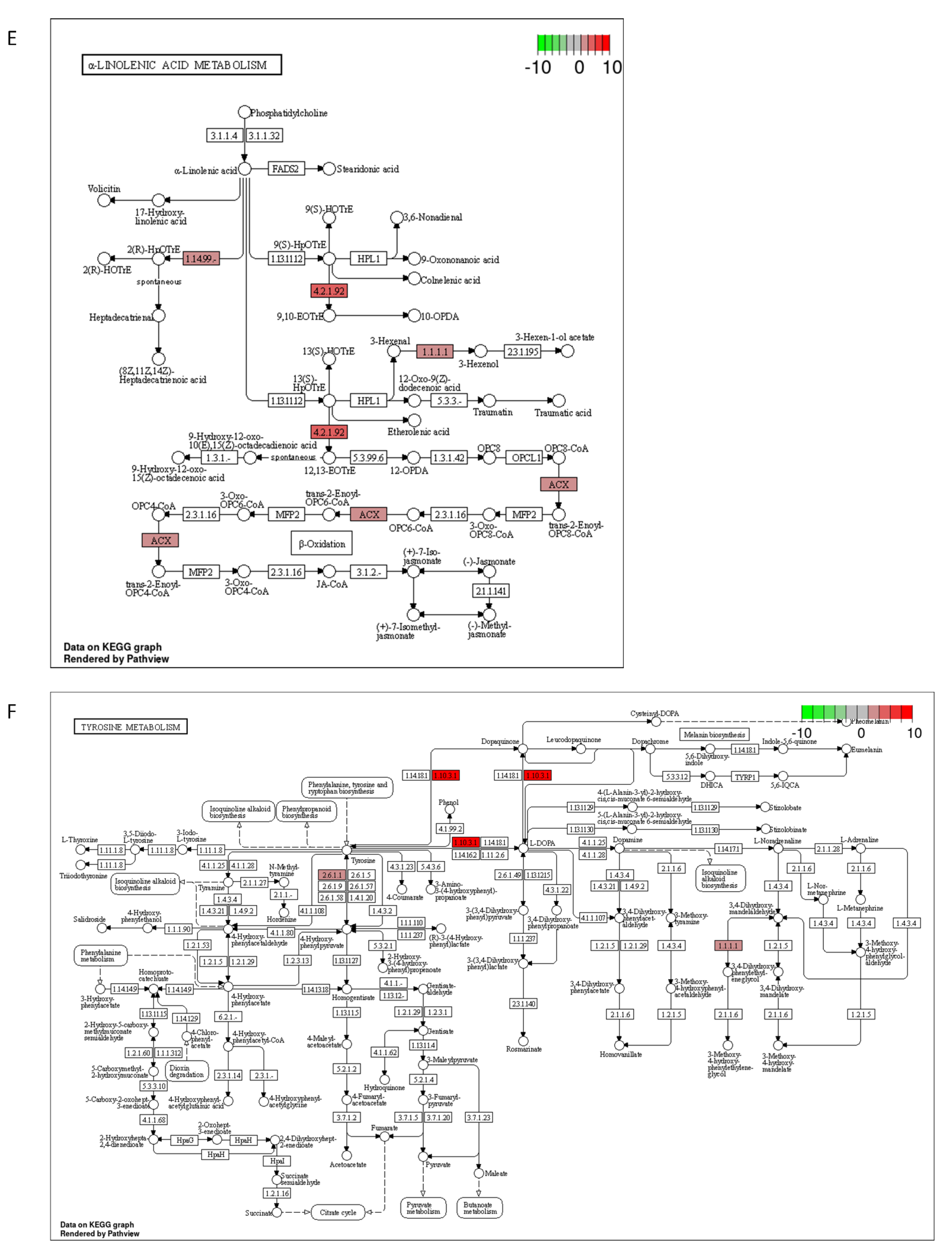
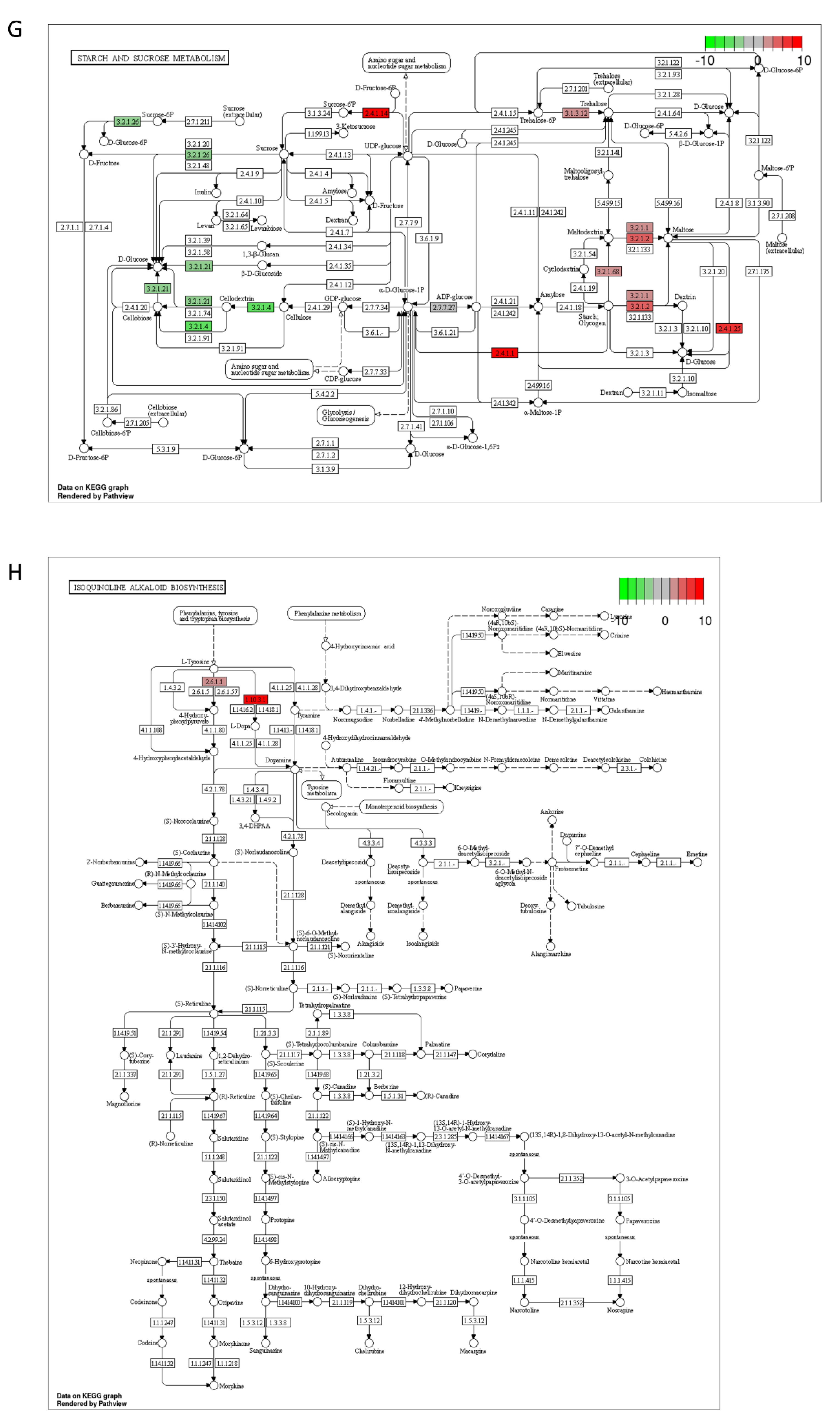
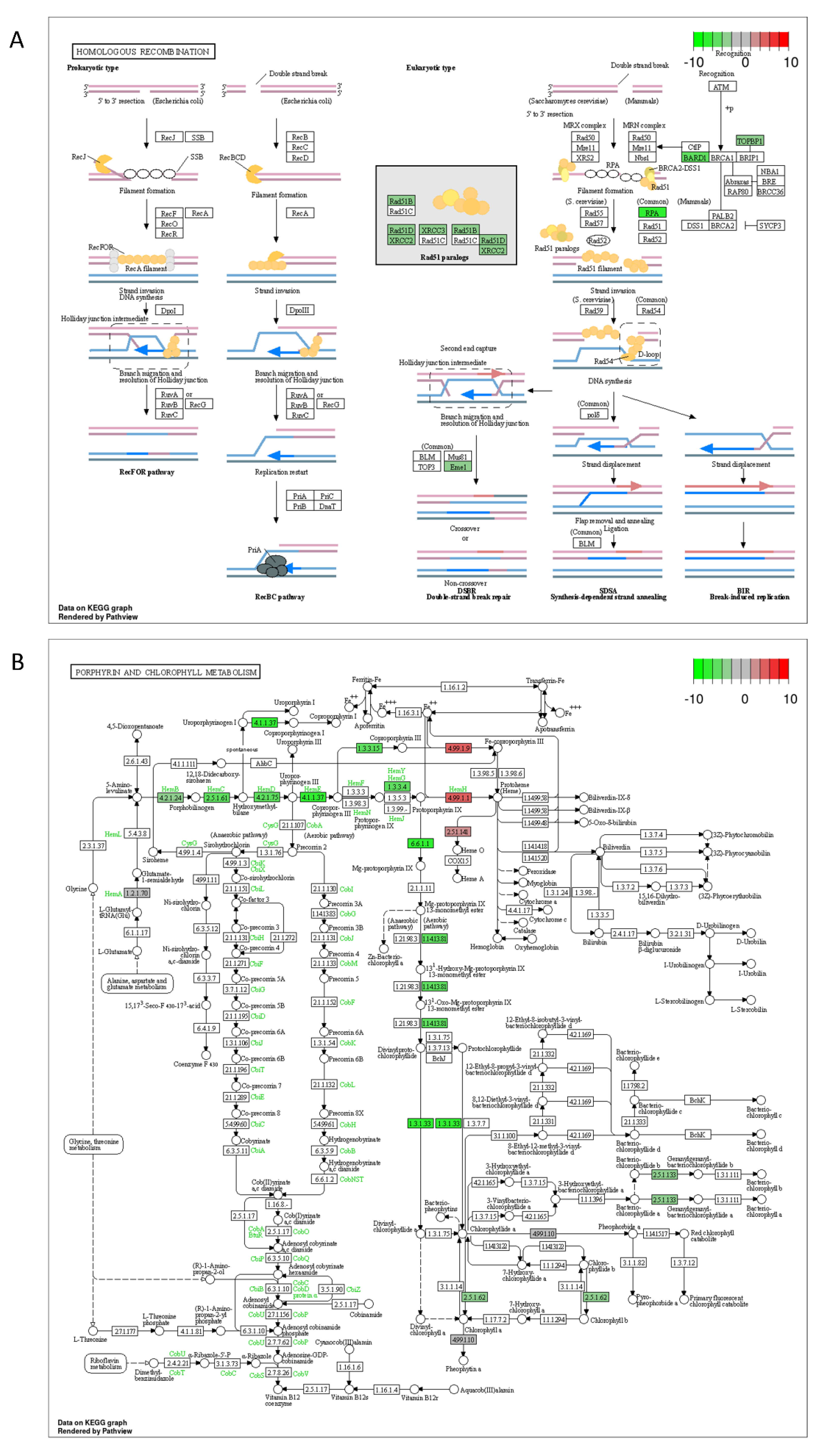
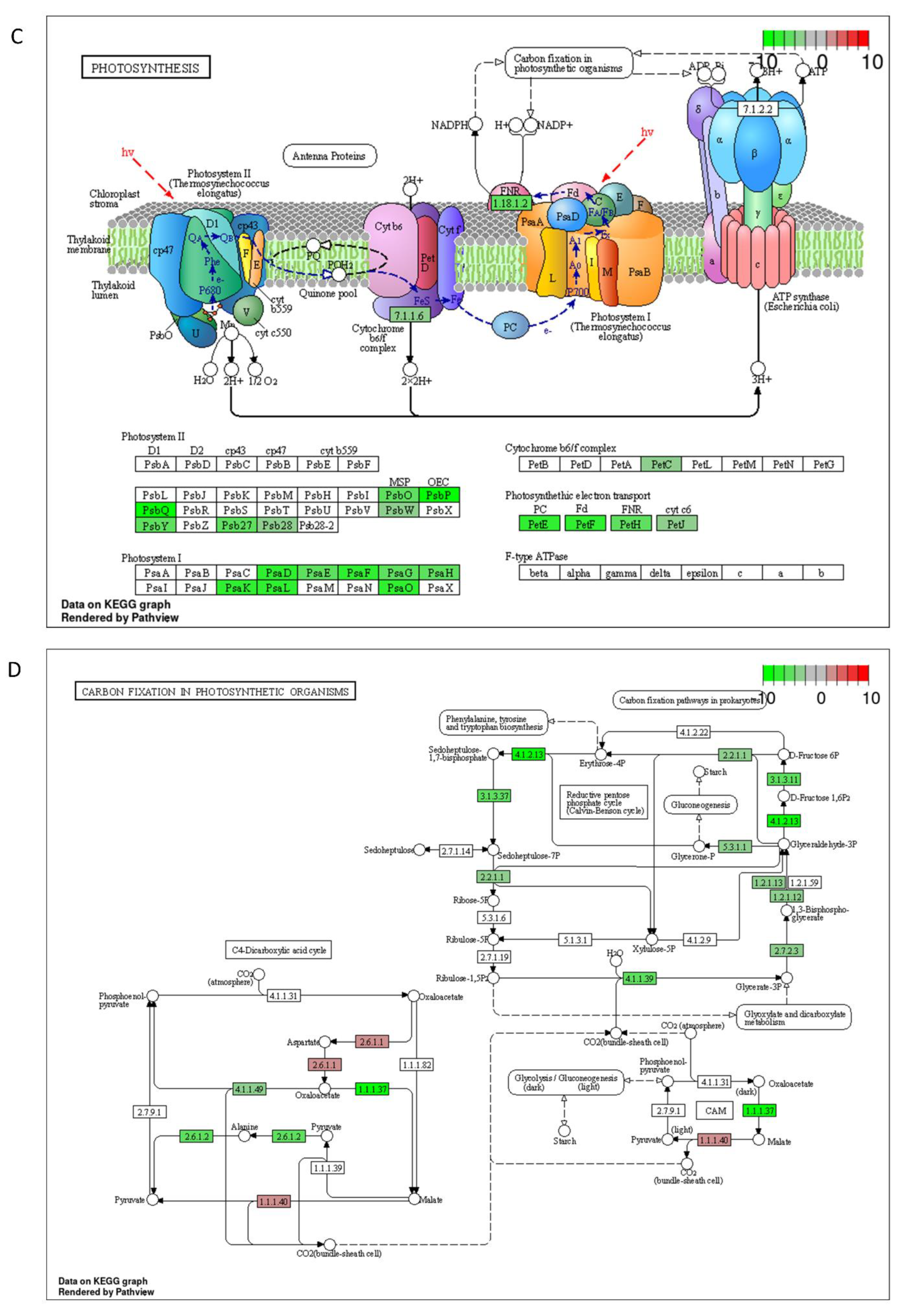
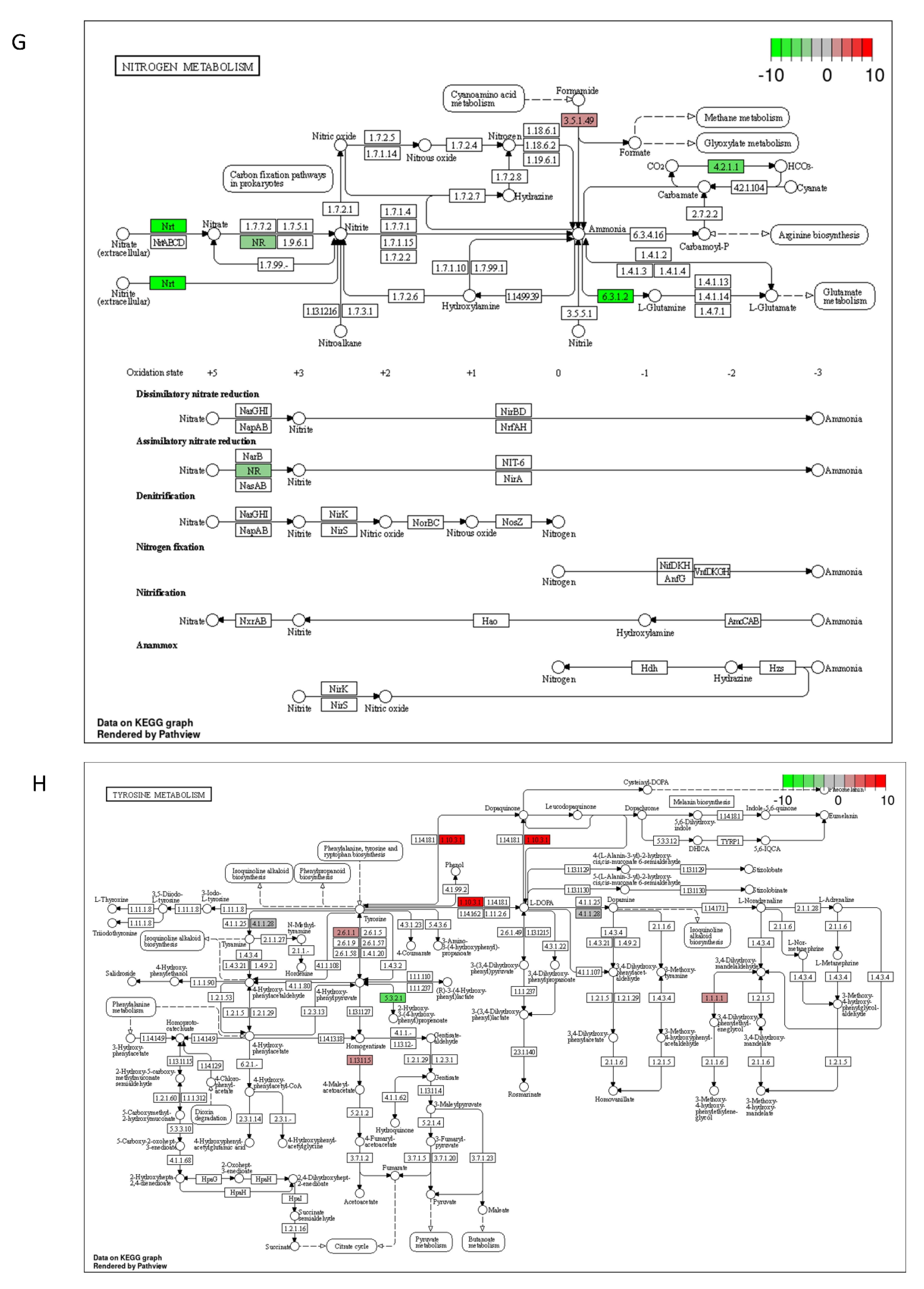
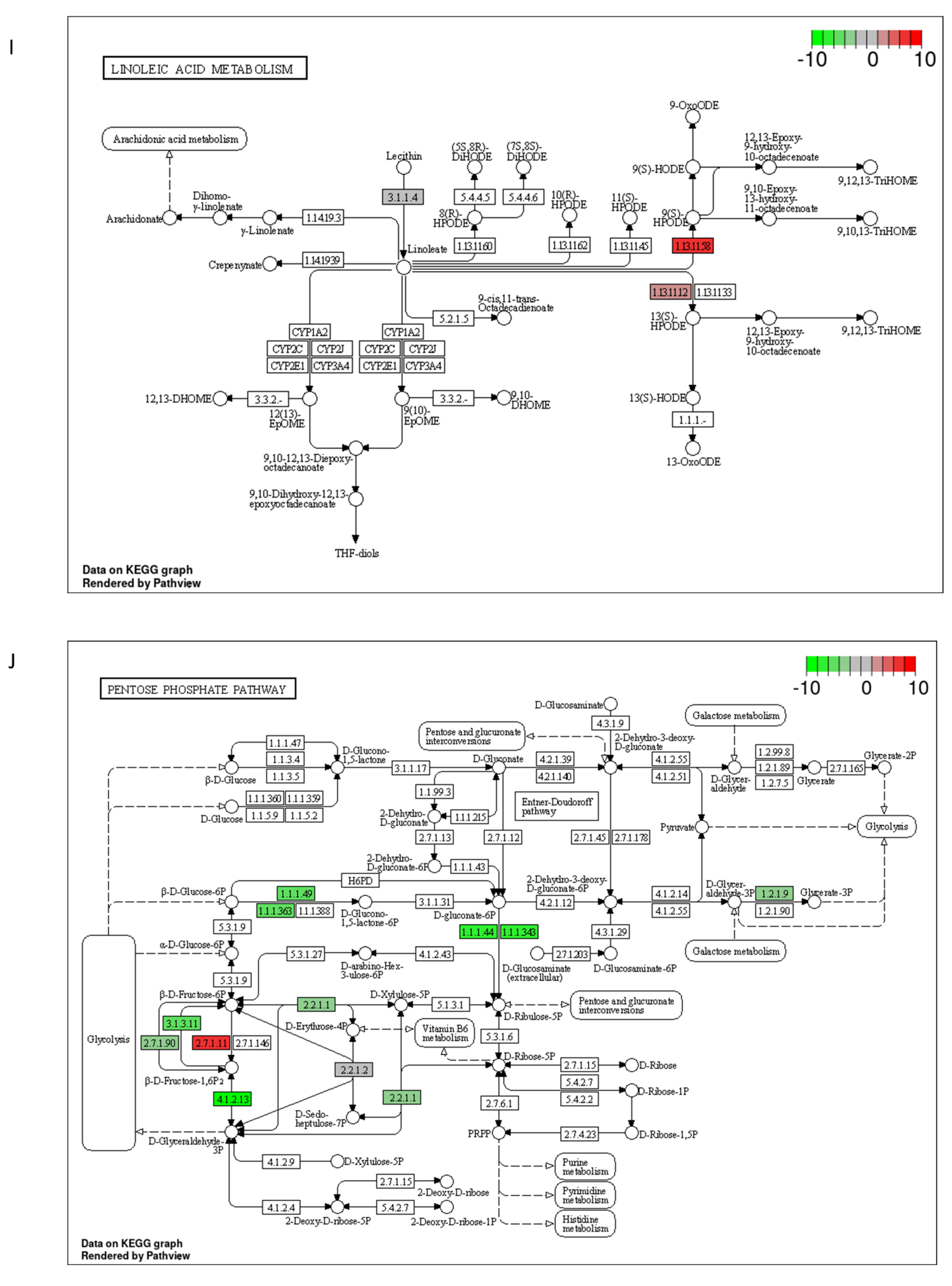
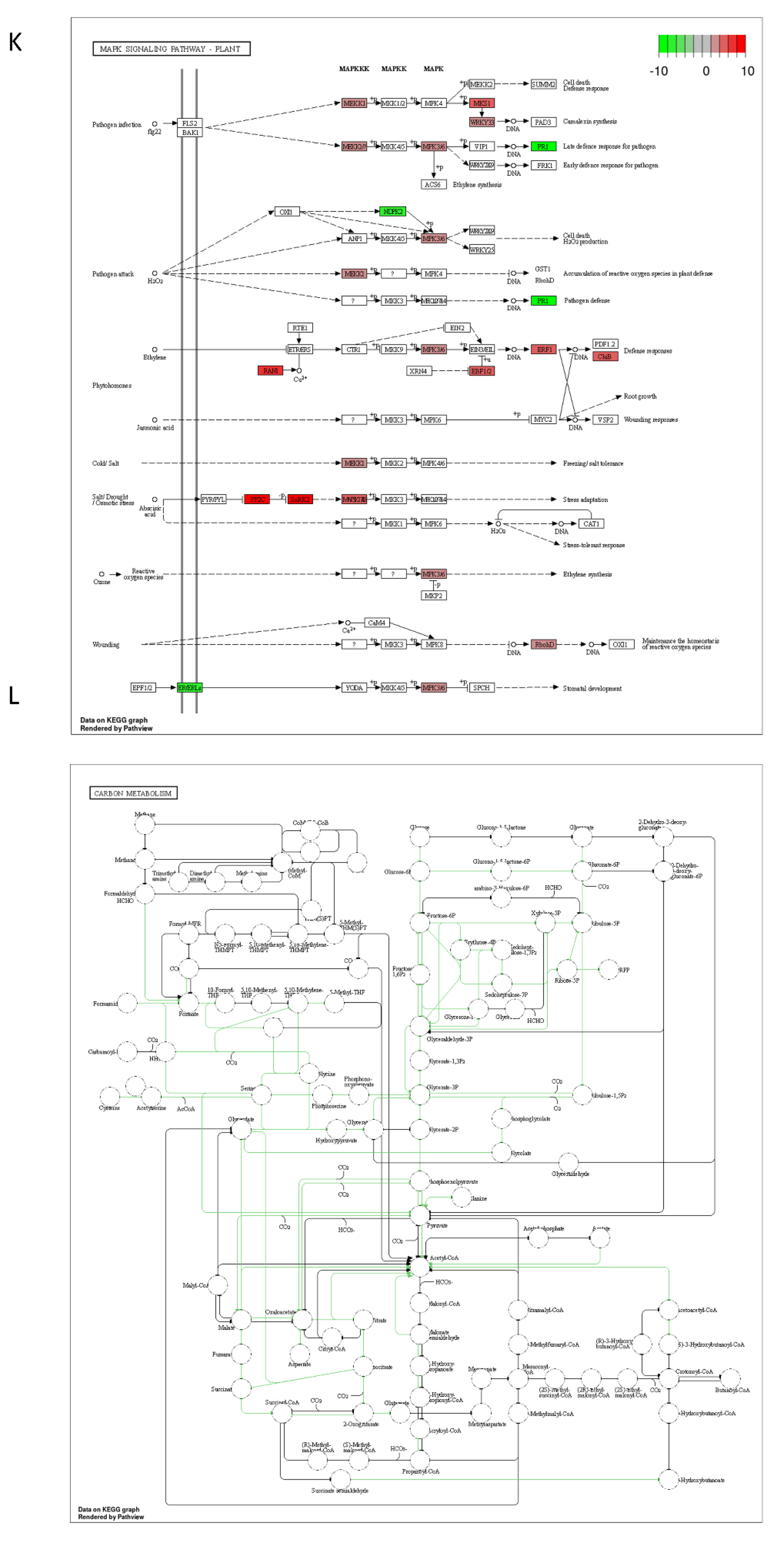
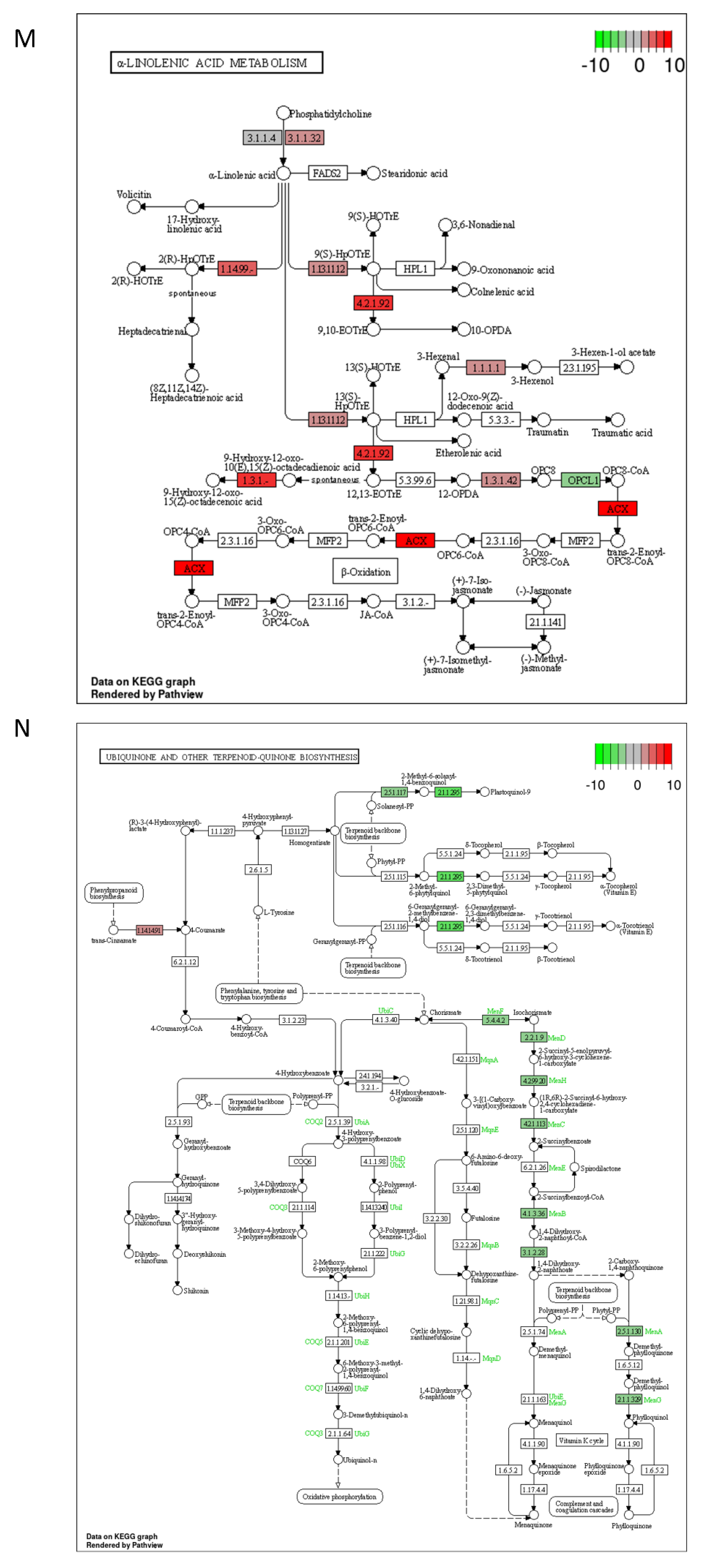
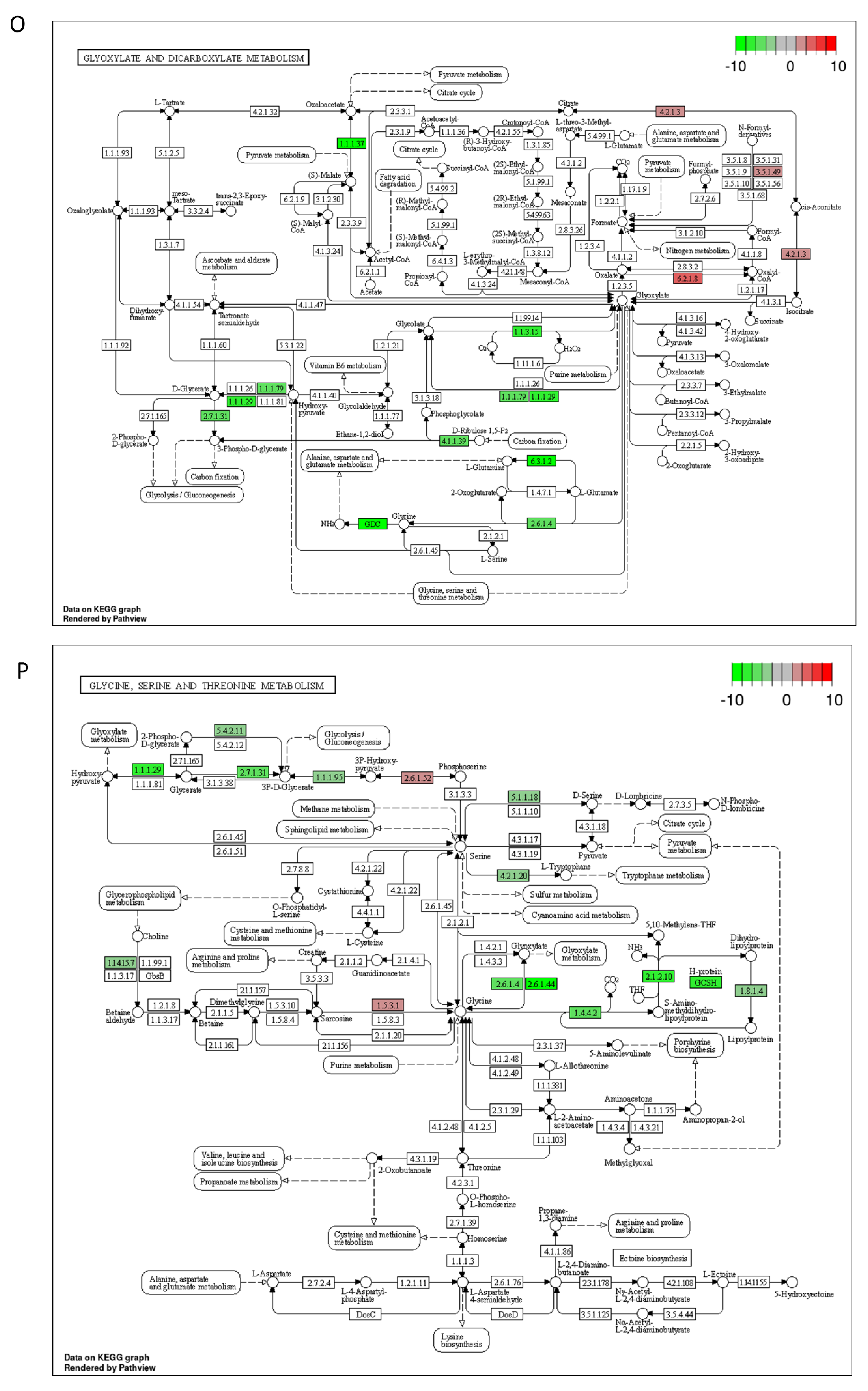
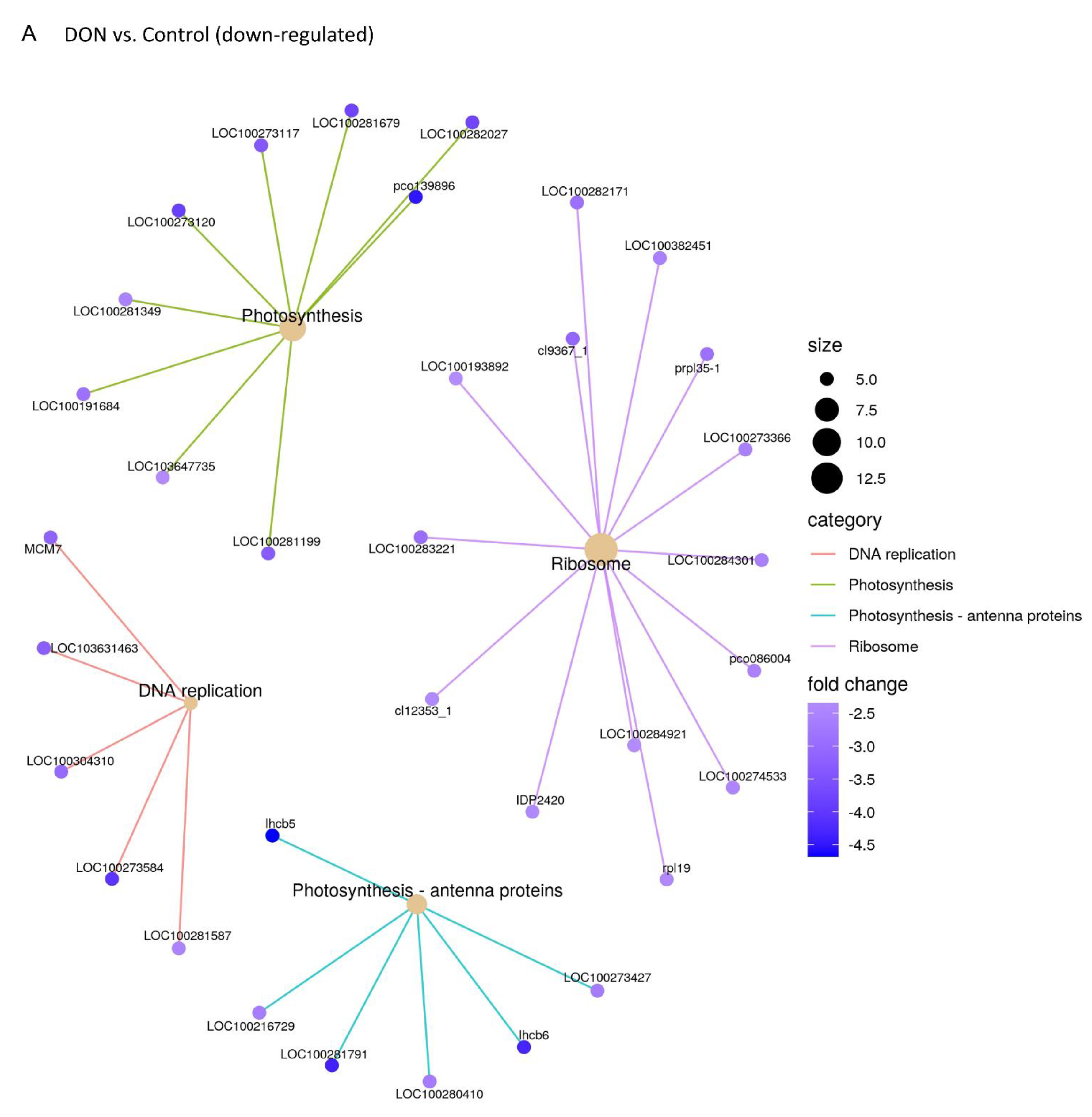
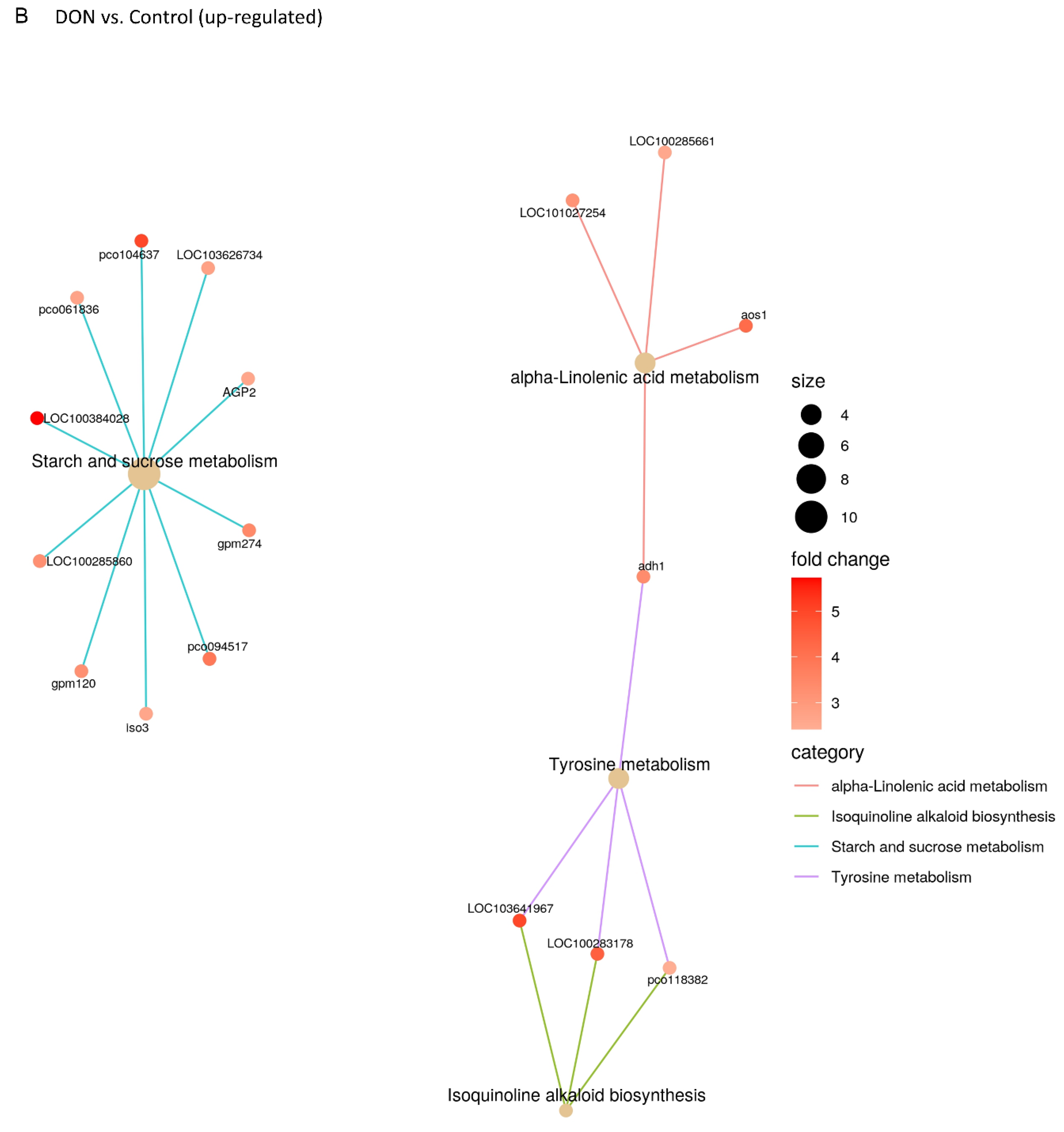
2.3. DON and 3-keto-DON, but Not 3-epi-DON, Significantly Modulated L. minor Transcriptome
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Duckweed (Lemna minor) Bioassays
4.3. Common Wheat (Triticum aestivum) Bioassays
4.4. HPLC Analysis of DON, 3-keto-DON, and 3-epi-DON
4.5. Lemna minor Transcriptome Sequencing and Analysis
4.6. Regression and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxins | A | B | R2adj | Model Significance |
|---|---|---|---|---|
| DON | −1.154 | 1.295 | 0.984 | <0.001 |
| 3-keto-DON | −2.615 | 1.383 | 0.931 | 0.005 |
| 3-epi-DON | −0.017 | 1.054 | 0.998 | <0.001 |
| Samples | Raw Reads | Clean Reads | Q30 (%) | GC Content (%) | Total Mapped (%) |
|---|---|---|---|---|---|
| Control 1 | 72,865,194 | 66,690,104 | 95.79 | 54.64 | 61.53 |
| Control 2 | 71,527,356 | 66,376,776 | 86.6 | 53.93 | 57 |
| Control 3 | 95,259,758 | 88,523,530 | 89.48 | 53.36 | 58.77 |
| DON 1 | 74,987,048 | 68,631,740 | 90.15 | 52.99 | 57.62 |
| DON 2 | 87,314,828 | 81,111,642 | 87.78 | 54.08 | 58.9 |
| DON 3 | 105,166,696 | 100,496,994 | 90.67 | 53.49 | 59.39 |
| 3-epi-DON 1 | 76,744,882 | 71,711,412 | 89.79 | 54.83 | 61.52 |
| 3-epi-DON 2 | 76,210,696 | 70,943,086 | 87.85 | 52.06 | 44.59 |
| 3-epi-DON 3 | 87,517,818 | 81,231,576 | 89.06 | 54.35 | 60.03 |
| 3-keto-DON 1 | 82,320,528 | 75,960,962 | 89.71 | 54.37 | 59.9 |
| 3-keto-DON 2 | 89,441,164 | 82,619,978 | 88.26 | 54.69 | 59.66 |
| 3-keto-DON 3 | 85,089,596 | 78,169,328 | 86.88 | 54.45 | 57.72 |
















References
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.; Zhou, T. Strategies and Methodologies for Developing Microbial Detoxification Systems to Mitigate Mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Megharaj, M.; Wu, C.Y.; Subashchandrabose, S.R.; Dai, C.C. Endophyte-assisted phytoremediation: Mechanisms and current application strategies for soil mixed pollutants. Crit. Rev. Biotechnol. 2020, 40, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Adegbeye, M.J.; Reddy, P.R.K.; Chilaka, C.A.; Balogun, O.B.; Elghandour, M.M.; Rivas-Caceres, R.R.; Salem, A.Z. Mycotoxin toxicity and residue in animal products: Prevalence, consumer exposure and reduction strategies—A review. Toxicon 2020, 177, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajslova, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Lana, F.D.; Madden, L.V.; Paul, P.A. Natural Occurrence of Maize Gibberella Ear Rot and Contamination of Grain with Mycotoxins in Association with Weather Variables. Plant Dis. 2021, 105, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Battilani, P.; Leggieri, M.C.; Jiang, Y.; Haesaert, G.; Lanubile, A.; Mahuku, G.; Akos, M.; Ortega-Beltran, A.; Pasti, M.; et al. Perspectives on global mycotoxin issues and management from the MycoKey Maize Working Group. Plant Dis. 2021, 105, 525–537. [Google Scholar] [CrossRef]
- Andrade, P.D.; Dias, J.V.; Souza, D.M.; Brito, A.P.; van Donkersgoed, G.; Pizzutti, I.R.; Caldas, E.D. Mycotoxins in cereals and cereal-based products: Incidence and probabilistic dietary risk assessment for the Brazilian population. Food Chem. Toxicol. 2020, 143, 111572. [Google Scholar] [CrossRef]
- Kim, D.B.; Song, N.E.; Nam, T.G.; Lee, S.; Seo, D.; Yoo, M. Occurrence of emerging mycotoxins in cereals and cereal-based products from the Korean market using LC-MS/MS. Food Addit. Contam. Part A 2019, 36, 289–295. [Google Scholar] [CrossRef]
- Shima, J.; Takase, S.; Takahashi, Y.; Iwai, Y.; Fujimoto, H.; Yamazaki, M.; Ochi, K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl. Environ. Microbiol. 1997, 63, 3825–3830. [Google Scholar] [CrossRef] [Green Version]
- Schweiger, W.; Pasquet, J.C.; Nussbaumer, T.; Paris, M.P.; Wiesenberger, G.; Macadre, C.; Ametz, C.; Berthiller, F.; Lemmens, M.; Saindrenan, P.; et al. Functional characterization of two clusters of Brachypodium distachyon UDP-glycosyltransferases encoding putative deoxynivalenol detoxification genes. Mol. Plant Microbe Interact. 2013, 26, 781–792. [Google Scholar] [CrossRef] [Green Version]
- Binder, J.; Horvath, E.M.; Schatzmayr, G.; Ellend, N.; Danner, H.; Krska, R.; Braun, R. Screening for deoxynivalenol-detoxifying anaerobic rumen microorganisms. Cereal Res. Commun. 1997, 25, 343–346. [Google Scholar] [CrossRef]
- Li, X.Z.; Zhu, C.; de Lange, C.F.; Zhou, T.; He, J.; Yu, H.; Gong, J.; Young, J.C. Efficacy of detoxification of deoxynivalenol-contaminated corn by Bacillus sp. LS100 in reducing the adverse effects of the mycotoxin on swine growth performance. Food Addit. Contam. Part A 2011, 28, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Mu, P.; Wen, J.; Sun, Y.; Chen, Q.; Deng, Y. Detoxification of trichothecene mycotoxins by a novel bacterium, Eggerthella sp. DII-9. Food Chem. Toxicol. 2018, 112, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Mu, P.; Zhu, X.; Chen, X.; Tang, S.; Wu, Y.; Miao, X.; Wang, X.; Wen, J.; Deng, Y. Dual Function of a Novel Bacterium, Slackia sp. D-G6: Detoxifying Deoxynivalenol and Producing the Natural Estrogen Analogue, Equol. Toxins 2020, 12, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.J.; Shi, M.M.; Yang, P.; Huang, T.; Yuan, Q.S.; Yi, S.Y.; Wu, A.B.; Li, H.P.; Gao, C.B.; Zhang, J.B.; et al. Novel Soil Bacterium Strain Desulfitobacterium sp. PGC-3-9 Detoxifies Trichothecene Mycotoxins in Wheat via De-Epoxidation under Aerobic and Anaerobic Conditions. Toxins 2020, 12, 363. [Google Scholar] [CrossRef]
- He, J.W.; Hassan, Y.I.; Perilla, N.; Li, X.Z.; Boland, G.J.; Zhou, T. Bacterial Epimerization as a Route for Deoxynivalenol Detoxification: The Influence of Growth and Environmental Conditions. Front. Microbiol. 2016, 7, 572. [Google Scholar] [CrossRef] [Green Version]
- Carere, J.; Hassan, Y.I.; Lepp, D.; Zhou, T. The enzymatic detoxification of the mycotoxin deoxynivalenol: Identification of DepA from the DON epimerization pathway. Microb. Biotechnol. 2018, 11, 1106–1111. [Google Scholar] [CrossRef]
- Carere, J.; Hassan, Y.I.; Lepp, D.; Zhou, T. The Identification of DepB: An Enzyme Responsible for the Final Detoxification Step in the Deoxynivalenol Epimerization Pathway in Devosia mutans 17-2-E-8. Front. Microbiol. 2018, 9, 1573. [Google Scholar] [CrossRef]
- Ito, M.; Sato, I.; Ishizaka, M.; Yoshida, S.; Koitabashi, M.; Yoshida, S.; Tsushima, S. Bacterial cytochrome P450 system catabolizing the Fusarium toxin deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- He, J.W.; Yang, R.; Zhou, T.; Boland, G.J.; Scott, P.M.; Bondy, G.S. An epimer of deoxynivalenol: Purification and structure identification of 3-epi-deoxynivalenol. Food Addit. Contam. Part A 2015, 32, 1523–1530. [Google Scholar] [CrossRef]
- Hassan, Y.I.; He, J.W.; Perilla, N.; Tang, K.; Karlovsky, P.; Zhou, T. The enzymatic epimerization of deoxynivalenol by Devosia mutans proceeds through the formation of 3-keto-DON intermediate. Sci. Rep. 2017, 7, 6929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Y.I.; Zhou, T. Addressing the mycotoxin deoxynivalenol contamination with soil-derived bacterial and enzymatic transformations targeting the C3 carbon. World Mycotoxin J. 2018, 11, 101–112. [Google Scholar] [CrossRef]
- Hassan, Y.I.; Watts, C.; Li, X.Z.; Zhou, T. A novel Peptide-binding motifs inference approach to understand deoxynivalenol molecular toxicity. Toxins 2015, 7, 1989–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [Green Version]
- He, J.W.; Bondy, G.S.; Zhou, T.; Caldwell, D.; Boland, G.J.; Scott, P.M. Toxicology of 3-epi-deoxynivalenol, a deoxynivalenol-transformation product by Devosia mutans 17-2-E-8. Food Chem. Toxicol. 2015, 84, 250–259. [Google Scholar] [CrossRef]
- Hassan, Y.I.; He, J.W.; Lepp, D.; Zhou, T. Understanding the Bacterial Response to Mycotoxins: The Transcriptomic Analysis of Deoxynivalenol-Induced Changes in Devosia mutans 17-2-E-8. Front. Pharmacol. 2019, 10, 1098. [Google Scholar] [CrossRef] [Green Version]
- Mayer, E.; Novak, B.; Springler, A.; Schwartz-Zimmermann, H.E.; Nagl, V.; Reisinger, N.; Hessenberger, S.; Schatzmayr, G. Effects of deoxynivalenol (DON) and its microbial biotransformation product deepoxy-deoxynivalenol (DOM-1) on a trout, pig, mouse, and human cell line. Mycotoxin Res. 2017, 33, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin Res. 2017, 33, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.; Liaubet, L.; Schatzmayr, G.; Berthiller, F.; Moll, W.D.; et al. Intestinal toxicity of the masked mycotoxin deoxynivalenol-3-beta-D-glucoside. Arch. Toxicol. 2016, 90, 2037–2046. [Google Scholar] [CrossRef]
- Pierron, A.; Bracarense, A.; Cossalter, A.M.; Laffitte, J.; Schwartz-Zimmermann, H.E.; Schatzmayr, G.; Pinton, P.; Moll, W.D.; Oswald, I.P. Deepoxy-deoxynivalenol retains some immune-modulatory properties of the parent molecule deoxynivalenol in piglets. Arch. Toxicol. 2018, 92, 3381–3389. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.W.; Currie, V.; Richardson, A.J.; Duncan, G.; Holtrop, G.; Farquharson, F.; Louis, P.; Pinton, P.; Oswald, I.P. Porcine Small and Large Intestinal Microbiota Rapidly Hydrolyze the Masked Mycotoxin Deoxynivalenol-3-Glucoside and Release Deoxynivalenol in Spiked Batch Cultures In Vitro. Appl. Environ. Microbiol. 2018, 84, e02106-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratz, S.W.; Dinesh, R.; Yoshinari, T.; Holtrop, G.; Richardson, A.J.; Duncan, G.; MacDonald, S.; Lloyd, A.; Tarbin, J. Masked trichothecene and zearalenone mycotoxins withstand digestion and absorption in the upper GI tract but are efficiently hydrolyzed by human gut microbiota in vitro. Mol. Nutr. Food Res. 2017, 61, 1600680. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, I.; De Mets, L.; De Boevre, M.; Uka, V.; Di Mavungu, J.D.; De Saeger, S.; De Gelder, L.; Audenaert, K. Microbial Detoxification of Deoxynivalenol (DON), Assessed via a Lemna minor L. Bioassay, through Biotransformation to 3-epi-DON and 3-epi-DOM-1. Toxins 2017, 9, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boenisch, M.J.; Schäfer, W. Fusarium graminearum forms mycotoxin producing infection structures on wheat. BMC Plant Biol. 2011, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Jacobsen, S.; Jorgensen, H.J.; Collinge, D.B.; Svensson, B.; Finnie, C. Fusarium graminearum and Its Interactions with Cereal Heads: Studies in the Proteomics Era. Front. Plant Sci. 2013, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Qin, X.; Guo, Y.; Zhang, Q.; Ma, Q.; Ji, C.; Zhao, L. Enzymatic degradation of deoxynivalenol by a novel bacterium, Pelagibacterium halotolerans ANSP101. Food Chem. Toxicol. 2020, 140, 111276. [Google Scholar] [CrossRef]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef]
- Bracarense, A.; Pierron, A.; Pinton, P.; Gerez, J.R.; Schatzmayr, G.; Moll, W.D.; Zhou, T.; Oswald, I.P. Reduced toxicity of 3-epi-deoxynivalenol and de-epoxy-deoxynivalenol through deoxynivalenol bacterial biotransformation: In vivo analysis in piglets. Food Chem. Toxicol. 2020, 140, 111241. [Google Scholar] [CrossRef]
- Bracarense, A.P.F.L.; Basso, K.M.; Da Silva, E.O.; Payros, D.; Oswald, I.P. Deoxynivalenol in the liver and lymphoid organs of rats: Effects of dose and duration on immunohistological changes. World Mycotoxin J. 2017, 10, 89–96. [Google Scholar] [CrossRef]
- He, Y.; Yin, X.; Dong, J.; Yang, Q.; Wu, Y.; Gong, Z. Transcriptome Analysis of Caco-2 Cells upon the Exposure of Mycotoxin Deoxynivalenol and Its Acetylated Derivatives. Toxins 2021, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wu, X.; Zhang, W.; Ye, P.; Wang, Y.; Zhu, W.; Tao, Q.; Xu, Y.; Shang, J.; Zhao, D.; et al. Transcriptional analysis of deoxynivalenol-induced apoptosis of sow ovarian granulosa cell. Reprod. Domest. Anim. 2020, 55, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yan, G.; Chang, J.; Wang, P.; Yin, Q.; Liu, C.; Zhu, Q.; Lu, F. Comparative Transcriptome Analysis Reveals the Protective Mechanism of Glycyrrhinic Acid for Deoxynivalenol-Induced Inflammation and Apoptosis in IPEC-J2 Cells. Oxidative Med. Cell. Longev. 2020, 2020, 5974157. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.E.; Jeong, J.Y.; Lee, Y.; Lee, H.J.; Kim, M.S.; Kim, D.W.; Jung, H.J.; Choe, C.; Oh, Y.K.; Lee, S.D. Deoxynivalenol- and zearalenone-contaminated feeds alter gene expression profiles in the livers of piglets. Asian Australas. J. Anim. Sci. 2018, 31, 595–606. [Google Scholar] [CrossRef] [Green Version]
- Di, R.; Zhang, H.; Lawton, M.A. Transcriptome Analysis of C. elegans Reveals Novel Targets for DON Cytotoxicity. Toxins 2018, 10, 262. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.J.; Carere, J.; Fitzgerald, T.L.; Stiller, J.; Covarelli, L.; Xu, Q.; Gubler, F.; Colgrave, M.L.; Gardiner, D.M.; Manners, J.M.; et al. The Fusarium crown rot pathogen Fusarium pseudograminearum triggers a suite of transcriptional and metabolic changes in bread wheat (Triticum aestivum L.). Ann. Bot. 2017, 119, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Puri, K.D.; Yan, C.; Leng, Y.; Zhong, S. RNA-Seq Revealed Differences in Transcriptomes between 3ADON and 15ADON Populations of Fusarium graminearum In Vitro and In Planta. PLoS ONE 2016, 11, e0163803. [Google Scholar] [CrossRef] [Green Version]
- Hofstad, A.N.; Nussbaumer, T.; Akhunov, E.; Shin, S.; Kugler, K.G.; Kistler, H.C.; Mayer, K.F.; Muehlbauer, G.J. Examining the Transcriptional Response in Wheat Near-Isogenic Lines to Infection and Deoxynivalenol Treatment. Plant Genome 2016, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Lim, W.; Park, S.; Kim, J.; You, S.; Song, G. Deoxynivalenol induces apoptosis and disrupts cellular homeostasis through MAPK signaling pathways in bovine mammary epithelial cells. Environ. Pollut. 2019, 252, 879–887. [Google Scholar] [CrossRef]
- Lucioli, J.; Pinton, P.; Callu, P.; Laffitte, J.; Grosjean, F.; Kolf-Clauw, M.; Oswald, I.P.; Bracarense, A.P. The food contaminant deoxynivalenol activates the mitogen activated protein kinases in the intestine: Interest of ex vivo models as an alternative to in vivo experiments. Toxicon 2013, 66, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Hassan, Y.I.; Zhu, H.; Zhu, Y.; Zhou, T. Beyond Ribosomal Binding: The Increased Polarity and Aberrant Molecular Interactions of 3-epi-deoxynivalenol. Toxins 2016, 8, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhu, Y.; Abraham, N.; Li, X.Z.; Kimber, M.; Zhou, T. The Ribosome-Binding Mode of Trichothecene Mycotoxins Rationalizes Their Structure-Activity Relationships. Int. J. Mol. Sci. 2021, 22, 1604. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.; Schatzmayr, G.; He, J.W.; Zhou, T.; Moll, W.D.; et al. Microbial biotransformation of DON: Molecular basis for reduced toxicity. Sci. Rep. 2016, 6, 29105. [Google Scholar] [CrossRef] [Green Version]
- Tamburic-Ilincic, L.; Falk, D.; Schaafsma, A. Fusarium ratings in Ontario Winter Wheat Performance Trial (OWWPT) using an index that combines Fusarium head blight symptoms and deoxynivalenol levels. Czech J. Genet. Plant Breed. 2011, 47, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Sari, E.; Berraies, S.; Knox, R.E.; Singh, A.K.; Ruan, Y.; Cuthbert, R.D.; Pozniak, C.J.; Henriquez, M.A.; Kumar, S.; Burt, A.J.; et al. High density genetic mapping of Fusarium head blight resistance QTL in tetraploid wheat. PLoS ONE 2018, 13, e0204362. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Powell, D.R. Degust: Interactive RNA-Seq Analysis; Version 4.1.1; Zenodo: London, UK, 2019. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [Green Version]






| Experiment | Fronds per Well (Mean ± SD) | Fresh-Weight in mg/Well (Mean ± SD) | |
|---|---|---|---|
| (A) DON concentration (µg/mL) | 0 | 24.56 ± 4.45 a | 18.33 ± 4.38 a |
| 0.5 | 18.88 ± 3.07 ab | 13.75 ± 2.23 ab | |
| 1 | 13.56 ± 2.54 bcd | 9.50 ± 1.47 bc | |
| 1.75 | 9.94 ± 1.73 cde | 6.22 ± 1.03 c | |
| 2.5 | 7.88 ± 1.11 cde | 5.64 ± 0.82 c | |
| 5 | 6.06 ± 0.13 de | 4.46 ± 0.70 c | |
| (B) 3-keto-DON concentration (µg/mL) | 0 | 21.75 ± 3.03 a | 14.57 ± 2.74 a |
| 0.5 | 10.67 ± 1.70 b | 7.19 ± 0.46 b | |
| 1 | 8.50 ± 1.30 bc | 5.95 ± 1.01 b | |
| 1.75 | 6.50 ± 0.66 bc | 4.84 ± 0.84 b | |
| 2.5 | 6.00 ± 0.00 c | 4.43 ±0.49 b | |
| 5 | 6.00 ± 0.00 c | 3.78 ± 0.95 b | |
| 3-epi-DON Concentration (µg/mL) | Fronds per Well | Percentage of Control (%) | Fresh Weight in mg/Well | Percentage of Control (%) |
|---|---|---|---|---|
| 0 | 24.83 | 100.0 (control) | 18.26 | 100.0 (control) |
| 5 | 25.13 | 101.2 | 18.05 | 98.8 |
| 25 | 18.75 | 75.5 | 12.37 | 67.7 |
| 50 | 14.5 | 58.4 | 8.5 | 46.6 |
| 125 | 8.25 | 33.2 | 5.38 | 29.5 |
| AC Morley (Moderately-Resistant) | |||
| Treatment/Measurements [mean ± SD] | Coleoptile’s Length (mm/seedling) | Coleoptile’s Fresh-Weight mg/seedling) | Root’s Fresh-Weight (mg/seedling) |
| Control | 130.75 ± 8.83 a | 73.54 ± 1.70 a | 36.60 ± 8.80 a |
| DON (30 µg/mL) | 60.73 ± 12.21 b | 48.29 ± 8.24 b | 17.49 ± 6.19 b |
| 3-keto-DON (30 µg/mL) | 68.57 ± 13.05 b | 55.95 ± 2.20 b | 21.90 ± 5.11 a |
| 3-epi-DON (30 µg/mL) | 130.21 ± 20.40 a | 70.92 ± 5.75 a | 33.92 ± 5.52 a |
| AC Sampson (Susceptible) | |||
| Treatment/Measurements [mean ± SD] | Coleoptile’s Length (mm/seedling) | Coleoptile’s Fresh-Weight mg/seedling) | Root’s Fresh-Weight (mg/seedling) |
| Control | 157.90 ± 3.92 a | 69.84 ± 5.37 a | 33.00 ± 4.60 a |
| DON (30 µg/mL) | 48.97 ± 3.95 b | 39.28 ± 1.44 b | 5.77 ± 0.50 b |
| 3-keto-DON (30 µg/mL) | 52.47 ± 3.00 b | 38.53 ± 4.93 b | 5.13 ± 1.16 b |
| 3-epi-DON (30 µg/mL) | 165.48 ± 5.41 a | 76.57 ± 4.96 a | 41.62 ± 9.45 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-Z.; Hassan, Y.I.; Lepp, D.; Zhu, Y.; Zhou, T. 3-keto-DON, but Not 3-epi-DON, Retains the in Planta Toxicological Potential after the Enzymatic Biotransformation of Deoxynivalenol. Int. J. Mol. Sci. 2022, 23, 7230. https://doi.org/10.3390/ijms23137230
Li X-Z, Hassan YI, Lepp D, Zhu Y, Zhou T. 3-keto-DON, but Not 3-epi-DON, Retains the in Planta Toxicological Potential after the Enzymatic Biotransformation of Deoxynivalenol. International Journal of Molecular Sciences. 2022; 23(13):7230. https://doi.org/10.3390/ijms23137230
Chicago/Turabian StyleLi, Xiu-Zhen, Yousef I. Hassan, Dion Lepp, Yan Zhu, and Ting Zhou. 2022. "3-keto-DON, but Not 3-epi-DON, Retains the in Planta Toxicological Potential after the Enzymatic Biotransformation of Deoxynivalenol" International Journal of Molecular Sciences 23, no. 13: 7230. https://doi.org/10.3390/ijms23137230