
Role of Sodium-Glucose Co-Transporter 2 Inhibitors in the Regulation of Inflammatory Processes in Animal Models

 , , , ,
, , , ,
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Effect | Tissue/Cell Type | Animal Model |
|---|---|---|
| Anti-inflammatory | Bladder | Diabetic Akita mice [15] |
| RAW 264.7 macrophages [16] | - | |
| 3T3-L1 cells differentiated into adipocytes [17] | - | |
| Brain Serum, liver, and cecum Gastrointestinal tract Brain Plasma | Aged mice [18] High-fat diet-fed mice [19] Rats with irritable bowel syndrome [20] Mice with Alzheimer’s disease [21] Ovariectomized rats under inflammation conditions [22] | |
| Body weight reduction | - | Diabetic db/db mice [23,24] High-fat diet-fed mice [19,25] |
| Lipolysis | 3T3-L1 cells differentiated into adipocytes [16] | - |
| Anti-oxidant | Brain and liver | Aged mice [18] |
| PC12 cells [18] | - | |
| Skeletal muscle | Mice with exercise-induced fatigue [26] | |
| HepG2 cells [27] | - | |
| Liver Serum, liver, and cecum | Rats with hepatic fibrosis [28] High-fat diet-fed mice [19] | |
| Renoprotective | Kidney | Diabetic db/db mice [23] |
| Hepatoprotective | Liver | Diabetic mice with NAFLD [29] |
| Liver | Rats with hepatic fibrosis [28] | |
| Liver | High-fat diet-fed mice [19] | |
| Regulation of gut microbiota homeostasis | Gut microbiota Gut microbiota | High-fat diet-fed mice [19,25] Diabetic db/db mice [24] |
| Bone homeostasis | MC3T3-E1 cells differentiated into osteoblasts and in bone Bone | SAM mice [30] Ovariectomized rats under inflammation conditions [22] |
| Improvement of cognitive function | Brain Brain | Mice with Alzheimer’s disease [21] Swiss mice [31] |
| Anti-cancer | KYSE450 and KYSE30 cell lines [32] | - |
| Cardioprotective | Heart ACBRI 5118 cells [33] HUVEC cells [34] Heart | Diabetic db/db mice [35] - Guinea Pig [36] |
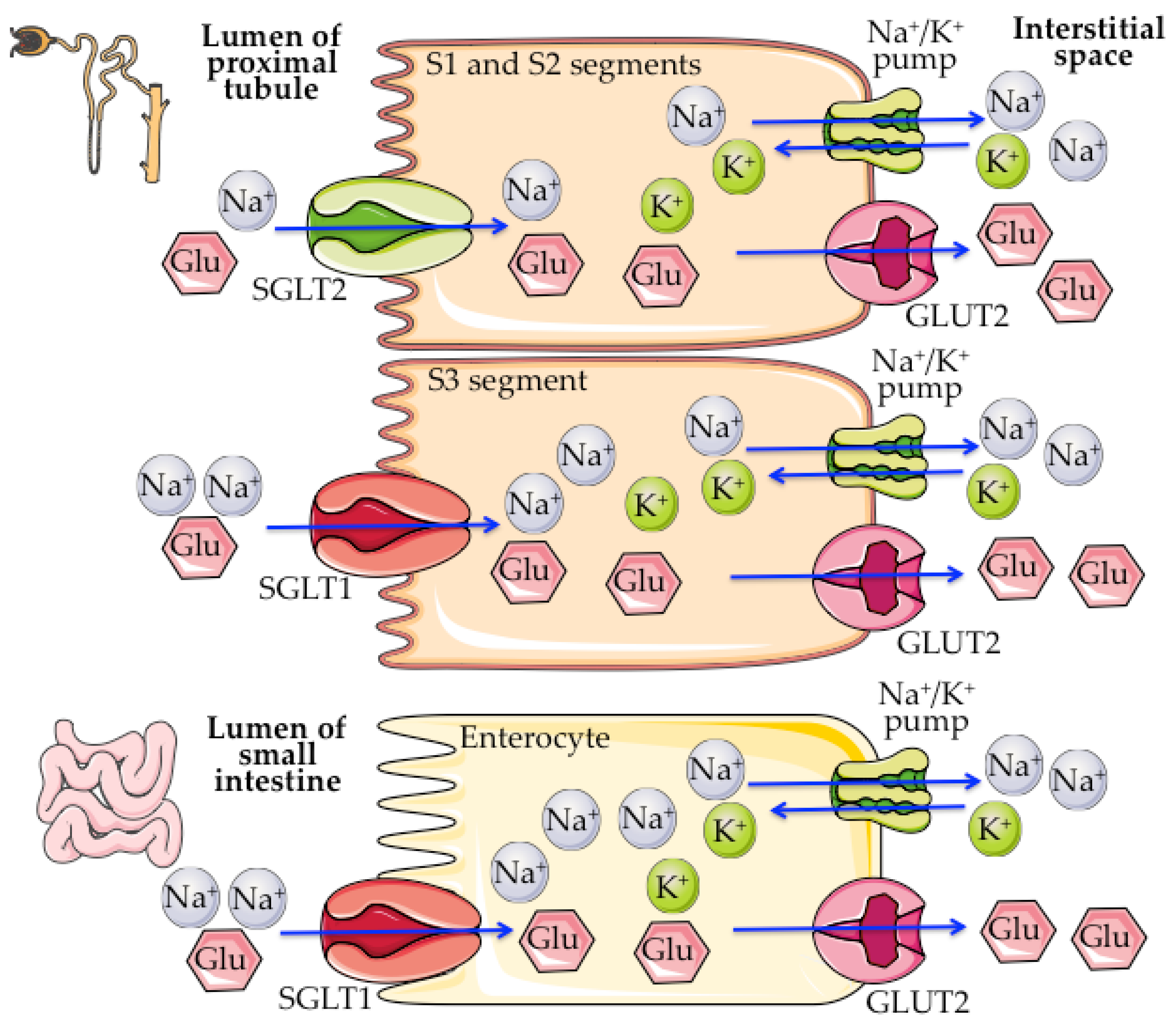
2. SGLT1/2 Biology
3. SGLT2 Inhibitors
| Generic Name | Brand Name | Company | SGLT2 Selectivity over SGLT1 | First Global Approval | Therapeutic Use |
|---|---|---|---|---|---|
| Tofogliflozin | Apleway® | Chugai Pharmaceutical Co. | ~2912 fold [74] | PMDA-2014 [75] | T2DM [75] |
| Empagliflozin | Jardiance® | Boehringer Ingelheim GmbH. and Eli Lilly and Company | ~2600 fold [76] | FDA-2014 [77] FDA-2016 [78] FDA-2021 [79] FDA-2022 [80] | T2DM [77] CV death in T2DM [78] HFrEF [79] Heart failure [80] |
| Bexagliflozin | - | Theracos Inc. | ~2435 fold [81] | Under clinical trials | - |
| Ertugliflozin | Steglatro™ | Merck & Co. and Pfizer Inc. | ~2200 fold [82] | FDA-2017 [83] | T2DM [83] |
| Luseogliflozin | Lusefi® | Taisho Pharmaceutical Holdings Co. | ~1730 fold [84] | PMDA-2014 [85] | T2DM [85] |
| Dapagliflozin | Forxiga® (E.U.) Farxiga® (U.S.) | AstraZeneca and Bristol-Myers Squibb Co. | ~1200 fold [86] | EMA-2012 [87] FDA-2020 [88] FDA-2021 [89] | T2DM [87] HFrEF [88] CKD [89] |
| # Remogliflozin | Remozen™ | Glenmark Pharmaceuticals Ltd. | ~902 fold [70] | CDSCO-2019 [90] | T2DM [90] |
| # Sergliflozin | - | Glaxo Smith Kline (GSK) Plc. | ~300 fold [69] | Discontinued after Phase II [43] | - |
| Ipragliflozin | Suglat® | Astellas Pharma Inc. and Kotobuki Pharmaceutical Co. | ~254 fold [91] | PMDA -2014 [92] | T2DM [92] |
| Canagliflozin | Invokana® | Janssen Global Services, L.L.C. and Mitsubishi Tanabe Pharma Co. | ~155 fold [93] | FDA-2013 [94] FDA-2018 [95] FDA-2019 [96] | T2DM [94] CV risk in T2DM [95] DKD and risk of hospitalization for HF in T2DM [96] |
| # T-1095 | - | Tanabe Seiyaku Company Ltd. | ~59 fold [76] | Discontinued after Phase II [97] | - |
| Licogliflozin | - | Novartis International A.G. | ~35 fold [98] | Under clinical trials | - |
| Sotagliflozin | Zynquista™ | Sanofi-Aventis Group S.A. and Lexicon Pharmaceuticals, Inc. | ~20 fold [99] | EMA-2019 [72] | T1DM [72] |
| # Phlorizin | - | - | ~13 fold [76] | - | - |
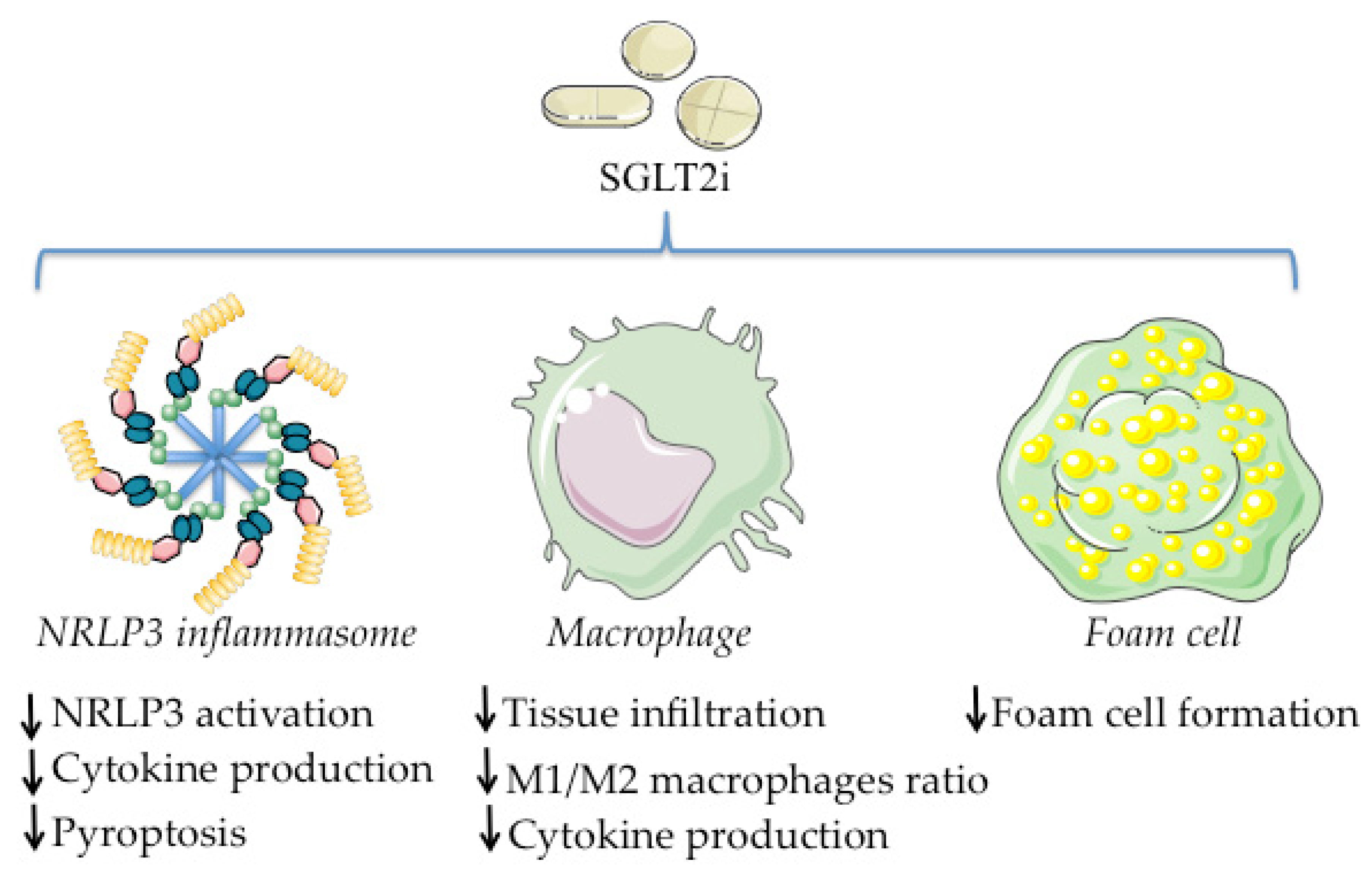
4. SGLT2i and Inflammation
4.1. Regulation of Macrophage Tissue Infiltration, Polarization, and Cytokine Production
4.1.1. Dapagliflozin
4.1.2. Canagliflozin
4.1.3. Empagliflozin
4.1.4. Other SGLT2i
4.2. Effects on NLRP3 Inflammasome
4.2.1. Dapagliflozin
4.2.2. Canagliflozin
4.2.3. Empagliflozin
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rossetti, L.; Smith, D.; Shulman, G.I.; Papachristou, D.; DeFronzo, R.A. Correction of Hyperglycemia with Phlorizin Normalizes Tissue Sensitivity to Insulin in Diabetic Rats. J. Clin. Investig. 1987, 79, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- Chasis, H.; Jolliffe, N.; Smith, H.W. The Action of Phlorizin on the Excretion of Glucose, Xylose, Sucrose, Creatinine and Urea by Man. J. Clin. Investig. 1933, 12, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.-I. Sodium-Glucose Cotransporter 2 (SGLT2) Inhibitors from Natural Products: Discovery of Next-Generation Antihyperglycemic Agents. Molecules 2016, 21, 1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.R. Apple Trees to Sodium Glucose Co-Transporter Inhibitors: A Review of SGLT2 Inhibition. Clin. Diabetes 2010, 28, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Cao, J.; Zhao, T.; Liu, Y.; Khan, A.; Cheng, G. The Bioavailability, Extraction, Biosynthesis and Distribution of Natural Dihydrochalcone: Phloridzin. Int. J. Mol. Sci. 2021, 22, 962. [Google Scholar] [CrossRef]
- Petersen, C. Analyse Des Phloridzins. Ann. Pharm. 1835, 15, 178. [Google Scholar] [CrossRef] [Green Version]
- Ehrenkranz, J.R.L.; Lewis, N.G.; Kahn, C.R.; Roth, J. Phlorizin: A Review. Diabetes Metab. Res. Rev. 2005, 21, 31–38. [Google Scholar] [CrossRef]
- Von Mering, J. Über Künstlichen Diabetes. Cent. Med. Wiss. 1886, 22, 31–35. [Google Scholar]
- Cai, W.; Jiang, L.; Xie, Y.; Liu, Y.; Liu, W.; Zhao, G. Design of SGLT2 Inhibitors for the Treatment of Type 2 Diabetes: A History Driven by Biology to Chemistry. Med. Chem. 2015, 11, 317–328. [Google Scholar] [CrossRef]
- Braunwald, E. SGLT2 Inhibitors: The Statins of the 21st Century. Eur. Heart J. 2022, 43, 1029–1030. [Google Scholar] [CrossRef]
- Kotas, M.E.; Medzhitov, R. Homeostasis, Inflammation, and Disease Susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libby, P. Inflammatory Mechanisms: The Molecular Basis of Inflammation and Disease. Nutr. Rev. 2007, 65, S140–S146. [Google Scholar] [CrossRef] [PubMed]
- Charlton, A.; Garzarella, J.; Jandeleit-Dahm, K.A.M.; Jha, J.C. Oxidative Stress and Inflammation in Renal and Cardiovascular Complications of Diabetes. Biology 2020, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, F.; Scheen, A.J. Effects of SGLT2 Inhibitors on Systemic and Tissue Low-Grade Inflammation: The Potential Contribution to Diabetes Complications and Cardiovascular Disease. Diabetes Metab. 2018, 44, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Inouye, B.M.; Hughes, F.M.; Jin, H.; Lütolf, R.; Potnis, K.C.; Routh, J.C.; Rouse, D.C.; Foo, W.C.; Todd Purves, J. Diabetic Bladder Dysfunction Is Associated with Bladder Inflammation Triggered through Hyperglycemia, not Polyuria. Res. Rep. Urol. 2018, 10, 219–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.C.; Chang, W.T.; Wu, S.J.; Xu, P.Y.; Ting, N.C.; Liou, C.J. Phloretin and Phlorizin Promote Lipolysis and Inhibit Inflammation in Mouse 3T3-L1 Cells and in Macrophage-Adipocyte Co-Cultures. Mol. Nutr. Food Res. 2013, 57, 1803–1813. [Google Scholar] [CrossRef] [PubMed]
- Liddle, D.M.; Kavanagh, M.E.; Wright, A.J.; Robinson, L.E. Apple Flavonols Mitigate Adipocyte Inflammation and Promote Angiogenic Factors in LPS- and Cobalt Chloride-Stimulated Adipocytes, in Part by a Peroxisome Proliferator-Activated Receptor-γ-Dependent Mechanism. Nutrients 2020, 12, 1386. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Guo, Y.; Xu, L.; Wang, H. Phlorizin Exerts Potent Effects against Aging Induced by D-Galactose in Mice and PC12 Cells. Food Funct. 2021, 12, 2148–2160. [Google Scholar] [CrossRef]
- Liu, D.; Ji, Y.; Guo, Y.; Wang, H.; Wu, Z.; Li, H.; Wang, H. Dietary Supplementation of Apple Phlorizin Attenuates the Redox State Related to Gut Microbiota Homeostasis in C57BL/6J Mice Fed with a High-Fat Diet. J. Agric. Food Chem. 2021, 69, 198–211. [Google Scholar] [CrossRef]
- Nozu, T.; Miyagishi, S.; Ishioh, M.; Takakusaki, K.; Okumura, T. Phlorizin Attenuates Visceral Hypersensitivity and Colonic Hyperpermeability in a Rat Model of Irritable Bowel Syndrome. Biomed. Pharmacother. 2021, 139, 111649. [Google Scholar] [CrossRef]
- Rani, R.; Kumar, A.; Jaggi, A.S.; Singh, N. Pharmacological Investigations on Efficacy of Phlorizin a Sodium-Glucose Co-Transporter (SGLT) Inhibitor in Mouse Model of Intracerebroventricular Streptozotocin Induced Dementia of AD Type. J. Basic Clin. Physiol. Pharmacol. 2021, 32, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Puel, C.; Quintin, A.; Mathey, J.; Obled, C.; Davicco, M.J.; Lebecque, P.; Kati-Coulibaly, S.; Horcajada, M.N.; Coxam, V. Prevention of Bone Loss by Phloridzin, an Apple Polyphenol, in Ovariectomized Rats under Inflammation Conditions. Calcif. Tissue Int. 2005, 77, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Pei, F.; Li, B.-Y.Y.; Zhang, Z.; Yu, F.; Li, X.-L.L.; Lu, W.D.; Cai, Q.; Gao, H.-Q.Q.; Shen, L. Beneficial Effects of Phlorizin on Diabetic Nephropathy in Diabetic Db/Db Mice. J. Diabetes Complicat. 2016, 28, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Zhang, X.; Wang, Z.; Gao, Z.; Liu, G.; Hu, H.; Zou, L.; Li, X. Insulin Sensitivity-Enhancing Activity of Phlorizin Is Associated with Lipopolysaccharide Decrease and Gut Microbiota Changes in Obese and Type 2 Diabetes (Db/Db) Mice. J. Agric. Food Chem. 2016, 64, 7502–7511. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Chen, J.; Yi, K.; Peng, L.; Xie, J.; Gou, X.; Peng, T.; Tang, L. Phlorizin Ameliorates Obesity-Associated Endotoxemia and Insulin Resistance in High-Fat Diet-Fed Mice by Targeting the Gut Microbiota and Intestinal Barrier Integrity. Gut Microbes 2020, 12, 1842990. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Deng, Y.; Xiao, R.; Xu, F.; Li, M.; Gong, Q.; Gao, J. Anti-Fatigue Effect of Phlorizin on Exhaustive Exercise-Induced Oxidative Injury Mediated by Nrf2/ARE Signaling Pathway in Mice. Eur. J. Pharmacol. 2022, 918, 174563. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, J.; Wang, H.; Wang, M.; Zhao, J.; Wu, Z. Protective Effect of Apple Phlorizin on Hydrogen Peroxide-Induced Cell Damage in HepG2 Cells. J. Food Biochem. 2019, 43, e13052. [Google Scholar] [CrossRef]
- Deng, G.; Wang, J.; Zhang, Q.; He, H.; Wu, F.; Feng, T.; Zhou, J.; Zou, K.; Hattori, M. Hepatoprotective Effects of Phloridzin on Hepatic Fibrosis Induced by Carbon Tetrachloride against Oxidative Stress-Triggered Damage and Fibrosis in Rats. Biol. Pharm. Bull. 2012, 35, 1118–1125. [Google Scholar] [CrossRef] [Green Version]
- David-Silva, A.; Esteves, J.V.; Morais, M.R.P.T.; Freitas, H.S.; Zorn, T.M.; Correa-Giannella, M.L.; Machado, U.F. Dual SGLT1/SGLT2 Inhibitor Phlorizin Ameliorates Non-Alcoholic Fatty Liver Disease and Hepatic Glucose Production in Type 2 Diabetic Mice. Diabetes. Metab. Syndr. Obes. 2020, 13, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Antika, L.D.; Lee, E.-J.; Kim, Y.-H.; Kang, M.-K.; Park, S.-H.; Kim, D.Y.; Oh, H.; Choi, Y.-J.; Kang, Y.-H. Dietary Phlorizin Enhances Osteoblastogenic Bone Formation through Enhancing β-Catenin Activity via GSK-3β Inhibition in a Model of Senile Osteoporosis. J. Nutr. Biochem. 2017, 49, 42–52. [Google Scholar] [CrossRef]
- Boccia, M.M.; Kopf, S.R.; Baratti, C.M. Phlorizin, a Competitive Inhibitor of Glucose Transport, Facilitates Memory Storage in Mice. Neurobiol. Learn. Mem. 1999, 71, 104–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Xie, Y.; Wu, H.; Wang, Z.; Li, A.; Li, Z.; Yang, Z.; Zhang, Z.; Xing, Z.; Zhang, X. Phlorizin from Sweet Tea Inhibits the Progress of Esophageal Cancer by Antagonizing the JAK2/STAT3 Signaling Pathway. Oncol. Rep. 2021, 46, 137. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Uzui, H.; Guo, H.; Tada, H. Role of SGLT1 in High Glucose Level-Induced MMP-2 Expression in Human Cardiac Fibroblasts. Mol. Med. Rep. 2018, 17, 6887–6892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.-Y.; Wang, L.-X.; Dong, S.-S.; Hong, Y.; Zhou, X.-H.; Zheng, W.-W.; Zheng, C. Phlorizin Exerts Direct Protective Effects on Palmitic Acid (PA)-Induced Endothelial Dysfunction by Activating the PI3K/AKT/ENOS Signaling Pathway and Increasing the Levels of Nitric Oxide (NO). Med. Sci. Monit. Basic Res. 2018, 24, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Li, B.; Yu, F.; Lu, W.; Zhang, Z.; Yin, M.; Gao, H. Investigation of the Protective Effects of Phlorizin on Diabetic Cardiomyopathy in Db/Db Mice by Quantitative Proteomics. J. Diabetes Res. 2013, 2013, 263845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, M.; Shibazaki, T.; Nakada, T.; Kashihara, T.; Yano, S.; Okamoto, Y.; Isaji, M.; Matsushita, N.; Taira, E.; Yamada, M. Phlorizin Prevents Electrically-Induced Ventricular Tachyarrhythmia during Ischemia in Langendorff-Perfused Guinea-Pig Hearts. Biol. Pharm. Bull. 2014, 37, 1168–1176. [Google Scholar] [CrossRef] [Green Version]
- Gyimesi, G.; Pujol-Giménez, J.; Kanai, Y.; Hediger, M.A. Sodium-Coupled Glucose Transport, the SLC5 Family, and Therapeutically Relevant Inhibitors: From Molecular Discovery to Clinical Application. Pflug. Arch. 2020, 472, 1177–1206. [Google Scholar] [CrossRef]
- Lam, J.T.; Martín, M.G.; Turk, E.; Hirayama, B.A.; Bosshard, N.U.; Steinmann, B.; Wright, E.M. Missense Mutations in SGLT1 Cause Glucose–Galactose Malabsorption by Trafficking Defects. Biochim. Biophys. Acta-Mol. Basis Dis. 1999, 1453, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Ottosson-Laakso, E.; Tuomi, T.; Forsén, B.; Gullström, M.; Groop, P.H.; Groop, L.; Vikman, P. Influence of Familial Renal Glycosuria Due to Mutations in the SLC5A2 Gene on Changes in Glucose Tolerance over Time. PLoS ONE 2016, 11, e0146114. [Google Scholar] [CrossRef]
- Ferrannini, E. Sodium-Glucose Co-Transporters and Their Inhibition: Clinical Physiology. Cell Metab. 2017, 26, 27–38. [Google Scholar] [CrossRef] [Green Version]
- DeFronzo, R.A.; Davidson, J.A.; Del Prato, S. The Role of the Kidneys in Glucose Homeostasis: A New Path towards Normalizing Glycaemia. Diabetes Obes. Metab. 2012, 14, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M. Renal Na(+)-Glucose Cotransporters. Am. J. Physiol. Ren. Physiol. 2001, 280, F10–F18. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Ghani, M.A.; Norton, L.; DeFronzo, R.A. Role of Sodium-Glucose Cotransporter 2 (SGLT 2) Inhibitors in the Treatment of Type 2 Diabetes. Endocr. Rev. 2011, 32, 515–531. [Google Scholar] [CrossRef] [Green Version]
- Isaji, M. SGLT2 Inhibitors: Molecular Design and Potential Differences in Effect. Kidney Int. Suppl. 2011, 1, S14–S19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koepsell, H. Glucose Transporters in the Small Intestine in Health and Disease. Pflug. Arch. 2020, 472, 1207–1248. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Xue, M.; Li, X.; Han, F.; Liu, X.; Xu, L.; Lu, Y.; Cheng, Y.; Li, T.; et al. SGLT2 Inhibition with Empagliflozin Attenuates Myocardial Oxidative Stress and Fibrosis in Diabetic Mice Heart. Cardiovasc. Diabetol. 2019, 18, 15. [Google Scholar] [CrossRef] [PubMed]
- Loo, D.D.F.; Hirayama, B.A.; Meinild, A.K.; Chandy, G.; Zeuthen, T.; Wright, E.M. Passive Water and Ion Transport by Cotransporters. J. Physiol. 1999, 518, 195–202. [Google Scholar] [CrossRef]
- Vallon, V.; Thomson, S.C. Targeting Renal Glucose Reabsorption to Treat Hyperglycaemia: The Pleiotropic Effects of SGLT2 Inhibition. Diabetologia 2017, 60, 215–225. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Hompesch, M.; Kasichayanula, S.; Liu, X.; Hong, Y.; Pfister, M.; Morrow, L.A.; Leslie, B.R.; Boulton, D.W.; Ching, A.; et al. Characterization of Renal Glucose Reabsorption in Response to Dapagliflozin in Healthy Subjects and Subjects with Type 2 Diabetes. Diabetes Care 2013, 36, 3169–3176. [Google Scholar] [CrossRef] [Green Version]
- Osorio, H.; Bautista, R.; Rios, A.; Franco, M.; Arellano, A.; Vargas-Robles, H.; Romo, E.; Escalante, B. Effect of Phlorizin on SGLT2 Expression in the Kidney of Diabetic Rats. J. Nephrol. 2010, 23, 541–546. [Google Scholar]
- Osorio, H.; Bautista, R.; Rios, A.; Franco, M.; Santamaría, J.; Escalante, B. Effect of Treatment with Losartan on Salt Sensitivity and SGLT2 Expression in Hypertensive Diabetic Rats. Diabetes Res. Clin. Pract. 2009, 86, e46–e49. [Google Scholar] [CrossRef]
- Tabatabai, N.M.; Sharma, M.; Blumenthal, S.S.; Petering, D.H. Enhanced Expressions of Sodium–Glucose Cotransporters in the Kidneys of Diabetic Zucker Rats. Diabetes Res. Clin. Pract. 2009, 83, e27–e30. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Watanabe, Y.; Fukuda, K.; Watanabe, M.; Onishi, A.; Ohara, K.; Imai, T.; Koepsell, H.; Muto, S.; Vallon, V.; et al. Unmasking a Sustained Negative Effect of SGLT2 Inhibition on Body Fluid Volume in the Rat. Am. J. Physiol. Ren. Physiol. 2018, 315, F653–F664. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Liao, Y.; Wang, J.; Wen, X.; Shu, L. Potential Impacts of Diabetes Mellitus and Anti-Diabetes Agents on Expressions of Sodium-Glucose Transporters (SGLTs) in Mice. Endocrine 2021, 74, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Albertoni Borghese, M.F.; Majowicz, M.P.; Ortiz, M.C.; del Passalacqua, M.R.; Sterin Speziale, N.B.; Vidal, N.A. Expression and Activity of SGLT2 in Diabetes Induced by Streptozotocin: Relationship with the Lipid Environment. Nephron. Physiol. 2009, 112, p45–p52. [Google Scholar] [CrossRef] [PubMed]
- Osorio, H.; Coronel, I.; Arellano, A.; Franco, M.; Escalante, B.; Bautista, R. Ursodeoxycholic Acid Decreases Sodium-Glucose Cotransporter (SGLT2) Expression and Oxidative Stress in the Kidney of Diabetic Rats. Diabetes Res. Clin. Pract. 2012, 97, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Solini, A.; Rossi, C.; Mazzanti, C.M.; Proietti, A.; Koepsell, H.; Ferrannini, E. Sodium-Glucose Co-Transporter (SGLT)2 and SGLT1 Renal Expression in Patients with Type 2 Diabetes. Diabetes Obes. Metab. 2017, 19, 1289–1294. [Google Scholar] [CrossRef]
- Zhao, S.; Lo, C.-S.; Miyata, K.N.; Ghosh, A.; Zhao, X.-P.; Chenier, I.; Cailhier, J.-F.; Ethier, J.; Lattouf, J.-B.; Filep, J.G.; et al. Overexpression of Nrf2 in Renal Proximal Tubular Cells Stimulates Sodium-Glucose Cotransporter 2 Expression and Exacerbates Dysglycemia and Kidney Injury in Diabetic Mice. Diabetes 2021, 70, 1388–1403. [Google Scholar] [CrossRef]
- Rahmoune, H.; Thompson, P.W.; Ward, J.M.; Smith, C.D.; Hong, G.; Brown, J. Glucose Transporters in Human Renal Proximal Tubular Cells Isolated from the Urine of Patients with Non-Insulin-Dependent Diabetes. Diabetes 2005, 54, 3427–3434. [Google Scholar] [CrossRef] [Green Version]
- Crespy, V.; Aprikian, O.; Morand, C.; Besson, C.; Manach, C.; Demigné, C.; Rémésy, C. Bioavailability of Phloretin and Phloridzin in Rats. J. Nutr. 2001, 131, 3227–3230. [Google Scholar] [CrossRef] [Green Version]
- LeFevre, P.G. Sugar Transport in the Red Blood Cell: Structure-Activity Relationships in Substrates and Antagonists. Pharmacol. Rev. 1961, 13, 39–70. [Google Scholar] [PubMed]
- Maher, F.; Vannucci, S.J.; Simpson, I.A. Glucose Transporter Proteins in Brain. FASEB J. 1994, 8, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, D.; Lin, H.; Jiang, S.; Han, L.; Hou, S.; Lin, S.; Cheng, Z.; Bian, W.; Zhang, X.; et al. Enhanced Oral Bioavailability and Bioefficacy of Phloretin Using Mixed Polymeric Modified Self-Nanoemulsions. Food Sci. Nutr. 2020, 8, 3545–3558. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, M.; Maeda, M.; Hayashi, S.; Mori, Y.; Abe, T. A Missense Mutation in the Na+/Glucose Cotransporter Gene SGLT1 in a Patient with Congenital Glucose-Galactose Malabsorption: Normal Trafficking but Inactivation of the Mutant Protein. Biochim. Biophys. Acta-Mol. Basis Dis. 2001, 1536, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.M.; Ghezzi, C.; Loo, D.D.F. Novel and Unexpected Functions of SGLTs. Physiology 2017, 32, 435. [Google Scholar] [CrossRef]
- Bisignano, P.; Ghezzi, C.; Jo, H.; Polizzi, N.F.; Althoff, T.; Kalyanaraman, C.; Friemann, R.; Jacobson, M.P.; Wright, E.M.; Grabe, M. Inhibitor Binding Mode and Allosteric Regulation of Na+-Glucose Symporters. Nat. Commun. 2018, 9, 5245. [Google Scholar] [CrossRef]
- Oku, A.; Ueta, K.; Arakawa, K.; Ishihara, T.; Nawano, M.; Kuronuma, Y.; Matsumoto, M.; Saito, A.; Tsujihara, K.; Anai, M.; et al. T-1095, an Inhibitor of Renal Na+-Glucose Cotransporters, May Provide a Novel Approach to Treating Diabetes. Diabetes 1999, 48, 1794–1800. [Google Scholar] [CrossRef]
- Adachi, T.; Yasuda, K.; Okamoto, Y.; Shihara, N.; Oku, A.; Ueta, K.; Kitamura, K.; Saito, A.; Iwakura, I.; Yamada, Y.; et al. T-1095, a Renal Na+-Glucose Transporter Inhibitor, Improves Hyperglycemia in Streptozotocin-Induced Diabetic Rats. Metab.-Clin. Exp. 2000, 49, 990–995. [Google Scholar] [CrossRef]
- Katsuno, K.; Fujimori, Y.; Takemura, Y.; Hiratochi, M.; Itoh, F.; Komatsu, Y.; Fujikura, H.; Isaji, M. Sergliflozin, a Novel Selective Inhibitor of Low-Affinity Sodium Glucose Cotransporter (SGLT2), Validates the Critical Role of SGLT2 in Renal Glucose Reabsorption and Modulates Plasma Glucose Level. J. Pharmacol. Exp. Ther. 2007, 320, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, K.; Fujikura, H.; Fushimi, N.; Nishimura, T.; Tatani, K.; Katsuno, K.; Fujimori, Y.; Watanabe, S.; Hiratochi, M.; Nakabayashi, T.; et al. Discovery of Remogliflozin Etabonate: A Potent and Highly Selective SGLT2 Inhibitor. Bioorg. Med. Chem. 2021, 34, 116033. [Google Scholar] [CrossRef]
- Atal, S.; Fatima, Z.; Singh, S.; Balakrishnan, S.; Joshi, R. Remogliflozin: The New Low Cost SGLT-2 Inhibitor for Type 2 Diabetes Mellitus. Diabetol. Int. 2020, 12, 247–253. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency Zynquista. 2019. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/zynquista (accessed on 11 April 2022).
- Osadebe, P.; Odoh, E.; Uzor, P. Natural Products as Potential Sources of Antidiabetic Drugs. Br. J. Pharm. Res. 2014, 4, 2075–2095. [Google Scholar] [CrossRef]
- Fediuk, D.J.; Nucci, G.; Dawra, V.K.; Cutler, D.L.; Amin, N.B.; Terra, S.G.; Boyd, R.A.; Krishna, R.; Sahasrabudhe, V. Overview of the Clinical Pharmacology of Ertugliflozin, a Novel Sodium-Glucose Cotransporter 2 (SGLT2) Inhibitor. Clin. Pharmacokinet. 2020, 59, 949–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, R.M.; Prossler, J.E. Tofogliflozin: First Global Approval. Drugs 2014, 74, 939–944. [Google Scholar] [CrossRef]
- Grempler, R.; Thomas, L.; Eckhardt, M.; Himmelsbach, F.; Sauer, A.; Sharp, D.E.; Bakker, R.A.; Mark, M.; Klein, T.; Eickelmann, P. Empagliflozin, a Novel Selective Sodium Glucose Cotransporter-2 (SGLT-2) Inhibitor: Characterisation and Comparison with Other SGLT-2 Inhibitors. Diabetes Obes. Metab. 2012, 14, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Fala, L. Jardiance (Empagliflozin), an SGLT2 Inhibitor, Receives FDA Approval for the Treatment of Patients with Type 2 Diabetes. Am. Heal. Drug Benefits 2015, 8, 92–95. [Google Scholar]
- U.S. Food and Drug Administration. FDA Approves Jardiance to Reduce Cardiovascular Death in Adults with Type 2 Diabetes. 2016. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-jardiance-reduce-cardiovascular-death-adults-type-2-diabetes (accessed on 9 April 2022).
- Boehringer-Ingelheim US FDA Approves Jardiance® (Empagliflozin) to Treat Adults Living with Heart Failure with Reduced Ejection Fraction. 2021. Available online: https://www.boehringer-ingelheim.us/press-release/us-fda-approves-jardiance-empagliflozin-treat-adults-living-heart-failure-reduced (accessed on 9 April 2022).
- U.S. Food and Drug Administration. FDA Approves Treatment for Wider Range of Patients with Heart Failure. 2022. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-treatment-wider-range-patients-heart-failure (accessed on 9 April 2022).
- Azzam, O.; Carnagarin, R.; Lugo-Gavidia, L.M.; Nolde, J.; Matthews, V.B.; Schlaich, M.P. Bexagliflozin for Type 2 Diabetes: An Overview of the Data. Expert Opin. Pharmacother. 2021, 22, 2095–2103. [Google Scholar] [CrossRef]
- Mascitti, V.; Maurer, T.S.; Robinson, R.P.; Bian, J.; Boustany-Kari, C.M.; Brandt, T.; Collman, B.M.; Kalgutkar, A.S.; Klenotic, M.K.; Leininger, M.T.; et al. Discovery of a Clinical Candidate from the Structurally Unique Dioxa-Bicyclo[3.2.1]Octane Class of Sodium-Dependent Glucose Cotransporter 2 Inhibitors. J. Med. Chem. 2011, 54, 2952–2960. [Google Scholar] [CrossRef]
- Markham, A. Ertugliflozin: First Global Approval. Drugs 2018, 78, 513–519. [Google Scholar] [CrossRef]
- Kakinuma, H.; Oi, T.; Hashimoto-Tsuchiya, Y.; Arai, M.; Kawakita, Y.; Fukasawa, Y.; Iida, I.; Hagima, N.; Takeuchi, H.; Chino, Y.; et al. (1S)-1,5-Anhydro-1-[5-(4-Ethoxybenzyl)-2-Methoxy-4-Methylphenyl]-1-Thio-D-Glucitol (TS-071) Is a Potent, Selective Sodium-Dependent Glucose Cotransporter 2 (SGLT2) Inhibitor for Type 2 Diabetes Treatment. J. Med. Chem. 2010, 53, 3247–3261. [Google Scholar] [CrossRef]
- Markham, A.; Elkinson, S. Luseogliflozin: First Global Approval. Drugs 2014, 74, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Ellsworth, B.A.; Nirschl, A.A.; McCann, P.J.; Patel, M.; Girotra, R.N.; Wu, G.; Sher, P.M.; Morrison, E.P.; Biller, S.A.; et al. Discovery of Dapagliflozin: A Potent, Selective Renal Sodium-Dependent Glucose Cotransporter 2 (SGLT2) Inhibitor for the Treatment of Type 2 Diabetes. J. Med. Chem. 2008, 51, 1145–1149. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency FORXIGATM (Dapagliflozin) Now Approved in European Union for Treatment of Type 2 Diabetes. 2014. Available online: https://www.astrazeneca.com/media-centre/press-releases/2012/FORXIGA-dapagliflozin-now-approved-in-European-Union-for-treatment-of-type-2-diabetes-14112012.html# (accessed on 11 April 2022).
- U.S. Food and Drug Administration. FDA Approves New Treatment for a Type of Heart Failure. 2020. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-new-treatment-type-heart-failure (accessed on 11 April 2022).
- U.S. Food and Drug Administration. FDA Approves Treatment for Chronic Kidney Disease. 2021. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-treatment-chronic-kidney-disease (accessed on 11 April 2022).
- Markham, A. Remogliflozin Etabonate: First Global Approval. Drugs 2019, 79, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Takasu, T.; Yokono, M.; Tahara, A.; Takakura, S. In Vitro Pharmacological Profile of Ipragliflozin, a Sodium Glucose Co-Transporter 2 Inhibitor. Biol. Pharm. Bull. 2019, 42, 507–511. [Google Scholar] [CrossRef] [Green Version]
- Poole, R.M.; Dungo, R.T. Ipragliflozin: First Global Approval. Drugs 2014, 74, 611–617. [Google Scholar] [CrossRef]
- Liang, Y.; Arakawa, K.; Ueta, K.; Matsushita, Y.; Kuriyama, C.; Martin, T.; Du, F.; Liu, Y.; Xu, J.; Conway, B.; et al. Effect of Canagliflozin on Renal Threshold for Glucose, Glycemia, and Body Weight in Normal and Diabetic Animal Models. PLoS ONE 2012, 7, e30555. [Google Scholar] [CrossRef] [Green Version]
- Elkinson, S.; Scott, L.J. Canagliflozin: First Global Approval. Drugs 2013, 73, 979–988. [Google Scholar] [CrossRef]
- Johnson, J.; U.S. FDA. Approves INVOKANA® (Canagliflozin) to Reduce the Risk of Heart Attack, Stroke or Cardiovascular Death in Adults with Type 2 Diabetes and Established Cardiovascular Disease. 2018. Available online: https://www.jnj.com/u-s-fda-approves-invokana-canagliflozin-to-reduce-the-risk-of-heart-attack-stroke-or-cardiovascular-death-in-adults-with-type-2-diabetes-and-established-cardiovascular-disease (accessed on 11 April 2022).
- Johnson, J.; U.S. FDA. Approves INVOKANA® (Canagliflozin) to Treat Diabetic Kidney Disease (DKD) and Reduce the Risk of Hospitalization for Heart Failure in Patients with Type 2 Diabetes (T2D) and DKD. 2019. Available online: https://www.jnj.com/u-s-fda-approves-invokana-canagliflozin-to-treat-diabetic-kidney-disease-dkd-and-reduce-the-risk-of-hospitalization-for-heart-failure-in-patients-with-type-2-diabetes-t2d-and-dkd (accessed on 11 April 2022).
- Hardman, T.C.; Dubrey, S.W. Development and Potential Role of Type-2 Sodium-Glucose Transporter Inhibitors for Management of Type 2 Diabetes. Diabetes Ther. 2011, 2, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Bays, H.E.; Kozlovski, P.; Shao, Q.; Proot, P.; Keefe, D. Licogliflozin, a Novel SGLT1 and 2 Inhibitor: Body Weight Effects in a Randomized Trial in Adults with Overweight or Obesity. Obesity 2020, 28, 870–881. [Google Scholar] [CrossRef] [Green Version]
- Cefalo, C.M.A.; Cinti, F.; Moffa, S.; Impronta, F.; Sorice, G.P.; Mezza, T.; Pontecorvi, A.; Giaccari, A. Sotagliflozin, the First Dual SGLT Inhibitor: Current Outlook and Perspectives. Cardiovasc. Diabetol. 2019, 18, 20. [Google Scholar] [CrossRef] [Green Version]
- Dennis, J.M. Precision Medicine in Type 2 Diabetes: Using Individualized Prediction Models to Optimize Selection of Treatment. Diabetes 2020, 69, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Gloyn, A.L.; Drucker, D.J. Precision Medicine in the Management of Type 2 Diabetes. Lancet Diabetes Endocrinol. 2018, 6, 891–900. [Google Scholar] [CrossRef]
- Kalra, S.; Shetty, K.K.; Nagarajan, V.B.; Ved, J.K. Basic and Clinical Pharmaco-Therapeutics of SGLT2 Inhibitors: A Contemporary Update. Diabetes Ther. 2020, 11, 813–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeoh, S.E.; Docherty, K.F.; Jhund, P.S.; Petrie, M.C.; Inzucchi, S.E.; Køber, L.; Kosiborod, M.N.; Martinez, F.A.; Ponikowski, P.; Sabatine, M.S.; et al. Relationship of Dapagliflozin With Serum Sodium. JACC Hear. Fail. 2022, 10, 306–318. [Google Scholar] [CrossRef]
- Masuda, T.; Muto, S.; Fukuda, K.; Watanabe, M.; Ohara, K.; Koepsell, H.; Vallon, V.; Nagata, D. Osmotic Diuresis by SGLT2 Inhibition Stimulates Vasopressin-Induced Water Reabsorption to Maintain Body Fluid Volume. Physiol. Rep. 2020, 8, e14360. [Google Scholar] [CrossRef] [Green Version]
- Upadhya, B.; Kozak, P.M.; Stacey, R.B.; Vasan, R.S. Newer Drugs to Reduce High Blood Pressure and Mitigate Hypertensive Target Organ Damage. Curr. Hypertens. Rep. 2022, 24, 1–20. [Google Scholar] [CrossRef]
- Saponaro, C.; Mühlemann, M.; Acosta-Montalvo, A.; Piron, A.; Gmyr, V.; Delalleau, N.; Moerman, E.; Thévenet, J.; Pasquetti, G.; Coddeville, A.; et al. Interindividual Heterogeneity of SGLT2 Expression and Function in Human Pancreatic Islets. Diabetes 2020, 69, 902–914. [Google Scholar] [CrossRef]
- Zhu, X.; Lin, C.; Li, L.; Hu, S.; Cai, X.; Ji, L. SGLT2i Increased the Plasma Fasting Glucagon Level in Patients with Diabetes: A Meta-Analysis. Eur. J. Pharmacol. 2021, 903, 174145. [Google Scholar] [CrossRef]
- Verma, S.; Rawat, S.; Ho, K.L.; Wagg, C.S.; Zhang, L.; Teoh, H.; Dyck, J.E.; Uddin, G.M.; Oudit, G.Y.; Mayoux, E.; et al. Empagliflozin Increases Cardiac Energy Production in Diabetes: Novel Translational Insights Into the Heart Failure Benefits of SGLT2 Inhibitors. JACC Basic Transl. Sci. 2018, 3, 575–587. [Google Scholar] [CrossRef]
- Marton, A.; Kaneko, T.; Kovalik, J.P.; Yasui, A.; Nishiyama, A.; Kitada, K.; Titze, J. Organ Protection by SGLT2 Inhibitors: Role of Metabolic Energy and Water Conservation. Nat. Rev. Nephrol. 2021, 17, 65–77. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lee, M.; Lee, J.Y.; Bae, J.; Shin, E.; Lee, Y.H.; Lee, B.W.; Kang, E.S.; Cha, B.S. Ipragliflozin, an SGLT2 Inhibitor, Ameliorates High-Fat Diet-Induced Metabolic Changes by Upregulating Energy Expenditure through Activation of the AMPK/ SIRT1 Pathway. Diabetes Metab. J. 2021, 45, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Wallenius, K.; Kroon, T.; Hagstedt, T.; Löfgren, L.; Sörhede-Winzell, M.; Boucher, J.; Lindén, D.; Oakes, N.D. The SGLT2 Inhibitor Dapagliflozin Promotes Systemic FFA Mobilization, Enhances Hepatic β-Oxidation, and Induces Ketosis. J. Lipid Res. 2022, 63, 100176. [Google Scholar] [CrossRef] [PubMed]
- Aragón-Herrera, A.; Feijóo-Bandín, S.; Otero Santiago, M.; Barral, L.; Campos-Toimil, M.; Gil-Longo, J.; Costa Pereira, T.M.; García-Caballero, T.; Rodríguez-Segade, S.; Rodríguez, J.; et al. Empagliflozin Reduces the Levels of CD36 and Cardiotoxic Lipids While Improving Autophagy in the Hearts of Zucker Diabetic Fatty Rats. Biochem. Pharmacol. 2019, 170, 113677. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Kim, S.H.; Kang, J.M.; Heo, J.H.; Kim, D.J.; Park, S.H.; Sung, M.J.; Kim, J.; Oh, J.; Yang, D.H.; et al. Empagliflozin Attenuates Diabetic Tubulopathy by Improving Mitochondrial Fragmentation and Autophagy. Am. J. Physiol.-Ren. Physiol. 2019, 317, F767–F780. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Kuno, A.; Yano, T.; Miki, T.; Oshima, H.; Sato, T.; Nakata, K.; Kimura, Y.; Tanno, M.; Miura, T. Empagliflozin Normalizes the Size and Number of Mitochondria and Prevents Reduction in Mitochondrial Size after Myocardial Infarction in Diabetic Hearts. Physiol. Rep. 2018, 6, e13741. [Google Scholar] [CrossRef]
- Song, Y.; Huang, C.; Sin, J.; de Germano, J.F.; Taylor, D.J.R.; Thakur, R.; Gottlieb, R.A.; Mentzer, R.M.; Andres, A.M. Attenuation of Adverse Postinfarction Left Ventricular Remodeling with Empagliflozin Enhances Mitochondria-Linked Cellular Energetics and Mitochondrial Biogenesis. Int. J. Mol. Sci. 2021, 23, 437. [Google Scholar] [CrossRef]
- Belosludtsev, K.N.; Starinets, V.S.; Belosludtsev, M.N.; Mikheeva, I.B.; Dubinin, M.V.; Belosludtseva, N.V. Chronic Treatment with Dapagliflozin Protects against Mitochondrial Dysfunction in the Liver of C57BL/6NCrl Mice with High-Fat Diet/Streptozotocin-Induced Diabetes Mellitus. Mitochondrion 2021, 59, 246–254. [Google Scholar] [CrossRef]
- Wei, D.; Liao, L.; Wang, H.; Zhang, W.; Wang, T.; Xu, Z. Canagliflozin Ameliorates Obesity by Improving Mitochondrial Function and Fatty Acid Oxidation via PPARα In Vivo and In Vitro. Life Sci. 2020, 247, 117414. [Google Scholar] [CrossRef]
- Croteau, D.; Luptak, I.; Chambers, J.M.; Hobai, I.; Panagia, M.; Pimentel, D.R.; Siwik, D.A.; Qin, F.; Colucci, W.S. Effects of Sodium-Glucose Linked Transporter 2 Inhibition With Ertugliflozin on Mitochondrial Function, Energetics, and Metabolic Gene Expression in the Presence and Absence of Diabetes Mellitus in Mice. J. Am. Heart Assoc. 2021, 10, e019995. [Google Scholar] [CrossRef]
- Yang, L.; Liang, B.; Li, J.; Zhang, X.; Chen, H.; Sun, J.; Zhang, Z. Dapagliflozin Alleviates Advanced Glycation End Product Induced Podocyte Injury through AMPK/MTOR Mediated Autophagy Pathway. Cell. Signal. 2022, 90, 110206. [Google Scholar] [CrossRef]
- Xu, J.; Kitada, M.; Ogura, Y.; Liu, H.; Koya, D. Dapagliflozin Restores Impaired Autophagy and Suppresses Inflammation in High Glucose-Treated HK-2 Cells. Cells 2021, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Q.; Huang, W.; Han, Y.; Tan, H.; An, M.; Xiang, Q.; Zhou, R.; Yang, L.; Cheng, Y. Dapagliflozin Alleviates Hepatic Steatosis by Restoring Autophagy via the AMPK-MTOR Pathway. Front. Pharmacol. 2021, 12, 1190. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Al-Shorbagy, M.Y.; Saad, M.A. Activation of Autophagy and Suppression of Apoptosis by Dapagliflozin Attenuates Experimental Inflammatory Bowel Disease in Rats: Targeting AMPK/MTOR, HMGB1/RAGE and Nrf2/HO-1 Pathways. Chem. Biol. Interact. 2021, 335, 109368. [Google Scholar] [CrossRef]
- Ala, M.; Khoshdel, M.R.F.; Dehpour, A.R. Empagliflozin Enhances Autophagy, Mitochondrial Biogenesis, and Antioxidant Defense and Ameliorates Renal Ischemia/Reperfusion in Nondiabetic Rats. Oxid. Med. Cell. Longev. 2022, 2022, 1197061. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rafei, M.; Thabet, N.; Rashed, L.; Moustafa, E. Canagliflozin, a SGLT-2 Inhibitor, Relieves ER Stress, Modulates Autophagy and Induces Apoptosis in Irradiated HepG2 Cells: Signal Transduction between PI3K/AKT/GSK-3β/MTOR and Wnt/β-Catenin Pathways; In Vitro. J. Cancer Res. Ther. 2021, 17, 1404–1418. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, W.; Zhong, J.; Lei, F.; Xu, N.; Zhang, Y.; Xie, W. Canagliflozin Exerts Anti-Inflammatory Effects by Inhibiting Intracellular Glucose Metabolism and Promoting Autophagy in Immune Cells. Biochem. Pharmacol. 2018, 152, 45–59. [Google Scholar] [CrossRef]
- Ashrafi Jigheh, Z.; Ghorbani Haghjo, A.; Argani, H.; Roshangar, L.; Rashtchizadeh, N.; Sanajou, D.; Nazari Soltan Ahmad, S.; Rashedi, J.; Dastmalchi, S.; Mesgari Abbasi, M. Empagliflozin Alleviates Renal Inflammation and Oxidative Stress in Streptozotocin-Induced Diabetic Rats Partly by Repressing HMGB1-TLR4 Receptor Axis. Iran. J. Basic Med. Sci. 2019, 22, 384–390. [Google Scholar] [CrossRef]
- Uthman, L.; Li, X.; Baartscheer, A.; Schumacher, C.A.; Baumgart, P.; Hermanides, J.; Preckel, B.; Hollmann, M.W.; Coronel, R.; Zuurbier, C.J.; et al. Empagliflozin Reduces Oxidative Stress through Inhibition of the Novel Inflammation/NHE/[Na+]c/ROS-Pathway in Human Endothelial Cells. Biomed. Pharmacother. 2022, 146, 112515. [Google Scholar] [CrossRef]
- Dasari, D.; Bhat, A.; Mangali, S.; Ghatage, T.; Lahane, G.P.; Sriram, D.; Dhar, A. Canagliflozin and Dapagliflozin Attenuate Glucolipotoxicity-Induced Oxidative Stress and Apoptosis in Cardiomyocytes via Inhibition of Sodium-Glucose Cotransporter-1. ACS Pharmacol. Transl. Sci. 2022, 5, 216–225. [Google Scholar] [CrossRef]
- Hazem, R.M.; Ibrahim, A.Z.; Ali, D.A.; Moustafa, Y.M. Dapagliflozin Improves Steatohepatitis in Diabetic Rats via Inhibition of Oxidative Stress and Inflammation. Int. Immunopharmacol. 2022, 104, 108503. [Google Scholar] [CrossRef]
- Chehrehgosha, H.; Sohrabi, M.R.; Ismail-Beigi, F.; Malek, M.; Reza Babaei, M.; Zamani, F.; Ajdarkosh, H.; Khoonsari, M.; Fallah, A.E.; Khamseh, M.E. Empagliflozin Improves Liver Steatosis and Fibrosis in Patients with Non-Alcoholic Fatty Liver Disease and Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Diabetes Ther. 2021, 12, 843–861. [Google Scholar] [CrossRef] [PubMed]
- Hayashizaki-Someya, Y.; Kurosaki, E.; Takasu, T.; Mitori, H.; Yamazaki, S.; Koide, K.; Takakura, S. Ipragliflozin, an SGLT2 Inhibitor, Exhibits a Prophylactic Effect on Hepatic Steatosis and Fibrosis Induced by Choline-Deficient l-Amino Acid-Defined Diet in Rats. Eur. J. Pharmacol. 2015, 754, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Heerspink, H.J.L.; Perco, P.; Mulder, S.; Leierer, J.; Hansen, M.K.; Heinzel, A.; Mayer, G. Canagliflozin Reduces Inflammation and Fibrosis Biomarkers: A Potential Mechanism of Action for Beneficial Effects of SGLT2 Inhibitors in Diabetic Kidney Disease. Diabetologia 2019, 62, 1154–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.; Zhang, M.; Suo, M.; Liu, D.; Wang, X.; Liu, M.; Pan, J.; Jin, T.; An, F. Dapagliflozin Alleviates Cardiac Fibrosis through Suppressing EndMT and Fibroblast Activation via AMPKα/TGF-β/Smad Signalling in Type 2 Diabetic Rats. J. Cell. Mol. Med. 2021, 25, 7642–7659. [Google Scholar] [CrossRef]
- Abdel-latif, R.G.; Rifaai, R.A.; Amin, E.F. Empagliflozin Alleviates Neuronal Apoptosis Induced by Cerebral Ischemia/Reperfusion Injury through HIF-1α/VEGF Signaling Pathway. Arch. Pharm. Res. 2020, 43, 514–525. [Google Scholar] [CrossRef]
- Tian, G.; Yu, Y.; Deng, H.; Yang, L.; Shi, X.; Yu, B. Empagliflozin Alleviates Ethanol-Induced Cardiomyocyte Injury through Inhibition of Mitochondrial Apoptosis via a SIRT1/PTEN/Akt Pathway. Clin. Exp. Pharmacol. Physiol. 2021, 48, 837–845. [Google Scholar] [CrossRef]
- Nakao, M.; Shimizu, I.; Katsuumi, G.; Yoshida, Y.; Suda, M.; Hayashi, Y.; Ikegami, R.; Hsiao, Y.T.; Okuda, S.; Soga, T.; et al. Empagliflozin Maintains Capillarization and Improves Cardiac Function in a Murine Model of Left Ventricular Pressure Overload. Sci. Rep. 2021, 11, 18384. [Google Scholar] [CrossRef]
- Faridvand, Y.; Kazemzadeh, H.; Vahedian, V.; Mirzajanzadeh, P.; Nejabati, H.R.; Safaie, N.; Maroufi, N.F.; Pezeshkian, M.; Nouri, M.; Jodati, A. Dapagliflozin Attenuates High Glucose-Induced Endothelial Cell Apoptosis and Inflammation through AMPK/SIRT1 Activation. Clin. Exp. Pharmacol. Physiol. 2022. [Google Scholar] [CrossRef]
- Hu, Y.; Xu, Q.; Li, H.; Meng, Z.; Hao, M.; Ma, X.; Lin, W.; Kuang, H. Dapagliflozin Reduces Apoptosis of Diabetic Retina and Human Retinal Microvascular Endothelial Cells Through ERK1/2/CPLA2/AA/ROS Pathway Independent of Hypoglycemic. Front. Pharmacol. 2022, 13, 497. [Google Scholar] [CrossRef]
- Shibusawa, R.; Yamada, E.; Okada, S.; Nakajima, Y.; Bastie, C.C.; Maeshima, A.; Kaira, K.; Yamada, M. Dapagliflozin Rescues Endoplasmic Reticulum Stress-Mediated Cell Death. Sci. Rep. 2019, 9, 9887. [Google Scholar] [CrossRef]
- Wang, C.-C.; Li, Y.; Qian, X.-Q.; Zhao, H.; Wang, D.; Zuo, G.-X.; Wang, K. Empagliflozin Alleviates Myocardial I/R Injury and Cardiomyocyte Apoptosis via Inhibiting ER Stress-Induced Autophagy and the PERK/ATF4/Beclin1 Pathway. J. Drug Target. 2022, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Nasiri-Ansari, N.; Nikolopoulou, C.; Papoutsi, K.; Kyrou, I.; Mantzoros, C.S.; Kyriakopoulos, G.; Chatzigeorgiou, A.; Kalotychou, V.; Randeva, M.S.; Chatha, K.; et al. Empagliflozin Attenuates Non-Alcoholic Fatty Liver Disease (NAFLD) in High Fat Diet Fed ApoE(−/−) Mice by Activating Autophagy and Reducing ER Stress and Apoptosis. Int. J. Mol. Sci. 2021, 22, 818. [Google Scholar] [CrossRef] [PubMed]
- Petito-da-Silva, T.I.; Souza-Mello, V.; Barbosa-da-Silva, S. Empaglifozin Mitigates NAFLD in High-Fat-Fed Mice by Alleviating Insulin Resistance, Lipogenesis and ER Stress. Mol. Cell. Endocrinol. 2019, 498, 110539. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.Y.; Lin, Y.W.; Fisch, S.; Cheng, J.T.; Kang, N.W.; Hong, C.S.; Chen, Z.C.; Chang, W.T. Dapagliflozin Suppresses ER Stress and Improves Subclinical Myocardial Function in Diabetes: From Bedside to Bench. Diabetes 2021, 70, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Uthman, L.; Kuschma, M.; Römer, G.; Boomsma, M.; Kessler, J.; Hermanides, J.; Hollmann, M.W.; Preckel, B.; Zuurbier, C.J.; Weber, N.C. Novel Anti-Inflammatory Effects of Canagliflozin Involving Hexokinase II in Lipopolysaccharide-Stimulated Human Coronary Artery Endothelial Cells. Cardiovasc. Drugs Ther. 2021, 35, 1083–1094. [Google Scholar] [CrossRef]
- Abdollahi, E.; Keyhanfar, F.; Delbandi, A.-A.; Falak, R.; Hajimiresmaiel, S.J.; Shafiei, M. Dapagliflozin Exerts Anti-Inflammatory Effects via Inhibition of LPS-Induced TLR-4 Overexpression and NF-ΚB Activation in Human Endothelial Cells and Differentiated Macrophages. Eur. J. Pharmacol. 2022, 918, 174715. [Google Scholar] [CrossRef]
- Tian, Y.; Gao, L.; Guo, Y.; Xu, Y. Short-Term Phlorizin Treatment Attenuates Adipose Tissue Inflammation without Alerting Obesity in High-Fat Diet Fed Mice. J. Food Biochem. 2017, 41, e12407. [Google Scholar] [CrossRef]
- Kamdi, S.P.; Raval, A.; Nakhate, K.T. Phloridzin Attenuates Lipopolysaccharide-Induced Cognitive Impairment via Antioxidant, Anti-Inflammatory and Neuromodulatory Activities. Cytokine 2021, 139, 155408. [Google Scholar] [CrossRef]
- Chang, W.T.; Huang, W.C.; Liou, C.J. Evaluation of the Anti-Inflammatory Effects of Phloretin and Phlorizin in Lipopolysaccharide-Stimulated Mouse Macrophages. Food Chem. 2012, 134, 972–979. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutierrez-Grijalva, E.; Ambriz-Perez, D.; Heredia, J. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef]
- Jasim, H.A.; Nahar, L.; Jasim, M.A.; Moore, S.A.; Ritchie, K.J.; Sarker, S.D. Chalcones: Synthetic Chemistry Follows Where Nature Leads. Biomolecules 2021, 11, 1203. [Google Scholar] [CrossRef] [PubMed]
- Nile, S.H.; Keum, Y.S.; Nile, A.S.; Jalde, S.S.; Patel, R.V. Antioxidant, Anti-Inflammatory, and Enzyme Inhibitory Activity of Natural Plant Flavonoids and Their Synthesized Derivatives. J. Biochem. Mol. Toxicol. 2018, 32, e22002. [Google Scholar] [CrossRef] [PubMed]
- De Nardo, D.; Latz, E. NLRP3 Inflammasomes Link Inflammation and Metabolic Disease. Trends Immunol. 2011, 32, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory Mechanisms Linking Obesity and Metabolic Disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549. [Google Scholar] [CrossRef]
- Blüher, M. Adipose Tissue Inflammation: A Cause or Consequence of Obesity-Related Insulin Resistance? Clin. Sci. 2016, 130, 1603–1614. [Google Scholar] [CrossRef]
- Mastrocola, R.; Aragno, M.; Alloatti, G.; Collino, M.; Penna, C.; Pagliaro, P. Metaflammation: Tissue-Specific Alterations of the NLRP3 Inflammasome Platform in Metabolic Syndrome. Curr. Med. Chem. 2017, 25, 1294–1310. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, Metaflammation and Immunometabolic Disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Scheen, A.J. Sodium-Glucose Cotransporter Type 2 Inhibitors for the Treatment of Type 2 Diabetes Mellitus. Nat. Rev. Endocrinol. 2020, 16, 556–577. [Google Scholar] [CrossRef]
- Wang, X.; Ni, J.; Guo, R.; Li, L.; Su, J.; He, F.; Fan, G. SGLT2 Inhibitors Break the Vicious Circle between Heart Failure and Insulin Resistance: Targeting Energy Metabolism. Heart Fail. Rev. 2021, 27, 961–980. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhou, H. SGLT2 Inhibitors: A Novel Player in the Treatment and Prevention of Diabetic Cardiomyopathy. Drug Des. Devel. Ther. 2020, 14, 4775. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Reeves, W.B.; Awad, A.S. Pathophysiology of Diabetic Kidney Disease: Impact of SGLT2 Inhibitors. Nat. Rev. Nephrol. 2021, 17, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Gharaibeh, N.E.; Rahhal, M.-N.; Rahimi, L.; Ismail-Beigi, F. SGLT-2 Inhibitors as Promising Therapeutics for Non-Alcoholic Fatty Liver Disease: Pathophysiology, Clinical Outcomes, and Future Directions. Diabetes Metab. Syndr. Obes. 2019, 12, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Alexander, M.; Misharin, A.V.; Budinger, G.R.S. The Role of Macrophages in the Resolution of Inflammation. J. Clin. Investig. 2019, 129, 2619–2628. [Google Scholar] [CrossRef] [Green Version]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Kuzmich, N.; Sivak, K.; Chubarev, V.; Porozov, Y.; Savateeva-Lyubimova, T.; Peri, F. TLR4 Signaling Pathway Modulators as Potential Therapeutics in Inflammation and Sepsis. Vaccines 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.; Martinez, F.O. Alternative Activation of Macrophages: Mechanism and Functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-M.; Chang, N.-C.; Lin, S.-Z. Dapagliflozin, a Selective SGLT2 Inhibitor, Attenuated Cardiac Fibrosis by Regulating the Macrophage Polarization via STAT3 Signaling in Infarcted Rat Hearts. Free Radic. Biol. Med. 2017, 104, 298–310. [Google Scholar] [CrossRef]
- Leng, W.; Ouyang, X.; Lei, X.; Wu, M.; Chen, L.; Wu, Q.; Deng, W.; Liang, Z. The SGLT-2 Inhibitor Dapagliflozin Has a Therapeutic Effect on Atherosclerosis in Diabetic ApoE −/− Mice. Mediat. Inflamm. 2016, 2016, 6305735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, M.; Hiromura, M.; Mori, Y.; Kohashi, K.; Nagashima, M.; Kushima, H.; Watanabe, T.; Hirano, T. Amelioration of Hyperglycemia with a Sodium-Glucose Cotransporter 2 Inhibitor Prevents Macrophage-Driven Atherosclerosis through Macrophage Foam Cell Formation Suppression in Type 1 and Type 2 Diabetic Mice. PLoS ONE 2015, 10, e0143396. [Google Scholar] [CrossRef] [PubMed]
- Gaspari, T.; Spizzo, I.; Liu, H.B.; Hu, Y.; Simpson, R.W.; Widdop, R.E.; Dear, A.E. Dapagliflozin Attenuates Human Vascular Endothelial Cell Activation and Induces Vasorelaxation: A Potential Mechanism for Inhibition of Atherogenesis. Diabetes Vasc. Dis. Res. 2018, 15, 64–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.G.; Lee, S.J.; Lee, J.J.; Kim, J.S.; Lee, O.H.; Kim, C.K.; Kim, D.; Lee, Y.H.; Oh, J.; Park, S.; et al. Anti-Inflammatory Effect for Atherosclerosis Progression by Sodium-Glucose Cotransporter 2 (SGLT-2) Inhibitor in a Normoglycemic Rabbit Model. Korean Circ. J. 2020, 50, 443–457. [Google Scholar] [CrossRef]
- Kohlmorgen, C.; Gerfer, S.; Feldmann, K.; Twarock, S.; Hartwig, S.; Lehr, S.; Klier, M.; Krüger, I.; Helten, C.; Keul, P.; et al. Dapagliflozin Reduces Thrombin Generation and Platelet Activation: Implications for Cardiovascular Risk Reduction in Type 2 Diabetes Mellitus. Diabetologia 2021, 64, 1834–1849. [Google Scholar] [CrossRef]
- Stelmaszyk, A.; Wesołowska, A.; Pomieczyńska, K.; Iskakova, S.; Frydrychowicz, M.; Dworacki, G.; Dworacka, M. The Impact of Dapagliflozin on Glucose Excursions Related to Early Proatherogenic Derangement in the Aortic Wall. Saudi Pharm. J. 2018, 26, 1192–1198. [Google Scholar] [CrossRef]
- Liu, H.; Wei, P.; Fu, W.; Xia, C.; Li, Y.; Tian, K.; Li, Y.; Cheng, D.; Sun, J.; Xu, Y.; et al. Dapagliflozin Ameliorates the Formation and Progression of Experimental Abdominal Aortic Aneurysms by Reducing Aortic Inflammation in Mice. Oxid. Med. Cell. Longev. 2022, 2022, 8502059. [Google Scholar] [CrossRef]
- Leng, W.; Wu, M.; Pan, H.; Lei, X.; Chen, L.; Wu, Q.; Ouyang, X.; Liang, Z. The SGLT2 Inhibitor Dapagliflozin Attenuates the Activity of ROS-NLRP3 Inflammasome Axis in Steatohepatitis with Diabetes Mellitus. Ann. Transl. Med. 2019, 7, 429. [Google Scholar] [CrossRef]
- Tang, L.; Wu, Y.; Tian, M.; Sjöström, C.D.; Johansson, U.; Peng, X.R.; Smith, D.M.; Huang, Y. Dapagliflozin Slows the Progression of the Renal and Liver Fibrosis Associated with Type 2 Diabetes. Am. J. Physiol.-Endocrinol. Metab. 2017, 313, E563–E576. [Google Scholar] [CrossRef]
- Terami, N.; Ogawa, D.; Tachibana, H.; Hatanaka, T.; Wada, J.; Nakatsuka, A.; Eguchi, J.; Sato Horiguchi, C.; Nishii, N.; Yamada, H.; et al. Long-Term Treatment with the Sodium Glucose Cotransporter 2 Inhibitor, Dapagliflozin, Ameliorates Glucose Homeostasis and Diabetic Nephropathy in Db/Db Mice. PLoS ONE 2014, 9, e100777. [Google Scholar] [CrossRef]
- Hatanaka, T.; Ogawa, D.; Tachibana, H.; Eguchi, J.; Inoue, T.; Yamada, H.; Takei, K.; Makino, H.; Wada, J. Inhibition of SGLT2 Alleviates Diabetic Nephropathy by Suppressing High Glucose-Induced Oxidative Stress in Type 1 Diabetic Mice. Pharmacol. Res. Perspect. 2016, 4, e00239. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.J.; Chung, S.; Kim, S.J.; Lee, E.M.; Yoo, Y.H.; Kim, J.W.; Ahn, Y.B.; Kim, E.S.; Moon, S.D.; Kim, M.J.; et al. Effect of Sodium-Glucose Co-Transporter 2 Inhibitor, Dapagliflozin, on Renal Renin-Angiotensin System in an Animal Model of Type 2 Diabetes. PLoS ONE 2016, 11, e0165703. [Google Scholar] [CrossRef] [PubMed]
- Cassis, P.; Locatelli, M.; Cerullo, D.; Corna, D.; Buelli, S.; Zanchi, C.; Villa, S.; Morigi, M.; Remuzzi, G.; Benigni, A.; et al. SGLT2 Inhibitor Dapagliflozin Limits Podocyte Damage in Proteinuric Nondiabetic Nephropathy. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Song, C.; Zeng, Y.; Li, Y.; Li, H.; Liu, B.; Dai, M.; Pan, P. Canagliflozin Alleviates LPS-Induced Acute Lung Injury by Modulating Alveolar Macrophage Polarization. Int. Immunopharmacol. 2020, 88, 106969. [Google Scholar] [CrossRef] [PubMed]
- Shiba, K.; Tsuchiya, K.; Komiya, C.; Miyachi, Y.; Mori, K.; Shimazu, N.; Yamaguchi, S.; Ogasawara, N.; Katoh, M.; Itoh, M.; et al. Canagliflozin, an SGLT2 Inhibitor, Attenuates the Development of Hepatocellular Carcinoma in a Mouse Model of Human NASH. Sci. Rep. 2018, 8, 2362. [Google Scholar] [CrossRef] [Green Version]
- Naznin, F.; Sakoda, H.; Okada, T.; Tsubouchi, H.; Waise, T.M.Z.; Arakawa, K.; Nakazato, M. Canagliflozin, a Sodium Glucose Cotransporter 2 Inhibitor, Attenuates Obesity-Induced Inflammation in the Nodose Ganglion, Hypothalamus, and Skeletal Muscle of Mice. Eur. J. Pharmacol. 2017, 794, 37–44. [Google Scholar] [CrossRef]
- Nasiri-Ansari, Ν.; Dimitriadis, G.K.; Agrogiannis, G.; Perrea, D.; Kostakis, I.D.; Kaltsas, G.; Papavassiliou, A.G.; Randeva, H.S.; Kassi, E. Canagliflozin Attenuates the Progression of Atherosclerosis and Inflammation Process in APOE Knockout Mice. Cardiovasc. Diabetol. 2018, 17, 106. [Google Scholar] [CrossRef] [Green Version]
- Woods, T.C.; Satou, R.; Miyata, K.; Katsurada, A.; Dugas, C.M.; Klingenberg, N.C.; Fonseca, V.A.; Navar, L.G. Canagliflozin Prevents Intrarenal Angiotensinogen Augmentation and Mitigates Kidney Injury and Hypertension in Mouse Model of Type 2 Diabetes Mellitus. Am. J. Nephrol. 2019, 49, 331–342. [Google Scholar] [CrossRef]
- Inoue, M.K.; Matsunaga, Y.; Nakatsu, Y.; Yamamotoya, T.; Ueda, K.; Kushiyama, A.; Sakoda, H.; Fujishiro, M.; Ono, H.; Iwashita, M.; et al. Possible Involvement of Normalized Pin1 Expression Level and AMPK Activation in the Molecular Mechanisms Underlying Renal Protective Effects of SGLT2 Inhibitors in Mice. Diabetol. Metab. Syndr. 2019, 11, 57. [Google Scholar] [CrossRef]
- Pennig, J.; Scherrer, P.; Gissler, M.C.; Anto-Michel, N.; Hoppe, N.; Füner, L.; Härdtner, C.; Stachon, P.; Wolf, D.; Hilgendorf, I.; et al. Glucose Lowering by SGLT2-Inhibitor Empagliflozin Accelerates Atherosclerosis Regression in Hyperglycemic STZ-Diabetic Mice. Sci. Rep. 2019, 9, 17937. [Google Scholar] [CrossRef]
- Lee, N.; Heo, Y.J.; Choi, S.-E.E.; Jeon, J.Y.; Han, S.J.; Kim, D.J.; Kang, Y.; Lee, K.W.; Kim, H.J. Anti-Inflammatory Effects of Empagliflozin and Gemigliptin on LPS-Stimulated Macrophage via the IKK/NF-ΚB, MKK7/JNK, and JAK2/STAT1 Signalling Pathways. J. Immunol. Res. 2021, 2021, 9944880. [Google Scholar] [CrossRef]
- Koyani, C.N.; Plastira, I.; Sourij, H.; Hallström, S.; Schmidt, A.; Rainer, P.P.; Bugger, H.; Frank, S.; Malle, E.; von Lewinski, D. Empagliflozin Protects Heart from Inflammation and Energy Depletion via AMPK Activation. Pharmacol. Res. 2020, 158, 104870. [Google Scholar] [CrossRef] [PubMed]
- Ganbaatar, B.; Fukuda, D.; Shinohara, M.; Yagi, S.; Kusunose, K.; Yamada, H.; Soeki, T.; Hirata, K.-I.; Sata, M. Empagliflozin Ameliorates Endothelial Dysfunction and Suppresses Atherogenesis in Diabetic Apolipoprotein E-Deficient Mice. Eur. J. Pharmacol. 2020, 875, 173040. [Google Scholar] [CrossRef]
- Ortega, R.; Collado, A.; Selles, F.; Gonzalez-Navarro, H.; Sanz, M.J.; Real, J.T.; Piqueras, L. SGLT-2 (Sodium-Glucose Cotransporter 2) Inhibition Reduces Ang II (Angiotensin II)-Induced Dissecting Abdominal Aortic Aneurysm in ApoE (Apolipoprotein E) Knockout Mice. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1614–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radlinger, B.; Hornsteiner, F.; Folie, S.; Salvenmoser, W.; Haubner, B.J.; Schuetz, T.; Haas, S.; Ress, C.; Adolph, T.E.; Salzmann, K.; et al. Cardioprotective Effects of Short-Term Empagliflozin Treatment in Db/Db Mice. Sci. Rep. 2020, 10, 19686. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Koibuchi, N.; Hasegawa, Y.; Sueta, D.; Toyama, K.; Uekawa, K.; Ma, M.; Nakagawa, T.; Kusaka, H.; Kim-Mitsuyama, S. Glycemic Control with Empagliflozin, a Novel Selective SGLT2 Inhibitor, Ameliorates Cardiovascular Injury and Cognitive Dysfunction in Obese and Type 2 Diabetic Mice. Cardiovasc. Diabetol. 2014, 13, 148. [Google Scholar] [CrossRef] [Green Version]
- Byrne, N.J.; Matsumura, N.; Maayah, Z.H.; Ferdaoussi, M.; Takahara, S.; Darwesh, A.M.; Levasseur, J.L.; Jahng, J.W.S.; Vos, D.; Parajuli, N.; et al. Empagliflozin Blunts Worsening Cardiac Dysfunction Associated With Reduced NLRP3 (Nucleotide-Binding Domain-Like Receptor Protein 3) Inflammasome Activation in Heart Failure. Circ. Heart Fail. 2020, 13, e006277. [Google Scholar] [CrossRef]
- Kräker, K.; Herse, F.; Golic, M.; Reichhart, N.; Crespo-Garcia, S.; Strauß, O.; Grune, J.; Kintscher, U.; Ebrahim, M.; Bader, M.; et al. Effects of Empagliflozin and Target-Organ Damage in a Novel Rodent Model of Heart Failure Induced by Combined Hypertension and Diabetes. Sci. Rep. 2020, 10, 14061. [Google Scholar] [CrossRef]
- Zhang, Q.-Q.; Li, G.-Q.; Zhong, Y.; Wang, J.; Wang, A.-N.; Zhou, X.; Mao, X.-M. Empagliflozin Improves Chronic Hypercortisolism-Induced Abnormal Myocardial Structure and Cardiac Function in Mice. Ther. Adv. Chronic Dis. 2020, 11, 2040622320974833. [Google Scholar] [CrossRef]
- Meng, Z.; Liu, X.; Li, T.; Fang, T.; Cheng, Y.; Han, L.; Sun, B.; Chen, L. The SGLT2 Inhibitor Empagliflozin Negatively Regulates IL-17/IL-23 Axis-Mediated Inflammatory Responses in T2DM with NAFLD via the AMPK/MTOR/Autophagy Pathway. Int. Immunopharmacol. 2021, 94, 107492. [Google Scholar] [CrossRef]
- Lee, N.; Heo, Y.J.; Choi, S.E.; Jeon, J.Y.; Han, S.J.; Kim, D.J.; Kang, Y.; Lee, K.W.; Kim, H.J. Hepatoprotective Effects of Gemigliptin and Empagliflozin in a Murine Model of Diet-Induced Non-Alcoholic Fatty Liver Disease. Biochem. Biophys. Res. Commun. 2022, 588, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Jojima, T.; Tomotsune, T.; Iijima, T.; Akimoto, K.; Suzuki, K.; Aso, Y. Empagliflozin (an SGLT2 Inhibitor), Alone or in Combination with Linagliptin (a DPP-4 Inhibitor), Prevents Steatohepatitis in a Novel Mouse Model of Non-Alcoholic Steatohepatitis and Diabetes. Diabetol. Metab. Syndr. 2016, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojima, A.; Matsui, T.; Nishino, Y.; Nakamura, N.; Yamagishi, S. Empagliflozin, an Inhibitor of Sodium-Glucose Cotransporter 2 Exerts Anti-Inflammatory and Antifibrotic Effects on Experimental Diabetic Nephropathy Partly by Suppressing AGEs-Receptor Axis. Horm. Metab. Res. 2015, 47, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Hudkins, K.L.; Li, X.; Holland, A.L.; Swaminathan, S.; Alpers, C.E. Regression of Diabetic Nephropathy by Treatment with Empagliflozin in BTBR Ob/Ob Mice. Nephrol. Dial. Transplant. 2022, 37, 847–859. [Google Scholar] [CrossRef]
- Maayah, Z.H.; Ferdaoussi, M.; Takahara, S.; Soni, S.; Dyck, J.R.B. Empagliflozin Suppresses Inflammation and Protects against Acute Septic Renal Injury. Inflammopharmacology 2021, 29, 269–279. [Google Scholar] [CrossRef]
- Castoldi, G.; Carletti, R.; Ippolito, S.; Colzani, M.; Barzaghi, F.; Stella, A.; Zerbini, G.; Perseghin, G.; Zatti, G.; di Gioia, C.R.T. Sodium-Glucose Cotransporter 2 Inhibition Prevents Renal Fibrosis in Cyclosporine Nephropathy. Acta Diabetol. 2021, 58, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Oh, T.J.; Lee, G.; Maeng, H.J.; Lee, D.H.; Kim, K.M.; Choi, S.H.; Jang, H.C.; Lee, H.S.; Park, K.S.; et al. The Beneficial Effects of Empagliflozin, an SGLT2 Inhibitor, on Atherosclerosis in ApoE −/− Mice Fed a Western Diet. Diabetologia 2017, 60, 364–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Nagata, N.; Nagashimada, M.; Zhuge, F.; Ni, Y.; Chen, G.; Mayoux, E.; Kaneko, S.; Ota, T. SGLT2 Inhibition by Empagliflozin Promotes Fat Utilization and Browning and Attenuates Inflammation and Insulin Resistance by Polarizing M2 Macrophages in Diet-Induced Obese Mice. EBioMedicine 2017, 20, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Nagata, N.; Chen, G.; Nagashimada, M.; Zhuge, F.; Ni, Y.; Sakai, Y.; Kaneko, S.; Ota, T. Empagliflozin Reverses Obesity and Insulin Resistance through Fat Browning and Alternative Macrophage Activation in Mice Fed a High-Fat Diet. BMJ Open Diabetes Res. Care 2019, 7, e000783. [Google Scholar] [CrossRef]
- Nakatsu, Y.; Kokubo, H.; Bumdelger, B.; Yoshizumi, M.; Yamamotoya, T.; Matsunaga, Y.; Ueda, K.; Inoue, Y.; Inoue, M.-K.; Fujishiro, M.; et al. The SGLT2 Inhibitor Luseogliflozin Rapidly Normalizes Aortic MRNA Levels of Inflammation-Related but Not Lipid-Metabolism-Related Genes and Suppresses Atherosclerosis in Diabetic ApoE KO Mice. Int. J. Mol. Sci. 2017, 18, 1704. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Tsuchiya, K.; Nakamura, S.; Miyachi, Y.; Shiba, K.; Ogawa, Y.; Kitamura, K. Ipragliflozin-Induced Adipose Expansion Inhibits Cuff-Induced Vascular Remodeling in Mice. Cardiovasc. Diabetol. 2019, 18, 83. [Google Scholar] [CrossRef] [PubMed]
- Salim, H.M.; Fukuda, D.; Yagi, S.; Soeki, T.; Shimabukuro, M.; Sata, M. Glycemic Control with Ipragliflozin, a Novel Selective SGLT2 Inhibitor, Ameliorated Endothelial Dysfunction in Streptozotocin-Induced Diabetic Mouse. Front. Cardiovasc. Med. 2016, 3, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, Y.; Terasaki, M.; Hiromura, M.; Saito, T.; Kushima, H.; Koshibu, M.; Osaka, N.; Ohara, M.; Fukui, T.; Ohtaki, H.; et al. Luseogliflozin Attenuates Neointimal Hyperplasia after Wire Injury in High-Fat Diet-Fed Mice via Inhibition of Perivascular Adipose Tissue Remodeling. Cardiovasc. Diabetol. 2019, 18, 143. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, Y.; Tsuchiya, K.; Shiba, K.; Mori, K.; Komiya, C.; Ogasawara, N.; Ogawa, Y. A Reduced M1-like/M2-like Ratio of Macrophages in Healthy Adipose Tissue Expansion during SGLT2 Inhibition. Sci. Rep. 2018, 8, 16113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komiya, C.; Tsuchiya, K.; Shiba, K.; Miyachi, Y.; Furuke, S.; Shimazu, N.; Yamaguchi, S.; Kanno, K.; Ogawa, Y. Ipragliflozin Improves Hepatic Steatosis in Obese Mice and Liver Dysfunction in Type 2 Diabetic Patients Irrespective of Body Weight Reduction. PLoS ONE 2016, 11, e0151511. [Google Scholar] [CrossRef] [Green Version]
- Okuma, H.; Mori, K.; Nakamura, S.; Sekine, T.; Ogawa, Y.; Tsuchiya, K. Ipragliflozin Ameliorates Diabetic Nephropathy Associated with Perirenal Adipose Expansion in Mice. Int. J. Mol. Sci. 2021, 22, 7329. [Google Scholar] [CrossRef]
- The Human Protein Atlas Single Cell Type—NLRP3. Available online: https://www.proteinatlas.org/ENSG00000162711-NLRP3/single+cell+type (accessed on 28 April 2022).
- Marcuzzi, A.; Melloni, E.; Zauli, G.; Romani, A.; Secchiero, P.; Maximova, N.; Rimondi, E. Autoinflammatory Diseases and Cytokine Storms-Imbalances of Innate and Adaptative Immunity. Int. J. Mol. Sci. 2021, 22, 11241. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 Inflammasome: Molecular Activation and Regulation to Therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Burdette, B.E.; Esparza, A.N.; Zhu, H.; Wang, S. Gasdermin D in Pyroptosis. Acta Pharm. Sin. B 2021, 11, 2768–2782. [Google Scholar] [CrossRef]
- Lee, H.M.; Kim, J.J.; Kim, H.J.; Shong, M.; Ku, B.J.; Jo, E.K. Upregulated NLRP3 Inflammasome Activation in Patients With Type 2 Diabetes. Diabetes 2013, 62, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Abbate, A.; Toldo, S.; Marchetti, C.; Kron, J.; Van Tassell, B.W.; Dinarello, C.A. Interleukin-1 and the Inflammasome as Therapeutic Targets in Cardiovascular Disease. Circ. Res. 2020, 126, 1260–1280. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Meng, X.-F.; Zhang, C. NLRP3 Inflammasome in Metabolic-Associated Kidney Diseases: An Update. Front. Immunol. 2021, 12, 714340. [Google Scholar] [CrossRef] [PubMed]
- Al Mamun, A.; Akter, A.; Hossain, S.; Sarker, T.; Safa, S.A.; Mustafa, Q.G.; Muhammad, S.A.; Munir, F. Role of NLRP3 Inflammasome in Liver Disease. J. Dig. Dis. 2020, 21, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Bajaj, M.; Yang, H.C.; Perez-Polo, J.R.; Birnbaum, Y. SGLT-2 Inhibition with Dapagliflozin Reduces the Activation of the Nlrp3/ASC Inflammasome and Attenuates the Development of Diabetic Cardiomyopathy in Mice with Type 2 Diabetes. Further Augmentation of the Effects with Saxagliptin, a DPP4 Inhibitor. Cardiovasc. Drugs Ther. 2017, 31, 119–132. [Google Scholar] [CrossRef] [PubMed]
- El-Rous, M.A.; Saber, S.; Raafat, E.M.; Ahmed, A.A.E. Dapagliflozin, an SGLT2 Inhibitor, Ameliorates Acetic Acid-Induced Colitis in Rats by Targeting NFκB/AMPK/NLRP3 Axis. Inflammopharmacology 2021, 29, 1169–1185. [Google Scholar] [CrossRef]
- Muhammad, R.N.; Ahmed, L.A.; Abdul Salam, R.M.; Ahmed, K.A.; Attia, A.S. Crosstalk Among NLRP3 Inflammasome, ET B R Signaling, and MiRNAs in Stress-Induced Depression-Like Behavior: A Modulatory Role for SGLT2 Inhibitors. Neurotherapeutics 2021, 18, 2664–2681. [Google Scholar] [CrossRef]
- Ke, Q.; Shi, C.; Lv, Y.; Wang, L.; Luo, J.; Jiang, L.; Yang, J.; Zhou, Y. SGLT2 Inhibitor Counteracts NLRP3 Inflammasome via Tubular Metabolite Itaconate in Fibrosis Kidney. FASEB J. 2022, 36, e22078. [Google Scholar] [CrossRef]
- Birnbaum, Y.; Bajaj, M.; Yang, H.C.; Ye, Y. Combined SGLT2 and DPP4 Inhibition Reduces the Activation of the Nlrp3/ASC Inflammasome and Attenuates the Development of Diabetic Nephropathy in Mice with Type 2 Diabetes. Cardiovasc. Drugs Ther. 2018, 32, 135–145. [Google Scholar] [CrossRef]
- Niu, Y.; Zhang, Y.; Zhang, W.; Lu, J.; Chen, Y.; Hao, W.; Zhou, J.; Wang, L.; Xie, W. Canagliflozin Ameliorates NLRP3 Inflammasome-Mediated Inflammation through Inhibiting NF-ΚB Signaling and Upregulating Bif-1. Front. Pharmacol. 2022, 13, 820541. [Google Scholar] [CrossRef]
- Kim, S.R.; Lee, S.-G.; Kim, S.H.; Kim, J.H.; Choi, E.; Cho, W.; Rim, J.H.; Hwang, I.; Lee, C.J.; Lee, M.; et al. SGLT2 Inhibition Modulates NLRP3 Inflammasome Activity via Ketones and Insulin in Diabetes with Cardiovascular Disease. Nat. Commun. 2020, 11, 2127. [Google Scholar] [CrossRef]
- Sukhanov, S.; Higashi, Y.; Yoshida, T.; Mummidi, S.; Aroor, A.R.; Jeffrey Russell, J.; Bender, S.B.; DeMarco, V.G.; Chandrasekar, B. The SGLT2 Inhibitor Empagliflozin Attenuates Interleukin-17A-Induced Human Aortic Smooth Muscle Cell Proliferation and Migration by Targeting TRAF3IP2/ROS/NLRP3/Caspase-1-Dependent IL-1β and IL-18 Secretion. Cell. Signal. 2021, 77, 109825. [Google Scholar] [CrossRef] [PubMed]
- Deguine, J.; Barton, G.M. MyD88: A Central Player in Innate Immune Signaling. F1000Prime Rep. 2014, 6, 97. [Google Scholar] [CrossRef]
- Quagliariello, V.; De Laurentiis, M.; Rea, D.; Barbieri, A.; Monti, M.G.; Carbone, A.; Paccone, A.; Altucci, L.; Conte, M.; Canale, M.L.; et al. The SGLT-2 Inhibitor Empagliflozin Improves Myocardial Strain, Reduces Cardiac Fibrosis and pro-Inflammatory Cytokines in Non-Diabetic Mice Treated with Doxorubicin. Cardiovasc. Diabetol. 2021, 20, 150. [Google Scholar] [CrossRef] [PubMed]
- Benetti, E.; Mastrocola, R.; Vitarelli, G.; Cutrin, J.C.; Nigro, D.; Chiazza, F.; Mayoux, E.; Collino, M.; Fantozzi, R. Empagliflozin Protects against Diet-Induced NLRP-3 Inflammasome Activation and Lipid Accumulation. J. Pharmacol. Exp. Ther. 2016, 359, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhang, Z.; Wang, J.; Zhang, X.; Yu, X.; Li, Y. Empagliflozin Protects Diabetic Pancreatic Tissue from Damage by Inhibiting the Activation of the NLRP3/Caspase-1/GSDMD Pathway in Pancreatic β Cells: In Vitro and in Vivo Studies. Bioengineered 2021, 12, 9356–9366. [Google Scholar] [CrossRef]
- Xue, M.; Li, T.; Wang, Y.; Chang, Y.; Cheng, Y.; Lu, Y.; Liu, X.; Xu, L.; Li, X.; Yu, X.; et al. Empagliflozin Prevents Cardiomyopathy via SGC-CGMP-PKG Pathway in Type 2 Diabetes Mice. Clin. Sci. 2019, 133, 1705–1720. [Google Scholar] [CrossRef] [Green Version]



| SGLT1 | SGLT2 | |
|---|---|---|
| Glucose affinity | High (Km~0.5–2 mM) | Low (Km~2–5 mM) |
| Glucose transport capacity | Low (2 nmol/mg·min) | High (10 nmol/mg·min) |
| Renal expression | S3 segment | S1 and S2 segments |
| Renal glucose reabsorption | 3–10% | 90–97% |
| Na+/glucose stoichiometry | 2:1 | 1:1 |
| Expression in the small intestine | Yes | No |
| Main function | Intestinal absorption of glucose and galactose (urea and water) | Renal reabsorption of glucose |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feijóo-Bandín, S.; Aragón-Herrera, A.; Otero-Santiago, M.; Anido-Varela, L.; Moraña-Fernández, S.; Tarazón, E.; Roselló-Lletí, E.; Portolés, M.; Gualillo, O.; González-Juanatey, J.R.; et al. Role of Sodium-Glucose Co-Transporter 2 Inhibitors in the Regulation of Inflammatory Processes in Animal Models. Int. J. Mol. Sci. 2022, 23, 5634. https://doi.org/10.3390/ijms23105634
Feijóo-Bandín S, Aragón-Herrera A, Otero-Santiago M, Anido-Varela L, Moraña-Fernández S, Tarazón E, Roselló-Lletí E, Portolés M, Gualillo O, González-Juanatey JR, et al. Role of Sodium-Glucose Co-Transporter 2 Inhibitors in the Regulation of Inflammatory Processes in Animal Models. International Journal of Molecular Sciences. 2022; 23(10):5634. https://doi.org/10.3390/ijms23105634
Chicago/Turabian StyleFeijóo-Bandín, Sandra, Alana Aragón-Herrera, Manuel Otero-Santiago, Laura Anido-Varela, Sandra Moraña-Fernández, Estefanía Tarazón, Esther Roselló-Lletí, Manuel Portolés, Oreste Gualillo, José Ramón González-Juanatey, and et al. 2022. "Role of Sodium-Glucose Co-Transporter 2 Inhibitors in the Regulation of Inflammatory Processes in Animal Models" International Journal of Molecular Sciences 23, no. 10: 5634. https://doi.org/10.3390/ijms23105634