
The Circadian Clock in the Retinal Pigment Epithelium Controls the Diurnal Rhythm of Phagocytic Activity

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
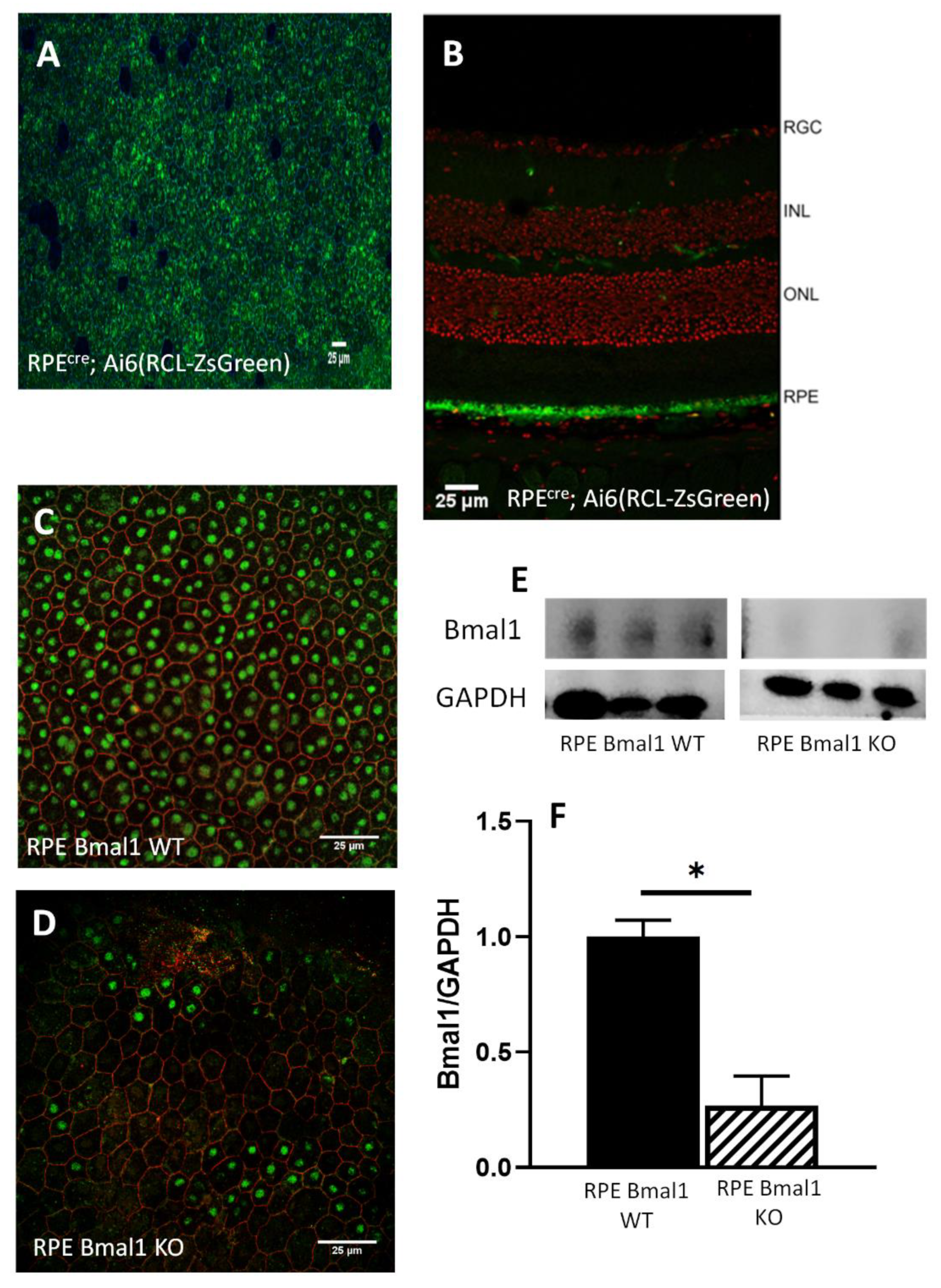
2.1. Removal of Bmal1 from RPE Cells
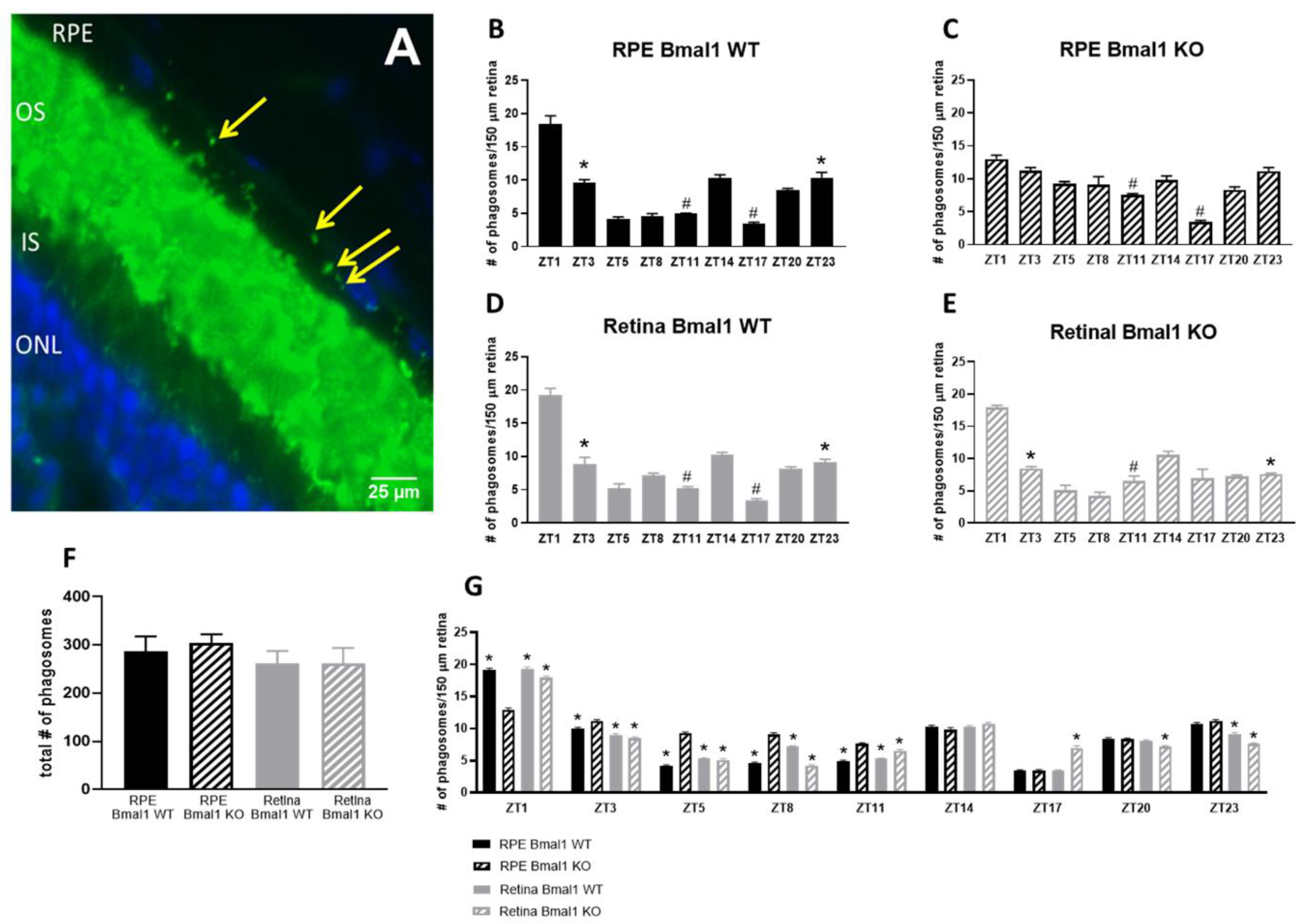
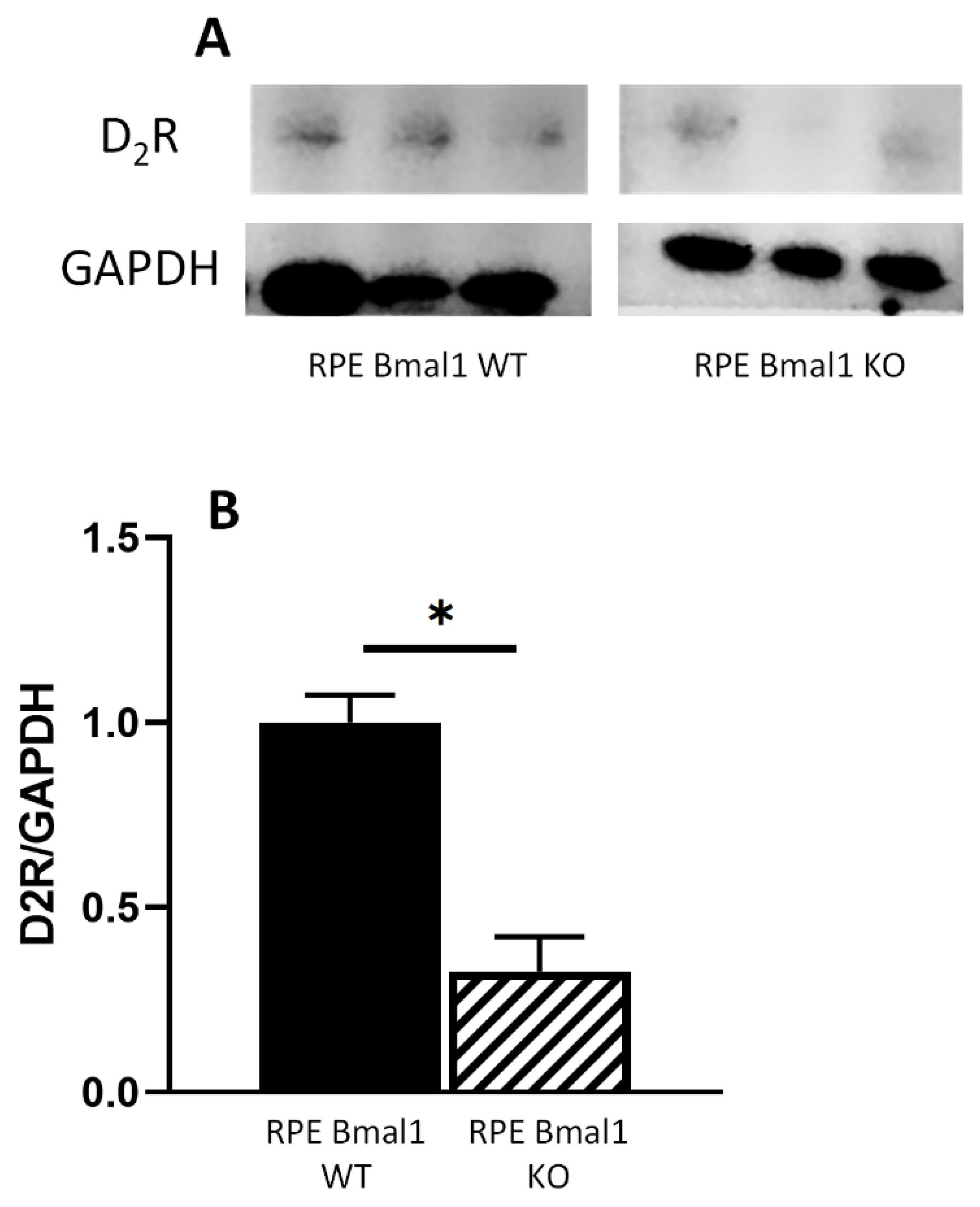
2.2. Loss of Bmal1 in the RPE Reduces the Daily Peak of Phagocytic Activity by the RPE
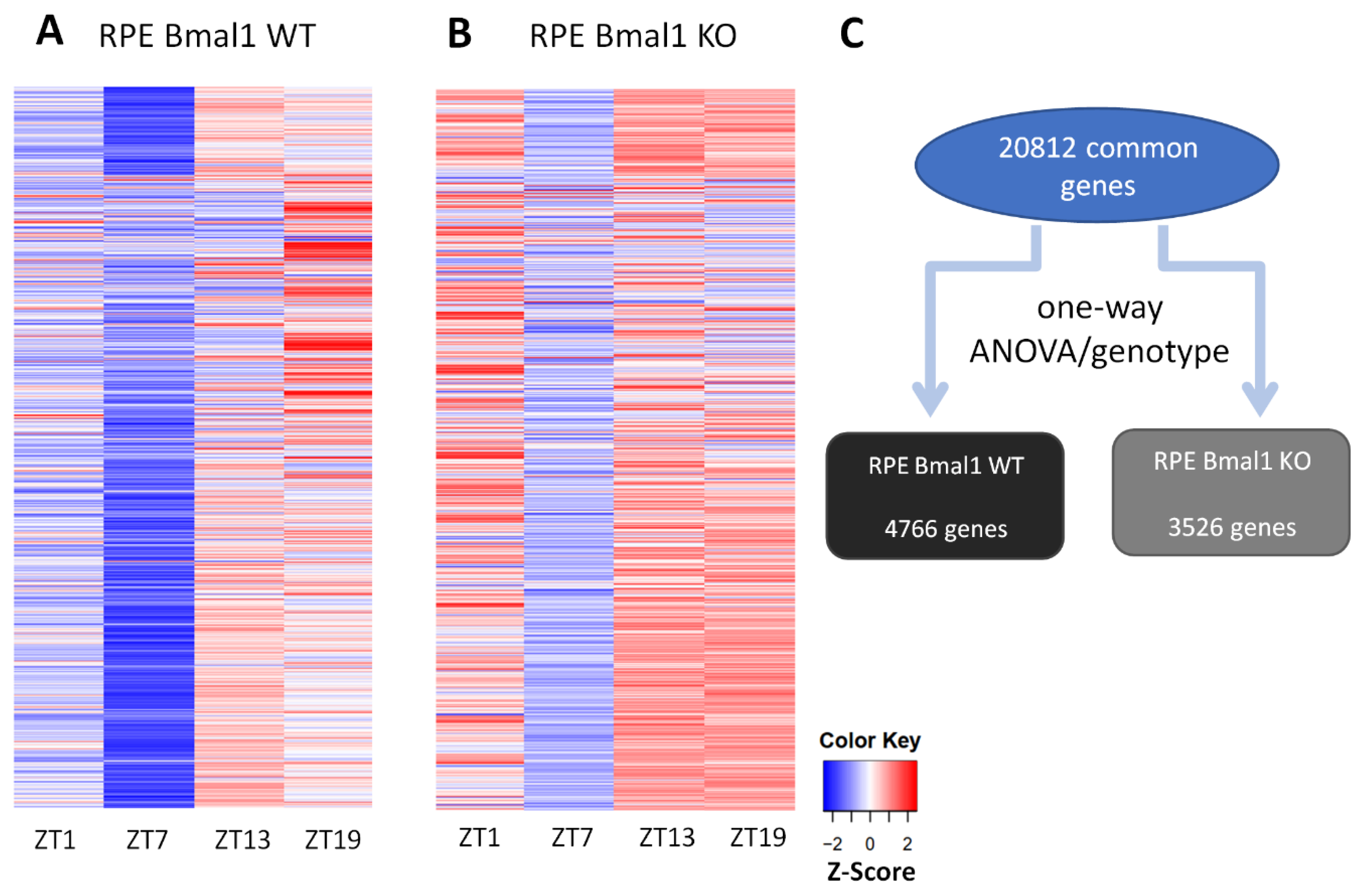
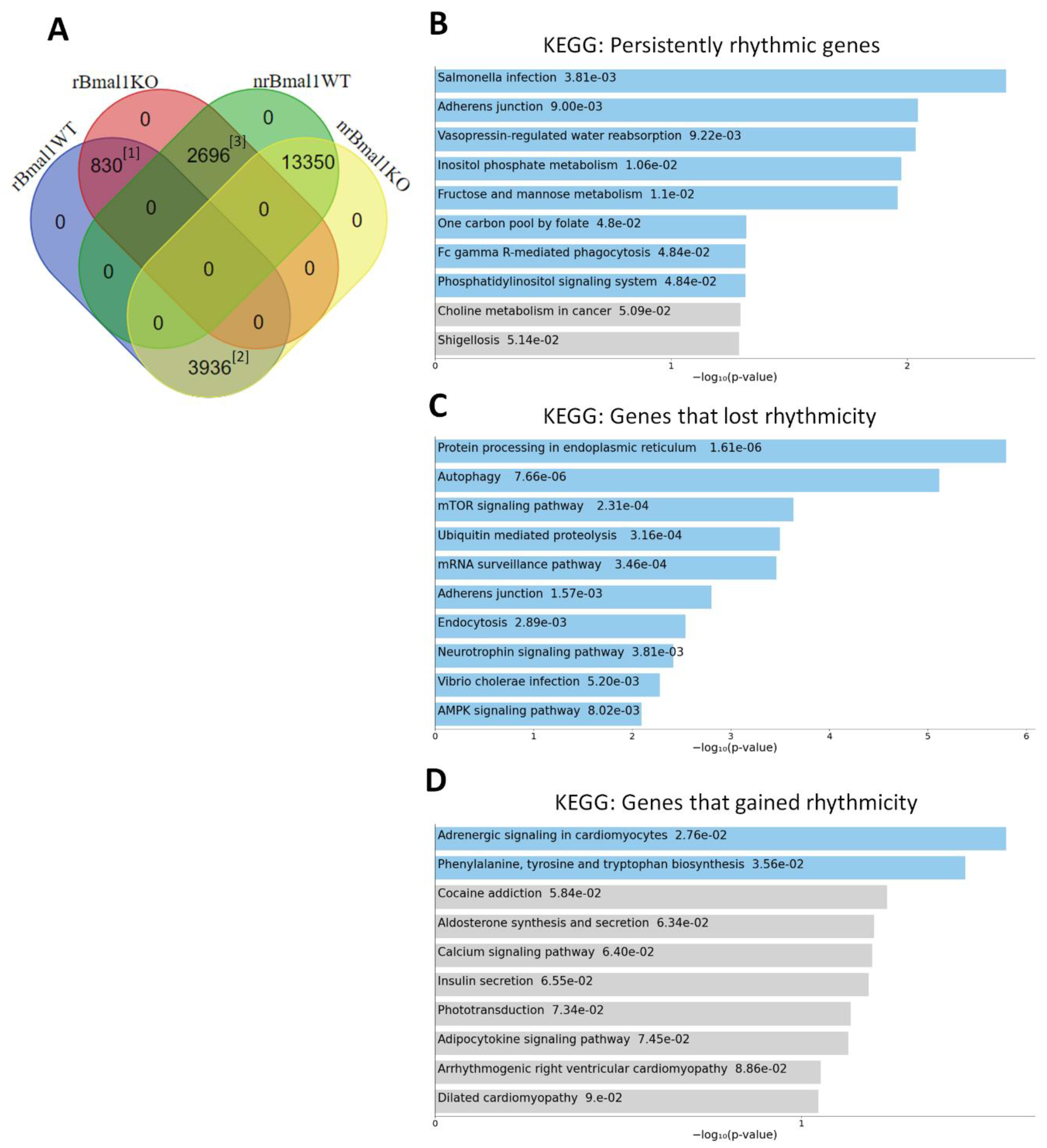
2.3. Loss of Bmal1 in RPE Alters the Daily Transcriptome
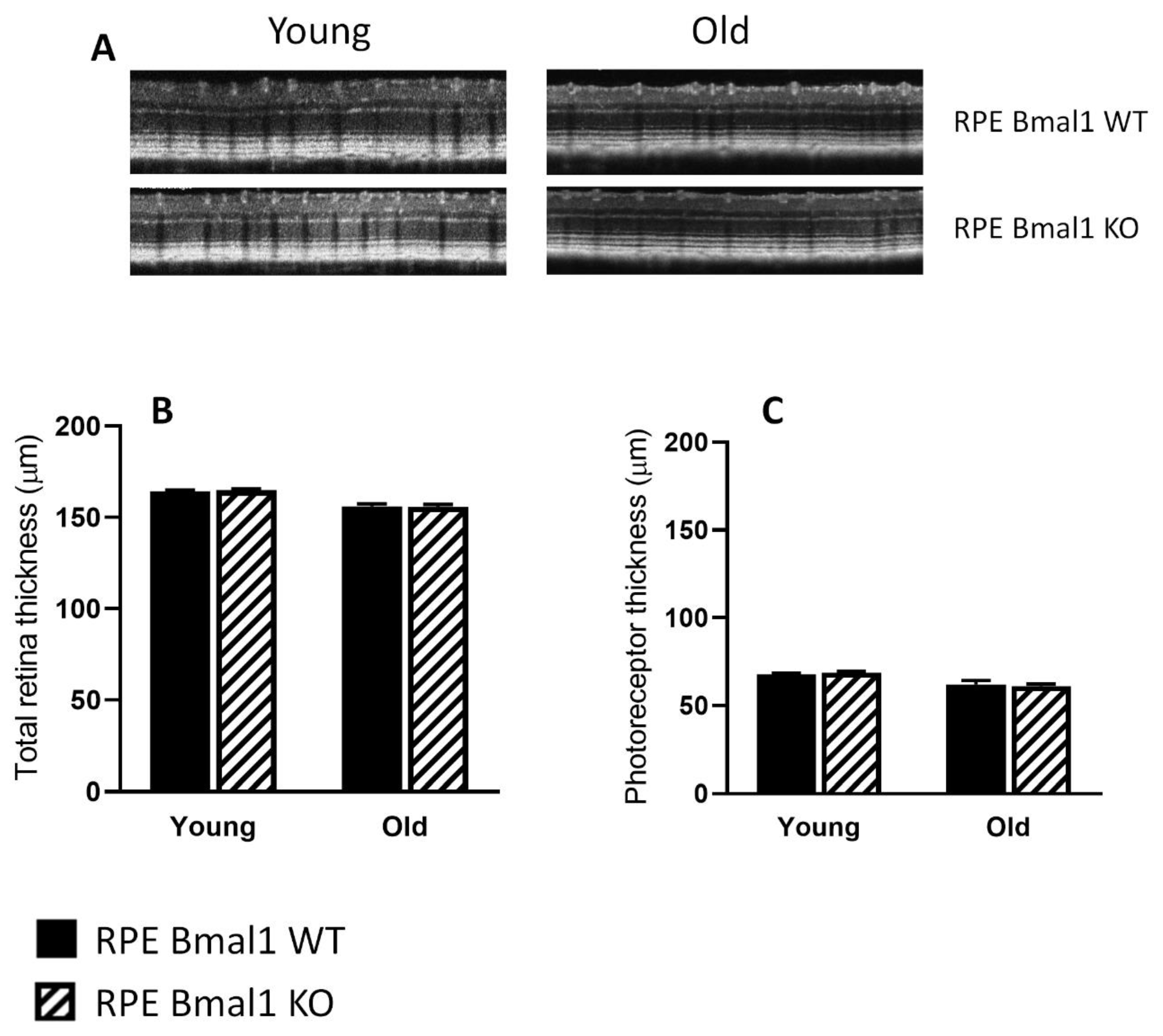
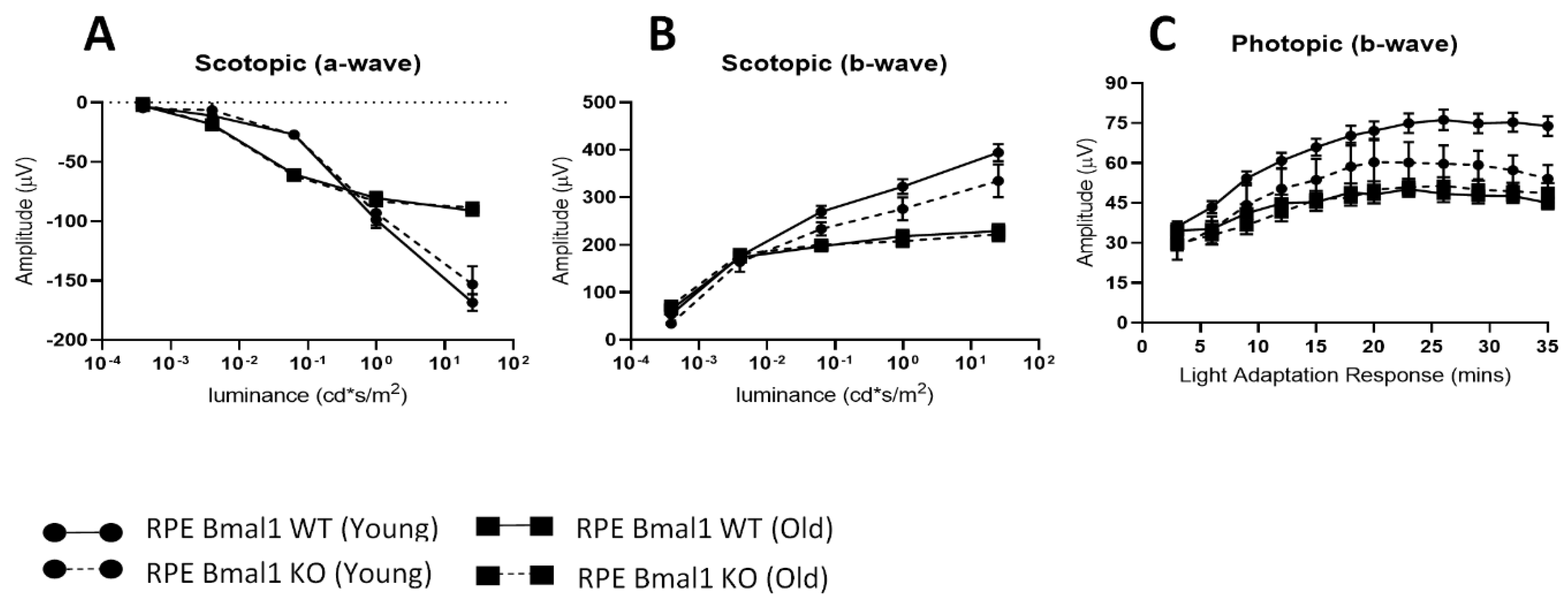
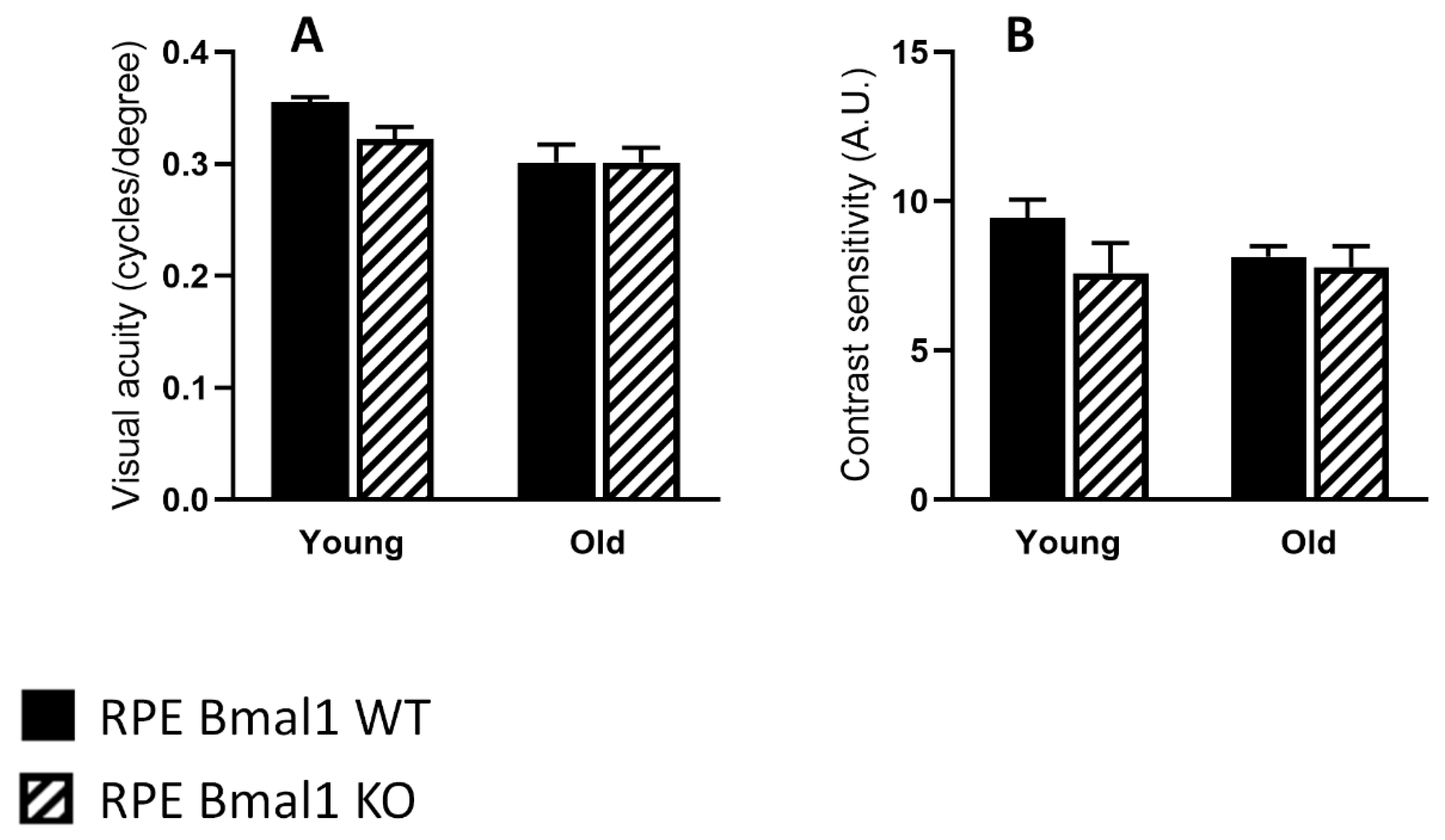
2.4. Removal of Bmal1 from the RPE Does Not Affect Retinal Structure during Aging
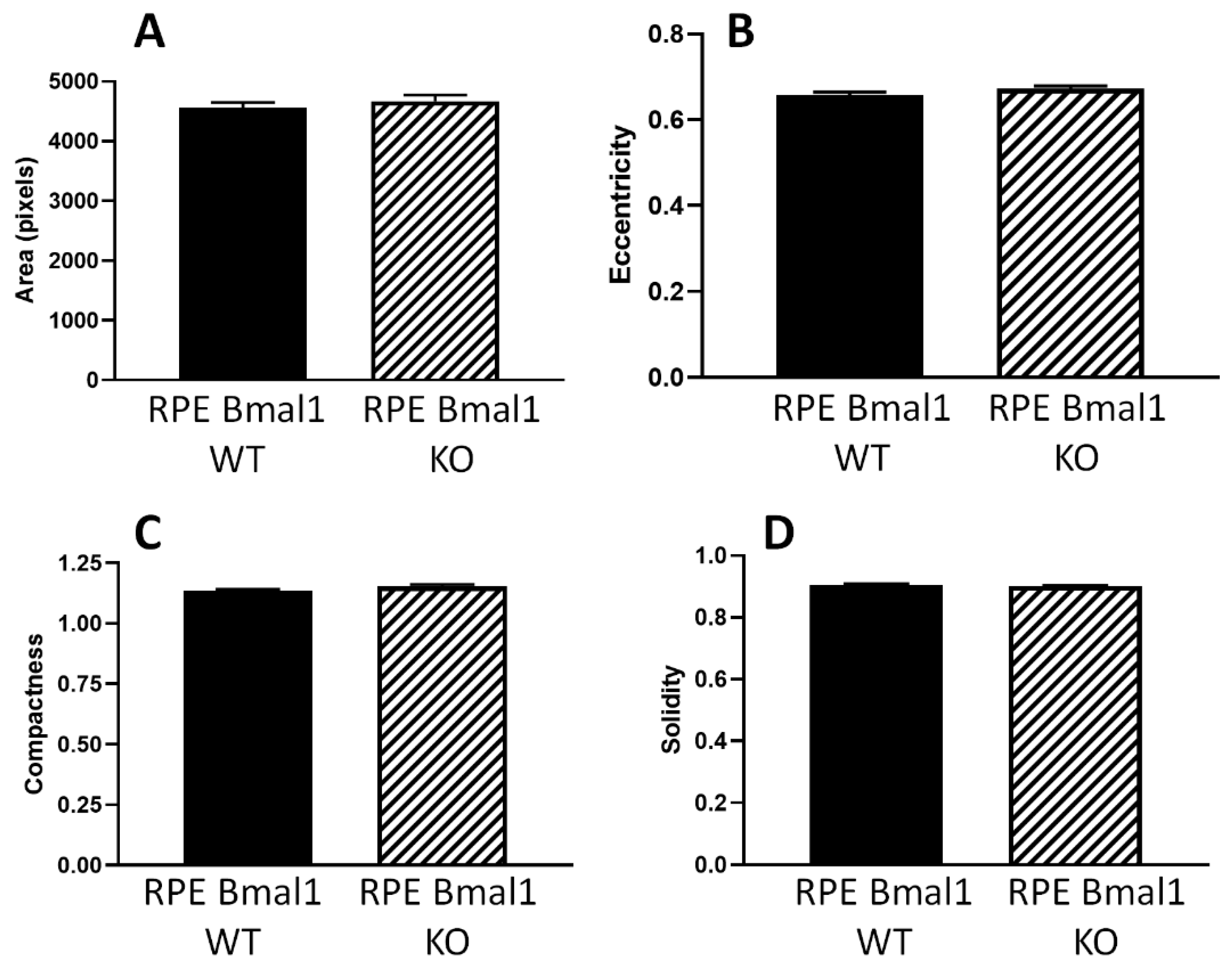
2.5. Removal of Bmal1 Does Not Produce Morphological Abnormalities in the RPE
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Phagosome Counting Assay
4.3. Western Blot
4.4. RPE RNA Library Preparation and Sequencing
4.5. Bioinformatics and Rhythmic Analysis
4.6. Spectral Domain-Optical Coherence Tomography (SD-OCT) and Fundus Imaging
4.7. Electroretinography (ERG)
4.8. Opto-Motor Response (OMR)
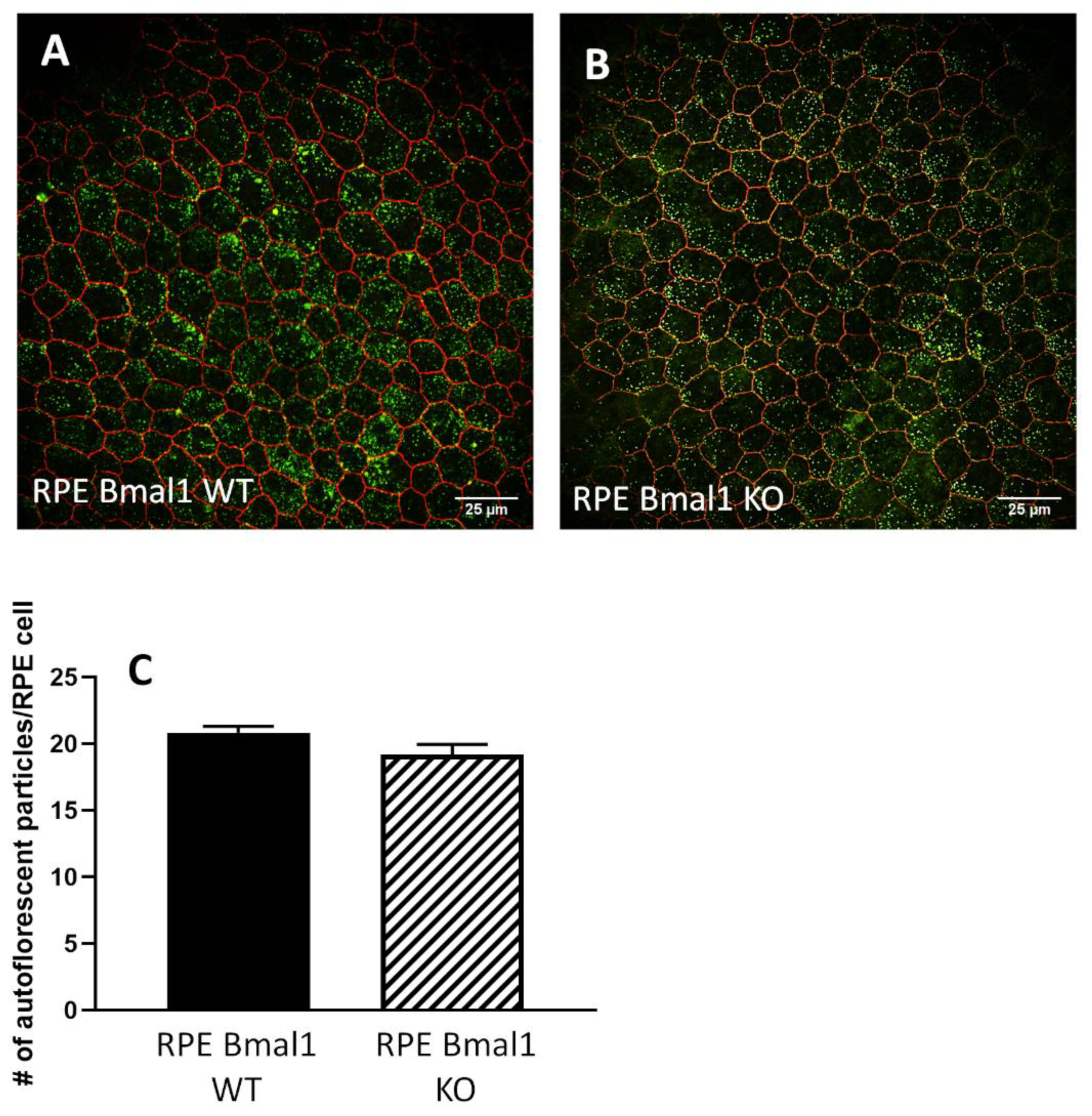
4.9. RPE Flat Mount Morphology and Autofluorescence Analysis
4.10. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunger, M.K.; Wilsbacher, L.D.; Moran, S.M.; Clendenin, C.; Radcliffe, L.A.; Hogenesch, J.B.; Simon, M.C.; Takahashi, J.S.; Bradfield, C.A. Mop3 Is an Essential Component of the Master Circadian Pacemaker in Mammals. Cell 2000, 103, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Kondratov, R.V.; Kondratova, A.A.; Gorbacheva, V.Y.; Vykhovanets, O.V.; Antoch, M.P. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006, 20, 1868–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musiek, E.; Lim, M.M.; Yang, G.; Bauer, A.Q.; Qi, L.; Lee, Y.; Roh, J.H.; Ortiz-Gonzalez, X.; Dearborn, J.; Culver, J.P.; et al. Circadian clock proteins regulate neuronal redox homeostasis and neurodegeneration. J. Clin. Investig. 2013, 123, 5389–5400. [Google Scholar] [CrossRef]
- DeVera, C.; Baba, K.; Tosini, G. Retinal Circadian Clocks are Major Players in the Modulation of Retinal Functions and Photoreceptor Viability. Yale J. Biol. Med. 2019, 92, 233–240. [Google Scholar]
- Baba, K.; Ribelayga, C.P.; Iuvone, P.M.; Tosini, G. The Retinal Circadian Clock and Photoreceptor Viability. Adv. Microbiol. Infect. Dis. Public Health 2018, 1074, 345–350. [Google Scholar] [CrossRef]
- Storch, K.-F.; Paz, C.; Signorovitch, J.; Raviola, E.; Pawlyk, B.; Li, T.; Weitz, C.J. Intrinsic Circadian Clock of the Mammalian Retina: Importance for Retinal Processing of Visual Information. Cell 2007, 130, 730–741. [Google Scholar] [CrossRef] [Green Version]
- Baba, K.; Piano, I.; Lyuboslavsky, P.; Chrenek, M.A.; Sellers, J.T.; Zhang, S.; Gargini, C.; He, L.; Tosini, G.; Iuvone, P.M. Removal of clock gene Bmal1 from the retina affects retinal development and accelerates cone photoreceptor degeneration during aging. Proc. Natl. Acad. Sci. USA 2018, 115, 13099–13104. [Google Scholar] [CrossRef] [Green Version]
- Sawant, O.B.; Horton, A.M.; Zucaro, O.F.; Chan, R.; Bonilha, V.L.; Samuels, I.S.; Rao, S. The Circadian Clock Gene Bmal1 Controls Thyroid Hormone-Mediated Spectral Identity and Cone Photoreceptor Function. Cell Rep. 2017, 21, 692–706. [Google Scholar] [CrossRef] [Green Version]
- Sawant, O.B.; Jidigam, V.K.; Fuller, R.D.; Zucaro, O.F.; Kpegba, C.; Yu, M.; Peachey, N.S.; Rao, S. The circadian clock gene Bmal1 is required to control the timing of retinal neurogenesis and lamination of Müller glia in the mouse retina. FASEB J. 2019, 33, 8745–8758. [Google Scholar] [CrossRef]
- Lakkaraju, A.; Umapathy, A.; Tan, L.X.; Daniele, L.; Philp, N.J.; Boesze-Battaglia, K.; Williams, D.S. The cell biology of the retinal pigment epithelium. Prog. Retin. Eye Res. 2020, 78, 100846. [Google Scholar] [CrossRef]
- Grace, M.S.; Chiba, A.; Menaker, M. Circadian control of photoreceptor outer segment membrane turnover in mice genetically incapable of melatonin synthesis. Vis. Neurosci. 1999, 16, 909–918. [Google Scholar] [CrossRef]
- LaVail, M.M. Rod Outer Segment Disk Shedding in Rat Retina: Relationship to Cyclic Lighting. Science 1976, 194, 1071–1074. [Google Scholar] [CrossRef]
- Lo, W.-K.; Bernstein, M.H. Daily patterns of the retinal pigment epithelium. Microperoxisomes and phagosomes. Exp. Eye Res. 1981, 32, 1–10. [Google Scholar] [CrossRef]
- Besharse, J.C.; Hollyfield, J.G.; Rayborn, M.E. Turnover of rod photoreceptor outer segments. II. Membrane addition and loss in relationship to light. J. Cell Biol. 1977, 75, 507–527. [Google Scholar] [CrossRef]
- Terman, J.S.; Reme, C.E.; Terman, M. Rod outer segment disk shedding in rats with lesions of the suprachiasmatic nucleus. Brain Res. 1993, 605, 256–264. [Google Scholar] [CrossRef]
- Teirstein, P.S.; Goldman, A.I.; O’Brien, P.J. Evidence for both local and central regulation of rat rod outer segment disc shedding. Investig. Ophthalmol. Vis. Sci. 1980, 19, 1268–1273. [Google Scholar]
- Baba, K.; Sengupta, A.; Tosini, M.; Contreras-Alcantara, S.; Tosini, G. Circadian regulation of the PERIOD 2::LUCIFERASE bioluminescence rhythm in the mouse retinal pigment epithelium-choroid. Mol. Vis. 2010, 16, 2605–2611. [Google Scholar]
- Baba, K.; Debruyne, J.P.; Tosini, G. Dopamine 2 Receptor Activation Entrains Circadian Clocks in Mouse Retinal Pigment Epithelium. Sci. Rep. 2017, 7, 5103. [Google Scholar] [CrossRef]
- Nandrot, E.F.; Kim, Y.; Brodie, S.; Huang, X.; Sheppard, D.; Finnemann, S.C. Loss of Synchronized Retinal Phagocytosis and Age-related Blindness in Mice Lacking αvβ5 Integrin. J. Exp. Med. 2004, 200, 1539–1545. [Google Scholar] [CrossRef]
- Goyal, V.; DeVera, C.; Laurent, V.; Sellers, J.; Chrenek, M.A.; Hicks, D.; Baba, K.; Iuvone, P.M.; Tosini, G. Dopamine 2 Receptor Signaling Controls the Daily Burst in Phagocytic Activity in the Mouse Retinal Pigment Epithelium. Investig. Opthalmol. Vis. Sci. 2020, 61, 10. [Google Scholar] [CrossRef]
- Nandrot, E.F.; Finnemann, S.C. Lack of αvβ5 Integrin Receptor or Its Ligand MFG-E8: Distinct Effects on Retinal Function. Ophthalmic Res. 2008, 40, 120–123. [Google Scholar] [CrossRef] [Green Version]
- DeVera, C.; Tosini, G. Circadian analysis of the mouse retinal pigment epithelium transcriptome. Exp. Eye Res. 2020, 193, 107988. [Google Scholar] [CrossRef]
- Hughes, M.E.; Abruzzi, K.C.; Allada, R.; Anafi, R.; Arpat, B.; Asher, G.; Baldi, P.; de Bekker, C.; Bell-Pedersen, D.; Blau, J.; et al. Guidelines for Genome-Scale Analysis of Biological Rhythms. J. Biol. Rhythm. 2017, 32, 380–393. [Google Scholar] [CrossRef] [Green Version]
- Louer, E.M.; Günzel, D.; Rosenthal, R.; Carmone, C.; Yi, G.; Stunnenberg, H.G.; Hollander, A.I.D.; Deen, P.M. Differential day-night expression of tight junction components in murine retinal pigment epithelium. Exp. Eye Res. 2020, 193, 107985. [Google Scholar] [CrossRef]
- Besharse, J.C.; Hollyfield, J.G. Turnover of mouse photoreceptor outer segments in constant light and darkness. Investig. Ophthalmol. Vis. Sci. 1979, 18, 1019–1024. [Google Scholar]
- Laurent, V.; Sengupta, A.; Sánchez-Bretaño, A.; Hicks, D.; Tosini, G. Melatonin signaling affects the timing in the daily rhythm of phagocytic activity by the retinal pigment epithelium. Exp. Eye Res. 2017, 165, 90–95. [Google Scholar] [CrossRef]
- Le, Y.-Z.; Zheng, W.; Rao, P.-C.; Zheng, L.; Anderson, R.E.; Esumi, N.; Zack, N.J.; Zhu, M. Inducible Expression of Cre Recombinase in the Retinal Pigmented Epithelium. Investig. Opthalmol. Vis. Sci. 2008, 49, 1248–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, T.R.; Kundinger, S.R.; Link, B.A.; Insinna, C.; Besharse, J.C. Kif17 phosphorylation regulates photoreceptor outer segment turnover. BMC Cell Biol. 2018, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Bulloj, A.; Duan, W.; Finnemann, S.C. PI 3-kinase independent role for AKT in F-actin regulation during outer segment phagocytosis by RPE cells. Exp. Eye Res. 2013, 113, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafi, D.; Kevany, B.M.; Genoud, C.; Bai, X.; Palczewski, K. Photoreceptor phagocytosis is mediated by phosphoinositide signaling. FASEB J. 2013, 27, 4585–4595. [Google Scholar] [CrossRef] [Green Version]
- Louer, E.M.M.; Yi, G.; Carmone, C.; Robben, J.; Stunnenberg, H.G.; Hollander, A.I.D.; Deen, P.M.T. Genes Involved in Energy Metabolism Are Differentially Expressed During the Day–Night Cycle in Murine Retinal Pigment Epithelium. Investig. Opthalmol. Vis. Sci. 2020, 61, 49. [Google Scholar] [CrossRef]
- Milićević, N.; Hakkari, O.A.; Bagchi, U.; Sandu, C.; Jongejan, A.; Moerland, P.D.; Brink, J.B.; Hicks, D.; Bergen, A.A.; Felder-Schmittbuhl, M. Core circadian clock genes Per1 and Per2 regulate the rhythm in photoreceptor outer segment phagocytosis. FASEB J. 2021, 35, e21722. [Google Scholar] [CrossRef]
- Weger, B.D.; Gobet, C.; David, F.P.A.; Atger, F.; Martin, E.; Phillips, N.E.; Charpagne, A.; Weger, M.; Naef, F.; Gachon, F. Systematic analysis of differential rhythmic liver gene expression mediated by the circadian clock and feeding rhythms. Proc. Natl. Acad. Sci. USA 2021, 118, e2015803118. [Google Scholar] [CrossRef]
- Yang, G.; Chen, L.; Grant, G.R.; Paschos, G.; Song, W.-L.; Musiek, E.S.; Lee, V.; McLoughlin, S.C.; Grosser, T.; Cotsarelis, G.; et al. Timing of expression of the core clock gene Bmal1 influences its effects on aging and survival. Sci. Transl. Med. 2016, 8, 324ra16. [Google Scholar] [CrossRef] [Green Version]
- Ray, S.; Valekunja, U.K.; Stangherlin, A.; Howell, S.A.; Snijders, A.P.; Damodaran, G.; Reddy, A.B. Circadian rhythms in the absence of the clock gene Bmal1. Science 2020, 367, 800–806. [Google Scholar] [CrossRef]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and Metabolic Syndrome in Circadian Clock Mutant Mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [Green Version]
- Bennis, A.; Gorgels, T.; Brink, J.T.; Van Der Spek, P.; Bossers, K.; Heine, V.; Bergen, A. Comparison of Mouse and Human Retinal Pigment Epithelium Gene Expression Profiles: Potential Implications for Age-Related Macular Degeneration. PLoS ONE 2015, 10, e0141597. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Goyal, V.; DeVera, C.; Baba, K.; Sellers, J.; Chrenek, M.A.; Iuvone, P.M.; Tosini, G. Photoreceptor Degeneration in Homozygous Male Per2luc Mice During Aging. J. Biol. Rhythm. 2020, 36, 137–145. [Google Scholar] [CrossRef]
- Boatright, J.H.; Dalal, N.; Chrenek, M.A.; Gardner, C.; Ziesel, A.; Jiang, Y.; Grossniklaus, H.E.; Nickerson, J.M. Methodologies for analysis of patterning in the mouse RPE sheet. Mol. Vis. 2015, 21, 40–60. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeVera, C.; Dixon, J.; Chrenek, M.A.; Baba, K.; Le, Y.Z.; Iuvone, P.M.; Tosini, G. The Circadian Clock in the Retinal Pigment Epithelium Controls the Diurnal Rhythm of Phagocytic Activity. Int. J. Mol. Sci. 2022, 23, 5302. https://doi.org/10.3390/ijms23105302
DeVera C, Dixon J, Chrenek MA, Baba K, Le YZ, Iuvone PM, Tosini G. The Circadian Clock in the Retinal Pigment Epithelium Controls the Diurnal Rhythm of Phagocytic Activity. International Journal of Molecular Sciences. 2022; 23(10):5302. https://doi.org/10.3390/ijms23105302
Chicago/Turabian StyleDeVera, Christopher, Jendayi Dixon, Micah A. Chrenek, Kenkichi Baba, Yun Z. Le, P. Michael Iuvone, and Gianluca Tosini. 2022. "The Circadian Clock in the Retinal Pigment Epithelium Controls the Diurnal Rhythm of Phagocytic Activity" International Journal of Molecular Sciences 23, no. 10: 5302. https://doi.org/10.3390/ijms23105302