
Microbial Consortia and Mixed Plastic Waste: Pangenomic Analysis Reveals Potential for Degradation of Multiple Plastic Types via Previously Identified PET Degrading Bacteria

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
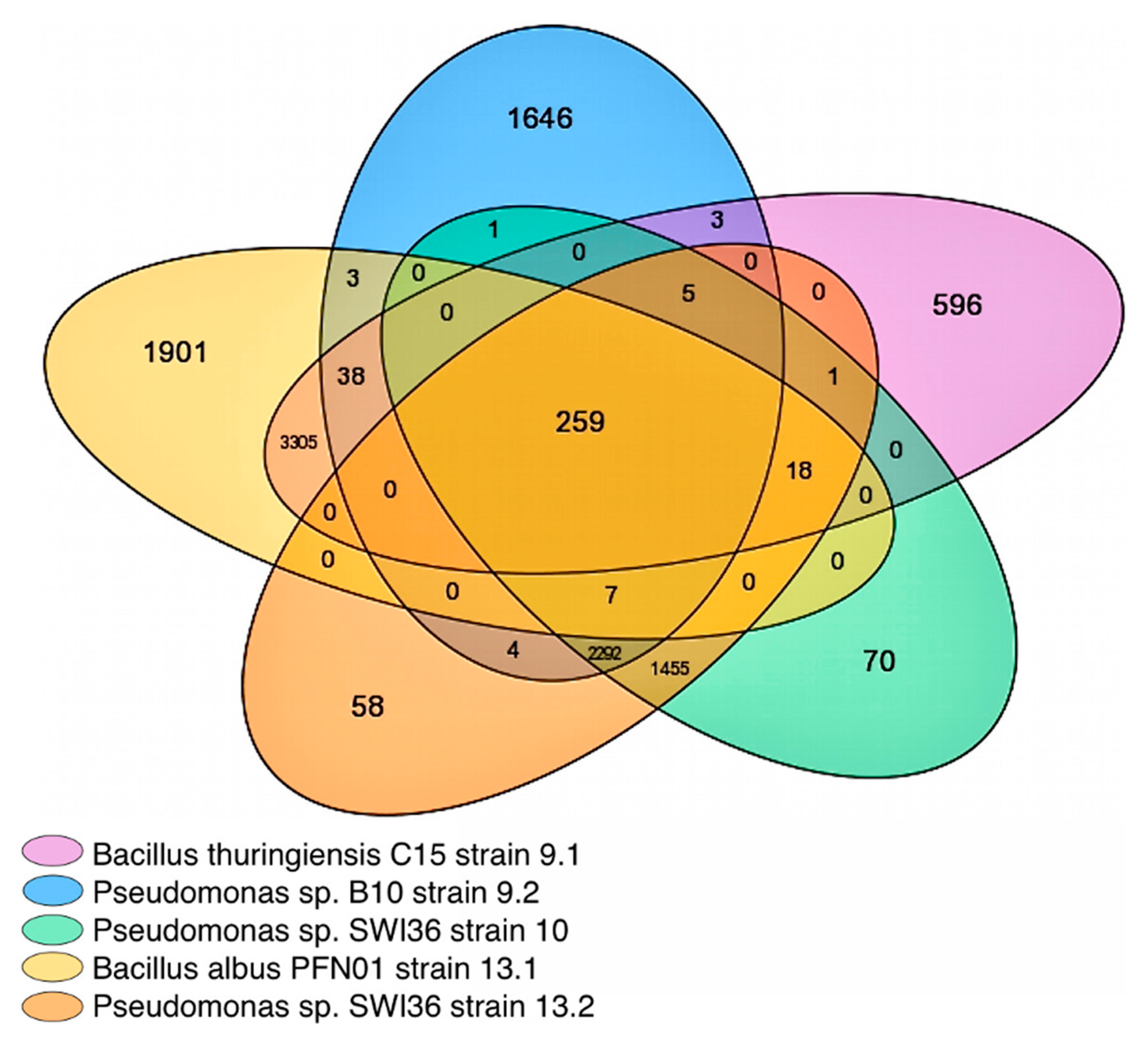
2.1. Employing Pangenomic Analysis toward Understanding Synergistic PET Degradation
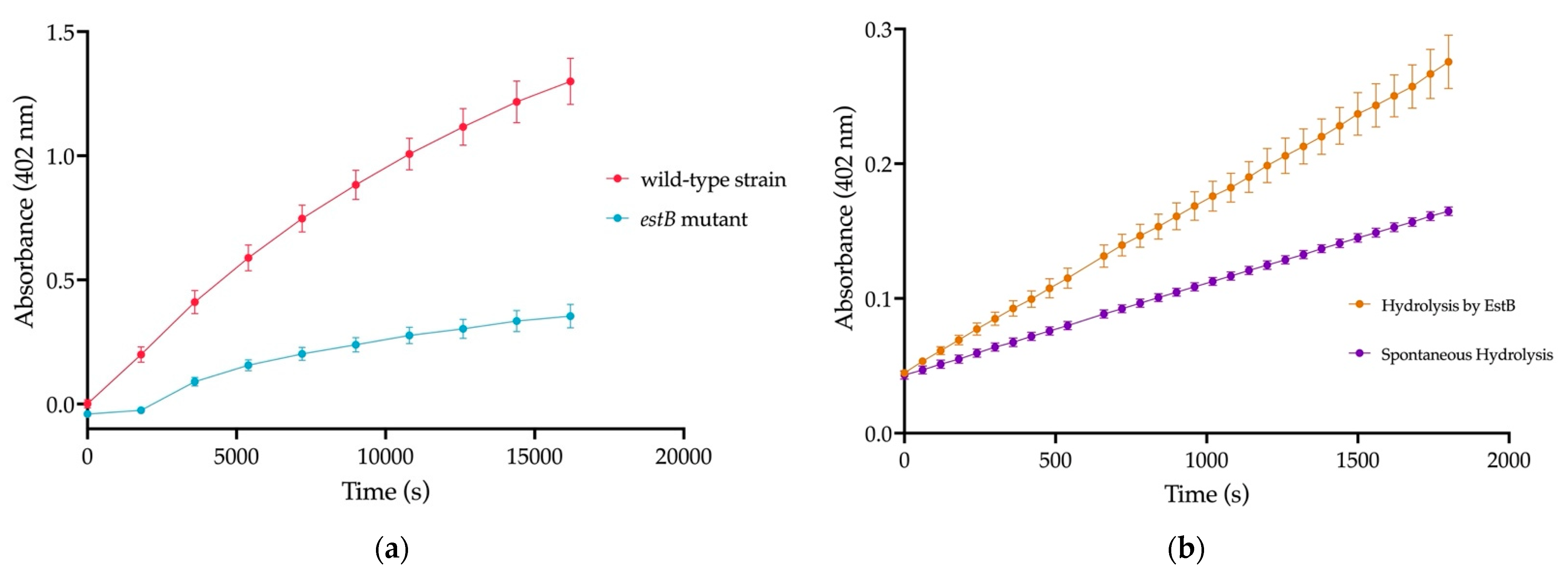
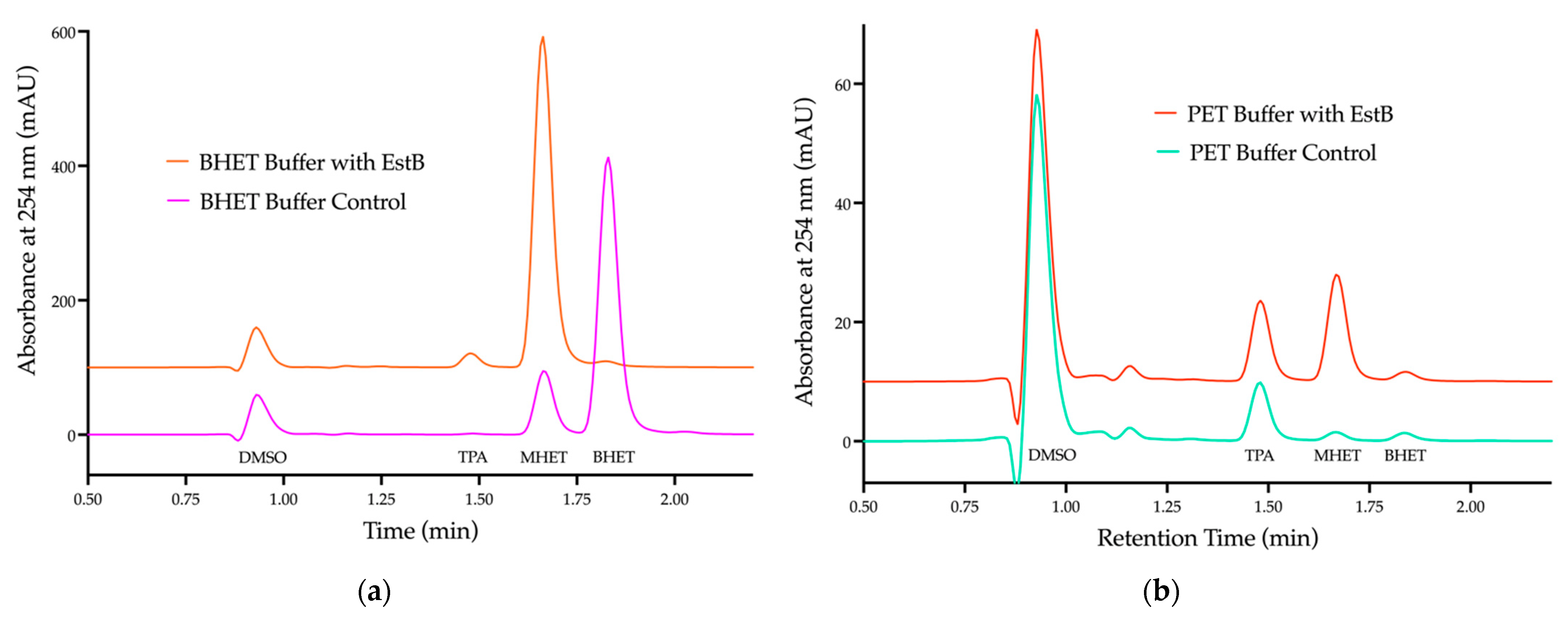
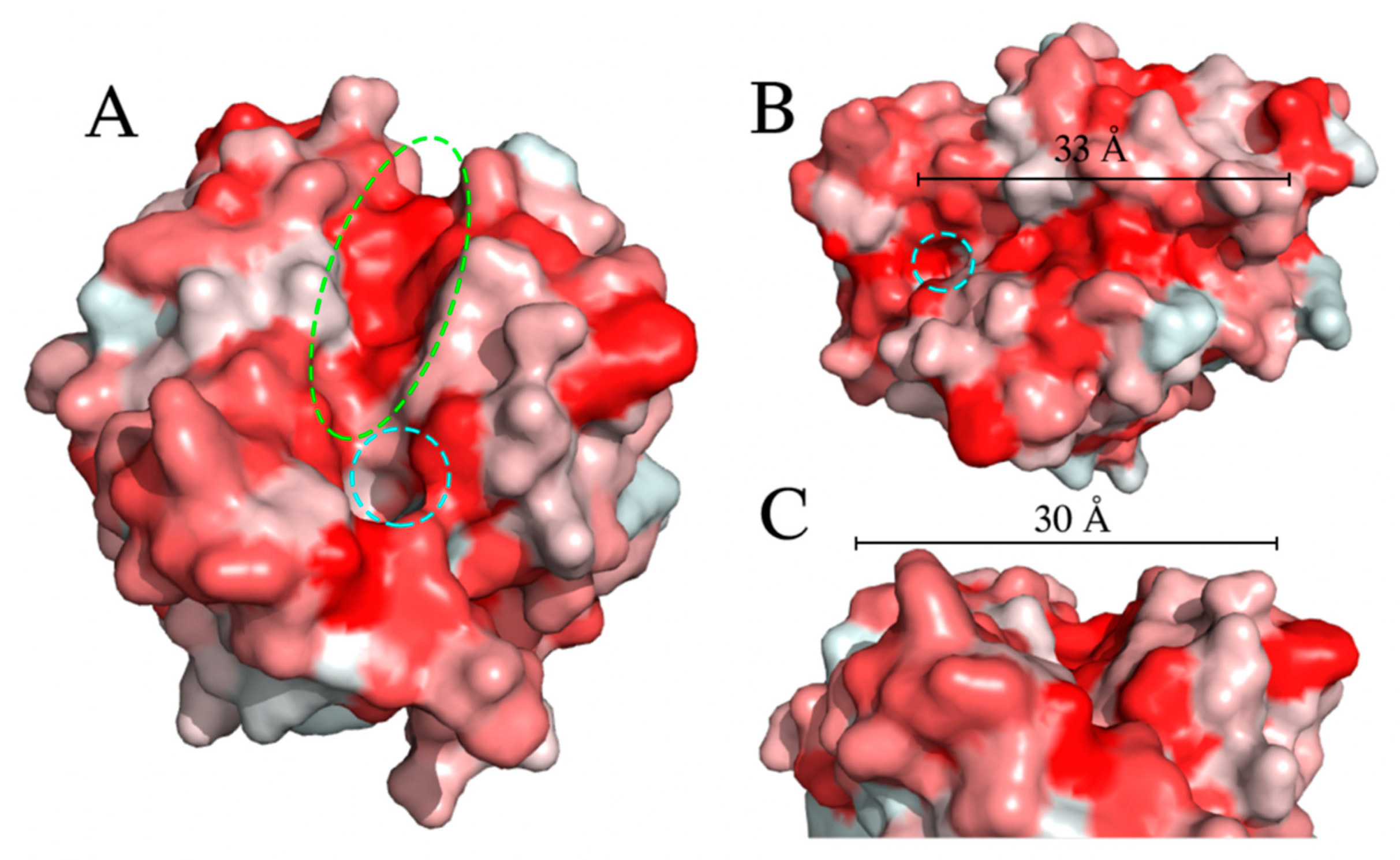
2.2. Esterase EstB in Strain 9.2 Is Active against PET
2.3. Identification of Predicted Plastic Degrading Genes Using PMBD
2.4. Genes Implicated in PET Degradation
2.5. Genes Implicated in the Biodegradation of Other Plastic Types
2.6. Genes Implicated in the Biodegradation of Plasticizer and Xenobiotics
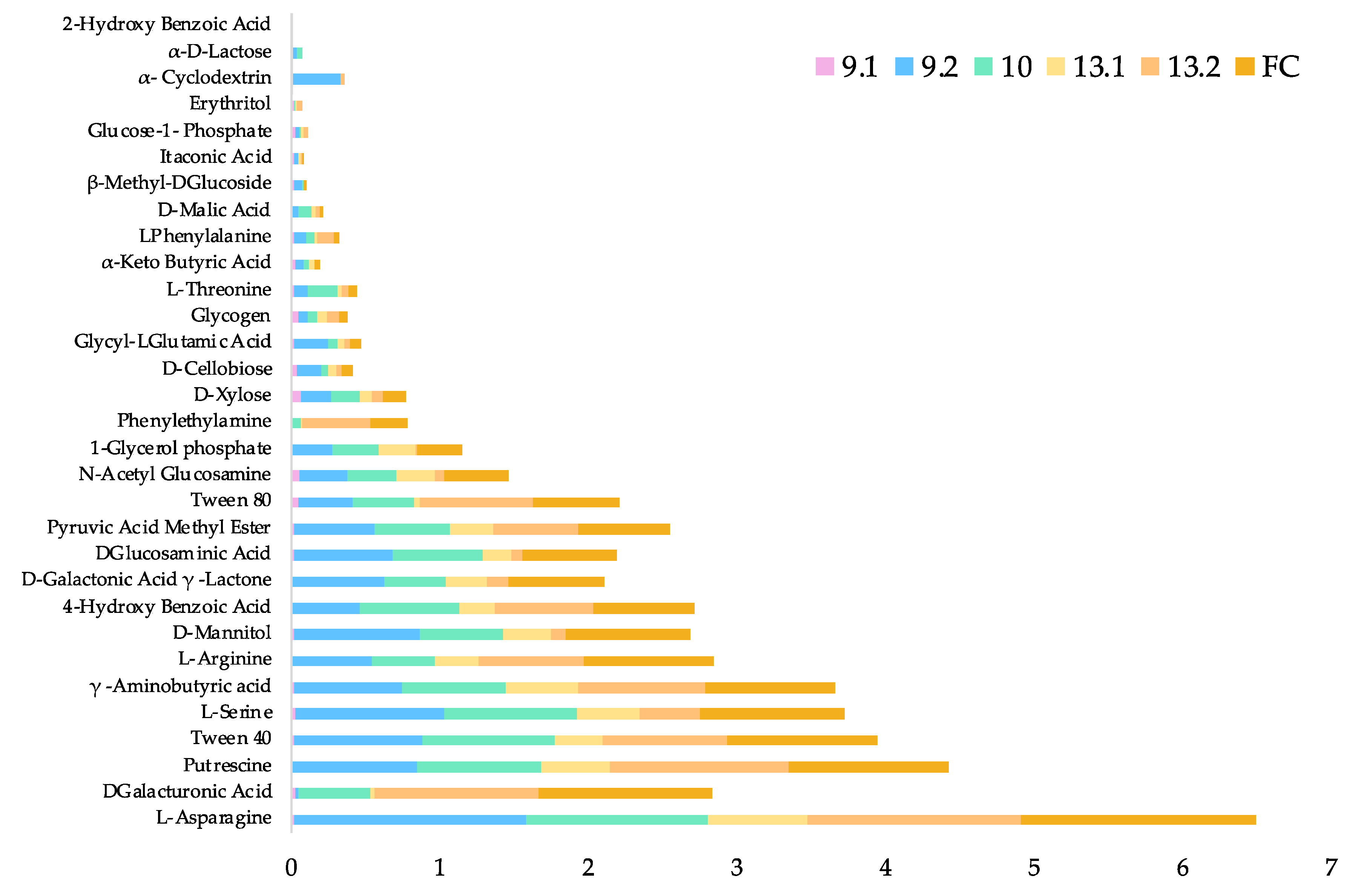
2.7. Strains within the Full Consortium Have Diverse Carbon Sources Utilization
2.8. Differential RNAseq Analysis Shows Diverse Transcriptional Activity Amongst the Individual Strains Related to PET Monomer Degradation and PHA Storage
3. Discussion
4. Materials and Methods
4.1. Omics Approaches
4.1.1. Pangenome Analysis
4.1.2. PMBD Analysis
4.2. RNA Sequencing to Determine Genes Implicated in PET Biodegradation
4.2.1. Total RNA Extraction and Sequencing
4.2.2. RNAseq Analysis
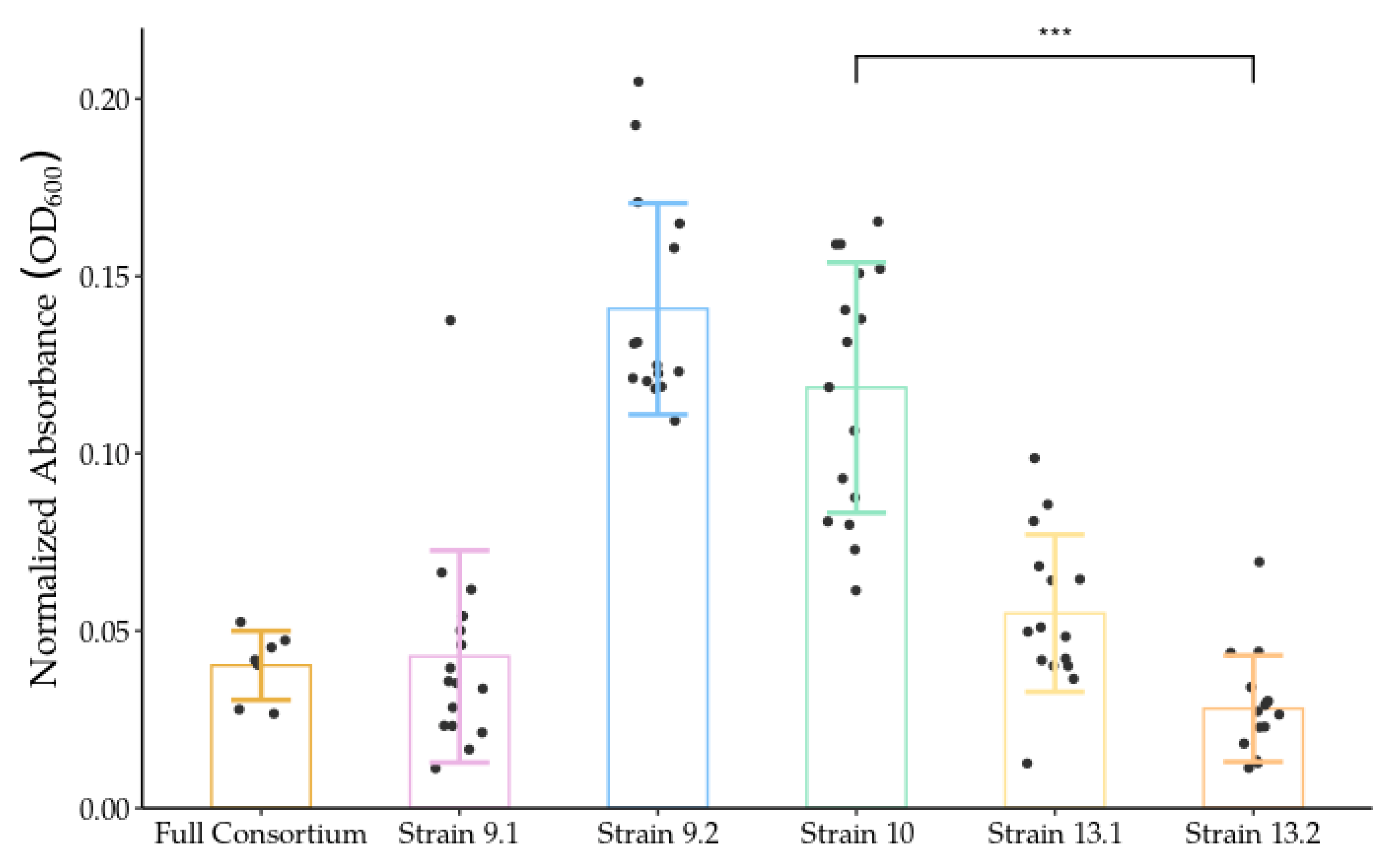
4.3. Evaluation of Carbon Source Utilization and Strain Biofilm Production
4.3.1. Biolog EcoPlates
4.3.2. Microtiter Plate Biofilm Quantification Assay
4.4. Mutant Preparation of ΔestB
4.5. EstB Purification
4.5.1. Amplification and Purification of estB Vector Insert
4.5.2. Assembly, Isolation, and Purification of RDpET24estB Expression Vector
4.5.3. Electroporation of BL21 E. Coli
4.5.4. Expression and Purification of EstB
4.6. Screening Purified EstB and Mutant ΔestB for Esterase Activity
4.7. Screening for PET and BHET Degradation
4.7.1. Synthesis of microPET and nanoPET
4.7.2. BHET and PET Hydrolysis Assay
4.7.3. PET Monomer Analysis Using HPLC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [Green Version]
- Plastic Oceans International. Available online: https://plasticoceans.org/the-facts/ (accessed on 27 April 2022).
- Peng, Y.; Wu, P.; Schartup, A.T.; Zhang, Y. Plastic Waste Release Caused by COVID-19 and Its Fate in the Global Ocean. Proc. Natl. Acad. Sci. USA 2021, 118, e2111530118. [Google Scholar] [CrossRef]
- Shams, M.; Alam, I.; Mahbub, M.S. Plastic Pollution during COVID-19: Plastic Waste Directives and Its Long-Term Impact on the Environment. Environ. Adv. 2021, 5, 100119. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic Waste Inputs from Land into the Ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef]
- Galloway, T.S. Micro- and Nano-Plastics and Human Health. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 343–366. [Google Scholar] [CrossRef] [Green Version]
- Jâms, I.B.; Windsor, F.M.; Poudevigne-Durance, T.; Ormerod, S.J.; Durance, I. Estimating the Size Distribution of Plastics Ingested by Animals. Nat. Commun. 2020, 11, 1594. [Google Scholar] [CrossRef] [Green Version]
- Chae, Y.; An, Y.-J. Current Research Trends on Plastic Pollution and Ecological Impacts on the Soil Ecosystem: A Review. Environ. Pollut. 2018, 240, 387–395. [Google Scholar] [CrossRef]
- Gross, L.; Enck, J. Confronting Plastic Pollution to Protect Environmental and Public Health. PLoS Biol. 2021, 19, e3001131. [Google Scholar] [CrossRef]
- Christman, G.D.; León-Zayas, R.I.; Zhao, R.; Summers, Z.M.; Biddle, J.F. Novel Clostridial Lineages Recovered from Metagenomes of a Hot Oil Reservoir. Sci. Rep. 2020, 10, 8048. [Google Scholar] [CrossRef]
- Brahney, J.; Hallerud, M.; Heim, E.; Hahnenberger, M.; Sukumaran, S. Plastic Rain in Protected Areas of the United States. Science 2020, 368, 1257–1260. [Google Scholar] [CrossRef]
- Danso, D.; Chow, J.; Streit, W.R. Plastics: Environmental and Biotechnological Perspectives on Microbial Degradation. Appl. Environ. Microbiol. 2019, 19, e01095-19. [Google Scholar] [CrossRef] [Green Version]
- Kale, S.; Deshmukh, A.G.; Dudhare, M.; Patil, V. Microbial Degradation Of Plastic—A Review. J. Biochem. Technol. 2015, 6, 952–961. [Google Scholar] [CrossRef]
- Mohanan, N.; Montazer, Z.; Sharma, P.K.; Levin, D.B. Microbial and Enzymatic Degradation of Synthetic Plastics. Front. Microbiol. 2020, 11, 580709. [Google Scholar] [CrossRef]
- Zrimec, J.; Kokina, M.; Jonasson, S.; Zorrilla, F.; Zelezniak, A. Plastic-Degrading Potential across the Global Microbiome Correlates with Recent Pollution Trends. MBio 2021, 12, e02155-21. [Google Scholar] [CrossRef]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A Bacterium That Degrades and Assimilates Poly(Ethylene Terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef]
- Tanasupawat, S.; Takehana, T.; Yoshida, S.; Hiraga, K.; Oda, K. Ideonella Sakaiensis sp. Nov., Isolated from a Microbial Consortium That Degrades Poly(Ethylene Terephthalate). Int. J. Syst. Evol. Microbiol. 2016, 66, 2813–2818. [Google Scholar] [CrossRef]
- Sulaiman, S.; Yamato, S.; Kanaya, E.; Kim, J.J.; Koga, Y.; Takano, K.; Kanaya, S. Isolation of a Novel Cutinase Homolog with Polyethylene Terephthalate-Degrading Activity from Leaf-Branch Compost by Using a Metagenomic Approach. Appl. Environ. Microbiol. 2012, 78, 1556–1562. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, H. Microbial Consortia Are Needed to Degrade Soil Pollutants. Microorganisms 2022, 10, 261. [Google Scholar] [CrossRef]
- Bharti, V.; Gupta, B.; Kaur, J. Novel Bacterial Strains Pseudomonas sp. and Bacillus sp. Isolated from Petroleum Oil Contaminated Soils for Degradation of Flourene and Phenanthrene. Pollution 2019, 5, 657–669. [Google Scholar] [CrossRef]
- Carniel, A.; Valoni, É.; Nicomedes, J.; da Conceição Gomes, A.; de Castro, A.M. Lipase from Candida Antarctica (CALB) and Cutinase from Humicola Insolens Act Synergistically for PET Hydrolysis to Terephthalic Acid. Process Biochem. 2017, 59, 84–90. [Google Scholar] [CrossRef]
- Meyer-Cifuentes, I.E.; Werner, J.; Jehmlich, N.; Will, S.E.; Neumann-Schaal, M.; Öztürk, B. Synergistic Biodegradation of Aromatic-Aliphatic Copolyester Plastic by a Marine Microbial Consortium. Nat. Commun. 2020, 11, 5790. [Google Scholar] [CrossRef]
- Huerta Lwanga, E.; Thapa, B.; Yang, X.; Gertsen, H.; Salánki, T.; Geissen, V.; Garbeva, P. Decay of Low-Density Polyethylene by Bacteria Extracted from Earthworm’s Guts: A Potential for Soil Restoration. Sci. Total Environ. 2018, 624, 753–757. [Google Scholar] [CrossRef]
- Shah, Z.; Gulzar, M.; Hasan, F.; Shah, A.A. Degradation of Polyester Polyurethane by an Indigenously Developed Consortium of Pseudomonas and Bacillus Species Isolated from Soil. Polym. Degrad. Stab. 2016, 134, 349–356. [Google Scholar] [CrossRef]
- Roberts, C.; Edwards, S.; Vague, M.; León-Zayas, R.; Scheffer, H.; Chan, G.; Swartz, N.A.; Mellies, J.L. Environmental Consortium Containing Pseudomonas and Bacillus Species Synergistically Degrades Polyethylene Terephthalate Plastic. mSphere 2020, 6, e01151-20. [Google Scholar] [CrossRef]
- Das, N.; Chandran, P. Microbial Degradation of Petroleum Hydrocarbon Contaminants: An Overview. Biotechnol. Res. Int. 2010, 2011, e941810. [Google Scholar] [CrossRef] [Green Version]
- Leon-Zayas, R.; Roberts, C.; Vague, M.; Mellies, J.L. Draft Genome Sequences of Five Environmental Bacterial Isolates That Degrade Polyethylene Terephthalate Plastic. Microbiol. Resour. Announc. 2019, 8, e00237-19. [Google Scholar] [CrossRef] [Green Version]
- Medini, D.; Donati, C.; Tettelin, H.; Masignani, V.; Rappuoli, R. The Microbial Pan-Genome. Curr. Opin. Genet. Dev. 2005, 15, 589–594. [Google Scholar] [CrossRef]
- Pacheco-Moreno, A.; Stefanato, F.L.; Ford, J.J.; Trippel, C.; Uszkoreit, S.; Ferrafiat, L.; Grenga, L.; Dickens, R.; Kelly, N.; Kingdon, A.D.; et al. Pan-Genome Analysis Identifies Intersecting Roles for Pseudomonas Specialized Metabolites in Potato Pathogen Inhibition. eLife 2021, 10, e71900. [Google Scholar] [CrossRef]
- Blaustein, R.A.; McFarland, A.G.; Ben Maamar, S.; Lopez, A.; Castro-Wallace, S.; Hartmann, E.M. Pangenomic Approach To Understanding Microbial Adaptations within a Model Built Environment, the International Space Station, Relative to Human Hosts and Soil. mSystems 2019, 4, e00281-18. [Google Scholar] [CrossRef] [Green Version]
- Polz, M.F.; Alm, E.J.; Hanage, W.P. Horizontal Gene Transfer and the Evolution of Bacterial and Archaeal Population Structure. Trends Genet. 2013, 29, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Gonzales-Siles, L.; Karlsson, R.; Schmidt, P.; Salvà-Serra, F.; Jaén-Luchoro, D.; Skovbjerg, S.; Moore, E.R.B.; Gomila, M. A Pangenome Approach for Discerning Species-Unique Gene Markers for Identifications of Streptococcus Pneumoniae and Streptococcus Pseudopneumoniae. Front. Cell. Infect. Microbiol. 2020, 10, 222. [Google Scholar] [CrossRef]
- Hara, H.; Eltis, L.D.; Davies, J.E.; Mohn, W.W. Transcriptomic Analysis Reveals a Bifurcated Terephthalate Degradation Pathway in Rhodococcus sp. Strain RHA1. J. Bacteriol. 2007, 189, 1641–1647. [Google Scholar] [CrossRef] [Green Version]
- Croucher, N.J.; Thomson, N.R. Studying Bacterial Transcriptomes Using RNA-Seq. Curr. Opin. Microbiol. 2010, 13, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Bano, N.; Bag, S.K.; Chaudhary, D.R.; Jha, B. Transcriptome-Guided Insights Into Plastic Degradation by the Marine Bacterium. Front. Microbiol. 2021, 12, 751571. [Google Scholar] [CrossRef]
- Gan, Z.; Zhang, H. PMBD: A Comprehensive Plastics Microbial Biodegradation Database. Database 2019, 2019, baz119. [Google Scholar] [CrossRef]
- Vallenet, D.; Calteau, A.; Dubois, M.; Amours, P.; Bazin, A.; Beuvin, M.; Burlot, L.; Bussell, X.; Fouteau, S.; Gautreau, G.; et al. MicroScope: An Integrated Platform for the Annotation and Exploration of Microbial Gene Functions through Genomic, Pangenomic and Metabolic Comparative Analysis. Nucleic Acids Res. 2020, 48, D579–D589. [Google Scholar] [CrossRef] [Green Version]
- Miele, V.; Penel, S.; Duret, L. Ultra-Fast Sequence Clustering from Similarity Networks with SiLiX. BMC Bioinform. 2011, 12, 116. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Yin, X.; Liu, T.; Zhang, H.; Chen, G.; Wu, S. Biodegradation of Bis(2-Hydroxyethyl) Terephthalate by a Newly Isolated Enterobacter sp. HY1 and Characterization of Its Esterase Properties. J. Basic Microbiol. 2020, 60, 699–711. [Google Scholar] [CrossRef]
- Huang, W.; Wilks, A. A Rapid Seamless Method for Gene Knockout in Pseudomonas Aeruginosa. BMC Microbiol. 2017, 17, 199. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Joo, S.; Cho, I.J.; Seo, H.; Son, H.F.; Sagong, H.-Y.; Shin, T.J.; Choi, S.Y.; Lee, S.Y.; Kim, K.-J. Structural Insight into Molecular Mechanism of Poly(Ethylene Terephthalate) Degradation. Nat. Commun. 2018, 9, 382. [Google Scholar] [CrossRef] [Green Version]
- Austin, H.P.; Allen, M.D.; Donohoe, B.S.; Rorrer, N.A.; Kearns, F.L.; Silveira, R.L.; Pollard, B.C.; Dominick, G.; Duman, R.; El Omari, K.; et al. Characterization and Engineering of a Plastic-Degrading Aromatic Polyesterase. Proc. Natl. Acad. Sci. USA 2018, 115, E4350–E4357. [Google Scholar] [CrossRef] [Green Version]
- Herold, D.A.; Keil, K.; Bruns, D.E. Oxidation of Polyethylene Glycols by Alcohol Dehydrogenase. Biochem. Pharmacol. 1989, 38, 73–76. [Google Scholar] [CrossRef]
- Rogers, J.D.; Thurman, E.M.; Ferrer, I.; Rosenblum, J.S.; Evans, M.V.; Mouser, P.J.; Ryan, J.N. Degradation of Polyethylene Glycols and Polypropylene Glycols in Microcosms Simulating a Spill of Produced Water in Shallow Groundwater. Environ. Sci. Process. Impacts 2019, 21, 256–268. [Google Scholar] [CrossRef]
- Sunny; Maurya, A.; Vats, M.K.; Khare, S.K.; Srivastava, K.R. Computational Exploration of Bio-Remediation Solution for Mixed Plastic Waste. BioRxiv 2022, 485065. [Google Scholar] [CrossRef]
- Hou, L.; Majumder, E.L.-W. Potential for and Distribution of Enzymatic Biodegradation of Polystyrene by Environmental Microorganisms. Materials 2021, 14, 503. [Google Scholar] [CrossRef]
- Gambarini, V.; Pantos, O.; Kingsbury, J.M.; Weaver, L.; Handley, K.M.; Lear, G. Phylogenetic Distribution of Plastic-Degrading Microorganisms. MSystems 2021, 6, e01112-20. [Google Scholar] [CrossRef]
- Rabodonirina, S.; Rasolomampianina, R.; Krier, F.; Drider, D.; Merhaby, D.; Net, S.; Ouddane, B. Degradation of Fluorene and Phenanthrene in PAHs-Contaminated Soil Using Pseudomonas and Bacillus Strains Isolated from Oil Spill Sites. J. Environ. Manage. 2019, 232, 1–7. [Google Scholar] [CrossRef]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Wu, G. Reorganization of Gene Network for Degradation of Polycyclic Aromatic Hydrocarbons (PAHs) in Pseudomonas Aeruginosa PAO1 under Several Conditions. J. Appl. Genet. 2017, 58, 545–563. [Google Scholar] [CrossRef] [Green Version]
- Boll, M.; Geiger, R.; Junghare, M.; Schink, B. Microbial Degradation of Phthalates: Biochemistry and Environmental Implications. Environ. Microbiol. Rep. 2020, 12, 3–15. [Google Scholar] [CrossRef]
- Valkova, N.; Lépine, F.; Valeanu, L.; Dupont, M.; Labrie, L.; Bisaillon, J.-G.; Beaudet, R.; Shareck, F.; Villemur, R. Hydrolysis of 4-Hydroxybenzoic Acid Esters (Parabens) and Their Aerobic Transformation into Phenol by the Resistant Enterobacter Cloacae Strain EM. Appl. Environ. Microbiol. 2001, 67, 2404–2409. [Google Scholar] [CrossRef] [Green Version]
- Nichols, N.N.; Harwood, C.S. PcaK, a High-Affinity Permease for the Aromatic Compounds 4-Hydroxybenzoate and Protocatechuate from Pseudomonas Putida. J. Bacteriol. 1997, 179, 5056–5061. [Google Scholar] [CrossRef] [Green Version]
- Harwood, C.S.; Nichols, N.N.; Kim, M.K.; Ditty, J.L.; Parales, R.E. Identification of the PcaRKF Gene Cluster from Pseudomonas Putida: Involvement in Chemotaxis, Biodegradation, and Transport of 4-Hydroxybenzoate. J. Bacteriol. 1994, 176, 6479–6488. [Google Scholar] [CrossRef] [Green Version]
- Ndahebwa Muhonja, C.; Magoma, G.; Imbuga, M.; Makonde, H.M. Molecular Characterization of Low-Density Polyethene (LDPE) Degrading Bacteria and Fungi from Dandora Dumpsite, Nairobi, Kenya. Int. J. Microbiol. 2018, 2018, 4167845. [Google Scholar] [CrossRef]
- Song, C.; Mazzola, M.; Cheng, X.; Oetjen, J.; Alexandrov, T.; Dorrestein, P.; Watrous, J.; van der Voort, M.; Raaijmakers, J.M. Molecular and Chemical Dialogues in Bacteria-Protozoa Interactions. Sci. Rep. 2015, 5, 12837. [Google Scholar] [CrossRef] [Green Version]
- Jendrossek, D.; Handrick, R. Microbial Degradation of Polyhydroxyalkanoates. Annu. Rev. Microbiol. 2002, 56, 403–432. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Chen, C.-C.; Han, X.; Ko, T.-P.; Liu, W.; Guo, R.-T. Structural Studies Reveal the Molecular Mechanism of PETase. FEBS J. 2018, 285, 3717–3723. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.-D. Microbial Colonization of Polymeric Materials for Space Applications and Mechanisms of Biodeterioration: A Review. Int. Biodeterior. Biodegrad. 2007, 59, 170–179. [Google Scholar] [CrossRef]
- Kyaw, B.M.; Champakalakshmi, R.; Sakharkar, M.K.; Lim, C.S.; Sakharkar, K.R. Biodegradation of Low Density Polythene (LDPE) by Pseudomonas Species. Indian J. Microbiol. 2012, 52, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Das, M.P.; Kumar, S. An Approach to Low-Density Polyethylene Biodegradation by Bacillus Amyloliquefaciens. 3 Biotech. 2015, 5, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubpachat, T.; Sombatsompop, N.; Prapagdee, B. Isolation and Role of Polylactic Acid-Degrading Bacteria on Degrading Enzymes Productions and PLA Biodegradability at Mesophilic Conditions. Polym. Degrad. Stab. 2018, 152, 75–85. [Google Scholar] [CrossRef]
- Bonifer, K.S.; Wen, X.; Hasim, S.; Phillips, E.K.; Dunlap, R.N.; Gann, E.R.; DeBruyn, J.M.; Reynolds, T.B. Bacillus Pumilus B12 Degrades Polylactic Acid and Degradation Is Affected by Changing Nutrient Conditions. Front. Microbiol. 2019, 10, 2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Ichihara, Y.; Yamada, M.; Tonomura, K. Some Characteristics of Pseudomonas 0–3 Which Utilizes Polyvinyl Alcohol. Agric. Biol. Chem. 1973, 37, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Rolsky, C.; Kelkar, V. Degradation of Polyvinyl Alcohol in US Wastewater Treatment Plants and Subsequent Nationwide Emission Estimate. Int. J. Environ. Res. Public. Health 2021, 18, 6027. [Google Scholar] [CrossRef] [PubMed]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An Overview of Chemical Additives Present in Plastics: Migration, Release, Fate and Environmental Impact during Their Use, Disposal and Recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef]
- Keresztes, S.; Tatár, E.; Czégény, Z.; Záray, G.; Mihucz, V.G. Study on the Leaching of Phthalates from Polyethylene Terephthalate Bottles into Mineral Water. Sci. Total Environ. 2013, 458–460, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Abboudi, M.; Odeh, A.; Aljoumaa, K. Carbonyl Compound Leaching from Polyethylene Terephthalate into Bottled Water under Sunlight Exposure. Toxicol. Environ. Chem. 2016, 98, 167–178. [Google Scholar] [CrossRef]
- Darbre, P.D.; Harvey, P.W. Paraben Esters: Review of Recent Studies of Endocrine Toxicity, Absorption, Esterase and Human Exposure, and Discussion of Potential Human Health Risks. J. Appl. Toxicol. 2008, 28, 561–578. [Google Scholar] [CrossRef]
- Dutta, S.; Haggerty, D.K.; Rappolee, D.A.; Ruden, D.M. Phthalate Exposure and Long-Term Epigenomic Consequences: A Review. Front. Genet. 2020, 11, 405. [Google Scholar] [CrossRef]
- Baloyi, N.D.; Tekere, M.; Maphangwa, K.W.; Masindi, V. Insights Into the Prevalence and Impacts of Phthalate Esters in Aquatic Ecosystems. Front. Environ. Sci. 2021, 9, 684190. [Google Scholar] [CrossRef]
- Wagner, S.; Schlummer, M. Legacy Additives in a Circular Economy of Plastics: Current Dilemma, Policy Analysis, and Emerging Countermeasures. Resour. Conserv. Recycl. 2020, 158, 104800. [Google Scholar] [CrossRef]
- Plass, C.; Hiessl, R.; Kleber, J.; Grimm, A.; Langsch, A.; Otter, R.; Liese, A.; Gröger, H. Towards Bio-Based Plasticizers with Reduced Toxicity: Synthesis and Performance Testing of a 3-Methylphthalate. Sustain. Chem. Pharm. 2020, 18, 100319. [Google Scholar] [CrossRef]
- Jamarani, R.; Erythropel, H.C.; Nicell, J.A.; Leask, R.L.; Marić, M. How Green Is Your Plasticizer? Polymers 2018, 10, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, L.; Giersch, T.; Wünschiers, R. Efficiency of RNA Extraction from Selected Bacteria in the Context of Biogas Production and Metatranscriptomics. Anaerobe 2014, 29, 85–90. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 11 February 2022).
- Chen, S.; Huang, T.; Zhou, Y.; Han, Y.; Xu, M.; Gu, J. AfterQC: Automatic Filtering, Trimming, Error Removing and Quality Control for Fastq Data. BMC Bioinform. 2017, 18, 80. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357-U54. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Hernández, A.G.; Muñoz-Tabares, J.A.; Aguilar-Guzmán, J.C.; Vazquez-Duhalt, R. A Novel and Simple Method for Polyethylene Terephthalate (PET) Nanoparticle Production. Environ. Sci. Nano 2019, 6, 2031–2036. [Google Scholar] [CrossRef]
- Palm, G.J.; Reisky, L.; Böttcher, D.; Müller, H.; Michels, E.A.P.; Walczak, M.C.; Berndt, L.; Weiss, M.S.; Bornscheuer, U.T.; Weber, G. Structure of the Plastic-Degrading Ideonella Sakaiensis MHETase Bound to a Substrate. Nat. Commun. 2019, 10, 1717. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, M.; Kawakami, N.; Tomizawa, A.; Miyamoto, K. Efficient Degradation of Poly(Ethylene Terephthalate) with Thermobifida Fusca Cutinase Exhibiting Improved Catalytic Activity Generated Using Mutagenesis and Additive-Based Approaches. Sci. Rep. 2019, 9, 16038. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, R.P.; Cunha, J.M.; Sousa, S.F. Perspectives on the Role of Enzymatic Biocatalysis for the Degradation of Plastic PET. Int. J. Mol. Sci. 2021, 22, 11257. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plastic | Enzyme | Species | Uniprot ID | % Identity | E-Value | Bit-Score | Strain |
|---|---|---|---|---|---|---|---|
| PET | Aldehyde dehydrogenase | Mycobacterium vanbaalenii | Q9KHU2 | 33.97 | 8.23 × 10−74 | 239 | all |
| Cellulose-binding protein | Micromonospora rifamycinica | A0A120F7D2 | 27.98 | 8.59 × 10−4 | 37.7 | 10/13.2 | |
| Feruloyl Esterase | Phialocephala subalpina | A0A1L7XXB0 | 32.00 | 1.43 × 10−4 | 39.7 | 9.2 | |
| Feruloyl Esterase | Rhynchosporium secalis | A0A1E1MSN7 | 27.89 | 5.65 × 10−5 | 42.7 | 9.1 | |
| Poly(ethylene terephthalate) hydrolase | Streptomyces sp. 111WW2 | A0A2P8AA05 | 23.77 | 2.93 × 10−5 | 43.1 | 10/13.2 | |
| Poly(ethylene terephthalate) hydrolase | Streptomyces sp. MH60 | A0A2S6X119 | 23.75 | 7.52 × 10−6 | 45.4 | 9.1 | |
| Putative terephthalate 1,2-dioxygenase | Rhodococcus sp. DK17 | Q6REK1 | 32.30 | 6.62 × 10−33 | 127 | 9.2/10/13.2 | |
| TPA 1,2-dioxygenase, reductase component 1 | Comamonas sp. | TPDR1 | 34.92 | 3.46 × 10−5 | 43.1 | 9.2/10/13.2 | |
| TPA 1,2-dioxygenase, reductase component 2 | Comamonas sp. | TPDR2 | 36.51 | 6.73 × 10−6 | 45.4 | 9.2/10/13.2 | |
| TPA 1,2-dioxygenase, oxygenase alpha 1 | Comamonas sp. | TPDA1 | 34.10 | 2.15 × 10−29 | 117 | 9.2/10/13.2 | |
| TPA 1,2-dioxygenase oxygenase alpha 2 | Comamonas sp. | TPDA2 | 34.10 | 2.21 × 10−29 | 117 | 9.2/10/13.2 | |
| Twin-arginine translocation pathway signal | Polaromonas sp. strain JS666 | Q12BN2 | 39.46 | 5.89 × 10−25 | 103 | 9.2/10/13.2 | |
| Glyoxalase | Azoarcus sp. PA01 | A0A0M0FWC0 | 26.471 | 1.19 × 10−4 | 37.7 | 9.1/13.1 | |
| PLA | PLA depolymerase | uncultured bacterium | A4UZ10 | 46.71 | 1.49 × 10−124 | 365 | 9.1/13.1 |
| PLA depolymerase | uncultured bacterium | A4UZ14 | 37.46 | 6.05 × 10−67 | 211 | 9.2/10/13.2 | |
| PLA depolymerase (Fragment) | uncultured bacterium | A4UZ11 | 48.5 | 1.80 × 10−123 | 362 | 9.1/9.2/13.1 | |
| PUR | Polyurethanase | Pseudomonas sp. FW305-BF6 | A0A2N8H9Y3 | 99.51 | 0.00 | 1225 | 9.2 |
| Polyurethanase | Pseudomonas fluorescens | A0A0C2A4M0 | 59.52 | 8.28 × 10−7 | 52 | 10/13.2 | |
| Polyurethanase (Fragment) | Pseudomonas sp. DrBHI1 | A0A246GXG4 | 66.61 | 0.00 | 733 | 9.2/10/13.2 | |
| Polyurethanase A | Pseudomonas sp. Os17 | A0A0D6BHI0 | 77.96 | 0.00 | 994 | 9.2 | |
| PVA | Polyvinyl alcohol dehydrogenase | Pseudomonas sp. FW305-BF6 | A0A2N8GY02 | 98.23 | 0.00 | 561 | 9.2 |
| Polyvinyl alcohol dehydrogenase | Xanthomonas arboricola | A0A2S6Y8G0 | 37.5 | 4.65 × 10−4 | 41.2 | 10/13.2 | |
| Polyvinyl alcohol dehydrogenase (Fragment) | Opitutae bacterium | A0A2D6VIL8 | 27.63 | 1.89 × 10−4 | 40.8 | 10/13.2 | |
| Oxidized polyvinyl alcohol hydrolase | Syntrophorhabdus sp. PtaB | A0A1V4WJI2 | 30.07 | 2.92 × 10−10 | 57 | 9.1/9.2/13.1 | |
| Probable polyvinyl alcohol dehydrogenase | Streptomyces rochei | Q83X81 | 36.15 | 1.60 × 10−6 | 49.3 | 9.2/10/13.2 | |
| PVA dehydrogenase PQQ dependent | Bradyrhizobium sp. | A0A160UKB5 | 28.36 | 4.92 × 10−4 | 41.6 | 9.2/10/13.2 | |
| PHA PHB | Poly(3-hydroxyalkanoate) depolymerase | Pseudomonas fluorescens | A0A0C1ZS59 | 100.00 | 0.00 | 577 | 9.2 |
| Poly(3-hydroxyalkanoate) depolymerase | Pseudomonas putida S12 | A0A0A7PVK5 | 99.65 | 0.00 | 574 | 10.13.2 | |
| Poly(3-hydroxybutyrate) depolymerase | Haladaptatus paucihalophilus | E7QQJ1 | 50.73 | 2.83 × 10−8 | 55.5 | 9.1 | |
| Poly(3-hydroxybutyrate) depolymerase | Marinobacter lutaoensis | A0A1V2DRR5 | 46.67 | 5.28 × 10−24 | 85.1 | 9.2 | |
| Poly(3-hydroxyalkanoate) depolymerase C | Paenibacillus polymyxa | A0A2X1WPU8 | 44.53 | 4.46 × 10−114 | 338 | 9.1 | |
| Poly(3-hydroxybutyrate) depolymerase | Marinobacter sp. AC-23 | A0A1S2CI13 | 44.44 | 6.19 × 10−17 | 67.4 | 10/13.2 | |
| Poly(3-hydroxyalkanoate) depolymerase | Pseudomonas fluorescens strain Pf0-1 | Q3KCH8 | 44.19 | 1.09 × 10−4 | 40.8 | 13.1 | |
| Poly(3-hydroxybutyrate) depolymerase | Bacillus megaterium ATCC12872 | D5DZL2 | 43.26 | 2.37 × 10−83 | 251 | 13.1 | |
| Poly(3-hydroxyalkanoate) synthase | Paracoccus denitrificans | Q9WX80 | 33.51 | 9.86 × 10−98 | 308 | all |
| Enzyme | Species | Uniprot ID | % Identity | E-Value | Bit-Score | Strain |
|---|---|---|---|---|---|---|
| Taurine dioxygenase | Gordonia phthalatica | A0A0N9N9P8 | 68.09 | 3.71 × 10−146 | 410 | 9.2/10/13.2 |
| 2-nitropropanedioxygenase | Gordonia phthalatica | A0A0N9N4Y4 | 51.10 | 1.02 × 10−83 | 254 | all |
| Tert-butyl alcohol monooxygenase reductase | Aquincola tertiaricarbonis | G8FRC6 | 44.59 | 2.50 × 10−84 | 255 | all |
| 4,4′-diaponeurosporenoateglycosyltransferase | Bacillus enclensis | A0A0V8HPX8 | 44.05 | 3.17 × 10−10 | 60.5 | all |
| Phenol hydrolase reductase | Methylibium petroleiphilum | A2SI47 | 41.38 | 4.97 × 10−11 | 61.2 | all |
| 2-hydroxy-6-oxo-6-(2′-carboxyphenyl)-hexa-2,4-dienoate hydrolase | Terrabacter sp. strain DBF63 | Q83ZF0 | 38.46 | 1.06 × 10−18 | 82.4 | all |
| Tert-butyl alcohol monooxygenase | Aquincola tertiaricarbonis | G8FRC5 | 38.18 | 1.27 × 10−4 | 37.7 | 9.2/10/13.1/13.2 |
| Quercetin 2,3-dioxygenase | Gordonia phthalatica | A0A0N9MT24 | 37.16 | 2.99 × 10−34 | 121 | all |
| NidB2 | Mycobacterium vanbaalenii | Q6H2J5 | 37.04 | 2.1 × 10−10 | 54.3 | 9.2/10/13.2 |
| Naphthalene inducible dioxygenase | Mycobacterium vanbaalenii | Q9KHU1 | 35.64 | 3.51 × 10−43 | 155 | 9.2/10/13.1/13.2 |
| 5,5′-dehydrodivanillateO-demethylase | Paraburkholderia tropica | A0A1A5XFM6 | 34.38 | 4.84 × 10−18 | 82 | all |
| Probable phenol hydrolase | Rhodococcus sp. EsD8 | N1M644 | 33.33 | 7.2 × 10−8 | 51.6 | all |
| Putative nitropropane dioxygenase | Rhodococcus sp. DK17 | Q6REN2 | 32.84 | 2.46 × 10−34 | 127 | all |
| 1-hydroxy-2-naphthoicaciddioxygenase | Mycobacterium sp. CH1 | C0KUL5 | 32.65 | 7.45 × 10−58 | 190 | 9.1 |
| Phenol hydrolase | Rhodococcus opacus M213 | K8XRS6 | 29.667 | 3.2 × 10−17 | 79 | all |
| 2-3DHBA3,4-dioxygenase | Pseudomonas stutzeri | A0A2Z5UC95 | 29.524 | 3.65 × 10−5 | 42.4 | 10/13.2 |
| Phenanthrene-4,5-dicarboxylate 5-decarboxylase | Pseudonocardia sp. Ae707 | A0A1Q8KNT8 | 27.727 | 2.02 × 10−9 | 54.3 | 9.1/9.2/10/13.2 |
| Enzyme | Species | Uniprot ID | % Identity | E-Value | Bit-Score | Strain |
|---|---|---|---|---|---|---|
| Phthalate 4,5-dioxygenase oxygenase reductase | Pseudomonas sp. 58R3 | A0A1B5EAD8 | 81.65 | 0.00 | 541 | 10/13.2 |
| Phthalate 4,5-dioxygenase | Pseudomonas fulva strain 12-X | F6AJ53 | 64.67 | 1.82 × 10−147 | 416 | 10/13.2 |
| Ferredoxin | Burkholderia cepacia | A0A1Z3YX76 | 57.05 | 2.73 × 10−128 | 367 | 9.2/10/13.2 |
| Reductase component of isophthalate dioxygenase | Comamonas sp. E6 | C4TNS5 | 54.05 | 2.47 × 10−5 | 43.1 | all |
| Putative phthalate dioxygenase reductase | Acinetobacter johnsonii SH046 | D0SH70 | 51.43 | 4.11 × 10−114 | 330 | 10/13.2 |
| Phthalate 4,5-dioxygenase | Hydrogenophaga sp. PBC | A0A1C9VA35 | 48.15 | 1.09 × 10−5 | 45.4 | 9.2 |
| Phthalate-dioxygenase | Hydrogenophaga intermedia | A0A1L1P942 | 48.15 | 1.09 × 10−5 | 45.4 | 9.2/10/13.2 |
| 4,5-dihydroxyphthalatedecarboxylase | Bacillus aquimaris | A0A1J6W284 | 46.43 | 1.42 × 10−11 | 64.7 | 9.1 |
| 4,5-dihydroxyphthalatedecarboxylase | Sporosarcina sp. P17b | A0A2G5XE92 | 44.90 | 2.82 × 10−5 | 43.1 | 13.1 |
| 4,5-dihydroxyphthalatedecarboxylase | Caballeronia megalochromosomata | A0A149R8D0 | 44.00 | 7.82 × 10−5 | 41.6 | 9.2 |
| Phthalate 4,5-dioxygenase oxygenase subunit | Novosphingobium sp. MBES04 | A0A0S6WTD6 | 43.93 | 2.44 × 10−82 | 250 | 10/13.2 |
| Phthalate 4,5-dioxygenase oxygenase reductase | Bordetella pertussis H921 | A0A0N2IN58 | 43.66 | 9.27 × 10−7 | 47.4 | 9.2 |
| Putative phthalate dioxygenase reductase | Bordetella pertussisH921 | Q2YM46 | 43.66 | 9.27 × 10−7 | 47.4 | 9.2 |
| Phthalate dioxygenase reductase | Pandoraea sputorum | A0A239SNB1 | 43.28 | 5.05 × 10−8 | 51.6 | 9.2 |
| Aromatic ring-opening dioxygenase LigA | Azoarcus sp. PA01 | A0A0M0FSY9 | 43.09 | 2.04 × 10−46 | 154 | all |
| Extradiol ring-cleavage dioxygenase | Gordonia phthalatica | A0A0N9NE56 | 43.03 | 4.80 × 10−51 | 166 | all |
| Putative phthalate dioxygenase reductase | Brucella abortus strain 2308 | Q2YM46 | 42.82 | 2.26 × 10−102 | 305 | 9.2 |
| Phthalate 4,5-dioxygenase (Phthalate dioxygenase) | Ramlibacter tataouinensis | F5XWD6 | 41.96 | 5.06 × 10−27 | 109 | all |
| Phthalate 4,5-dioxygenase oxygenase (OhpA2) | Paraburkholderia xenovorans | Q13QM0 | 41.82 | 8.89 × 10−32 | 122 | all |
| Phthalate 4,5-dioxygenase | Mycolicibacterium wolinskyi | A0A1X2FHI8 | 40.74 | 4.99 × 10−8 | 52 | 9.1/13.1 |
| Phthalate 4,5-dioxygenase oxygenase (OhpA2) | Variovorax sp. WDL1 | A0A109CIC4 | 39.54 | 1.06 × 10−4 | 37.7 | 9.1/10/13.1/13.2 |
| Phthalate 3,4-dioxygenase alpha subunit | Klenkia soli | A0A1H0Q6Z9 | 38.89 | 1.89 × 10−11 | 62.4 | 13.1 |
| Phthalate 4,5-dioxygenase oxygenase reductase | Gibberella fujikuroi | A0A0I9YA52 | 38.73 | 2.29 × 10−22 | 92.4 | 9.1/13.1 |
| Ferredoxin | Brevirhabdus pacifica | A0A1U7DHI8 | 38.21 | 1.30 × 10−9 | 57 | 9.1/13.1 |
| Phthalate 3,4-dioxygenase alpha subunit | Rhodococcus sp. OK302 | A0A235G3V7 | 38.18 | 2.65 × 10−4 | 38.5 | 9.1/13.1 |
| Oxygenase large subunit of phthalate dioxygenase | Terrabacter sp. strain DBF63 | Q8GI63 | 38.18 | 8.64 × 10−4 | 37 | all |
| Phthalate 4,5-dioxygenase oxygenase subunit | Thalassobius gelatinovorus | A0A0P1FRT5 | 37.84 | 5.95 × 10−20 | 82.8 | 13.1 |
| 4,5-dihydroxyphthalate decarboxylase | Pseudoruegeria lutimaris | A0A1G9AEN9 | 37.50 | 9.18 × 10−5 | 41.6 | 10/13.2 |
| Phthalate dioxygenase reductase | Gibberella subglutinans | A0A109QSZ1 | 37.27 | 1.64 × 10−26 | 104 | 9.1 |
| Phthalate 4,5-dioxygenase oxygenase subunit | Alphaproteobacteria bacterium | A0A2S6QA16 | 36.84 | 4.36 × 10−4 | 35.8 | 9.1 |
| Phthalate 3,4-dioxygenase alpha subunit | Mycolicibacterium rutilum | A0A1H6J828 | 35.48 | 7.71 × 10−6 | 45.4 | 9.2 |
| Oxygenase component of isophthalate dioxygenase | Comamonas sp. E6 | C4TNS2 | 34.43 | 1.37 × 10−28 | 113 | 9.2/10/13.1/13.2 |
| Putative phthalate dioxygenase reductase | Providenciaal califaciens PAL-3 | W3YHJ5 | 33.64 | 6.31 × 10−9 | 54.7 | 9.1/13.1 |
| 3,4-dihydroxyphthalatedecarboxylase | Arthrobacter sp. strain FB24 | A0AWN5 | 33.49 | 2.26 × 10−17 | 75.9 | 9.1/13.1 |
| 3,4-dihydroxy-3,4-dihydrophthalate dehydrogenase | Terrabacter sp. strain DBF63 | Q8GI60 | 33.15 | 2.80 × 10−8 | 51.6 | all |
| 3,4-dihydroxyphthalatedecarboxylase | Nocardioides terrae | A0A1I1EG61 | 33.15 | 3.34 × 10−17 | 76.6 | 10/13.2 |
| 3,4-dihydroxyphthalatedecarboxylase | Klenkia soli | A0A1H0Q8Z1 | 32.52 | 1.20 × 10−14 | 69.3 | 9.2 |
| Phthalate-dioxygenase | Bradyrhizobium sp. ORS3257 | A0A2U3Q6T0 | 32.22 | 1.48 × 10−23 | 97.8 | 13.1 |
| Cis-phthalate dihydrodiol dehydrogenase | Comamonas sp. E6 | A0A0M2DHI3 | 31.90 | 2.50 × 10−18 | 82.8 | 9.2 |
| Phthalate 3,4-dioxygenase alpha subunit | Rhodococcus rhodnii LMG5362 | R7WIP7 | 31.63 | 1.31 × 10−38 | 140 | 10/13.2 |
| Cis-phthalate dihydrodiol dehydrogenase | Burkholderia multivorans | A0A0H3KKN4 | 28.89 | 2.13 × 10−8 | 52.8 | 9.1/10/13.1/13.2 |
| Strain | Gene | logFC | logCPM | p-Value |
|---|---|---|---|---|
| 9.1 | Poly(3-hydroxyalkanoate) depolymerase | 3.42 | 2.09 | 3.64 × 10−2 |
| Phthalate 4,5-dioxygenase, reductase subunit | 2.60 | 8.24 | 2.20 × 10−9 | |
| Short-chain dehydrogenase/reductase | 1.90 | 3.59 | 6.32 × 10−3 | |
| Alcohol dehydrogenase 2 | 1.47 | 3.50 | 3.79 × 10−2 | |
| Poly(3-hydroxyalkanoate) depolymerase | 1.35 | 10.69 | 4.09 × 10−3 | |
| Phenol hydrolase | 1.31 | 10.94 | 1.68 × 10−4 | |
| Poly(3-hydroxybutyrate) depolymerase | 1.14 | 12.18 | 8.31 × 10−4 | |
| Glyoxal reductase | 0.89 | 7.14 | 1.58 × 10−2 | |
| Gentisate transporter | 0.70 | 9.60 | 4.30 × 10−2 | |
| 9.2 | Taurine dioxygenase | 3.67 | 2.06 | 4.91 × 10−3 |
| Putative regulatory protein | 2.90 | 2.32 | 1.75 × 10−2 | |
| Short-chain dehydrogenase/reductase | 1.21 | 5.19 | 2.51 × 10−2 | |
| Beta-carboxy-cis,cis-muconate cycloisomerase | 1.05 | 10.95 | 2.52 × 10−3 | |
| Beta-ketoadipyl CoA thiolase | 0.97 | 8.61 | 1.10 × 10−2 | |
| Aldehyde dehydrogenase | 0.89 | 8.35 | 3.30 × 10−2 | |
| Poly(3-hydroxyalkanoate) depolymerase | 0.78 | 9.45 | 3.32 × 10−2 | |
| 10 | Poly(3-hydroxybutyrate) depolymerase | 2.80 | −0.54 | 3.93 × 10−3 |
| 3,4-dihydroxyphthalate decarboxylase | 2.48 | 7.72 | 2.19 × 10−8 | |
| Terephthalate 1,2-dioxygenase, oxygenase | 1.32 | 1.20 | 3.09 × 10−2 | |
| Putative regulatory protein | 1.19 | 6.38 | 4.34 × 10−4 | |
| Putative regulatory protein | 1.09 | 5.72 | 9.15 × 10−4 | |
| 4,5-dihydroxyphthalate decarboxylase | 1.07 | 5.63 | 8.44 × 10−4 | |
| Poly(3-hydroxybutyrate) depolymerase (nlhH) | 1.04 | 5.15 | 2.35 × 10−3 | |
| Beta-carboxy-cis,cis-muconate cycloisomerase | 0.84 | 9.74 | 7.32 × 10−3 | |
| Putative regulatory protein | 0.78 | 9.89 | 3.17 × 10−2 | |
| Poly(3-hydroxybutyrate) depolymerase | 0.74 | 10.89 | 1.74 × 10−2 | |
| Quercetin 2,3-dioxygenase | 0.68 | 6.18 | 3.37 × 10−2 | |
| Surfactin synthase subunit 3 | 1.03 | 4.21 | 4.71 × 10−3 | |
| 13.2 | Poly(3-hydroxybutyrate) depolymerase | 2.80 | −0.54 | 3.89 × 10−3 |
| 3,4-dihydroxyphthalate decarboxylase | 2.48 | 7.72 | 2.11 × 10−8 | |
| Terephthalate 1,2-dioxygenase, terminal oxygenase | 1.32 | 1.20 | 3.09 × 10−2 | |
| Putative regulatory protein | 1.23 | 5.24 | 2.26 × 10−3 | |
| Putative regulatory protein | 1.19 | 6.38 | 4.31 × 10−4 | |
| Poly(3-hydroxybutyrate) depolymerase (nlhH) | 1.13 | 5.11 | 8.26 × 10−4 | |
| Beta-carboxy-cis,cis-muconate cycloisomerase | 0.84 | 9.74 | 7.31 × 10−3 | |
| Putative regulatory protein | 0.78 | 9.89 | 3.14 × 10−2 | |
| Poly(3-hydroxybutyrate) depolymerase | 0.74 | 10.90 | 1.73 × 10−2 | |
| Poly(3-hydroxyalkanoate) depolymerase | 0.73 | 8.09 | 2.62 × 10−2 | |
| Quercetin 2,3-dioxygenase | 0.68 | 6.18 | 3.32 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edwards, S.; León-Zayas, R.; Ditter, R.; Laster, H.; Sheehan, G.; Anderson, O.; Beattie, T.; Mellies, J.L. Microbial Consortia and Mixed Plastic Waste: Pangenomic Analysis Reveals Potential for Degradation of Multiple Plastic Types via Previously Identified PET Degrading Bacteria. Int. J. Mol. Sci. 2022, 23, 5612. https://doi.org/10.3390/ijms23105612
Edwards S, León-Zayas R, Ditter R, Laster H, Sheehan G, Anderson O, Beattie T, Mellies JL. Microbial Consortia and Mixed Plastic Waste: Pangenomic Analysis Reveals Potential for Degradation of Multiple Plastic Types via Previously Identified PET Degrading Bacteria. International Journal of Molecular Sciences. 2022; 23(10):5612. https://doi.org/10.3390/ijms23105612
Chicago/Turabian StyleEdwards, Sabrina, Rosa León-Zayas, Riyaz Ditter, Helen Laster, Grace Sheehan, Oliver Anderson, Toby Beattie, and Jay L. Mellies. 2022. "Microbial Consortia and Mixed Plastic Waste: Pangenomic Analysis Reveals Potential for Degradation of Multiple Plastic Types via Previously Identified PET Degrading Bacteria" International Journal of Molecular Sciences 23, no. 10: 5612. https://doi.org/10.3390/ijms23105612