
Identification of an Epigenetically Marked Locus within the Sex Determination Region of Channel Catfish

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
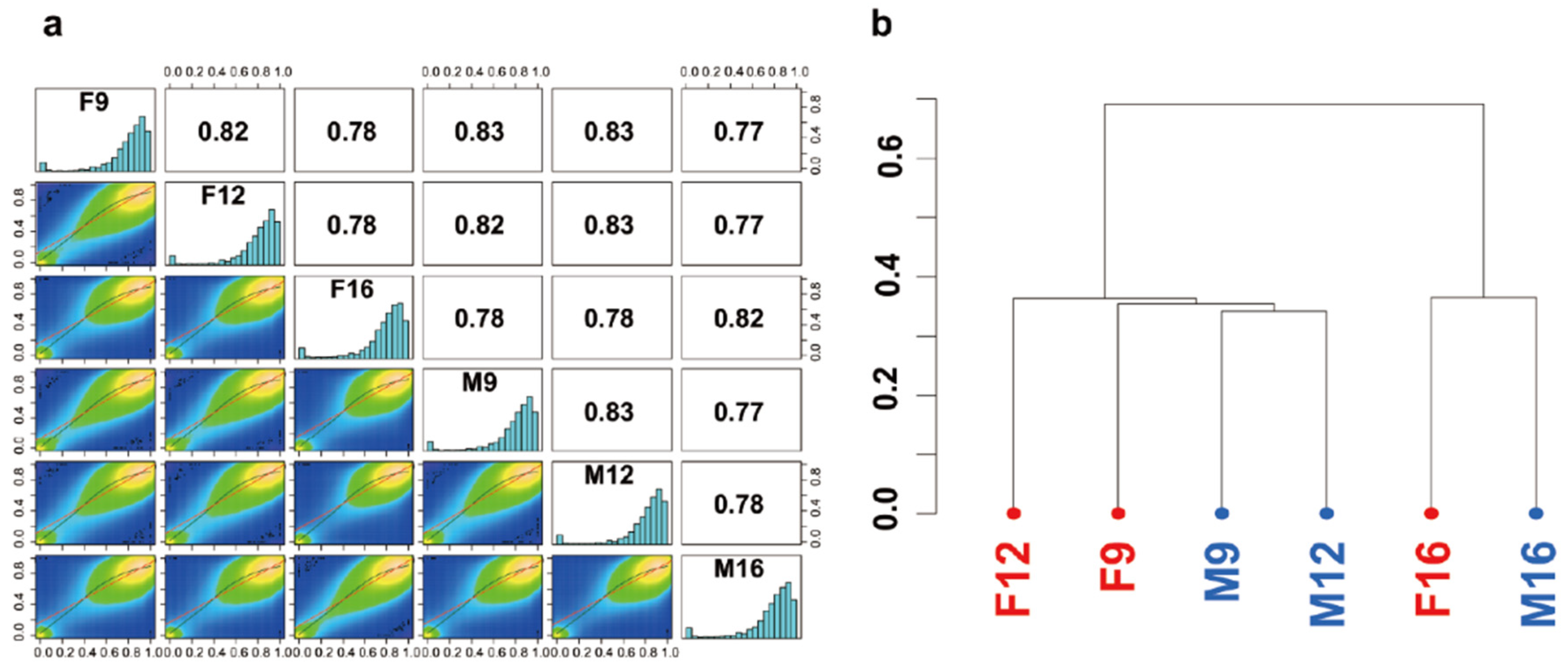
2.1. Genomic Landscape of Methylation
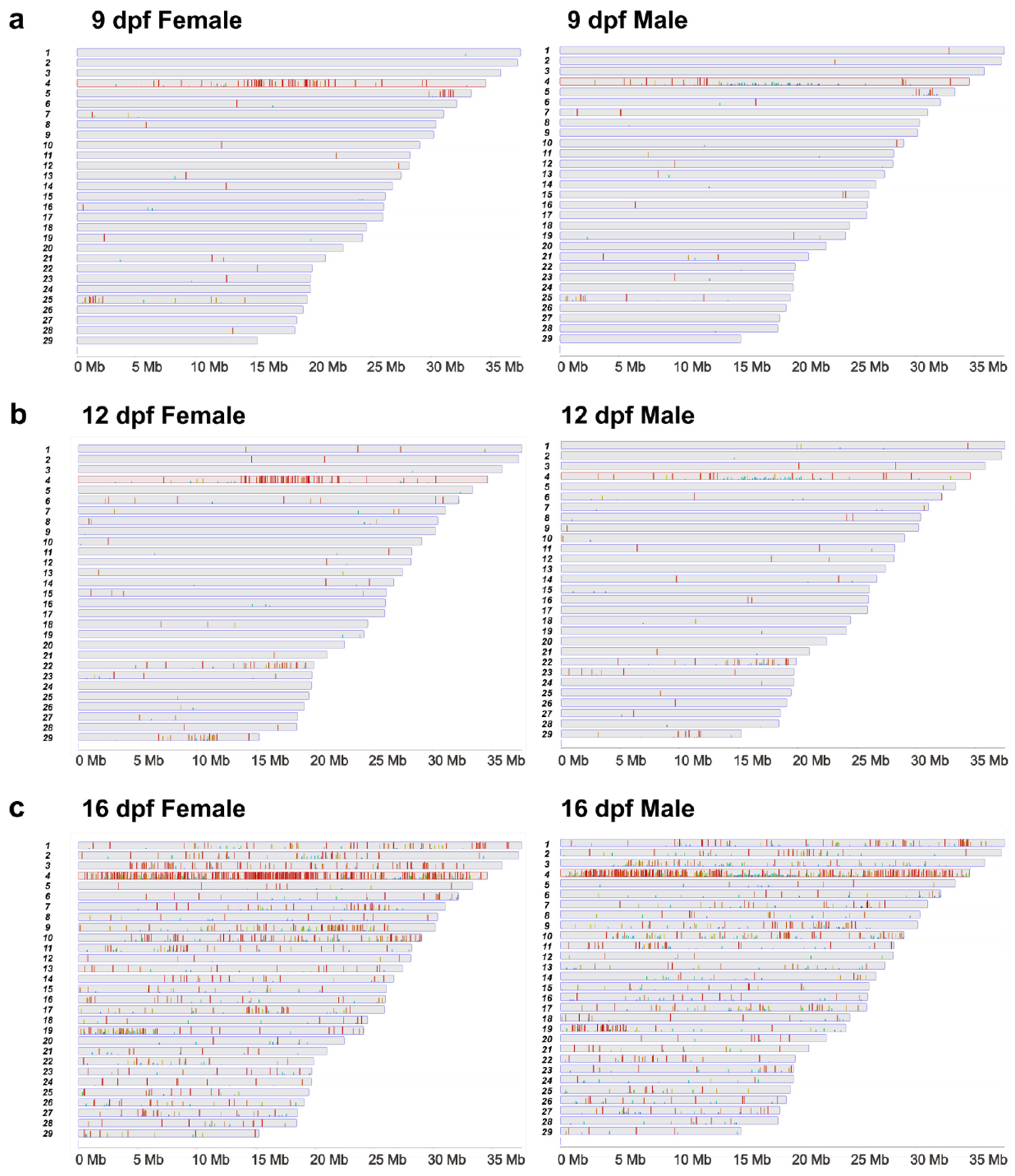
2.2. Differentially Methylated CpG Sites between Females and Males
2.3. Differentially Methylated CpG Sites between Females and Males
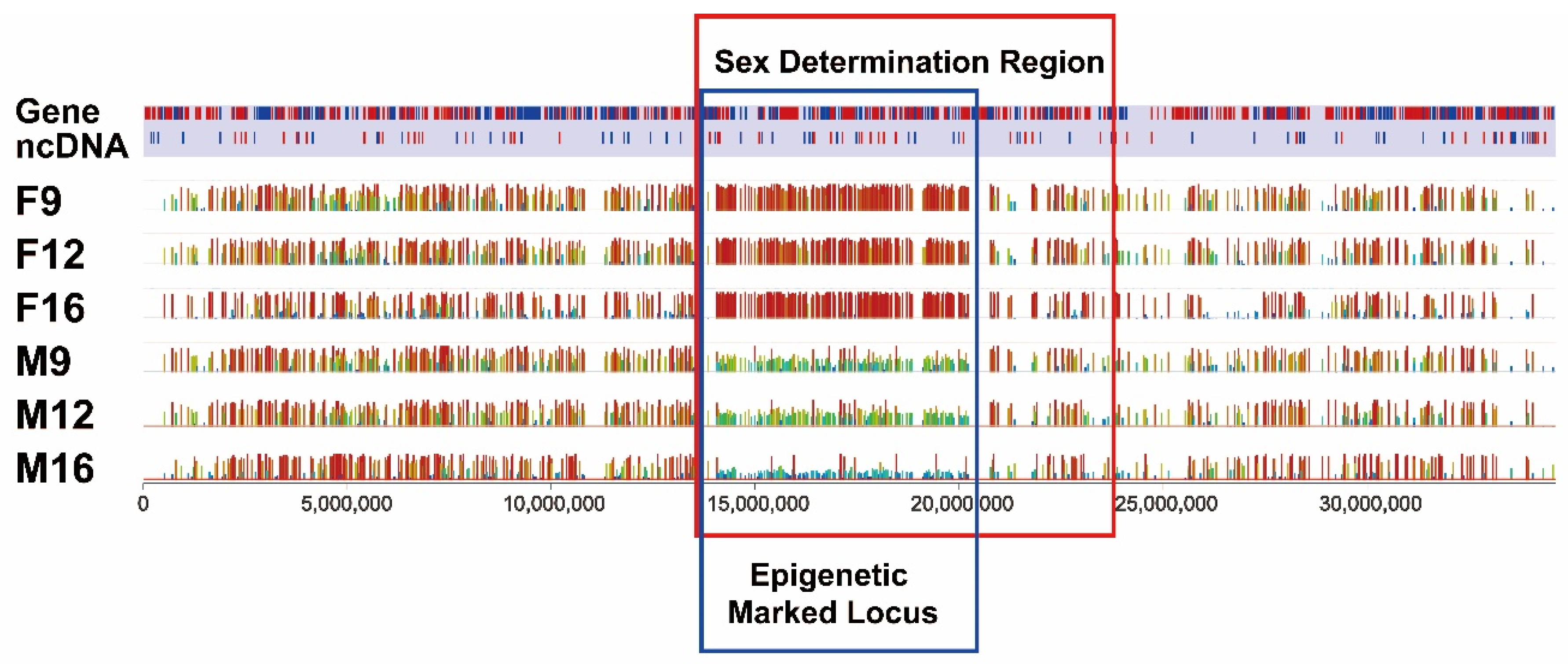
2.4. Identification of Differentially Methylated Genes within SDR
2.5. Differentially Expressed Genes within the SDR
3. Discussion
4. Materials and Methods
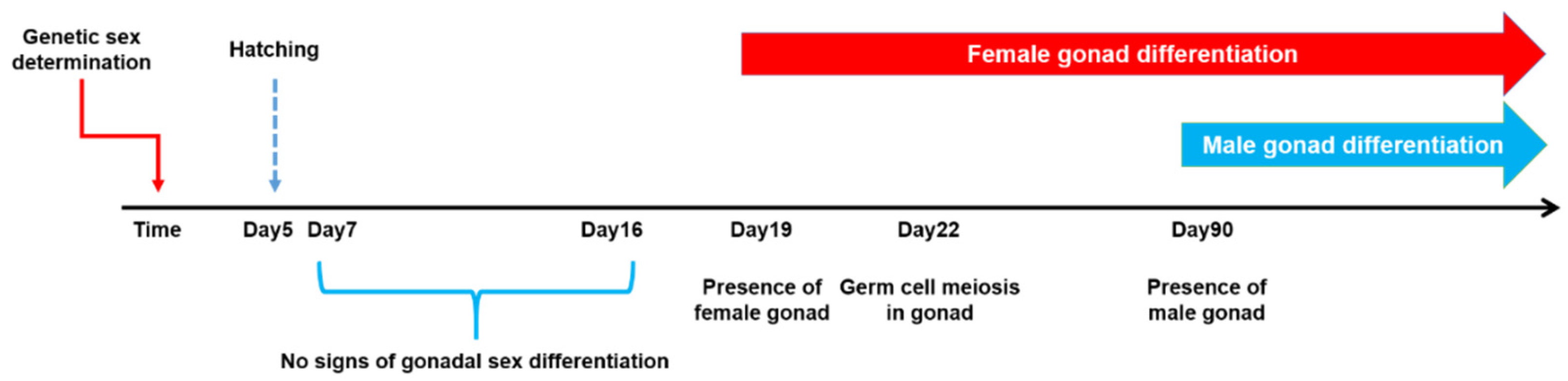
4.1. Experimental Fish
4.2. Tissue Collection and DNA Isolation
4.3. Bisulfite Conversion, Library Preparation, and Sequencing
4.4. Alignment of Reads and Calculation of the Methylation Level
4.5. Differential Methylation Analysis
4.6. High-Throughput RNA Sequencing
4.7. Identification of Sexually Dimorphic Expressed Genes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Berta, P.; Hawkins, J.B.; Sinclair, A.H.; Taylor, A.; Griffiths, B.L.; Goodfellow, P.N.; Fellous, M. Genetic evidence equating SRY and the testis-determining factor. Nature 1990, 348, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Ray-Chaudhuri, S.; Guha, A. X and neo-Y mechanism of sex-determination in the grasshopper, Thisiocetrus pulcher. Nature 1952, 169, 78–79. [Google Scholar] [CrossRef] [PubMed]
- Mesa, A.; García-Novo, P.; dos Santos, D. X1X2O (male)–X1X1X2X2 (female) chromosomal sex determining mechanism in the cricket Cicloptyloides americanus (Orthoptera, Grylloidea, Mogoplistidae). J. Orthoptera Res. 2002, 11, 87–90. [Google Scholar] [CrossRef]
- Cave, M.D. Absence of rDNA amplification in the uninucleolate oocyte of the cockroach Blattella germanica (Oorthoptera: Blattidae). J. Cell Biol. 1976, 71, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Kuroiwa, A.; Handa, S.; Nishiyama, C.; Chiba, E.; Yamada, F.; Abe, S.; Matsuda, Y. Additional copies of CBX2 in the genomes of males of mammals lacking SRY, the Amami spiny rat (Tokudaia osimensis) and the Tokunoshima spiny rat (Tokudaia tokunoshimensis). Chromosome Res. 2011, 19, 635–644. [Google Scholar] [CrossRef]
- Villeneuve, A.M.; Meyer, B.J. sdc-1: A link between sex determination and dosage compensation in C. elegans. Cell 1987, 48, 25–37. [Google Scholar] [CrossRef]
- Alves, A.L.; Oliveira, C.; Nirchio, M.; Granado, A.; Foresti, F. Karyotypic relationships among the tribes of Hypostominae (Siluriformes: Loricariidae) with description of XO sex chromosome system in a Neotropical fish species. Genetica 2006, 128, 1–9. [Google Scholar] [CrossRef]
- Steinmann-Zwicky, M.; Nöthiger, R. The hierarchical relation between X-chromosomes and autosomal sex determining genes in Drosophila. EMBO J. 1985, 4, 163–166. [Google Scholar] [CrossRef]
- Solovei, I.; Gaginskaya, E.; Hutchison, N.; Macgregor, H. Avian sex chromosomes in the lampbrush form: The ZW lampbrush bivalents from six species of bird. Chromosome Res. 1993, 1, 153–166. [Google Scholar] [CrossRef]
- Moreira-Filho, O.; Bertollo, L.A.; Galetti, P.M., Jr. Distribution of sex chromosome mechanisms in neotropical fish and description of a ZZ/ZW system in Parodon hilarii (Parodontidae). Caryologia 1993, 46, 115–125. [Google Scholar] [CrossRef]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.-L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed]
- Ravi, V.; Venkatesh, B. The divergent genomes of teleosts. Annu. Rev. Anim. Biosci. 2018, 6, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.; Avise, J. Evolutionary diversity and turn-over of sex determination in teleost fishes. Sex. Dev. 2009, 3, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Yano, A.; Guyomard, R.; Nicol, B.; Jouanno, E.; Quillet, E.; Klopp, C.; Cabau, C.; Bouchez, O.; Fostier, A.; Guiguen, Y. An immune-related gene evolved into the master sex-determining gene in rainbow trout, Oncorhynchus mykiss. Curr. Biol. 2012, 22, 1423–1428. [Google Scholar] [CrossRef]
- Hattori, R.S.; Murai, Y.; Oura, M.; Masuda, S.; Majhi, S.K.; Sakamoto, T.; Fernandino, J.I.; Somoza, G.M.; Yokota, M.; Strüssmann, C.A. A Y-linked anti-Müllerian hormone duplication takes over a critical role in sex determination. Proc. Natl. Acad. Sci. USA 2012, 109, 2955–2959. [Google Scholar] [CrossRef]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559. [Google Scholar] [CrossRef]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778–11783. [Google Scholar] [CrossRef]
- Myosho, T.; Otake, H.; Masuyama, H.; Matsuda, M.; Kuroki, Y.; Fujiyama, A.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Tracing the emergence of a novel sex-determining gene in medaka, Oryzias luzonensis. Genetics 2012, 191, 163–170. [Google Scholar] [CrossRef]
- Pandian, T.J.; Sheela, S.G. Hormonal induction of sex reversal in fish. Aquaculture 1995, 138, 1–22. [Google Scholar] [CrossRef]
- Patiño, R.; Davis, K.B.; Schoore, J.E.; Uguz, C.; Strüssmann, C.A.; Parker, N.C.; Simco, B.A.; Goudie, C.A. Sex differentiation of channel catfish gonads: Normal development and effects of temperature. J. Exp. Zool. 1996, 276, 209–218. [Google Scholar] [CrossRef]
- Baroiller, J.-F.; D’Cotta, H.; Saillant, E. Environmental effects on fish sex determination and differentiation. Sex. Dev. 2009, 3, 118–135. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Troadec, C.; Boualem, A.; Rajab, M.; Fernandez, R.; Morin, H.; Pitrat, M.; Dogimont, C.; Bendahmane, A. A transposon-induced epigenetic change leads to sex determination in melon. Nature 2009, 461, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Vyskot, B.; Araya, A.; Veuskens, J.; Negrutiu, I.; Mouras, A. DNA methylation of sex chromosomes in a dioecious plant, Melandrium album. Mol. Gen. Genet. 1993, 239, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-S.; Zhang, X.; Zhang, W.; Shen, D.; Wang, H.; Xia, Y.; Qiu, Y.; Song, J.; Wang, C.; Li, X. The association of changes in DNA methylation with temperature-dependent sex determination in cucumber. J. Exp. Bot. 2017, 68, 2899–2912. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, X.; Yu, Q.; Ming, R.; Jiang, J. DNA methylation and heterochromatinization in the male-specific region of the primitive Y chromosome of papaya. Genome Res. 2008, 18, 1938–1943. [Google Scholar] [CrossRef]
- Bräutigam, K.; Soolanayakanahally, R.; Champigny, M.; Mansfield, S.; Douglas, C.; Campbell, M.M.; Cronk, Q. Sexual epigenetics: Gender-specific methylation of a gene in the sex determining region of Populus balsamifera. Sci. Rep. 2017, 7, 45388. [Google Scholar] [CrossRef]
- Venegas, D.; Marmolejo-Valencia, A.; Valdes-Quezada, C.; Govenzensky, T.; Recillas-Targa, F.; Merchant-Larios, H. Dimorphic DNA methylation during temperature-dependent sex determination in the sea turtle Lepidochelys olivacea. Gen. Comp. Endocrinol. 2016, 236, 35–41. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Buemio, A.; Chu, R.; Vafaee, M.; Crews, D. Epigenetic control of gonadal aromatase (cyp19a1) in temperature-dependent sex determination of red-eared slider turtles. PLoS ONE 2013, 8, e63599. [Google Scholar] [CrossRef]
- Radhakrishnan, S.; Literman, R.; Mizoguchi, B.; Valenzuela, N. MeDIP-seq and nCpG analyses illuminate sexually dimorphic methylation of gonadal development genes with high historic methylation in turtle hatchlings with temperature-dependent sex determination. Epigenetics Chromatin 2017, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Parrott, B.B.; Kohno, S.; Cloy-McCoy, J.A.; Guillette, L.J., Jr. Differential incubation temperatures result in dimorphic DNA methylation patterning of the SOX9 and aromatase promoters in gonads of alligator (Alligator mississippiensis) embryos. Biol. Reprod. 2014, 90, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Martín, L.; Viñas, J.; Ribas, L.; Díaz, N.; Gutiérrez, A.; Di Croce, L.; Piferrer, F. DNA methylation of the gonadal aromatase (cyp19a) promoter is involved in temperature-dependent sex ratio shifts in the European sea bass. PLoS Genet. 2011, 7, e1002447. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadi, D.; Vandeputte, M.; Sánchez-Baizán, N.; Allal, F.; Piferrer, F. Dynamic epimarks in sex-related genes predict gonad phenotype in the European sea bass, a fish with mixed genetic and environmental sex determination. Epigenetics 2018, 13, 988–1011. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Li, Q.; Chen, S.; Zhang, P.; Lian, J.; Hu, Q.; Sun, B.; Jin, L.; Liu, S.; Wang, Z. Epigenetic modification and inheritance in sexual reversal of fish. Genome Res. 2014, 24, 604–615. [Google Scholar] [CrossRef]
- Sun, L.-X.; Wang, Y.-Y.; Zhao, Y.; Wang, H.; Li, N.; Ji, X.S. Global DNA methylation changes in Nile tilapia gonads during high temperature-induced masculinization. PLoS ONE 2016, 11, 8. [Google Scholar] [CrossRef]
- Wen, A.; You, F.; Sun, P.; Li, J.; Xu, D.; Wu, Z.; Ma, D.; Zhang, P. CpG methylation of dmrt1 and cyp19a promoters in relation to their sexual dimorphic expression in the Japanese flounder Paralichthys olivaceus. J. Fish Biol. 2014, 84, 193–205. [Google Scholar] [CrossRef]
- Metzger, D.C.; Schulte, P.M. The DNA methylation landscape of stickleback reveals patterns of sex chromosome evolution and effects of environmental salinity. Genome Biol. Evol. 2018, 10, 775–785. [Google Scholar] [CrossRef]
- Piferrer, F. Epigenetics of sex determination and differentiation in fish. In Sex Control in Aquaculture; Wang, H., Chen, S., Shen, Z.-G., Piferrer, F., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2018; pp. 65–83. [Google Scholar] [CrossRef]
- Piferrer, F.; Anastasiadi, D.; Valdivieso, A.; Sánchez, N.; Moraleda, J.; Ribas, L. The model of the conserved epigenetic regulation of sex. Front. Genet. 2019, 10, 857. [Google Scholar] [CrossRef]
- Ross, J.A.; Peichel, C.L. Molecular cytogenetic evidence of rearrangements on the Y chromosome of the threespine stickleback fish. Genetics 2008, 179, 2173–2182. [Google Scholar] [CrossRef]
- Peichel, C.L.; Ross, J.A.; Matson, C.K.; Dickson, M.; Grimwood, J.; Schmutz, J.; Myers, R.M.; Mori, S.; Schluter, D.; Kingsley, D.M. The master sex-determination locus in threespine sticklebacks is on a nascent Y chromosome. Curr. Biol. 2004, 14, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Ospina-Alvarez, N.; Piferrer, F. Temperature-dependent sex determination in fish revisited: Prevalence, a single sex ratio response pattern, and possible effects of climate change. PLoS ONE 2008, 3, e2837. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.B.; Simco, B.A.; Goudie, C.A.; Parker, N.C.; Cauldwell, W.; Snellgrove, R. Hormonal sex manipulation and evidence for female homogamety in channel catfish. Gen. Comp. Endocrinol. 1990, 78, 218–223. [Google Scholar] [CrossRef]
- Goudie, C.A.; Redner, B.D.; Simco, B.A.; Davis, K.B. Feminization of channel catfish by oral administration of steroid sex hormones. Trans. Am. Fish. Soc. 1983, 112, 670–672. [Google Scholar] [CrossRef]
- LeGrande, W.H.; Dunham, R.A.; Smitherman, R. Karyology of three species of catfishes (Ictaluridae: Ictalurus) and four hybrid combinations. Copeia 1984, 1984, 873–878. [Google Scholar] [CrossRef]
- Tiersch, T.; Simco, B.; Davis, K.; Wachtel, S. Molecular genetics of sex determination in channel catfish: Studies on SRY, ZFY, Bkm, and human telomeric repeats. Biol. Reprod. 1992, 47, 185–192. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Qin, Z.; Waldbieser, G.; Wang, R.; Sun, L.; Bao, L.; Danzmann, R.G.; Dunham, R.; Liu, Z. Construction of a high-density, high-resolution genetic map and its integration with BAC-based physical map in channel catfish. DNA Res. 2014, 22, 39–52. [Google Scholar] [CrossRef]
- Bao, L.; Tian, C.; Liu, S.; Zhang, Y.; Elaswad, A.; Yuan, Z.; Khalil, K.; Sun, F.; Yang, Y.; Zhou, T. The Y chromosome sequence of the channel catfish suggests novel sex determination mechanisms in teleost fish. BMC Biol. 2019, 17, 6. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, S.; Yao, J.; Bao, L.; Zhang, J.; Li, Y.; Jiang, C.; Sun, L.; Wang, R.; Zhang, Y. The channel catfish genome sequence provides insights into the evolution of scale formation in teleosts. Nat. Commun. 2016, 7, 11757. [Google Scholar] [CrossRef]
- Feng, S.; Cokus, S.J.; Zhang, X.; Chen, P.-Y.; Bostick, M.; Goll, M.G.; Hetzel, J.; Jain, J.; Strauss, S.H.; Halpern, M.E. Conservation and divergence of methylation patterning in plants and animals. Proc. Natl. Acad. Sci. USA 2010, 107, 8689–8694. [Google Scholar] [CrossRef]
- Wan, Z.Y.; Xia, J.H.; Lin, G.; Wang, L.; Lin, V.C.; Yue, G.H. Genome-wide methylation analysis identified sexually dimorphic methylated regions in hybrid tilapia. Sci. Rep. 2016, 6, 35903. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhuang, Z.-X.; Sun, Y.-Q.; Chen, Q.; Zheng, X.-Y.; Liang, Y.-T.; Mahboob, S.; Wang, Q.; Zhang, R.; Al-Ghanim, K.A. Changes in DNA methylation during epigenetic-associated sex reversal under low temperature in Takifugu rubripes. PLoS ONE 2019, 14, e0221641. [Google Scholar] [CrossRef] [PubMed]
- Kenchanmane Raju, S.K.; Ritter, E.J.; Niederhuth, C.E. Establishment, maintenance, and biological roles of non-CG methylation in plants. Essays Biochem. 2019, 63, 743–755. [Google Scholar] [PubMed]
- Sassaman, C.; Weeks, S.C. The genetic mechanism of sex determination in the conchostracan shrimp Eulimnadia texana. Am. Nat. 1993, 141, 314–328. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.; Double, M.C.; Orr, K.; Dawson, R.J. A DNA test to sex most birds. Mol. Ecol. 1998, 7, 1071–1075. [Google Scholar] [CrossRef]
- Valdez, M.B., Jr.; Mizutani, M.; Kinoshita, K.; Fujiwara, A.; Yazawa, H.; Shimada, K.; Namikawa, T.; Yamagata, T. Differential development of sex-related characters of chickens from the GSP and PNP/DO inbred lines after left ovariectomy. J. Reprod. Dev. 2010, 56, 154–161. [Google Scholar] [CrossRef]
- Piprek, R.P.; Podkowa, D.; Kubiak, J.Z. Global analysis of cell adhesion genes in sexually differentiating gonads of mouse. Acta Biol. Crac. Ser. Bot. Suppl. 2016, 58, 1. [Google Scholar]
- Liu, Z.; Miao, D.; Xia, Q.; Hermo, L.; Wing, S.S. Regulated expression of the ubiquitin protein ligase, E3Histone/LASU1/Mule/ARF-BP1/HUWE1, during spermatogenesis. Dev. Dyn. 2007, 236, 2889–2898. [Google Scholar] [CrossRef]
- Geng, X.; Sha, J.; Liu, S.; Bao, L.; Zhang, J.; Wang, R.; Yao, J.; Li, C.; Feng, J.; Sun, F. A genome-wide association study in catfish reveals the presence of functional hubs of related genes within QTLs for columnaris disease resistance. BMC Genom. 2015, 16, 196. [Google Scholar] [CrossRef]
- Liang, C.; He, Y.; Liu, Y.; Gao, Y.; Han, Y.; Li, X.; Zhao, Y.; Wang, J.; Zhang, J. Fluoride exposure alters the ultra-structure of sperm flagellum via reducing key protein expressions in testis. Chemosphere 2020, 246, 125772. [Google Scholar] [CrossRef]
- Sun, F.; Liu, S.; Gao, X.; Jiang, Y.; Perera, D.; Wang, X.; Li, C.; Sun, L.; Zhang, J.; Kaltenboeck, L. Male-biased genes in catfish as revealed by RNA-Seq analysis of the testis transcriptome. PLoS ONE 2013, 8, e68452. [Google Scholar] [CrossRef] [PubMed]
- Zhai, G.; Shu, T.; Yu, G.; Tang, H.; Shi, C.; Jia, J.; Lou, Q.; Dai, X.; Jin, X.; He, J. Augmentation of progestin signaling rescues testis organization and spermatogenesis in zebrafish with the depletion of androgen signaling. eLife 2022, 11, e66118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-L.; Wang, Z.-W.; Guo, X.-F.; Wu, J.-J.; Lu, W.-J.; Zhou, L.; Gui, J.-F. Construction of a high-density genetic linkage map and fine mapping of QTLs for growth and sex-related traits in red-tail catfish (Hemibagrus wyckioides). Aquaculture 2021, 531, 735892. [Google Scholar] [CrossRef]
- Lande-Diner, L.; Boyault, C.; Kim, J.Y.; Weitz, C.J. A positive feedback loop links circadian clock factor CLOCK-BMAL1 to the basic transcriptional machinery. Proc. Natl. Acad. Sci. USA 2013, 110, 16021–16026. [Google Scholar] [CrossRef]
- Shen, X.; Yan, H.; Jiang, J.; Li, W.; Xiong, Y.; Liu, Q.; Liu, Y. Profile of gene expression changes during estrodiol-17β-induced feminization in the Takifugu rubripes brain. BMC Genom. 2021, 22, 851. [Google Scholar] [CrossRef]
- Ma, L.; Buchold, G.M.; Greenbaum, M.P.; Roy, A.; Burns, K.H.; Zhu, H.; Han, D.Y.; Harris, R.A.; Coarfa, C.; Gunaratne, P.H. GASZ is essential for male meiosis and suppression of retrotransposon expression in the male germline. PLoS Genet. 2009, 5, e1000635. [Google Scholar] [CrossRef]
- Luitjens, C.; Gallegos, M.; Kraemer, B.; Kimble, J.; Wickens, M. CPEB proteins control two key steps in spermatogenesis in C. elegans. Genes Dev. 2000, 14, 2596–2609. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Liu, Z.; Zhang, L.; Zhang, W. Epigenetic modifications during sex change repress gonadotropin stimulation of cyp19a1a in a teleost ricefield eel (Monopterus albus). Endocrinology 2013, 154, 2881–2890. [Google Scholar] [CrossRef]
- Dunham, R.A.; Lambert, D.M.; Argue, B.J.; Ligeon, C.; Yant, D.R.; Liu, Z. Comparison of manual stripping and pen spawning for production of channel catfish × blue catfish hybrids and aquarium spawning of channel catfish. N. Am. J. Aquac. 2000, 62, 260–265. [Google Scholar] [CrossRef]
- Ninwichian, P.; Peatman, E.; Perera, D.; Liu, S.; Kucuktas, H.; Dunham, R.; Liu, Z. Identification of a sex-linked marker for channel catfish. Anim. Genet. 2012, 43, 476–477. [Google Scholar] [CrossRef]
- Khoueiry, R.; Sohni, A.; Thienpont, B.; Luo, X.; Velde, J.V.; Bartoccetti, M.; Boeckx, B.; Zwijsen, A.; Rao, A.; Lambrechts, D. Lineage-specific functions of TET1 in the postimplantation mouse embryo. Nat. Genet. 2017, 49, 1061. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Krueger, F.; Andrews, S.R. Bismark: A flexible aligner and methylation caller for Bisulfite-Seq applications. Bioinformatics 2011, 27, 1571–1572. [Google Scholar] [CrossRef] [PubMed]
- Akalin, A.; Kormaksson, M.; Li, S.; Garrett-Bakelman, F.E.; Figueroa, M.E.; Melnick, A.; Mason, C.E. methylKit: A comprehensive R package for the analysis of genome-wide DNA methylation profiles. Genome Biol. 2012, 13, R87. [Google Scholar] [CrossRef]
- Wagih, O. ggseqlogo: A versatile R package for drawing sequence logos. Bioinformatics 2017, 33, 3645–3647. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Methylated CpG | Methylated CHG | Methylated CHH |
|---|---|---|---|
| Female | |||
| 9 dpf | 258,326,088 (77.1%) | 2,817,165 (0.4%) | 7,706,815 (0.3%) |
| 12 dpf | 217,250,994 (77.2%) | 2,156,312 (0.3%) | 6,284,900 (0.3%) |
| 16 dpf | 284,031,704 (75.7%) | 2,814,003 (0.3%) | 8,120,484 (0.3%) |
| Male | |||
| 9 dpf | 253,621,156 (77.5%) | 2,718,707 (0.4%) | 7,540,415 (0.3%) |
| 12 dpf | 277,521,207 (78.4%) | 3,124,962 (0.4%) | 8,583,702 (0.4%) |
| 16 dpf | 290,874,272 (74.5%) | 2,922,346 (0.3%) | 8,239,077 (0.3%) |
| Chr | 9 dpf | 12 dpf | 16dpf | |||
|---|---|---|---|---|---|---|
| ♀ Hyper | ♂ Hyper | ♀ Hyper | ♂ Hyper | ♀ Hyper | ♂ Hyper | |
| 1 | 0 | 1 | 3 | 3 | 62 | 63 |
| 2 | 0 | 1 | 2 | 2 | 32 | 26 |
| 3 | 0 | 0 | 0 | 0 | 81 | 52 |
| 4 | 59 | 21 | 64 | 21 | 569 | 402 |
| 5 | 9 | 6 | 0 | 1 | 9 | 4 |
| 6 | 1 | 1 | 9 | 3 | 16 | 11 |
| 7 | 2 | 3 | 4 | 1 | 28 | 18 |
| 8 | 1 | 0 | 3 | 2 | 10 | 11 |
| 9 | 0 | 0 | 0 | 1 | 90 | 39 |
| 10 | 1 | 1 | 1 | 1 | 70 | 64 |
| 11 | 1 | 1 | 1 | 2 | 30 | 25 |
| 12 | 1 | 1 | 1 | 2 | 6 | 4 |
| 13 | 1 | 1 | 2 | 0 | 18 | 11 |
| 14 | 1 | 0 | 2 | 2 | 19 | 9 |
| 15 | 0 | 2 | 4 | 0 | 12 | 11 |
| 16 | 2 | 1 | 1 | 2 | 20 | 12 |
| 17 | 0 | 0 | 0 | 0 | 31 | 38 |
| 18 | 0 | 0 | 4 | 1 | 6 | 10 |
| 19 | 1 | 2 | 2 | 1 | 80 | 48 |
| 20 | 0 | 0 | 0 | 0 | 3 | 8 |
| 21 | 2 | 4 | 1 | 1 | 7 | 9 |
| 22 | 1 | 0 | 28 | 18 | 12 | 17 |
| 23 | 1 | 1 | 4 | 6 | 10 | 10 |
| 24 | 0 | 0 | 0 | 1 | 9 | 4 |
| 25 | 16 | 8 | 0 | 1 | 19 | 10 |
| 26 | 0 | 0 | 1 | 1 | 18 | 12 |
| 27 | 0 | 0 | 2 | 1 | 22 | 15 |
| 28 | 1 | 0 | 2 | 0 | 12 | 4 |
| 29 | 0 | 0 | 25 | 7 | 9 | 4 |
| Total | 101 | 55 | 166 | 81 | 1310 | 951 |
| Stage | Differentially Methylated CpG Sites | Differentially Methylated Genes | ||||
|---|---|---|---|---|---|---|
| Whole Genome | Chr4 | SDR | Whole Genome | Chr4 | SDR | |
| 9 dpf | 156 | 80 | 47 | 96 | 43 | 23 |
| 12 dpf | 247 | 85 | 60 | 153 | 50 | 31 |
| 16 dpf | 2261 | 971 | 316 | 1120 | 308 | 77 |
| Gene | Differential Expression FPKM Male/FPKM Female | Differentially Methylation | ||||
|---|---|---|---|---|---|---|
| 7 d | 12 d | 17 d | 9 d | 12 d | 16 d | |
| hydin-1 | 8.96 | 15.05 | 41.56 | nd | X Hyper | X Hyper |
| LOC108264027 | 2.08 | 2.29 | nd | nd | ♀ Hyper | ♀ Hyper |
| gtf2ird2b | 2.83 | 2.35 | 2.70 | nd | ♀ Hyper | ♀ Hyper |
| kcnj1 | nd | nd | 2.75 | nd | nd | nd |
| LOC108264687 | nd | nd | 2.24 | nd | nd | nd |
| actrt3 | nd | nd | 2.07 | nd | nd | nd |
| cipc | −2.18 | −2.78 | −2.61 | nd | ♀ Hyper | ♀ Hyper |
| LOC108264260 | nd | nd | −2.29 | nd | nd | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhou, T.; Liu, Y.; Tian, C.; Bao, L.; Wang, W.; Zhang, Y.; Liu, S.; Shi, H.; Tan, S.; et al. Identification of an Epigenetically Marked Locus within the Sex Determination Region of Channel Catfish. Int. J. Mol. Sci. 2022, 23, 5471. https://doi.org/10.3390/ijms23105471
Yang Y, Zhou T, Liu Y, Tian C, Bao L, Wang W, Zhang Y, Liu S, Shi H, Tan S, et al. Identification of an Epigenetically Marked Locus within the Sex Determination Region of Channel Catfish. International Journal of Molecular Sciences. 2022; 23(10):5471. https://doi.org/10.3390/ijms23105471
Chicago/Turabian StyleYang, Yujia, Tao Zhou, Yang Liu, Changxu Tian, Lisui Bao, Wenwen Wang, Yu Zhang, Shikai Liu, Huitong Shi, Suxu Tan, and et al. 2022. "Identification of an Epigenetically Marked Locus within the Sex Determination Region of Channel Catfish" International Journal of Molecular Sciences 23, no. 10: 5471. https://doi.org/10.3390/ijms23105471