
Transcriptional Contribution of Transposable Elements in Relation to Salinity Conditions in Teleosts and Silencing Mechanisms Involved

 , ,
, ,  , ,
, ,  , , and
, , and 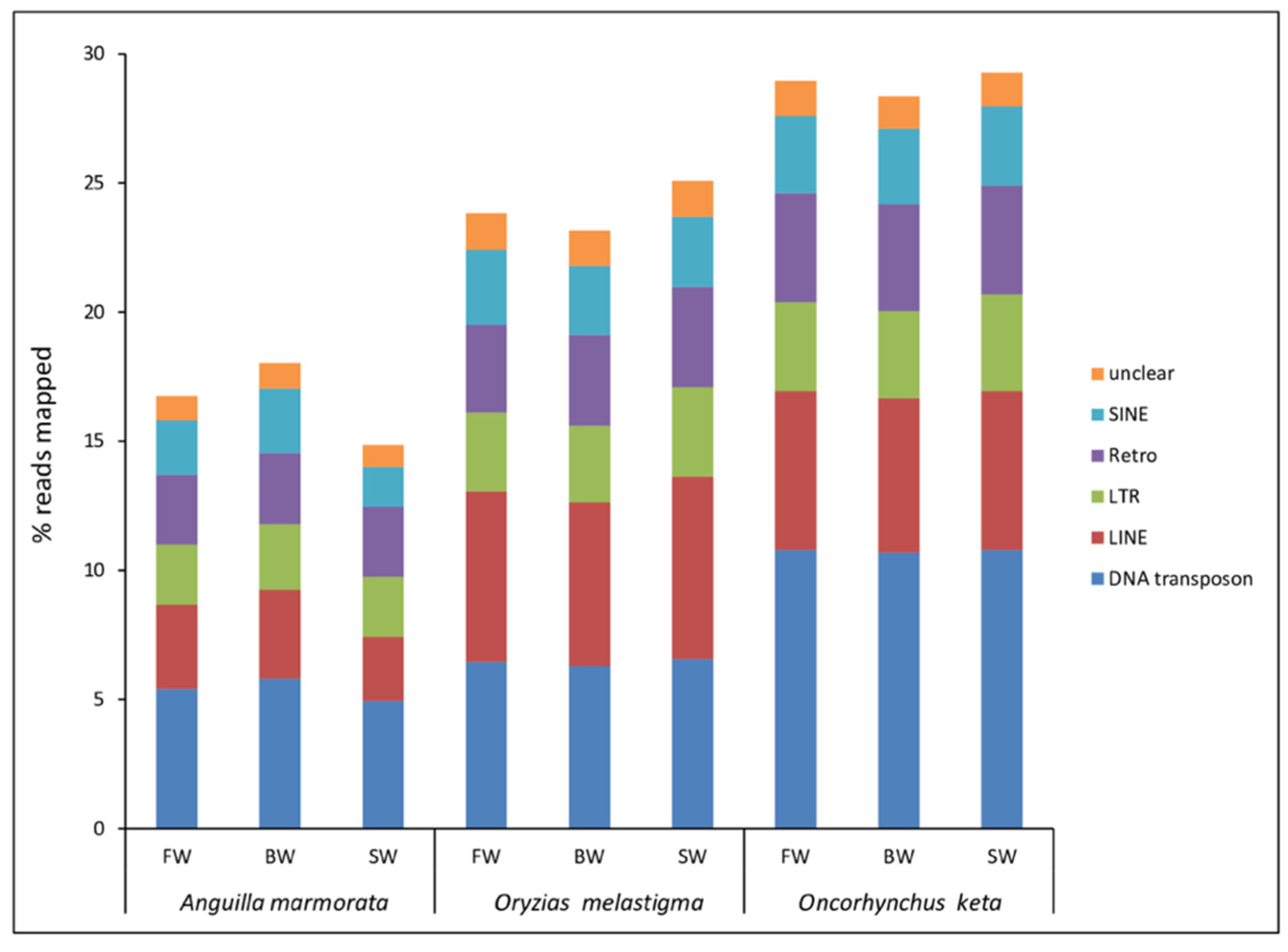{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
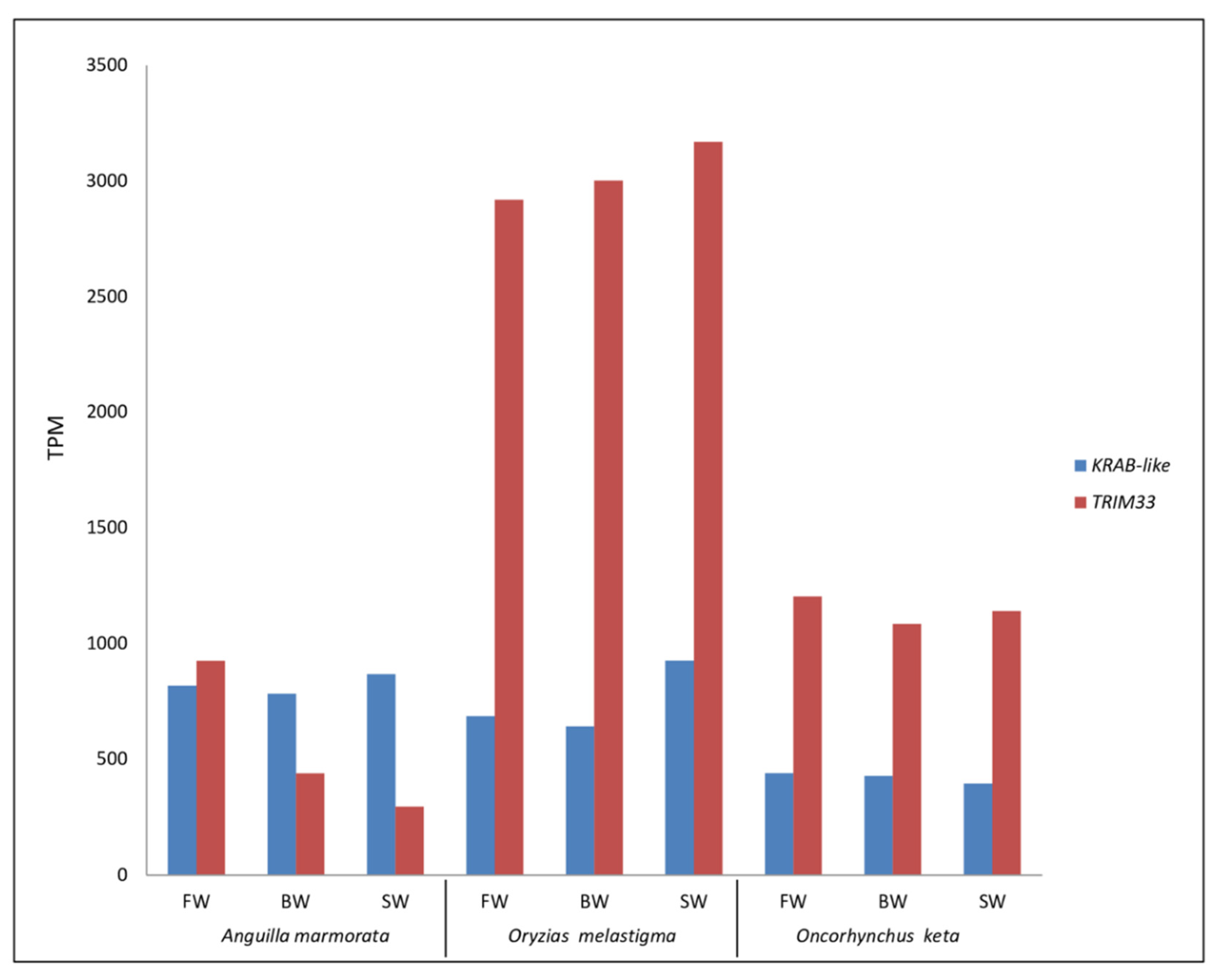
2.1. Transcriptional Contribution of Transposable Elements in Gill Transcriptomes of A. marmorata, O. melastigma, and O. keta
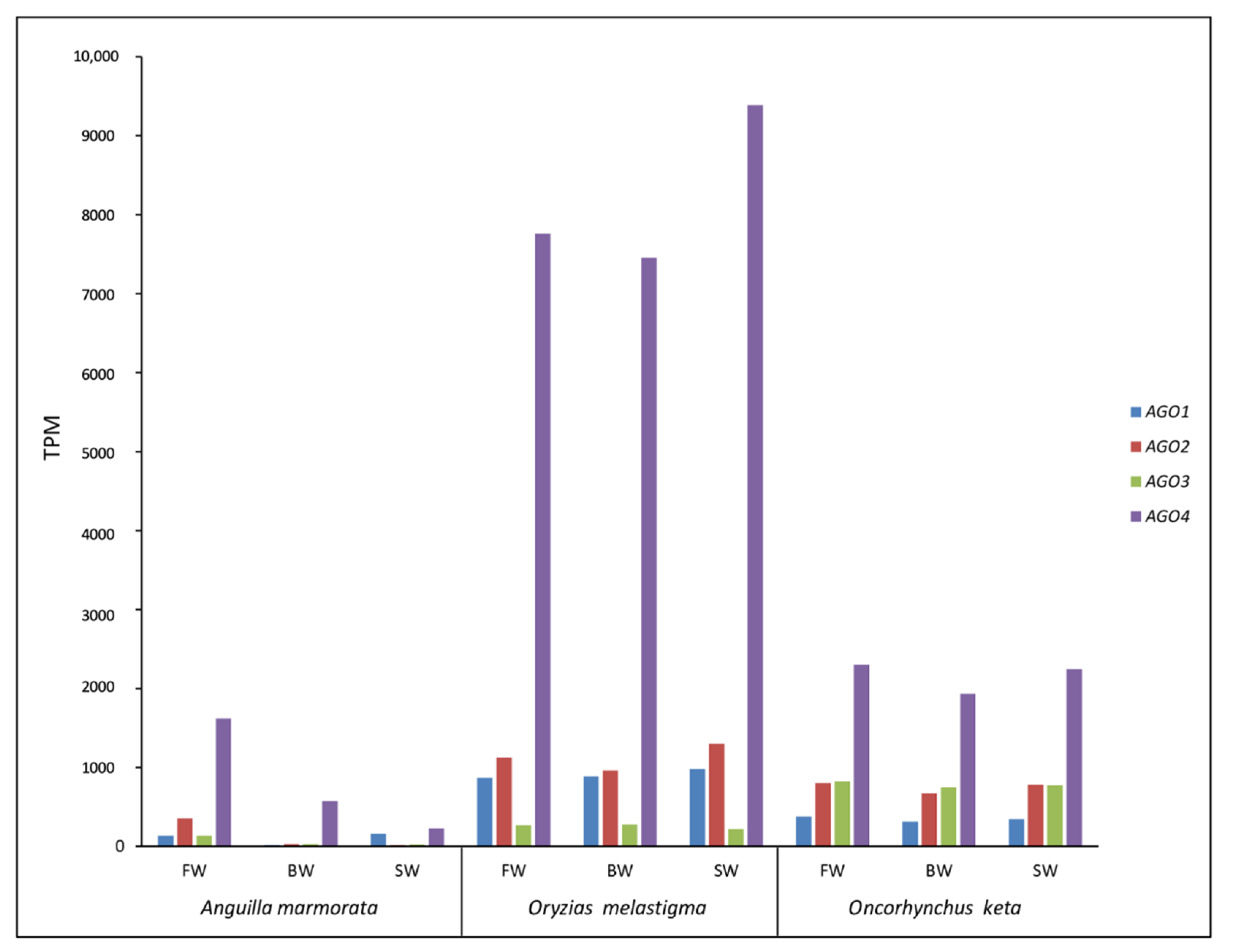
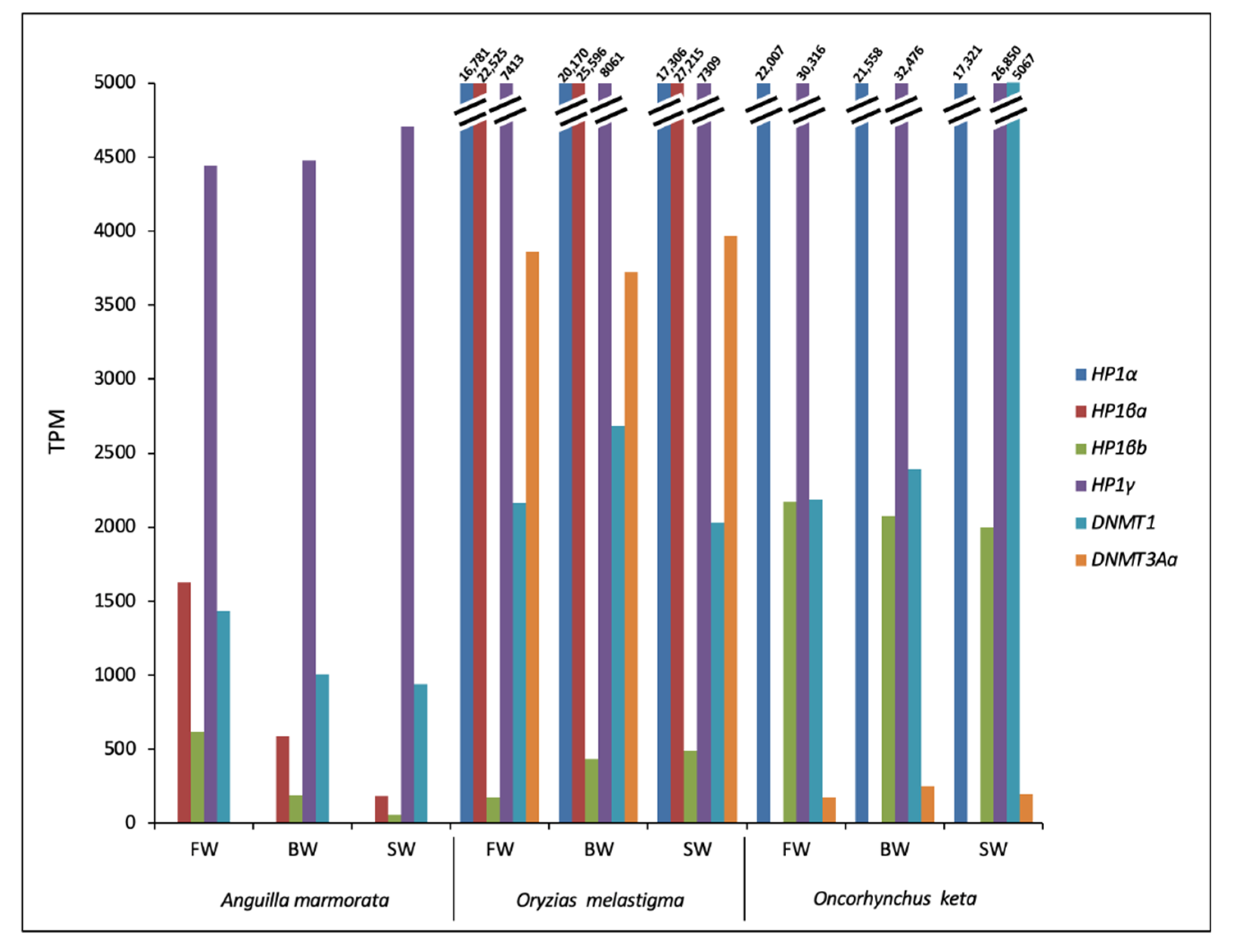
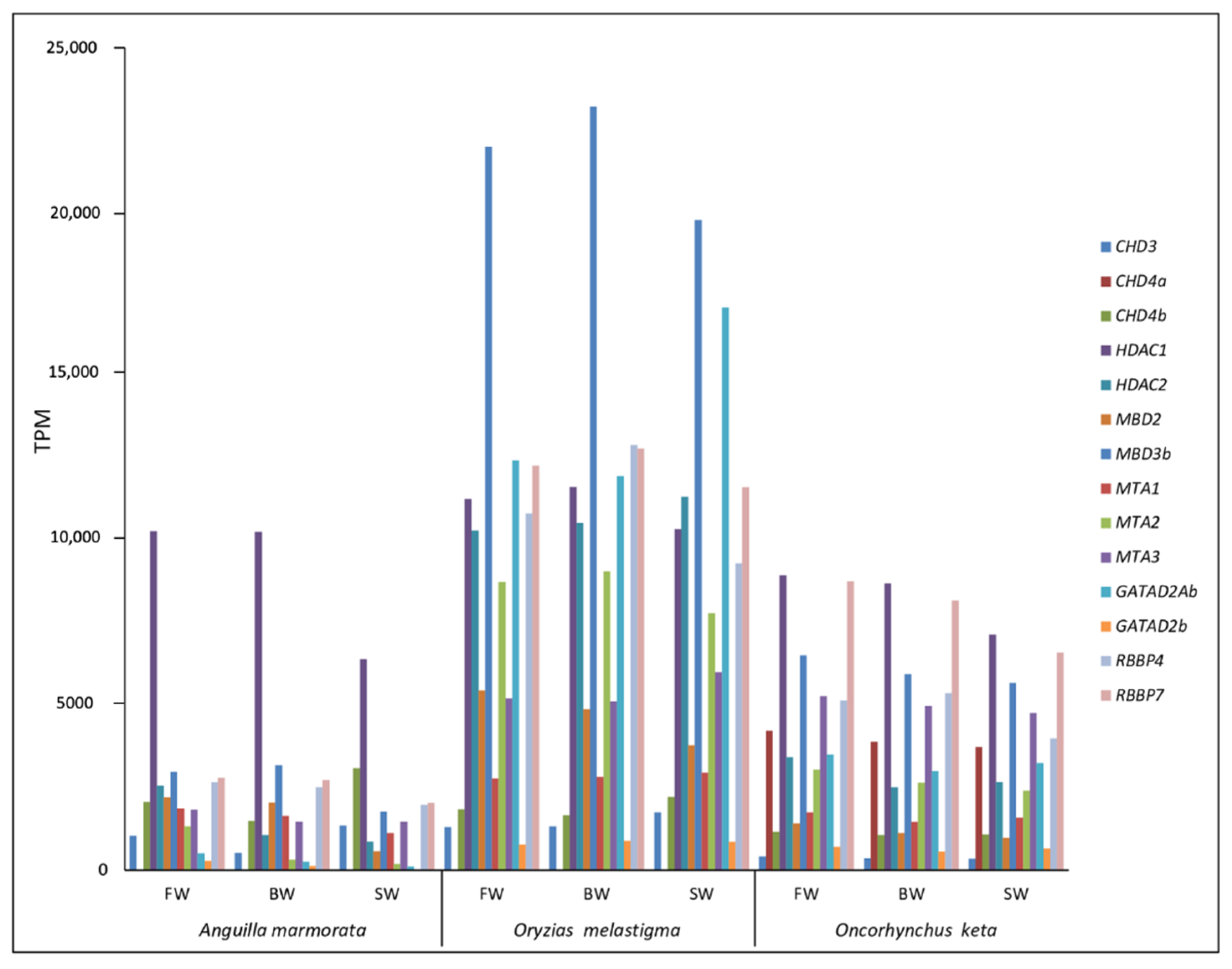
2.2. Identification and Transcriptional Activity of Genes Involved in TE Silencing Mechanisms
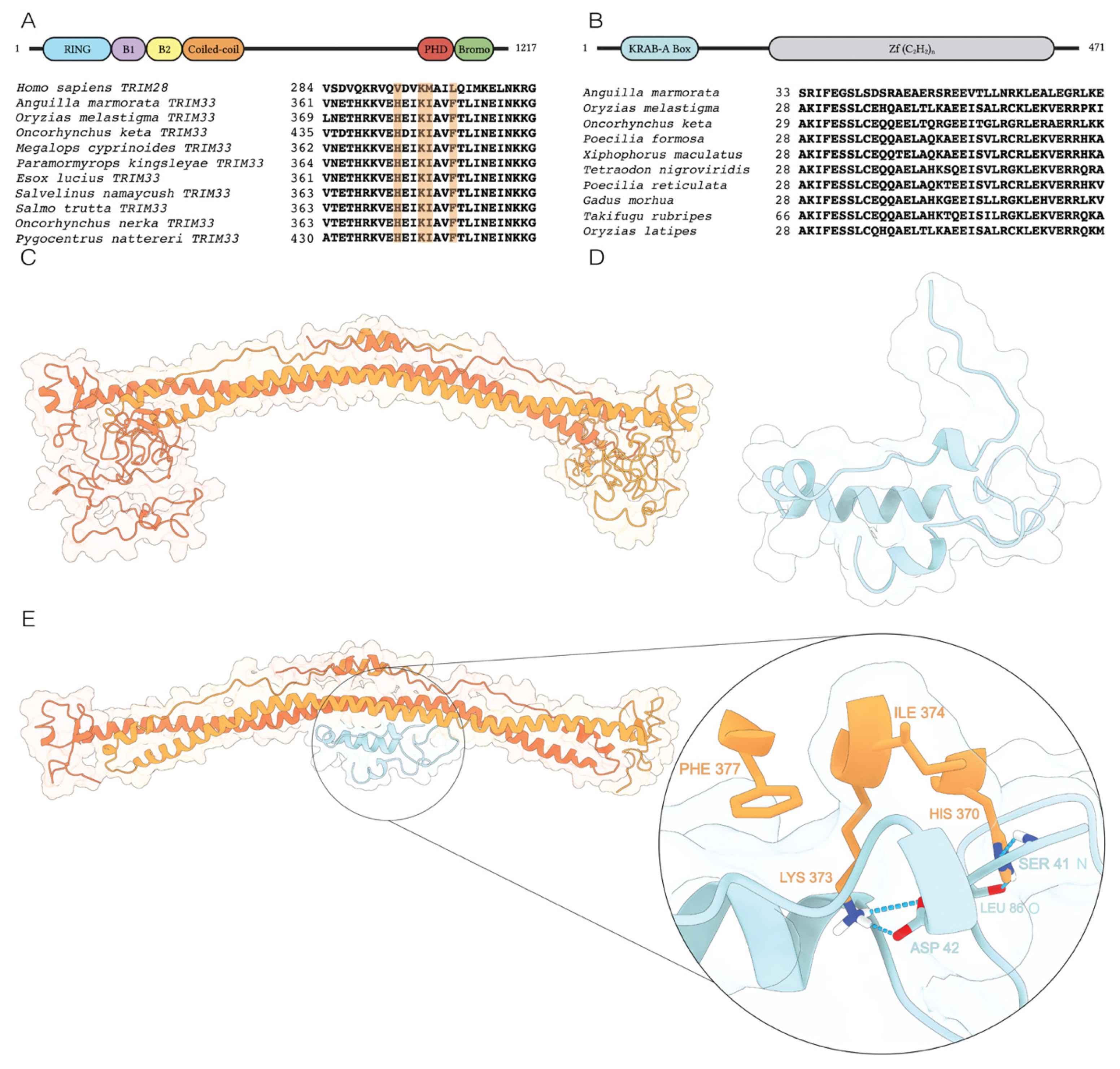
2.3. Structural Prediction of the TRIM33/KRAB-like Complex
3. Discussion
4. Materials and Methods
4.1. Estimation of TE Activity
4.2. Identification and Expression Analysis of Genes of Interest
4.3. Molecular Modelling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fujino, K.; Hashida, S.N.; Ogawa, T.; Natsume, T.; Uchiyama, T.; Mikami, T.; Kishima, Y. Temperature controls nuclear import of Tam3 transposase in Antirrhinum. Plant J. 2011, 65, 146–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casacuberta, E.; González, J. The impact of transposable elements in environmental adaptation. Mol. Ecol. 2013, 6, 1503–1517. [Google Scholar] [CrossRef] [PubMed]
- Makarevitch, I.; Waters, A.J.; West, P.T.; Stitzer, M.; Hirsch, C.N.; Ross-Ibarra, J.; Springer, N.M. Transposable elements contribute to activation of maize genes in response to abiotic stress. PLoS Genet. 2015, 11, e1004915. [Google Scholar]
- Schmidt, A.L.; Anderson, L.M. Repetitive DNA elements as mediators of genomic change in response to environmental cues. Biol. Rev. Camb. Philos. Soc. 2006, 81, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Fablet, M.; Vieira, C. Evolvability, epigenetics and transposable elements. Biomol. Concepts 2011, 2, 331–341. [Google Scholar] [CrossRef]
- Canapa, A.; Biscotti, M.A.; Barucca, M.; Carducci, F.; Carotti, E.; Olmo, E. Shedding light upon the complex net of genome size, genome composition and environment in chordates. Eur. Zool. J. 2020, 87, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.G.; Tan, C.T.; Yu, G.T.; Zhong, S.; Xu, S.S.; Yan, L. A novel retrotransposon inserted in the dominant Vrn-B1 allele confers spring growth habit in tetraploid wheat (Triticum turgidum L.). G3 2011, 1, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Carducci, F.; Barucca, M.; Canapa, A.; Carotti, E.; Biscotti, M.A. Mobile elements in ray-finned fish genomes. Life 2020, 10, 221. [Google Scholar] [CrossRef]
- Carducci, F.; Biscotti, M.A.; Forconi, M.; Barucca, M.; Canapa, A. An intriguing relationship between teleost Rex3 retroelement and environmental temperature. Biol. Lett. 2019, 15, 20190279. [Google Scholar] [CrossRef] [Green Version]
- Carotti, E.; Carducci, F.; Canapa, A.; Barucca, M.; Greco, S.; Gerdol, M.; Biscotti, M.A. Transposable elements and teleost migratory behaviour. Int. J. Mol. Sci. 2021, 22, 602. [Google Scholar] [CrossRef]
- Pappalardo, A.M.; Ferrito, V.; Biscotti, M.A.; Canapa, A.; Capriglione, T. Transposable elements and stress in vertebrates: An overview. Int. J. Mol. Sci. 2021, 22, 1970. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Yang, J.; Tang, W.; Zhang, X.; Royster, C.M.; Zhang, M. SINE Retrotransposon variation drives Ecotypic disparity in natural populations of Coilia nasus. Mob. DNA 2020, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Steendam, C.; Verhelst, P.; Van Wassenbergh, S.; De Meyer, J. Burrowing behaviour of the European eel (Anguilla anguilla): Effects of life stage. J. Fish Biol. 2020, 97, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Lee, H.J.; Kim, Y.K. Comparative transcriptome profiling of selected osmotic regulatory proteins in the gill during seawater acclimation of chum salmon (Oncorhynchus keta) fry. Sci. Rep. 2020, 10, 1987. [Google Scholar] [CrossRef] [PubMed]
- Beechie, T.J.; Nicol, C.; Fogel, C.; Jorgensen, J.; Timpane-Padgham, B. Chum Salmon Life-Cycle Model Description and Results for the Chehalis River Basin; NOAA Contract Report NMFS-NWFSC-CR-2021-02; U.S. Department of Commerce: Washington, DC, USA, 2021.
- Cao, Q.; Wang, H.; Fan, C.; Sun, Y.; Li, J.; Cheng, J.; Chu, P.; Yin, S. Environmental salinity influences the branchial expression of TCR pathway related genes based on transcriptome of a catadromous fish. Comp. Biochem. Physiol. D 2021, 38, 100815. [Google Scholar] [CrossRef]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef]
- Boudinot, P.; van der Aa, L.M.; Jouneau, L.; Du Pasquier, L.; Pontarotti, P.; Briolat, V.; Benmansour, A.; Levraud, J.P. Origin and evolution of TRIM proteins: New insights from the complete TRIM repertoire of zebrafish and pufferfish. PLoS ONE 2011, 6, 7. [Google Scholar] [CrossRef]
- Stoll, G.A.; Oda, S.I.; Chong, Z.S.; Yu, M.; McLaughlin, S.H.; Modis, Y. Structure of KAP1 tripartite motif identifies molecular interfaces required for retroelement silencing. Proc. Natl. Acad. Sci. USA 2019, 116, 15042–15051. [Google Scholar] [CrossRef] [Green Version]
- Di Marino, D.; Chillemi, G.; De Rubeis, S.; Tramontano, A.; Achsel, T.; Bagni, C. MD and Docking Studies Reveal That the Functional Switch of CYFIP1 is mediated by a Butterfly-like Motion. J. Chem. Theory Comput. 2015, 11, 3401–3410. [Google Scholar] [CrossRef]
- Trucchi, E.; Gratton, P.; Mafessoni, F.; Motta, S.; Cicconardi, F.; Mancia, F.; Bertorelle, G.; D’Annessa, I.; Di Marino, D. Population dynamics and structural effects at short and long range support the hypothesis of the selective advantage of the G614 SARS-CoV-2 Spike Variant. Mol. Biol. Evol. 2021, 38, 1966–1979. [Google Scholar] [CrossRef]
- Schmidt, D.; Schwalie, P.C.; Wilson, M.D.; Ballester, B.; Gonçalves, A.; Kutter, C.; Brown, G.D.; Marshall, A.; Flicek, P.; Odom, D.T. Waves of retrotransposon expansion remodel genome organization and CTCF binding in multiple mammalian lineages. Cell 2012, 148, 335–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crepaldi, L.; Policarpi, C.; Coatti, A.; Sherlock, W.T.; Jongbloets, B.C.; Down, T.A.; Riccio, A. Binding of TFIIIC to sine elements controls the relocation of activity-dependent neuronal genes to transcription factories. PLoS Genet. 2013, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miousse, I.R.; Chalbot, M.C.; Lumen, A.; Ferguson, A.; Kavouras, I.G.; Koturbash, I. Response of transposable elements to environmental stressors. Mutat. Res. Rev. Mutat. Res. 2015, 765, 19–39. [Google Scholar] [CrossRef] [Green Version]
- Fedoroff, N.V. Transposable elements, epigenetics, and genome evolution. Science 2012, 338, 758–767. [Google Scholar] [CrossRef] [Green Version]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef]
- Leebonoi, W.; Sukthaworn, S.; Panyim, S.; Udomkit, A. A novel gonad-specific Argonaute 4 serves as a defense against transposons in the black tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2015, 42, 280–288. [Google Scholar] [CrossRef]
- Berrens, R.V.; Andrews, S.; Spensberger, D.; Santos, F.; Dean, W.; Gould, P.; Sharif, J.; Olova, N.; Chandra, T.; Koseki, H.; et al. An endosiRNA-based repression mechanism counteracts transposon activation during global DNA demethylation in embryonic stem cells. Cell Stem Cell 2017, 21, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Pfefferli, C.; Jaźwińska, A. The art of fin regeneration in zebrafish. Regeneration 2015, 2, 72–83. [Google Scholar] [CrossRef]
- Wang, J.; Huang, J.; Shi, G. Retrotransposons in pluripotent stem cells. Cell Regen. 2020, 9, 4. [Google Scholar] [CrossRef]
- Grassi, D.A.; Jönsson, M.E.; Brattås, P.L.; Jakobsson, J. TRIM28 and the control of transposable elements in the brain. Brain Res. 2019, 1705, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Seah, M.; Wang, Y.; Goy, P.A.; Loh, H.M.; Peh, W.J.; Low, D.; Han, B.Y.; Wong, E.; Leong, E.L.; Wolf, G.; et al. The KRAB-zinc-finger protein ZFP708 mediates epigenetic repression at RMER19B retrotransposons. Development 2019, 146, dev170266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbeault, M.; Helleboid, P.Y.; Trono, D. KRAB zinc-finger proteins contribute to the evolution of gene regulatory networks. Nature 2017, 543, 550–554. [Google Scholar] [CrossRef]
- Ecco, G.; Cassano, M.; Kauzlaric, A.; Duc, J.; Coluccio, A.; Offner, S.; Imbeault, M.; Rowe, H.M.; Turelli, P.; Trono, D. Transposable elements and their KRAB-ZFP controllers regulate gene expression in adult tissues. Dev. Cell 2016, 36, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Carducci, F.; Carotti, E.; Gerdol, M.; Greco, S.; Canapa, A.; Barucca, M.; Biscotti, M.A. Investigation of the activity of transposable elements and genes involved in their silencing in the newt Cynops orientalis, a species with a giant genome. Sci. Rep. 2021, 11, 14743. [Google Scholar] [CrossRef] [PubMed]
- Helleboid, P.Y.; Heusel, M.; Duc, J.; Piot, C.; Thorball, C.W.; Coluccio, A.; Pontis, J.; Imbeault, M.; Turelli, P.; Aebersold, R.; et al. The interactome of KRAB zinc finger proteins reveals the evolutionary history of their functional diversification. EMBO J. 2019, 38, 18. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Saqib, H.; Lin, Z.; Zheng, R.; Qiu, Y.; Xie, Y.; Ma, D.; Shen, Y. RNA-seq analyses of marine medaka (Oryzias melastigma) reveals salinity responsive transcriptomes in the gills and livers. Aquat. Toxicol. 2021, 240, 105970. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Zdobnov, E.M. BUSCO: Assessing Genomic Data Quality and Beyond. Curr. Protoc. 2021, 1, e323. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Biscotti, M.A.; Gerdol, M.; Canapa, A.; Forconi, M.; Olmo, E.; Pallavicini, A.; Barucca, M.; Schartl, M. The lungfish transcriptome: A glimpse into molecular evolution events at the transition from water to land. Sci. Rep. 2016, 6, 21571. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, 296–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, C.; Boelens, R.; Bonvin, A.M. HADDOCK: A protein-protein docking approach based on biochemical or biophysical information. J. Am. Chem. Soc. 2003, 125, 1731–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Zundert, G.C.P.; Rodrigues, J.; Trellet, M.; Schmitz, C.; Kastritis, P.L.; Karaca, E.; Melquiond, A.; van Dijk, M.; de Vries, S.J.; Bonvin, A. The HADDOCK2.2 Web Server: User-Friendly Integrative Modeling of Biomolecular Complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honorato, R.V.; Koukos, P.I.; Jiménez-García, B.; Tsaregorodtsev, A.; Verlato, M.; Giachetti, A.; Rosato, A.; Bonvin, A. Structural biology in the clouds: The WeNMR-EOSC Ecosystem. Front. Mol. Biosci. 2021, 8, 729513. [Google Scholar] [CrossRef]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Friedli, M.; Trono, D. The developmental control of transposable elements and the evolution of higher species. Annu. Rev. Cell. Dev. Biol. 2015, 31, 429–451. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carotti, E.; Carducci, F.; Greco, S.; Gerdol, M.; Di Marino, D.; Perta, N.; La Teana, A.; Canapa, A.; Barucca, M.; Biscotti, M.A. Transcriptional Contribution of Transposable Elements in Relation to Salinity Conditions in Teleosts and Silencing Mechanisms Involved. Int. J. Mol. Sci. 2022, 23, 5215. https://doi.org/10.3390/ijms23095215
Carotti E, Carducci F, Greco S, Gerdol M, Di Marino D, Perta N, La Teana A, Canapa A, Barucca M, Biscotti MA. Transcriptional Contribution of Transposable Elements in Relation to Salinity Conditions in Teleosts and Silencing Mechanisms Involved. International Journal of Molecular Sciences. 2022; 23(9):5215. https://doi.org/10.3390/ijms23095215
Chicago/Turabian StyleCarotti, Elisa, Federica Carducci, Samuele Greco, Marco Gerdol, Daniele Di Marino, Nunzio Perta, Anna La Teana, Adriana Canapa, Marco Barucca, and Maria Assunta Biscotti. 2022. "Transcriptional Contribution of Transposable Elements in Relation to Salinity Conditions in Teleosts and Silencing Mechanisms Involved" International Journal of Molecular Sciences 23, no. 9: 5215. https://doi.org/10.3390/ijms23095215