
The Distinct Role of the HDL Receptor SR-BI in Cholesterol Homeostasis of Human Placental Arterial and Venous Endothelial Cells

 , , ,
, , ,  , and
, and 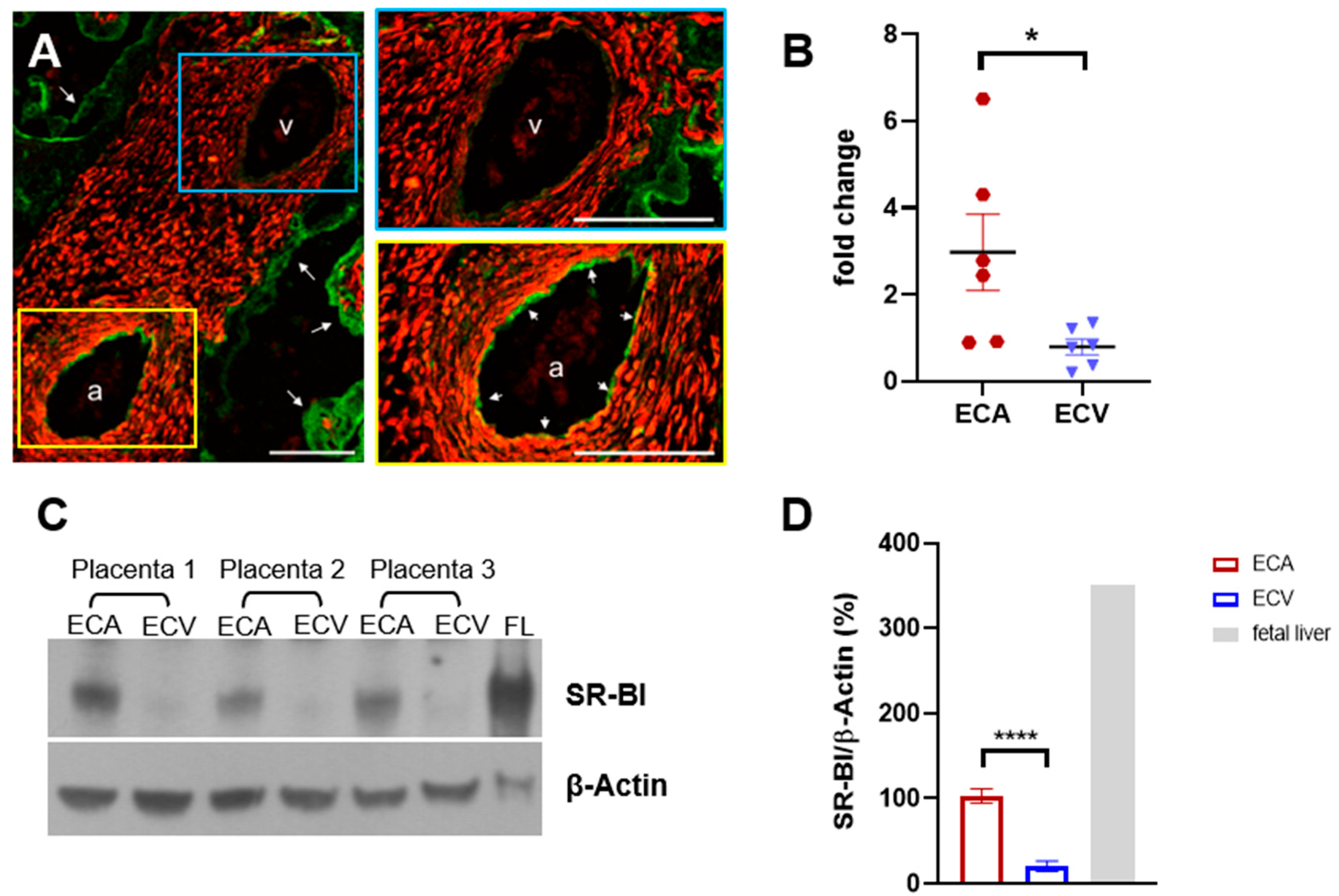{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SR-BI Protein Is Elevated in ECA Compared to ECV
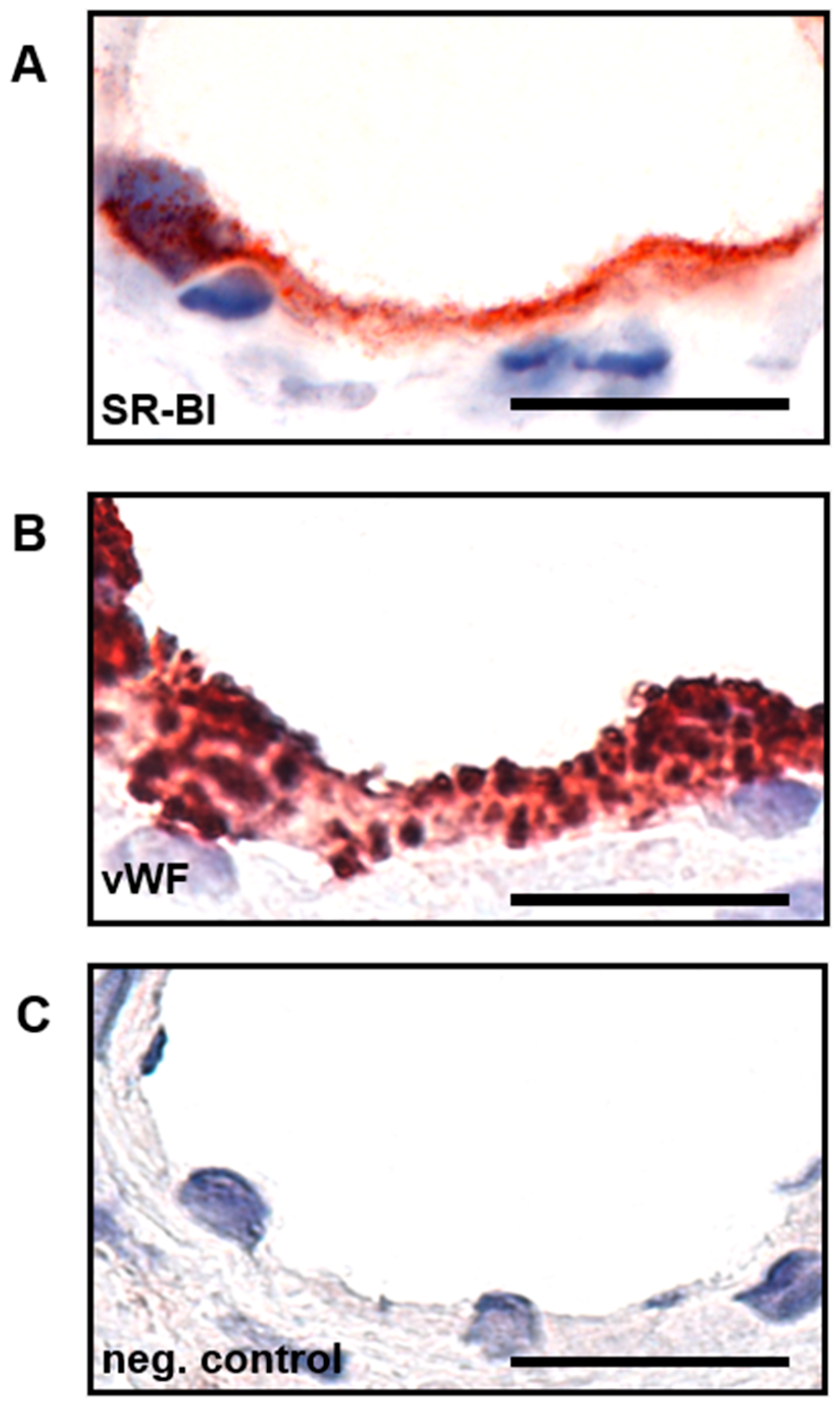
2.2. SR-BI Is Predominantly Expressed on the Apical Side of Placental Endothelial Cells
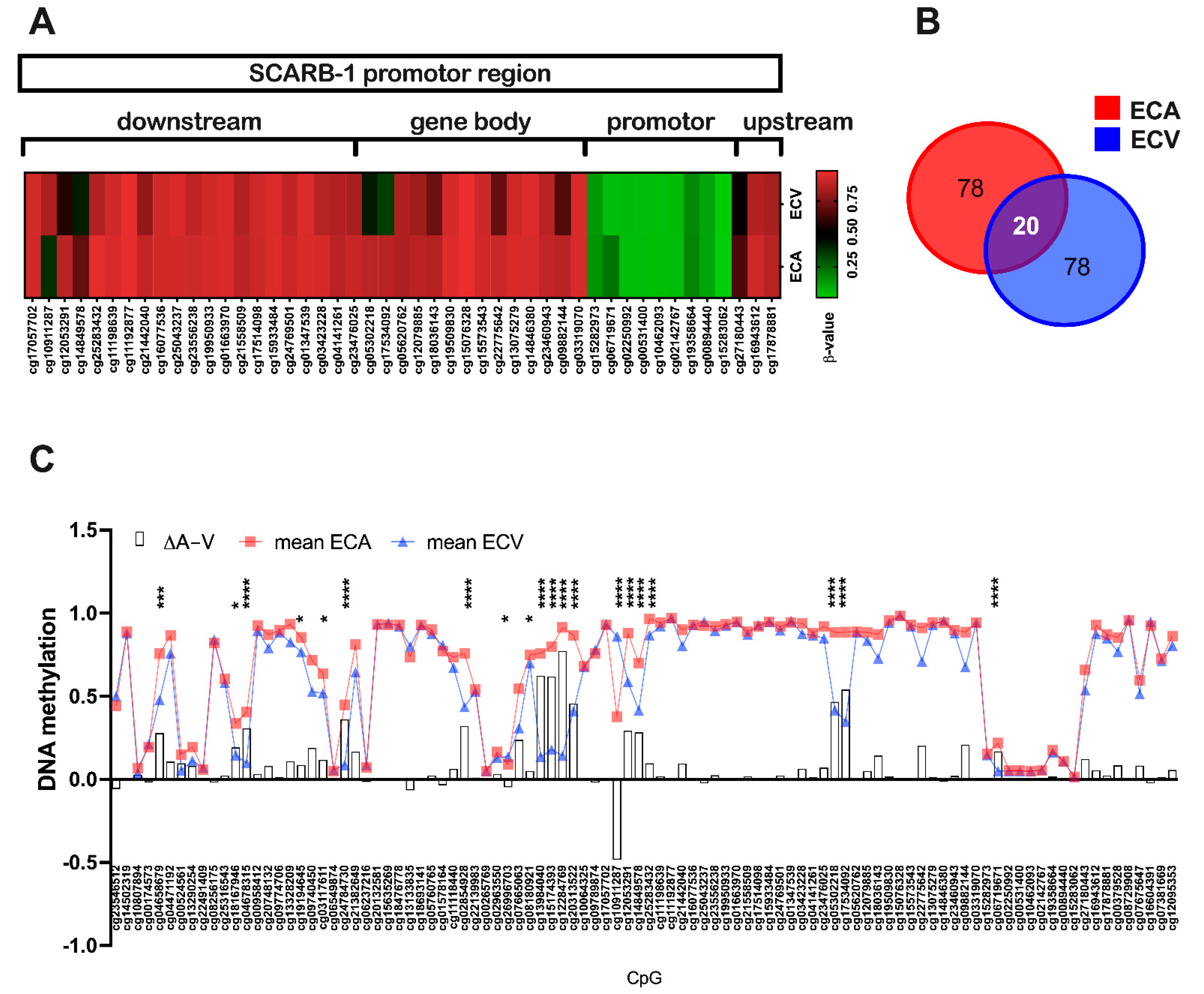
2.3. SR-BI Promoter Methylation Differs between ECA and ECV
2.4. GATA3 Transcription Factor Might Contribute to Differential SR-BI Expression
2.5. LXR Transcription Is Not Involved in SR-BI Regulation between Vascular Beds
2.6. PDZ Domain-Containing Protein PDZK1 Is Equally Expressed in ECA and ECV
2.7. Neither Oxygen nor Shear Stress Regulate SR-BI Protein in Primary Placental Endothelial Cells
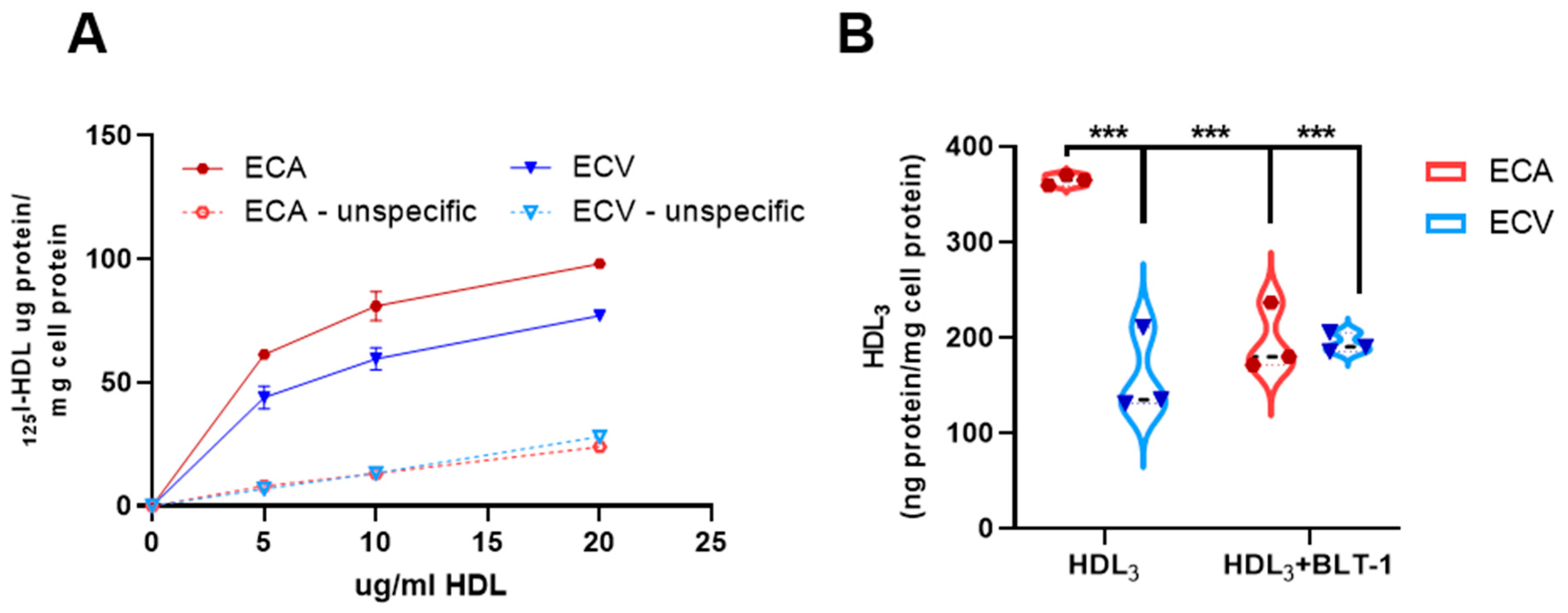
2.8. Selective HDL CE-Uptake Is Impaired in ECV Compared to ECA
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Immunohistochemistry
4.3. Double-Fluorescence Microscopy
4.4. Light Microscopy
4.5. Isolation of Human Feto-Placental Endothelial Cells from Arteries and Veins
4.6. Cell Culture Experiments
4.7. Simulation of Shear Stress
4.8. Immunoblot Analyses
4.9. Quantitative RT-PCR
4.10. Genome-Scale DNA Methylation Analysis: Data Acquisition and Processing
4.11. HDL-Isolation and Labelling
4.12. HDL Lipid-Uptake
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patton, S. Correlative relationship of cholesterol and sphingomyelin in cell membranes. J. Theor. Biol. 1970, 29, 489–491. [Google Scholar] [CrossRef]
- Ohvo, H.; Olsio, C.; Slotte, J.P. Effects of sphingomyelin and phosphatidylcholine degradation on cyclodextrin-mediated cholesterol efflux in cultured fibroblasts. Biochim. Biophys. Acta 1997, 1349, 131–141. [Google Scholar] [CrossRef]
- Fielding, C.J.; Fielding, P.E. Membrane cholesterol and the regulation of signal transduction. Biochem. Soc. Trans. 2004, 32, 65–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, E.J.; Graf, G.A.; McNiven, M.A.; Sessa, W.C.; Engelman, J.A.; Scherer, P.E.; Okamoto, T.; Lisanti, M.P. Caveolins, liquid-ordered domains, and signal transduction. Mol. Cell. Biol. 1999, 19, 7289–7304. [Google Scholar] [CrossRef] [Green Version]
- Lopez, D.; McLean, M.P. Estrogen regulation of the scavenger receptor class B gene: Anti-atherogenic or steroidogenic, is there a priority? Mol. Cell. Endocrinol. 2006, 247, 22–33. [Google Scholar] [CrossRef]
- Porter, J.A.; Young, K.E.; Beachy, P.A. Cholesterol Modification of Hedgehog Signaling Proteins in Animal Development. Science 1996, 274, 255. [Google Scholar] [CrossRef]
- Cooper, M.K.; Wassif, C.A.; Krakowiak, P.A.; Taipale, J.; Gong, R.; Kelley, R.I.; Porter, F.D.; Beachy, P.A. A defective response to Hedgehog signaling in disorders of cholesterol biosynthesis. Nat. Genet. 2003, 33, 508–513. [Google Scholar] [CrossRef]
- Carr, B.R.; Simpson, E.R. Cholesterol Synthesis in Human Fetal Tissues. J. Clin. Endocrinol. Metab. 1982, 55, 447–452. [Google Scholar] [CrossRef]
- Cummings, S.W.; Hatley, W.; Simpson, E.R.; Ohashi, M. The binding of high and low density lipoproteins to human placental membrane fractions. J. Clin. Endocrinol. Metab. 1982, 54, 903–908. [Google Scholar] [CrossRef]
- Wyne, K.L.; Woollett, L.A. Transport of maternal LDL and HDL to the fetal membranes and placenta of the Golden Syrian hamster is mediated by receptor-dependent and receptor-independent processes. J. Lipid Res. 1998, 39, 518–530. [Google Scholar] [CrossRef]
- Schmid, K.E.; Davidson, W.S.; Myatt, L.; Woollett, L.A. Transport of cholesterol across a BeWo cell monolayer: Implications for net transport of sterol from maternal to fetal circulation. J. Lipid Res. 2003, 44, 1909–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woollett, L. a Maternal cholesterol in fetal development: Transport of cholesterol from the maternal to the fetal circulation. Am. J. Clin. Nutr. 2005, 82, 1155–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasaka, H.; Chiba, H.; Kikuta, H.; Akita, H.; Takahashi, Y.; Yanai, H.; Hui, S.-P.; Fuda, H.; Fujiwara, H.; Kobayashi, K. Unique character and metabolism of high density lipoprotein (HDL) in fetus. Atherosclerosis 2002, 161, 215–223. [Google Scholar] [CrossRef]
- Sreckovic, I.; Birner-Gruenberger, R.; Obrist, B.; Stojakovic, T.; Scharnagl, H.; Holzer, M.; Scholler, M.; Philipose, S.; Marsche, G.; Lang, U.; et al. Distinct composition of human fetal HDL attenuates its anti-oxidative capacity. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2013, 1831, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Augsten, M.; Hackl, H.; Ebner, B.; Chemelli, A.; Glatter, O.; Marsche, G.; Lang, U.; Desoye, G.; Wadsack, C.; Lang, U.; et al. Fetal HDL/apoE: A novel regulator of gene expression in human placental endothelial cells. Physiol. Genom. 2011, 43, 1255–1262. [Google Scholar] [CrossRef] [Green Version]
- Al-Jarallah, A.; Trigatti, B.L. A role for the scavenger receptor, class B type I in high density lipoprotein dependent activation of cellular signaling pathways. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2010, 1801, 1239–1248. [Google Scholar] [CrossRef]
- Acton, S.; Rigotti, A.; Landschulz, K.T.; Xu, S.; Hobbs, H.H.; Krieger, M. Identification of Scavenger Receptor SR-BI as a High Density Lipoprotein Receptor. Science 1996, 271, 518. [Google Scholar] [CrossRef]
- Krieger, M. Scavenger receptor class B type I is a multiligand HDL receptor that influences diverse physiologic systems. J. Clin. Investig. 2001, 108, 793–797. [Google Scholar] [CrossRef]
- Rigotti, A.; Miettinen, H.E.; Krieger, M. The Role of the High-Density Lipoprotein Receptor SR-BI in the Lipid Metabolism of Endocrine and Other Tissues. Endocr. Rev. 2003, 24, 357–387. [Google Scholar] [CrossRef] [Green Version]
- Stein, Y.; Dabach, Y.; Hollander, G.; Halperin, G.; Stein, O. Metabolism of HDL-cholesteryl ester in the rat, studied with a nonhydrolyzable analog, cholesteryl linoleyl ether. Biochim. Biophys. Acta-Lipids Lipid Metab. 1983, 752, 98–105. [Google Scholar] [CrossRef]
- Glass, C.; Pittman, R.C.; Weinstein, D.B.; Steinberg, D. Dissociation of tissue uptake of cholesterol ester from that of apoprotein A-I of rat plasma high density lipoprotein: Selective delivery of cholesterol ester to liver, adrenal, and gonad. Proc. Natl. Acad. Sci. USA 1983, 80, 5435–5439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing-Fei, D.D.; Spin, J.M.; Anya, T.; Aditya, V.; Amir, B.-D.; Zohar, Y.; Phil, T.; Laurakay, B.; Thomas, Q. Molecular Signatures Determining Coronary Artery and Saphenous Vein Smooth Muscle Cell Phenotypes. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1058–1065. [Google Scholar] [CrossRef] [Green Version]
- Lang, I.; Schweizer, A.; Hiden, U.; Ghaffari-Tabrizi, N.; Hagendorfer, G.; Bilban, M.; Pabst, M.A.; Korgun, E.T.; Dohr, G.; Desoye, G. Human fetal placental endothelial cells have a mature arterial and a juvenile venous phenotype with adipogenic and osteogenic differentiation potential. Differentiation 2008, 76, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.D.; Geary, R.L.; McManus, B.; Schwartz, S.M. A comparison of aorta and vena cava medial message expression by cDNA array analysis identifies a set of 68 consistently differentially expressed genes, all in aortic media. Circ. Res. 2000, 87, 623–631. [Google Scholar] [CrossRef] [Green Version]
- Hatzopoulos, A.K.; Rigotti, A.; Rosenberg, R.D.; Krieger, M. Temporal and spatial pattern of expression of the HDL receptor SR-BI during murine embryogenesis. J. Lipid Res. 1998, 39, 495–508. [Google Scholar] [CrossRef]
- Napoli, C.; Witztum, J.L.; de Nigris, F.; Palumbo, G.; D’Armiento, F.P.; Palinski, W. Intracranial Arteries of Human Fetuses Are More Resistant to Hypercholesterolemia-Induced Fatty Streak Formation Than Extracranial Arteries. Circulation 1999, 99, 2003–2010. [Google Scholar] [CrossRef] [Green Version]
- Napoli, C.; D’Armiento, F.P.; Mancini, F.P.; Postiglione, A.; Witztum, J.L.; Palumbo, G.; Palinski, W. Fatty streak formation occurs in human fetal aortas and is greatly enhanced by maternal hypercholesterolemia. Intimal accumulation of low density lipoprotein and its oxidation precede monocyte recruitment into early atherosclerotic lesions. J. Clin. Investig. 1997, 100, 2680–2690. [Google Scholar] [CrossRef]
- Stangl, H.; Hyatt, M.; Hobbs, H.H. Transport of Lipids from High and Low Density Lipoproteins via Scavenger Receptor-BI. J. Biol. Chem. 1999, 274, 32692–32698. [Google Scholar] [CrossRef] [Green Version]
- Stefulj, J.; Panzenboeck, U.; Becker, T.; Hirschmugl, B.; Schweinzer, C.; Lang, I.; Marsche, G.; Sadjak, A.; Lang, U.; Desoye, G.; et al. Human endothelial cells of the placental barrier efficiently deliver cholesterol to the fetal circulation via ABCA1 and ABCG1. Circ. Res. 2009, 104, 600–608. [Google Scholar] [CrossRef]
- Sun, Y.; Kopp, S.; Strutz, J.; Gali, C.C.; Zandl-Lang, M.; Fanaee-Danesh, E.; Kirsch, A.; Cvitic, S.; Frank, S.; Saffery, R.; et al. Gestational diabetes mellitus modulates cholesterol homeostasis in human fetoplacental endothelium. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2018, 1863, 968–979. [Google Scholar] [CrossRef]
- Palinski, W.; Napoli, C. Pathophysiological Events during Pregnancy Influence the Development of Atherosclerosis in Humans. Trends Cardiovasc. Med. 1999, 9, 205–214. [Google Scholar] [CrossRef]
- Gnasso, A.; Carallo, C.; Irace, C.; Spagnuolo, V.; De Novara, G.; Mattioli Pier, L.; Pujia, A. Association Between Intima-Media Thickness and Wall Shear Stress in Common Carotid Arteries in Healthy Male Subjects. Circulation 1996, 94, 3257–3262. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2012, 38, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [Green Version]
- Bogdanović, O.; Lister, R. DNA methylation and the preservation of cell identity. Curr. Opin. Genet. Dev. 2017, 46, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhang, X.; Huang, C.-C.; Jafari, N.; Kibbe, W.A.; Hou, L.; Lin, S.M. Comparison of Beta-value and M-value methods for quantifying methylation levels by microarray analysis. BMC Bioinform. 2010, 11, 587. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Suehiro, J.I.; Kanki, Y.; Kawai, Y.; Inoue, K.; Daida, H.; Yano, K.; Ohhashi, T.; Oettgen, P.; Aird, W.C.; et al. Critical role for GATA3 in mediating Tie2 expression and function in large vessel endothelial cells. J. Biol. Chem. 2009, 284, 29109–29124. [Google Scholar] [CrossRef] [Green Version]
- Minami, T.; Aird, W.C. Thrombin Stimulation of the Vascular Cell Adhesion Molecule-1 Promoter in Endothelial Cells Is Mediated by Tandem Nuclear Factor-κB and GATA Motifs. J. Biol. Chem. 2001, 276, 47632–47641. [Google Scholar] [CrossRef] [Green Version]
- Umetani, M.; Mataki, C.; Minegishi, N.; Yamamoto, M.; Hamakubo, T.; Kodama, T. Function of GATA transcription factors in induction of endothelial vascular cell adhesion molecule-1 by tumor necrosis factor-alpha. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Kocher, O.; Yesilaltay, A.; Cirovic, C.; Pal, R.; Rigotti, A.; Krieger, M. Targeted Disruption of the PDZK1 Gene in Mice Causes Tissue-specific Depletion of the High Density Lipoprotein Receptor Scavenger Receptor Class B Type I and Altered Lipoprotein Metabolism. J. Biol. Chem. 2003, 278, 52820–52825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikemoto, M.; Arai, H.; Feng, D.; Tanaka, K.; Aoki, J.; Dohmae, N.; Takio, K.; Adachi, H.; Tsujimoto, M.; Inoue, K. Identification of a PDZ-domain-containing protein that interacts with the scavenger receptor class B type I. Proc. Natl. Acad. Sci. USA 2000, 97, 6538–6543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonika, S.; Chieko, M.; Shaul, P.W. Signaling by the High-Affinity HDL Receptor Scavenger Receptor B Type I. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Steiner, H.; Schneider, K.-T. Dopplersonographie in der Gynäkologie und Geburtshilfe; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 978-3-540-72371-4. [Google Scholar]
- Williams, D.L.; de la Llera-Moya, M.; Thuahnai, S.T.; Lund-Katz, S.; Connelly, M.A.; Azhar, S.; Anantharamaiah, G.M.; Phillips, M.C. Binding and Cross-linking Studies Show That Scavenger Receptor BI Interacts with Multiple Sites in Apolipoprotein A-I and Identify the Class A Amphipathic α-Helix as a Recognition Motif. J. Biol. Chem. 2000, 275, 18897–18904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temel, R.E.; Walzem, R.L.; Banka, C.L.; Williams, D.L. Apolipoprotein A-I Is Necessary for thein Vivo Formation of High Density Lipoprotein Competent for Scavenger Receptor BI-mediated Cholesteryl Ester-selective Uptake. J. Biol. Chem. 2002, 277, 26565–26572. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Heesom, K.; Phillips, R.; Chen, L.; Trinder, J.; Bernal, A.L. Low abundance plasma proteins in labour. Reproduction 2012, 144, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Xing-Fei, D.D.; Anya, T.; Aditya, V.; Amir, B.-D.; Ramendra, K.; Ivette, E.; Raymond, T.; Robert, K.; Zohar, Y.; Laurakay, B.; et al. Differences in Vascular Bed Disease Susceptibility Reflect Differences in Gene Expression Response to Atherogenic Stimuli. Circ. Res. 2006, 98, 200–208. [Google Scholar] [CrossRef]
- Wang, H.U.; Chen, Z.-F.; Anderson, D.J. Molecular Distinction and Angiogenic Interaction between Embryonic Arteries and Veins Revealed by ephrin-B2 and Its Receptor Eph-B4. Cell 1998, 93, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Clapp, C.; Thebault, S.; Jeziorski, M.C.; Martínez De La Escalera, G. Peptide Hormone Regulation of Angiogenesis. Physiol. Rev. 2009, 89, 1177–1215. [Google Scholar] [CrossRef]
- Hu, Y.-W.; Wang, Q.; Ma, X.; Li, X.-X.; Liu, X.-H.; Xiao, J.; Liao, D.-F.; Xiang, J.; Tang, C.-K. TGFβ1 Up-Regulates Expression of ABCA1, ABCG1 and SR-BI through Liver X Receptor α Signaling Pathway in THP-1 Macrophage-Derived Foam Cells. J. Atheroscler. Thromb. 2010, 17, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Charlet, J.; Duymich, C.E.; Lay, F.D.; Mundbjerg, K.; Dalsgaard Sørensen, K.; Liang, G.; Jones, P.A. Bivalent Regions of Cytosine Methylation and H3K27 Acetylation Suggest an Active Role for DNA Methylation at Enhancers. Mol. Cell 2016, 62, 422–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aran, D.; Hellman, A. DNA methylation of transcriptional enhancers and cancer predisposition. Cell 2013, 154, 11–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hon, G.C.; Rajagopal, N.; Shen, Y.; McCleary, D.F.; Yue, F.; Dang, M.D.; Ren, B. Epigenetic memory at embryonic enhancers identified in DNA methylation maps from adult mouse tissues. Nat. Genet. 2013, 45, 1198–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholler, M.; Wadsack, C.; Metso, J.; Chirackal Manavalan, A.P.; Sreckovic, I.; Schweinzer, C.; Hiden, U.; Jauhiainen, M.; Desoye, G.; Panzenboeck, U. Phospholipid transfer protein is differentially expressed in human arterial and venous placental endothelial cells and enhances cholesterol efflux to fetal HDL. J. Clin. Endocrinol. Metab. 2012, 97, 2466–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grefhorst, A.; Oosterveer, M.H.; Brufau, G.; Boesjes, M.; Kuipers, F.; Groen, A.K. Pharmacological LXR activation reduces presence of SR-B1 in liver membranes contributing to LXR-mediated induction of HDL-cholesterol. Atherosclerosis 2012, 222, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Shen, W.-J.; Kraemer, F.B.; Azhar, S. MicroRNAs 125a and 455 Repress Lipoprotein-Supported Steroidogenesis by Targeting Scavenger Receptor Class B Type I in Steroidogenic Cells. Mol. Cell. Biol. 2012, 32, 5035–5045. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Jia, X.-J.; Jiang, H.-J.; Du, Y.; Yang, F.; Si, S.-Y.; Hong, B. MicroRNAs 185, 96, and 223 Repress Selective High-Density Lipoprotein Cholesterol Uptake through Posttranscriptional Inhibition. Mol. Cell. Biol. 2013, 33, 1956–1964. [Google Scholar] [CrossRef] [Green Version]
- Ren, K.; Zhu, X.; Zheng, Z.; Mo, Z.C.; Peng, X.S.; Zeng, Y.Z.; Ou, H.X.; Zhang, Q.H.; Qi, H.Z.; Zhao, G.J.; et al. MicroRNA-24 aggravates atherosclerosis by inhibiting selective lipid uptake from HDL cholesterol via the post-transcriptional repression of scavenger receptor class B type I. Atherosclerosis 2018, 270, 57–67. [Google Scholar] [CrossRef]
- Nye, G.A.; Ingram, E.; Johnstone, E.D.; Jensen, O.E.; Schneider, H.; Lewis, R.M.; Chernyavsky, I.L.; Brownbill, P. Human placental oxygenation in late gestation: Experimental and theoretical approaches. J. Physiol. 2018, 596, 5523–5534. [Google Scholar] [CrossRef]
- Bianchi, C.; Taricco, E.; Cardellicchio, M.; Mandò, C.; Massari, M.; Savasi, V.; Cetin, I. The role of obesity and gestational diabetes on placental size and fetal oxygenation. Placenta 2021, 103, 59–63. [Google Scholar] [CrossRef]
- Traub, O.; Berk, B.C. Laminar Shear Stress: Mechanisms by which ebdothelial cells transduce an atheroprotective force. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 677–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yinong, J.; Katsuhiko, K.; Kunio, H. Association Between Risk Factors for Atherosclerosis and Mechanical Forces in Carotid Artery. Stroke 2000, 31, 2319–2324. [Google Scholar] [CrossRef]
- Nieland, T.J.F.; Penman, M.; Dori, L.; Krieger, M.; Kirchhausen, T. Discovery of chemical inhibitors of the selective transfer of lipids mediated by the HDL receptor SR-BI. Proc. Natl. Acad. Sci. USA 2002, 99, 15422–15427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brundert, M.; Heeren, J.; Bahar-Bayansar, M.; Ewert, A.; Moore, K.J.; Rinninger, F. Selective uptake of HDL cholesteryl esters and cholesterol efflux from mouse peritoneal macrophages independent of SR-BI. J. Lipid Res. 2006, 47, 2408–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammad, S.M.; Barth, J.L.; Knaak, C.; Argraves, W.S. Megalin acts in concert with cubilin to mediate endocytosis of high density lipoproteins. J. Biol. Chem. 2000, 275, 12003–12008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, K.A.; Jauniaux, E.; Burton, G.J.; Cindrova-Davies, T. Expression and immunolocalisation of the endocytic receptors megalin and cubilin in the human yolk sac and placenta across gestation. Placenta 2013, 34, 1105–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storm, T.; Christensen, E.I.; Christensen, J.N.; Kjaergaard, T.; Uldbjerg, N.; Larsen, A.; Honoré, B.; Madsen, M. Megalin Is Predominantly Observed in Vesicular Structures in First and Third Trimester Cytotrophoblasts of the Human Placenta. J. Histochem. Cytochem. 2016, 64, 769–784. [Google Scholar] [CrossRef] [Green Version]
- Sreckovic, I.; Birner-Gruenberger, R.; Besenboeck, C.; Miljkovic, M.; Stojakovic, T.; Scharnagl, H.; Marsche, G.; Lang, U.; Kotur-Stevuljevic, J.; Jelic-Ivanovic, Z.; et al. Gestational diabetes mellitus modulates neonatal high-density lipoprotein composition and its functional heterogeneity. Biochim. Biophys. Acta (BBA)/Lipids Lipid Metab. 2014, 1841, 1619–1627. [Google Scholar] [CrossRef]
- Connelly, M.A.; Klein, S.M.; Azhar, S.; Abumrad, N.A.; Williams, D.L. Comparison of class B scavenger receptors, CD36 and scavenger receptor BI (SR-BI), shows that both receptors mediate high density lipoprotein-cholesteryl ester selective uptake but SR-BI exhibits a unique enhancement of cholesteryl ester uptake. J. Biol. Chem. 1999, 274, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Brundert, M.; Heeren, J.; Merkel, M.; Carambia, A.; Herkel, J.; Groitl, P.; Dobner, T.; Ramakrishnan, R.; Moore, K.J.; Rinninger, F. Scavenger receptor CD36 mediates uptake of high density lipoproteins in mice and by cultured cells. J. Lipid Res. 2011, 52, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Fox, H.; Sebire, N. Pathology of the Placenta; Elsevier: Amsterdam, The Netherlands, 2007; ISBN 9781416025924. [Google Scholar]
- Salafia, C.M.; Pezzullo, J.C.; Charles, A.K.; Ernst, L.M.; Maas, E.M.; Gross, B.; Pijnenborg, R. Morphometry of the Basal Plate Superficial Uteroplacental Vasculature in Normal Midtrimester and at Term. Pediatr. Dev. Pathol. 2005, 8, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Uittenbogaard, A.; Shaul, P.W.; Yuhanna, I.S.; Blair, A.; Smart, E.J. High density lipoprotein prevents oxidized low density lipoprotein-induced inhibition of endothelial nitric-oxide synthase localization and activation in caveolae. J. Biol. Chem. 2000, 275, 11278–11283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malek, A.M.; Ahlquist, R.; Gibbons, G.H.; Dzau, V.J.; Izumo, S. A cone-plate apparatus for the in vitro biochemical and molecular analysis of the effect of shear stress on adherent cells. Methods Cell Sci. 1995, 17, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Maksimovic, J.; Gordon, L.; Oshlack, A. SWAN: Subset-quantile Within Array Normalization for Illumina Infinium HumanMethylation450 BeadChips. Genome Biol. 2012, 13, R44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattler, W.; Mohr, D.; Stocker, R. Rapid isolation of lipoproteins and assessment of their peroxidation by high-performance liquid chromatography postcolumn chemiluminescence. Methods Enzymol. 1994, 233, 469–489. [Google Scholar]
- Artl, A.; Marsche, G.; Pussinen, P.; Knipping, G.; Sattler, W.; Malle, E. Impaired capacity of acute-phase high density lipoprotein particles to deliver cholesteryl ester to the human HUH-7 hepatoma cell line. Int. J. Biochem. Cell Biol. 2002, 34, 370–381. [Google Scholar] [CrossRef]
- Artl, A.; Marsche, G.; Lestavel, S.; Sattler, W.; Malle, E. Role of Serum Amyloid A During Metabolism of Acute-Phase HDL by Macrophages. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Benoist, F.; Lau, P.; McDonnell, M.; Doelle, H.; Milne, R.; McPherson, R. Cholesteryl ester transfer protein mediates selective uptake of high density lipoprotein cholesteryl esters by human adipose tissue. J. Biol. Chem. 1997, 272, 23572–23577. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strahlhofer-Augsten, M.; Schliefsteiner, C.; Cvitic, S.; George, M.; Lang-Olip, I.; Hirschmugl, B.; Marsche, G.; Lang, U.; Novakovic, B.; Saffery, R.; et al. The Distinct Role of the HDL Receptor SR-BI in Cholesterol Homeostasis of Human Placental Arterial and Venous Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 5364. https://doi.org/10.3390/ijms23105364
Strahlhofer-Augsten M, Schliefsteiner C, Cvitic S, George M, Lang-Olip I, Hirschmugl B, Marsche G, Lang U, Novakovic B, Saffery R, et al. The Distinct Role of the HDL Receptor SR-BI in Cholesterol Homeostasis of Human Placental Arterial and Venous Endothelial Cells. International Journal of Molecular Sciences. 2022; 23(10):5364. https://doi.org/10.3390/ijms23105364
Chicago/Turabian StyleStrahlhofer-Augsten, Manuela, Carolin Schliefsteiner, Silvija Cvitic, Meekha George, Ingrid Lang-Olip, Birgit Hirschmugl, Gunther Marsche, Uwe Lang, Boris Novakovic, Richard Saffery, and et al. 2022. "The Distinct Role of the HDL Receptor SR-BI in Cholesterol Homeostasis of Human Placental Arterial and Venous Endothelial Cells" International Journal of Molecular Sciences 23, no. 10: 5364. https://doi.org/10.3390/ijms23105364