
Comprehensive Analyses of Four PtoNF-YC Genes from Populus tomentosa and Impacts on Flowering Timing


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Cloning and Sequence Analysis of the Four PtoNF-YC Genes
2.3. PtoNF-YC Expressing Vector Construction and Subcellular Localization Assay
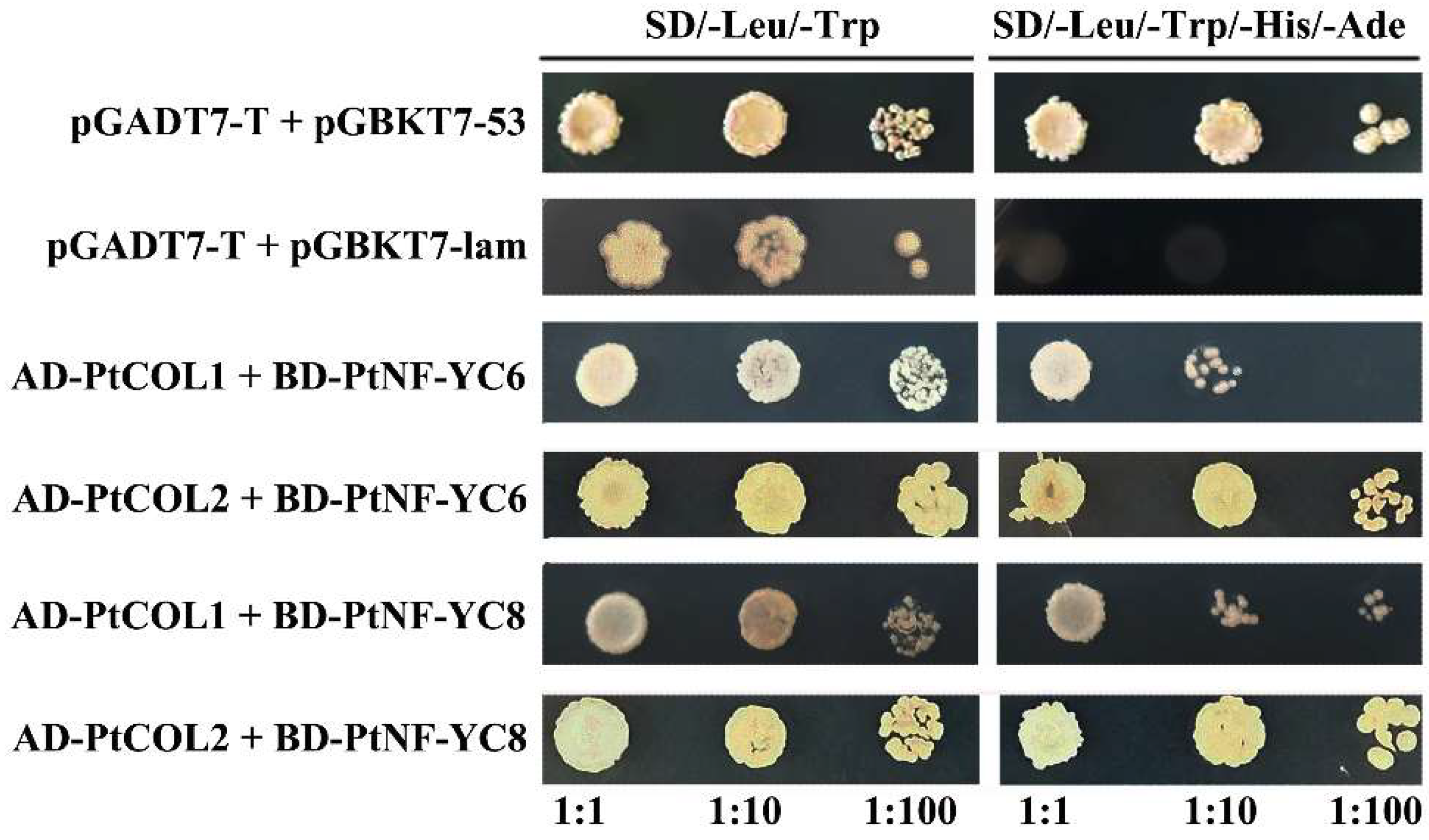
2.4. Analysis of the Interaction Mode of PtoNF-YCs and PtoCOL
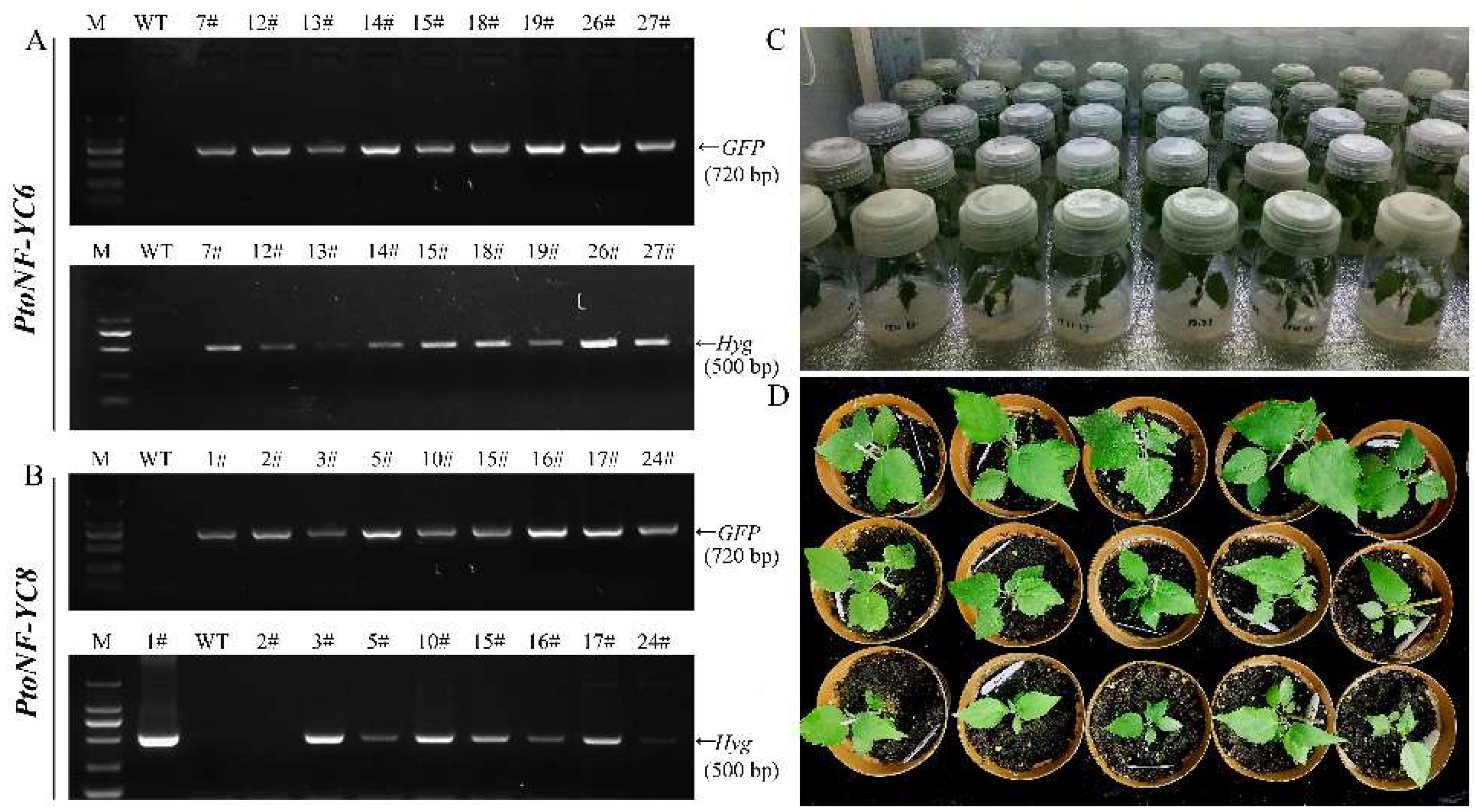
2.5. Arabidopsis and Poplar Transformation
2.6. Extraction of RNA and Quantitative PCR Analysis
2.7. Phenotypic Analysis and Biomass Measurements
2.8. Statistical Analyses
3. Results
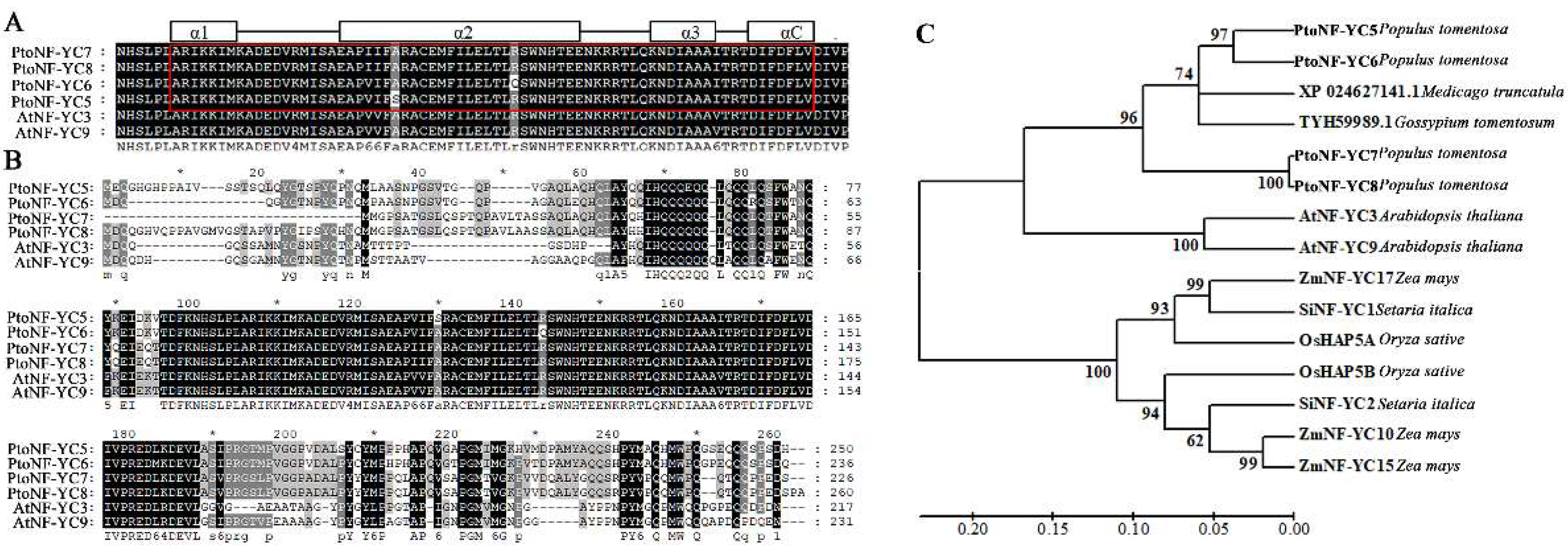
3.1. Cloning and Sequence Analysis of the Four PtoNF-YC Genes
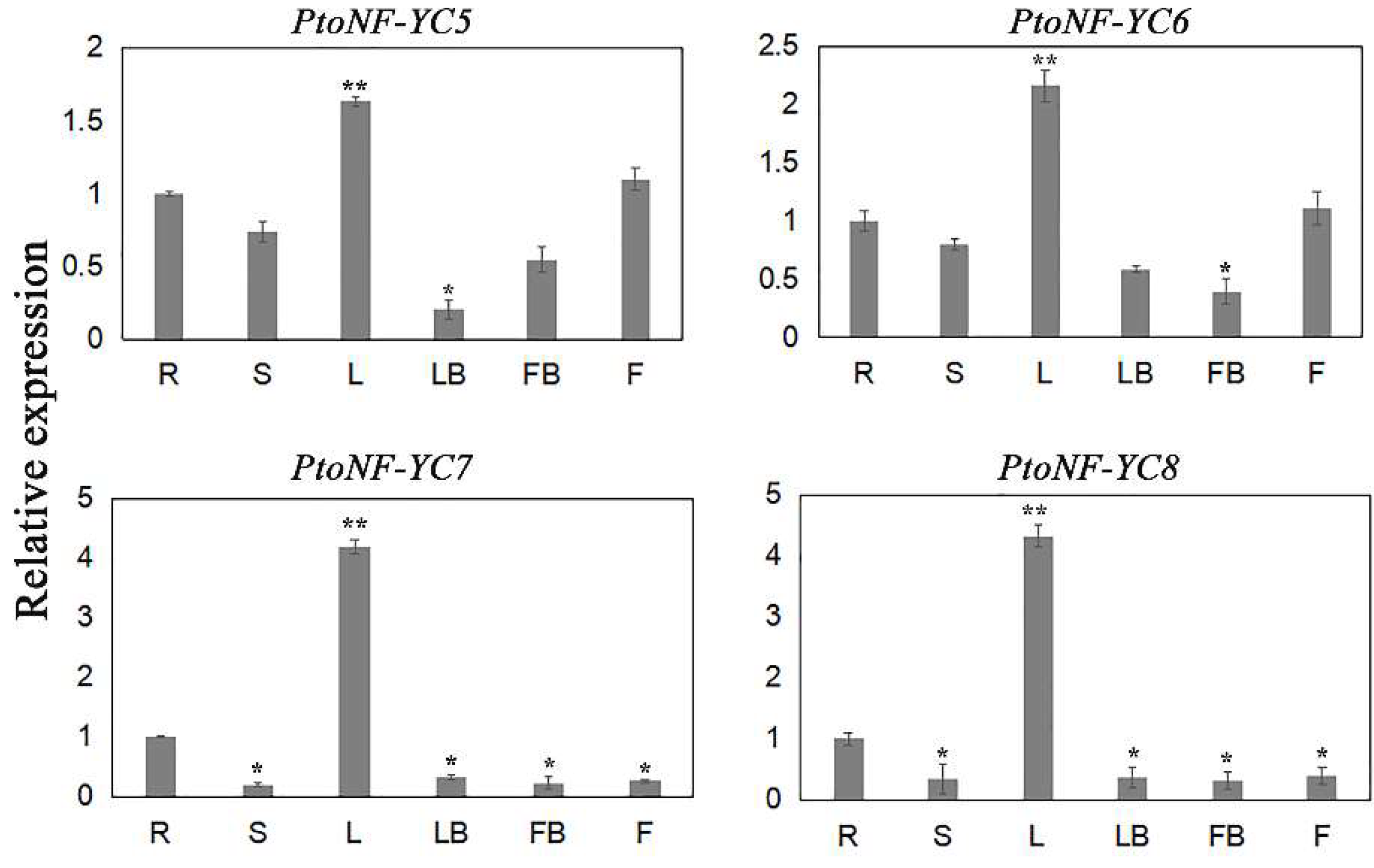
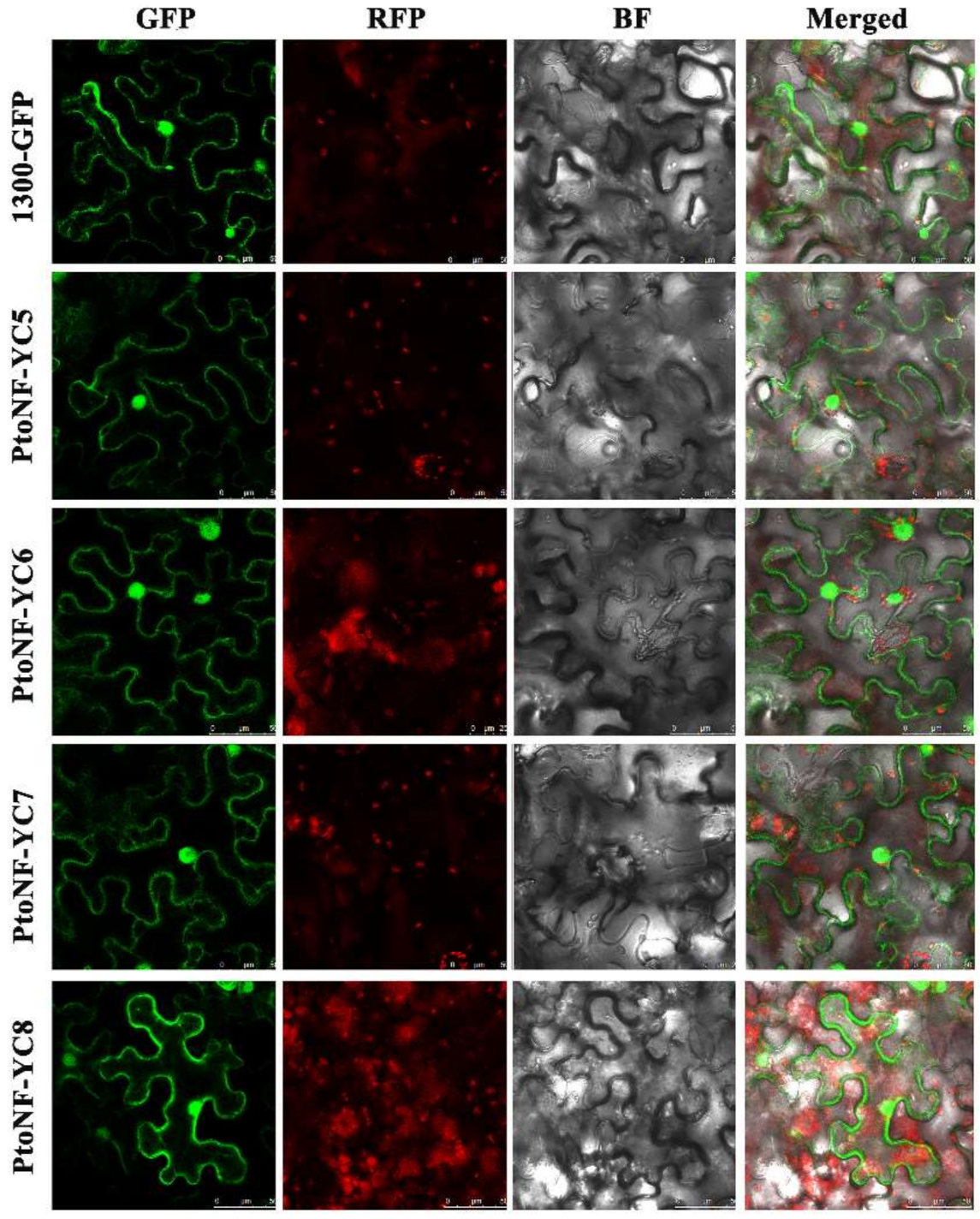
3.2. Tissue-Specific Expression and Subcellular Localization Analysis of the Four PtoNF-YC Genes
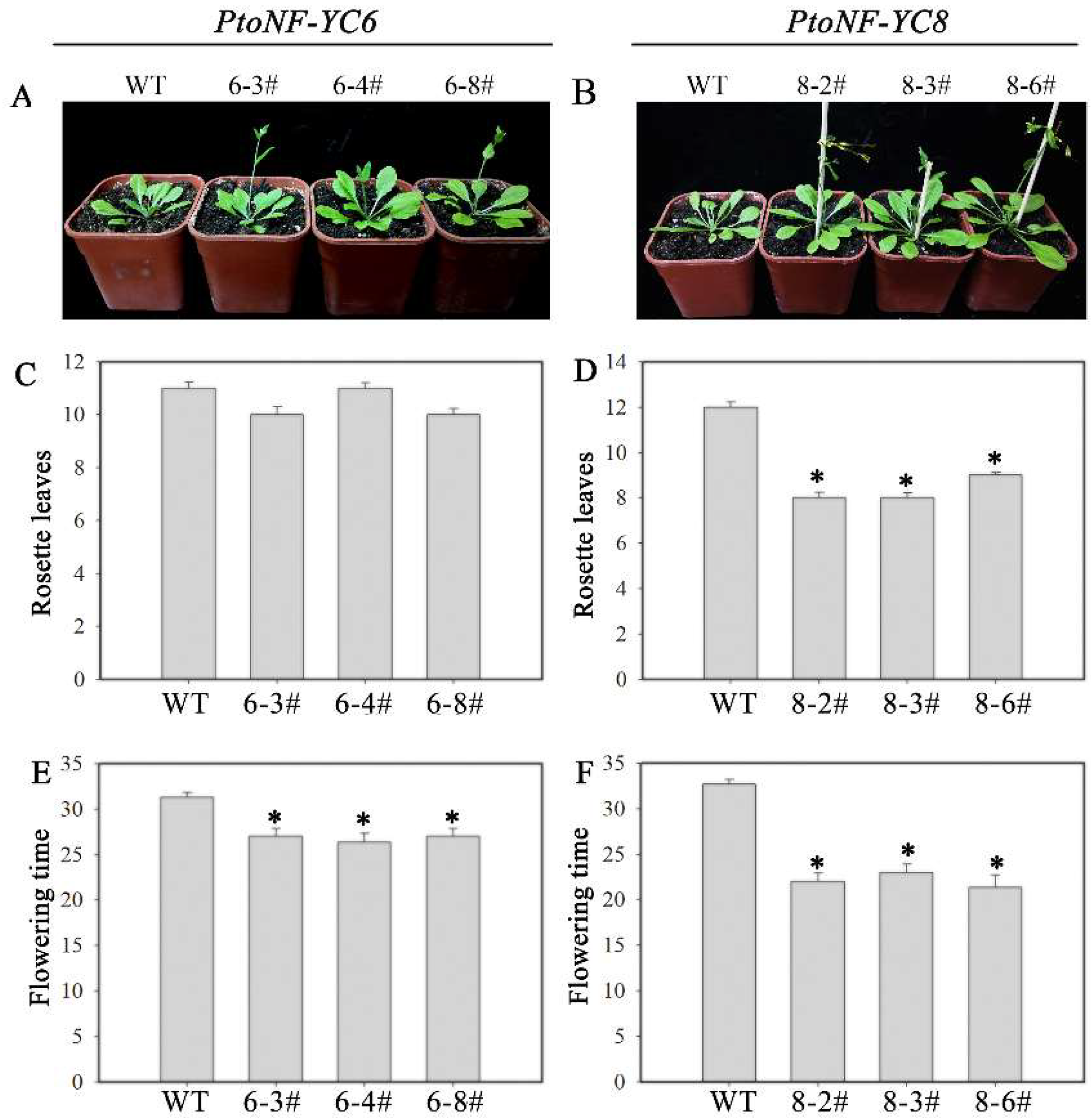
3.3. Effect of Ectopic Expression of PtoNF-YC6/8 on Early Flowering in Transgenic Arabidopsis
3.4. Analysis of the Effects of PtoNF-YC6 and PtoNF-YC8 on the Floral Organs of Arabidopsis
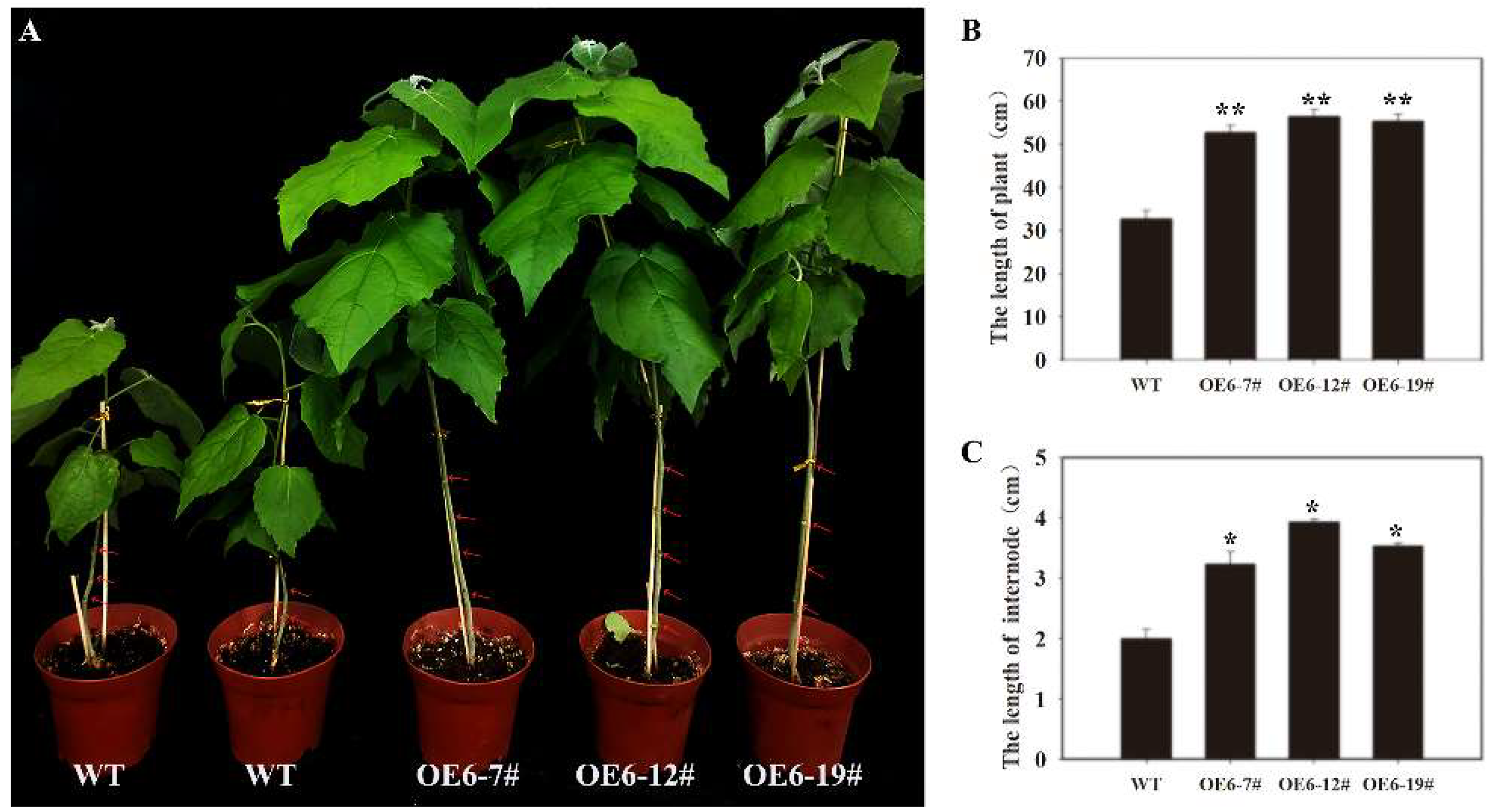
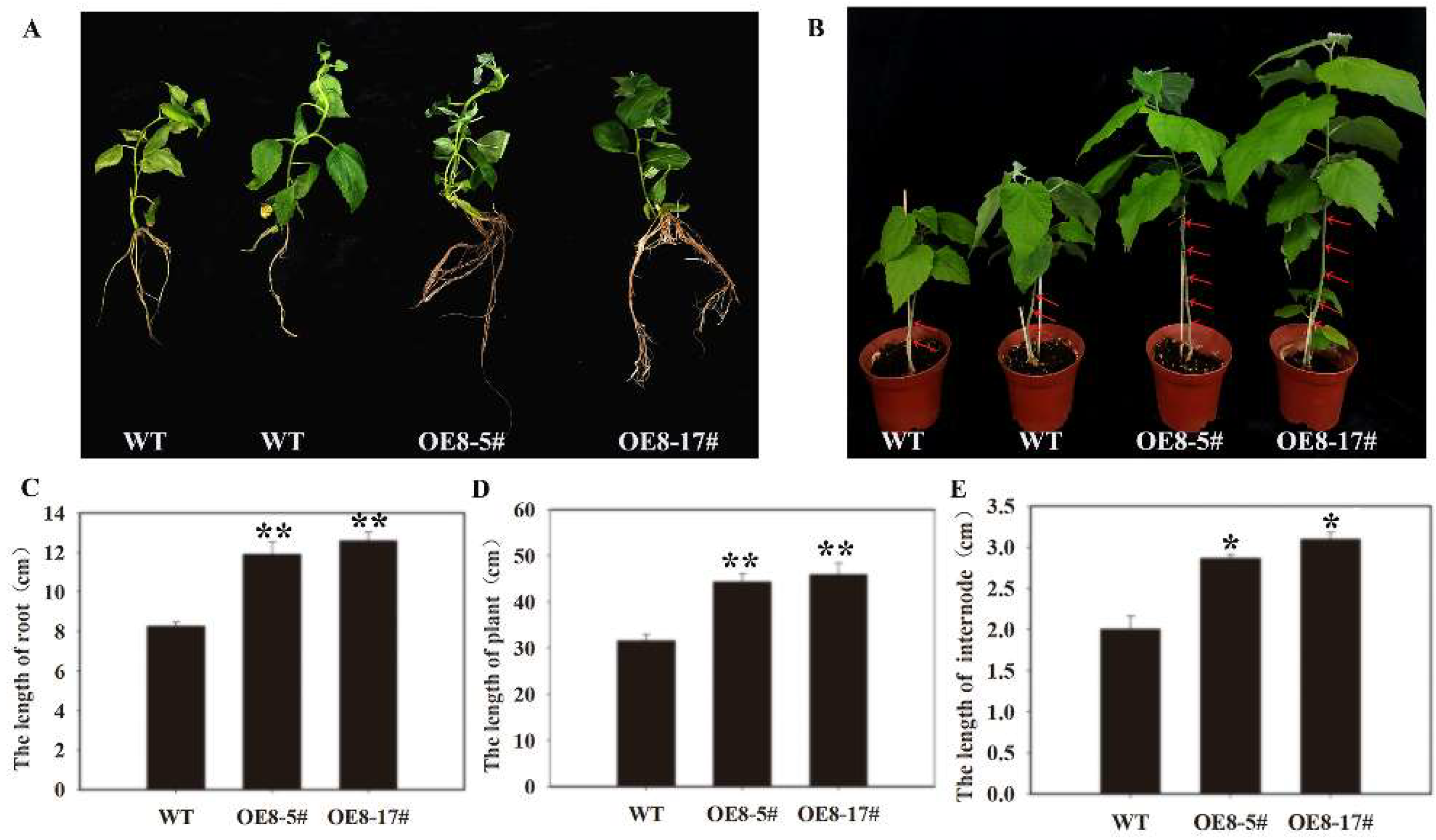
3.5. Effects of PtoNF-YC6 and PtoNF-YC8 on the Growth of P. tomentosa
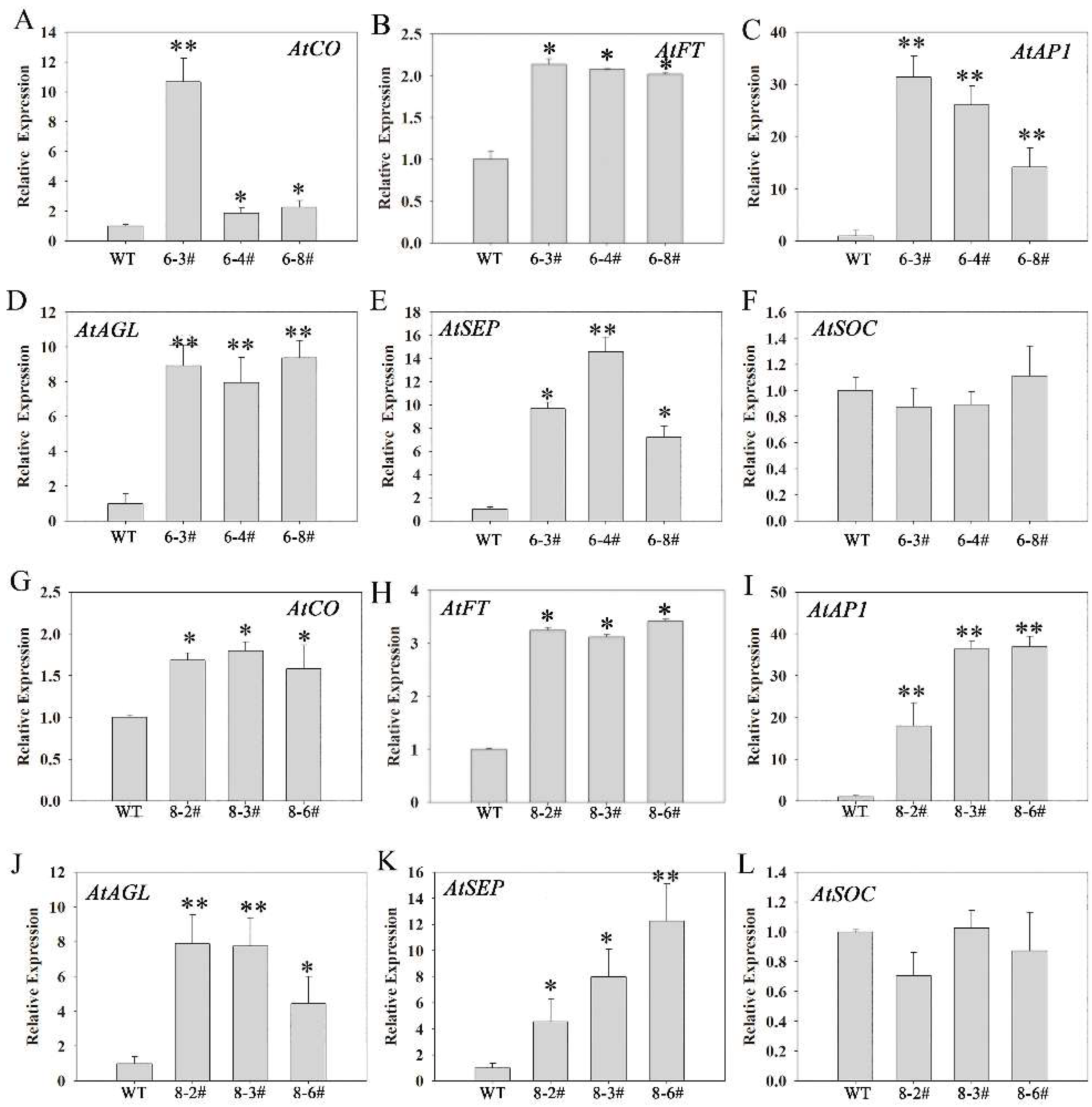
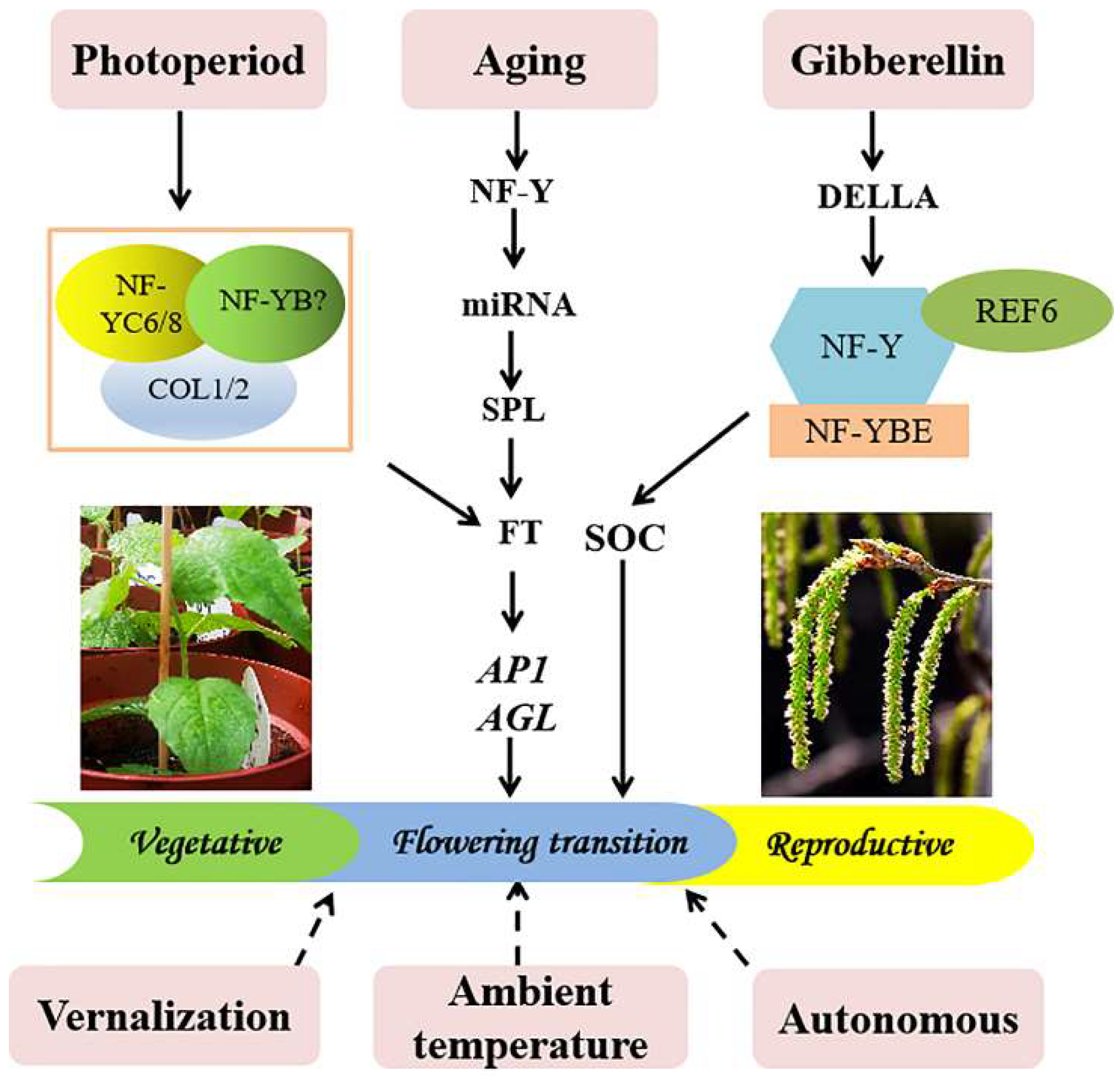
3.6. The Possible Molecular Mechanism of PtoNF-YC6/8 Expression in P. tomentosa Promoting Early Flowering
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsu, C.Y.; Adams, J.P.; Kim, H.; No, K.; Ma, C.; Strauss, S.H.; Drnevich, J.; Vandervelde, L.; Ellis, J.D.; Rice, B.M.; et al. FLOWERING LOCUS T duplication coordinates reproductive and vegetative growth in perennial poplar. Proc. Natl. Acad. Sci. USA 2011, 108, 10756–10761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, X.M.; Wang, D.M.; Wang, Z.L.; Li, B.; Bo, W.H.; Cao, G.L.; Zhang, Z.Y. Isolation of a LEAFY homolog from Populus tomentosa: Expression of PtLFY in P. tomentosa floral buds and PtLFY-IR-mediated gene silencing in tobacco (Nicotiana tabacum). Plant Cell Rep. 2011, 30, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhu, L.; Zhang, Y.; Fan, J.; Li, L. Genome-wide analysis of poplar NF-YB gene family and identified PtNF-YB1 important in regulate flowering timing in transgenic plants. BMC Plant Biol. 2019, 19, 251. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Rao, P.; Yang, X.; Su, X.; Zhao, T.; Gao, K.; Yang, X.; An, X. A Global view of transcriptome dynamics during male floral bud development in Populus tomentosa. Sci. Rep. 2018, 8, 722. [Google Scholar] [CrossRef] [Green Version]
- Bucher, P.; Trifonov, E.N. CCAAT box revisited: Bidirectionality, location and context. J. Biomol. Struct. Dyn. 1988, 5, 1231–1236. [Google Scholar] [CrossRef] [PubMed]
- Roberto, M. A survey of 178 NF-Y binding CCAAT boxes. Nucleic Acids Res. 1998, 5, 1135–1143. [Google Scholar]
- McNabb, D.S.; Xing, Y.; Guarente, L. Cloning of yeast HAP5: A novel subunit of a heterotrimeric complex required for CCAAT binding. Genes Dev. 1995, 9, 47–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, D.; Murray, A.H.; Smith, A.G. Multiple genes encoding the conserved CCAAT-Box transcription factor complex are expressed in Arabidopsis. Plant Physiol. 1998, 117, 1015–1022. [Google Scholar] [CrossRef] [Green Version]
- Maruyuma, K.; Todaka, D.; Mizoi, J.; Yoshida, T.; Kidokoro, S.; Matsukura, S.; Takasaki, H.; Sakurai, T.; Yamamoto, Y.Y.; Yoshiwawa, K. Identification of Cis-acting promoter elements in cold and dehydration-induced transcriptional pathways in Arabidopsis, rice, and soybean. DNA Res. 2011, 19, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Laloum, T.; Mita, D.S.; Gamas, P.; Baudin, M.; Niebel, A. CCAAT-box binding transcription factors in plants: Y so many? Trends Plant Sci. 2013, 18, 157–166. [Google Scholar] [CrossRef]
- Li, X.Y.; Huijsduijnen, R.; Mantovani, R.; Benoist, C.; Mathis, D. Intron-exon organization of the NF-Y genes. Tissue-specific splicing modifies an activation domain. J. Biol. Chem. 1992, 267, 8984. [Google Scholar] [CrossRef]
- Siefers, N.; Dang, K.K.; Kumimoto, R.W.; Bynum, W.E., IV; Tayrose, G.; Holt, B.F., III. Tissue-specific expression patterns of Arabidopsis NF-Y transcription factors suggest potential for extensive combinatorial complexity. Plant Physiol. 2008, 149, 625–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirumurugan, T.; Ito, Y.; Kubo, T.; Serizawa, A.; Kurata, N. Identification, characterization and interaction of HAP family genes in rice. J. Recept. Res. 2008, 7, 279–289. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, X.; Zhang, C.; Zou, H.; Wu, Z. Isolation, structural analysis, and expression characteristics of the maize nuclear factor Y gene families. Biochem. Biophys. Res. Commun. 2016, 478, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, K.; Ju, Z.; Cao, D.; Fu, D.; Zhu, H.; Zhu, B.; Luo, Y. Genome-wide analysis of tomato NF-Y factors and their role in fruit ripening. BMC Genom. 2016, 17, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gao, K.; Khan, W.U.; Yang, X.; Yang, X.; Zhao, T.; Chen, Z.; An, X. Genome-wide analysis of the poplar NF-Y gene family and its expression in floral bud development of Populus tomentosa. Trees 2019, 34, 285–296. [Google Scholar] [CrossRef]
- Guo, Y.; Niu, S.; El-Kassaby, Y.A.; Li, W. Transcriptome-wide isolation and expression of NF-Y gene family in male cone development and hormonal treatment of Pinus tabuliformis. Physiol. Plant. 2021, 171, 34–47. [Google Scholar] [CrossRef]
- Calvenzani, V.; Testoni, B.; Gusmaroli, G.; Lorenzo, M.; Gnesutta, N.; Petroni, K.; Mantovani, R.; Tonelli, C. Interactions and CCAAT-Binding of Arabidopsis thaliana NF-Y Subunits. PLoS ONE 2012, 7, e42902. [Google Scholar] [CrossRef] [Green Version]
- Petroni, K.; Kumimoto, R.W.; Gnesutta, N.; Calvenzani, V.; Fornari, M.; Tonelli, C.; Holt, B.F.; Mantovani, R. The promiscuous life of plant NUCLEAR FACTOR Y transcription factors. Plant Cell 2012, 24, 4777–4792. [Google Scholar] [CrossRef] [Green Version]
- Fornari, M.; Calvenzani, V.; Masiero, S.; Tonelli, C.; Petroni, K.J.P.O. The Arabidopsis NF-YA3 and NF-YA8 genes are functionally redundant and are required in early embryogenesis. PLoS ONE 2013, 8, e82043. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hu, P.; Huang, M.; Tang, Y.; Li, Y.; Li, L.; Hou, X. The NF-YC-RGL2 module integrates GA and ABA signalling to regulate seed germination in Arabidopsis. Nat. Commun. 2016, 7, 12768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumimoto, R.W.; Zhang, Y.; Siefers, N.; Holt, B.F. NF-YC3, NF-YC4 and NF-YC9 are required for CONSTANS-mediated, photoperiod-dependent flowering in Arabidopsis thaliana. Plant J. 2010, 63, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Ma, C.; Xu, Y.; Wang, T.; Chen, Y.; Lü, J.; Zhang, L.; Jiang, C.-Z.; Hong, B.; Gao, J. Control of chrysanthemum flowering through integration with an aging pathway. Nat. Commun. 2017, 8, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, G.; Li, C.; Zhang, C.; Cui, L.; Ai, G.; Wang, X.; Zheng, F.; Zhang, D.; Larkin, R.M.; et al. NF-Y plays essential roles in flavonoid biosynthesis by modulating histone modifications in tomato. New Phytol. 2021, 229, 3237–3252. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Oono, Y.; Zhu, J.; He, X.J.; Wu, J.; Iida, K.; Lu, X.Y.; Cui, X.; Jin, H.; Zhu, J.K. The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and posttranscriptionally to promote drought resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, W.; Chen, Z.; Han, B.; Haque, M.E.; Liu, A. Gene structure, expression pattern and interaction of Nuclear Factor-Y family in castor bean (Ricinus communis). Planta 2018, 247, 559–572. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, T.; Takahashi, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K.J.P.P. NF-YB2 and NF-YB3 have functionally diverged and differentially induce drought and heat stress-specific genes. Plant Physiol. 2019, 80, 1677–1690. [Google Scholar] [CrossRef] [Green Version]
- Hou, X. Nuclear factor Y-mediated H3K27me3 demethylation of the SOC1 locus orchestrates flowering responses of Arabidopsis. Nat. Commun. 2014, 5, 4601. [Google Scholar] [CrossRef] [Green Version]
- Lian, C.; Li, Q.; Yao, K.; Zhang, Y.; Meng, S.; Yin, W.; Xia, X. Populus trichocarpa PtNF-YA9, a multifunctional transcription factor, regulates seed germination, abiotic stress, plant growth and development in Arabidopsis. Front. Plant Sci. 2018, 9, 954. [Google Scholar] [CrossRef]
- Li, J.; Gao, K.; Lei, B.; Zhou, J.; Guo, T.; An, X. Altered sucrose metabolism and plant growth in transgenic Populus tomentosa with altered sucrose synthase PtSS3. Transgenic Res. 2020, 29, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Mangalam, A.; Dwivedi, S.; Naik, S.J.B. Primer premier: Program for design of degenerate primers from a protein sequence. Biotechniques 1998, 24, 318–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Li, J.; Lu, J.; Tian, B.; Liu, X.; Yang, G.; Pei, Y. Cloning and functional analysis of four O-Acetylserine (thiol) lyase family genes from foxtail millet. Plant Physiol. Biochem. 2019, 139, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Heijne, G.; Sonnhammer, E.L.L. Bioinformatics SJJoMB. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 7, 1870. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 2010, 6, 735–743. [Google Scholar]
- Zhang, Q.; Zhang, Z.Y.; Lin, S.Z.; Zheng, H.Q.; Lin, Y.Z.; An, X.M.; Li, Y.; Li, H.X. Characterization of resistance gene analogs with a nucleotide binding site isolated from a triploid white poplar. Plant Biol. 2008, 10, 310–322. [Google Scholar] [CrossRef]
- Zheng, H.; Lin, S.; Zhang, Q.; Lei, Y.; Zhang, Z. Functional analysis of 5′untranslated region of a TIR-NBS-encoding gene from triploid white poplar. Mol. Gen. Genet. 2009, 282, 381–394. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 −ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Putterill, J.; Robson, F.; Lee, K.; Simon, R.; Coupland, G.J.C. The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell 1995, 80, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Gao, K.; Yang, X.; Khan, W.U.; Guo, B.; Guo, T.; An, X. Identification and characterization of the CONSTANS-like gene family and its expression profiling under light treatment in Populus. Int. J. Biol. Macromol. 2020, 161, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz, C.; Gu, Q.; Martienssen, R.; Yanofsky, M.F.J.D. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Zhao, Y.; Zhuo, C.; Lu, S.; Guo, Z.J.P.B.J. Overexpression of a NF-YC transcription factor from bermudagrass confers tolerance to drought and salinity in transgenic rice. Plant Biotechnol. J. 2015, 13, 482–491. [Google Scholar]
- Ma, X.; Li, C.; Wang, M.J.B. Wheat NF-YA10 functions independently in salinity and drought stress. Bioengineered 2015, 6, 245–247. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Wu, M.; Liu, H.L.; Gao, Y.M.; Chen, J.; Yan, H.W.; Xiang, Y. Genome-wide identification and expression analysis of the NF-Y transcription factor family in Populus. Physiol. Plant. 2021, 171, 309–327. [Google Scholar] [CrossRef]
- Hackenberg, D.; Keetman, U.; Grimm, B. Homologous NF-YC2 subunit from Arabidopsis and tobacco is activated by photooxidative stress and induces flowering. Int. J. Mol. Sci. 2012, 13, 3458–3477. [Google Scholar] [CrossRef] [Green Version]
- Ben-Naim, O.; Eshed, R.; Parnis, A.; Teper-Bamnolker, P.; Shalit, A.; Coupland, G.; Samach, A.; Lifschitz, E. The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. Plant J. 2006, 46, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, T.J.; McIntyre, C.L.; Collet, C.; Xue, G.P. TaNF-YC11, one of the light-upregulated NF-YC members in Triticum aestivum, is co-regulated with photosynthesis-related genes. Funct. Integr. Genom. 2010, 10, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.F.; Wang, Z.Y. Overexpression of COL9, a CONSTANS-LIKE gene, delays flowering by reducing expression of CO and FT in Arabidopsis thaliana. Plant J. 2005, 43, 758–768. [Google Scholar] [CrossRef]
- Zicola, J.; Liu, L.; Tänzler, P.; Turck, F. Targeted DNA methylation represses two enhancers of FLOWERING LOCUS T in Arabidopsis thaliana. Nat. Plants 2019, 5, 300–307. [Google Scholar] [CrossRef]
- Myers, Z.A.; Holt, B.F. NUCLEAR FACTOR-Y: Still complex after all these years? Curr. Opin. Plant Biol. 2018, 45, 96–102. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Gao, K.; Yang, X.; Guo, B.; Xue, Y.; Miao, D.; Huang, S.; An, X. Comprehensive Analyses of Four PtoNF-YC Genes from Populus tomentosa and Impacts on Flowering Timing. Int. J. Mol. Sci. 2022, 23, 3116. https://doi.org/10.3390/ijms23063116
Li J, Gao K, Yang X, Guo B, Xue Y, Miao D, Huang S, An X. Comprehensive Analyses of Four PtoNF-YC Genes from Populus tomentosa and Impacts on Flowering Timing. International Journal of Molecular Sciences. 2022; 23(6):3116. https://doi.org/10.3390/ijms23063116
Chicago/Turabian StyleLi, Juan, Kai Gao, Xiaoyu Yang, Bin Guo, Yinxuan Xue, Deyu Miao, Sai Huang, and Xinmin An. 2022. "Comprehensive Analyses of Four PtoNF-YC Genes from Populus tomentosa and Impacts on Flowering Timing" International Journal of Molecular Sciences 23, no. 6: 3116. https://doi.org/10.3390/ijms23063116