
Chronic Monocular Deprivation Reveals MMP9-Dependent and -Independent Aspects of Murine Visual System Plasticity

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Extracellular Proteolysis Enables Plasticity in Immature and Mature Circuits
2. Results
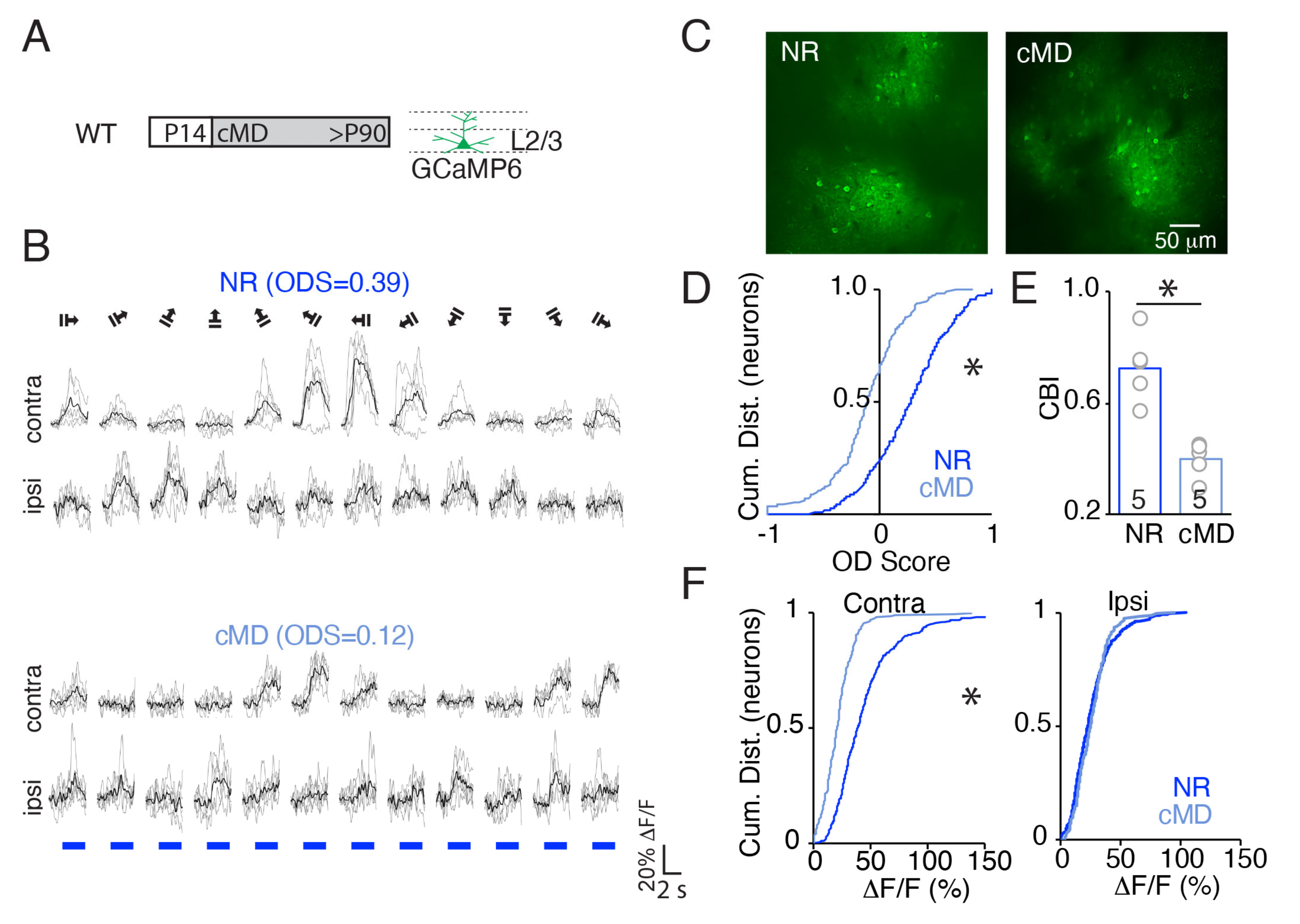
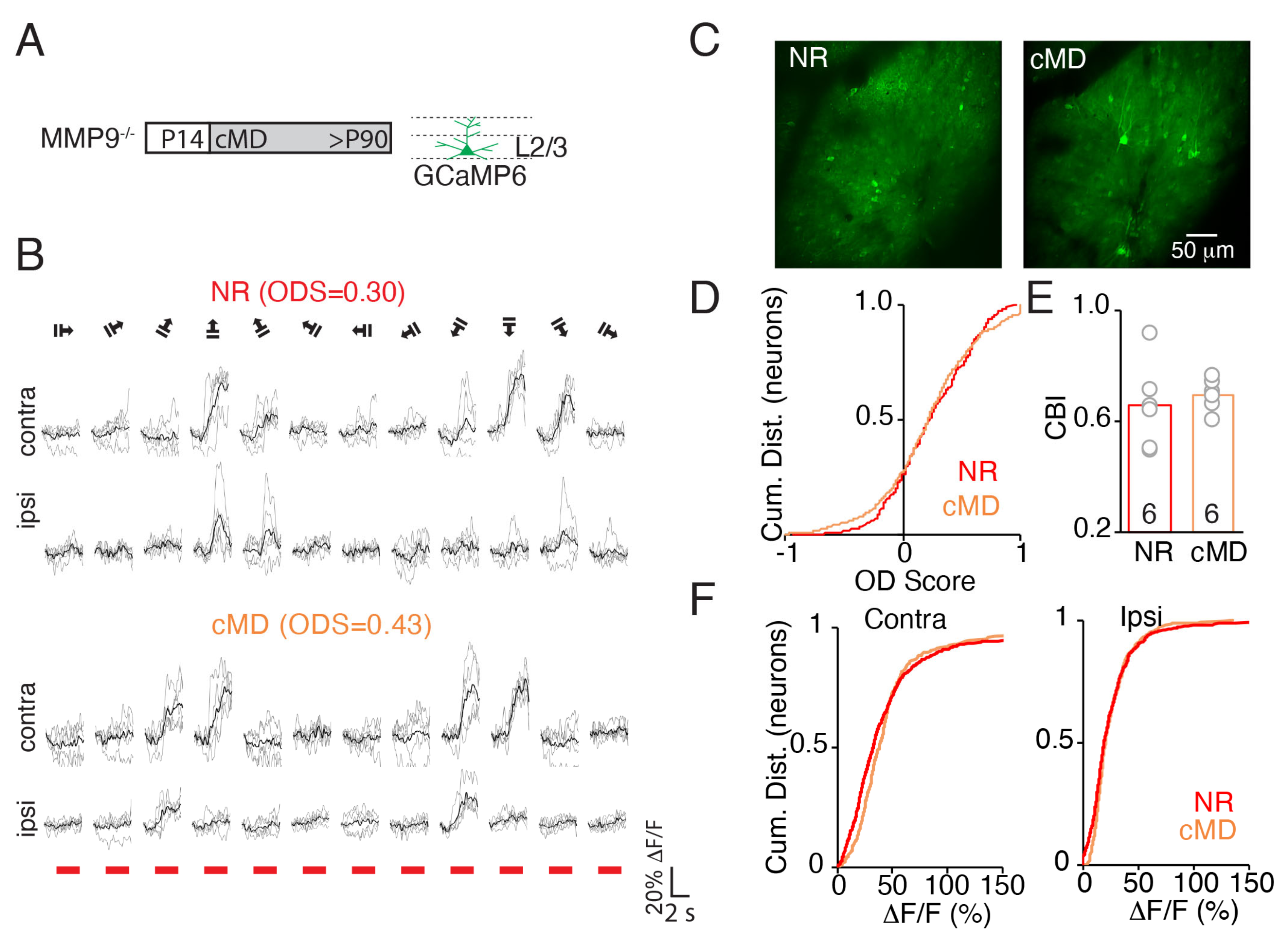
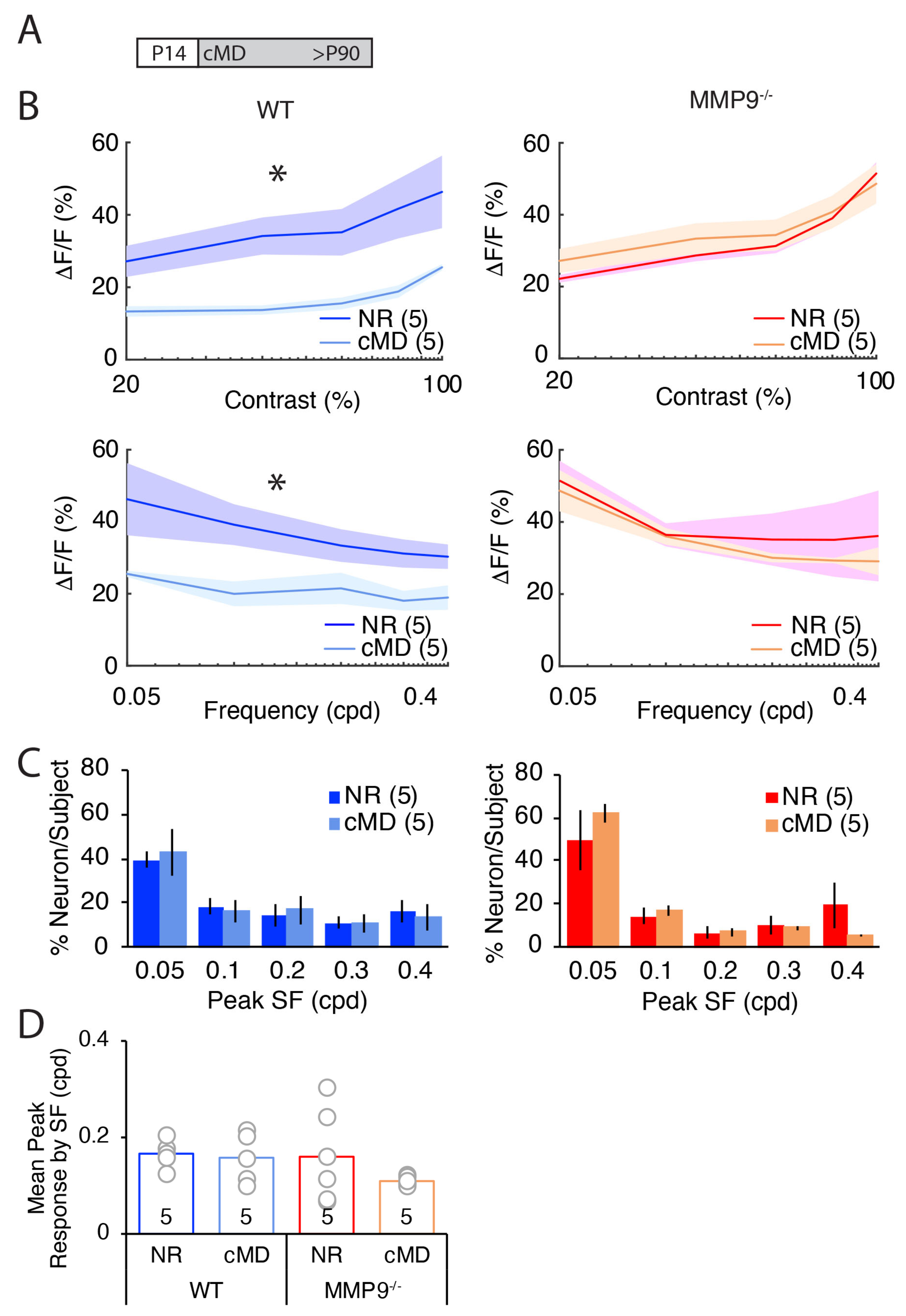
2.1. cMD Decreases the Strength of the Deprived Eye Pathway of WT but Not MMP9−/− Mice
2.2. cMD Decreases the Strength, but Not the Selectivity of the Deprived Eye Pathway of WT but Not MMP9−/− Mice
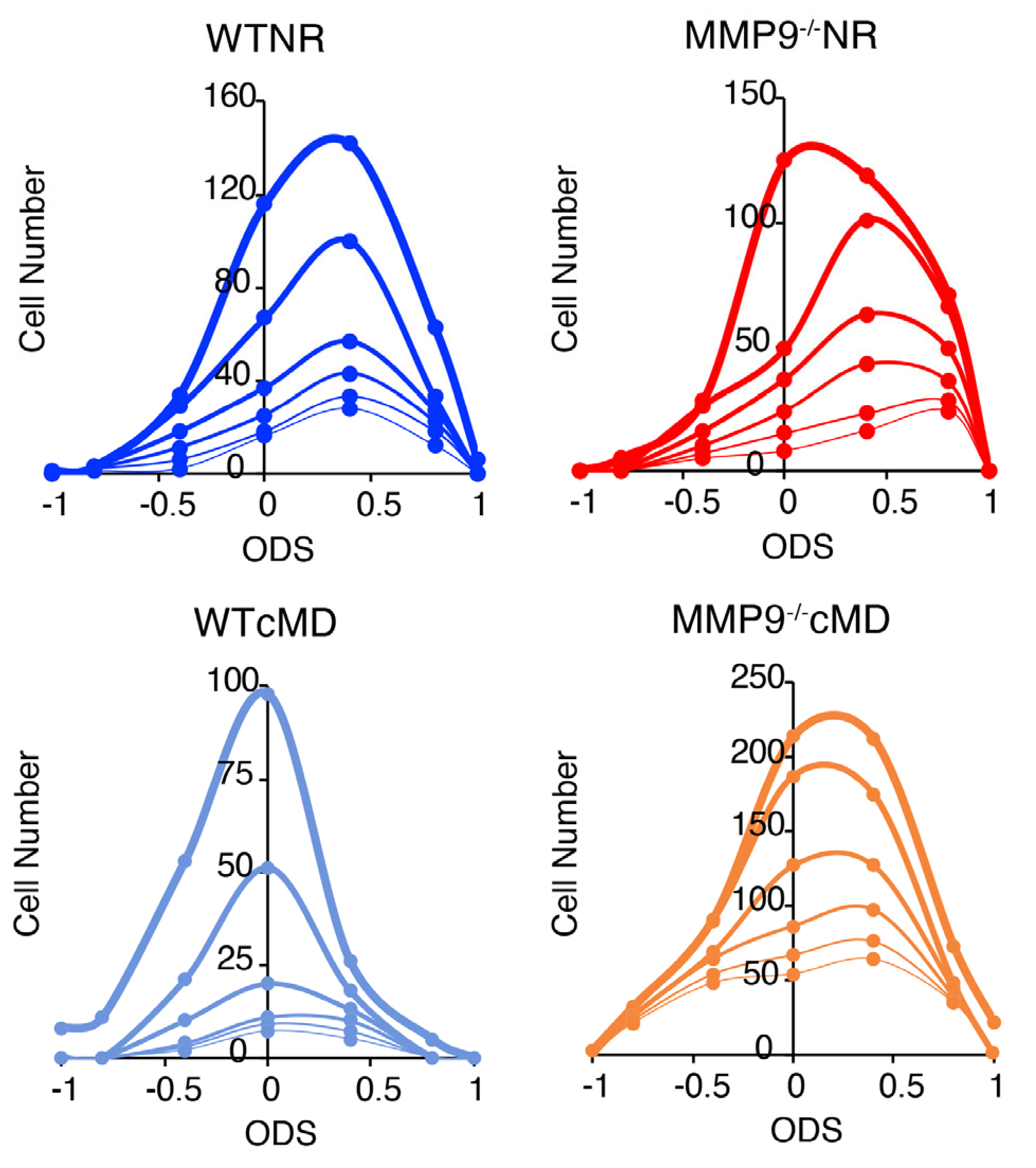
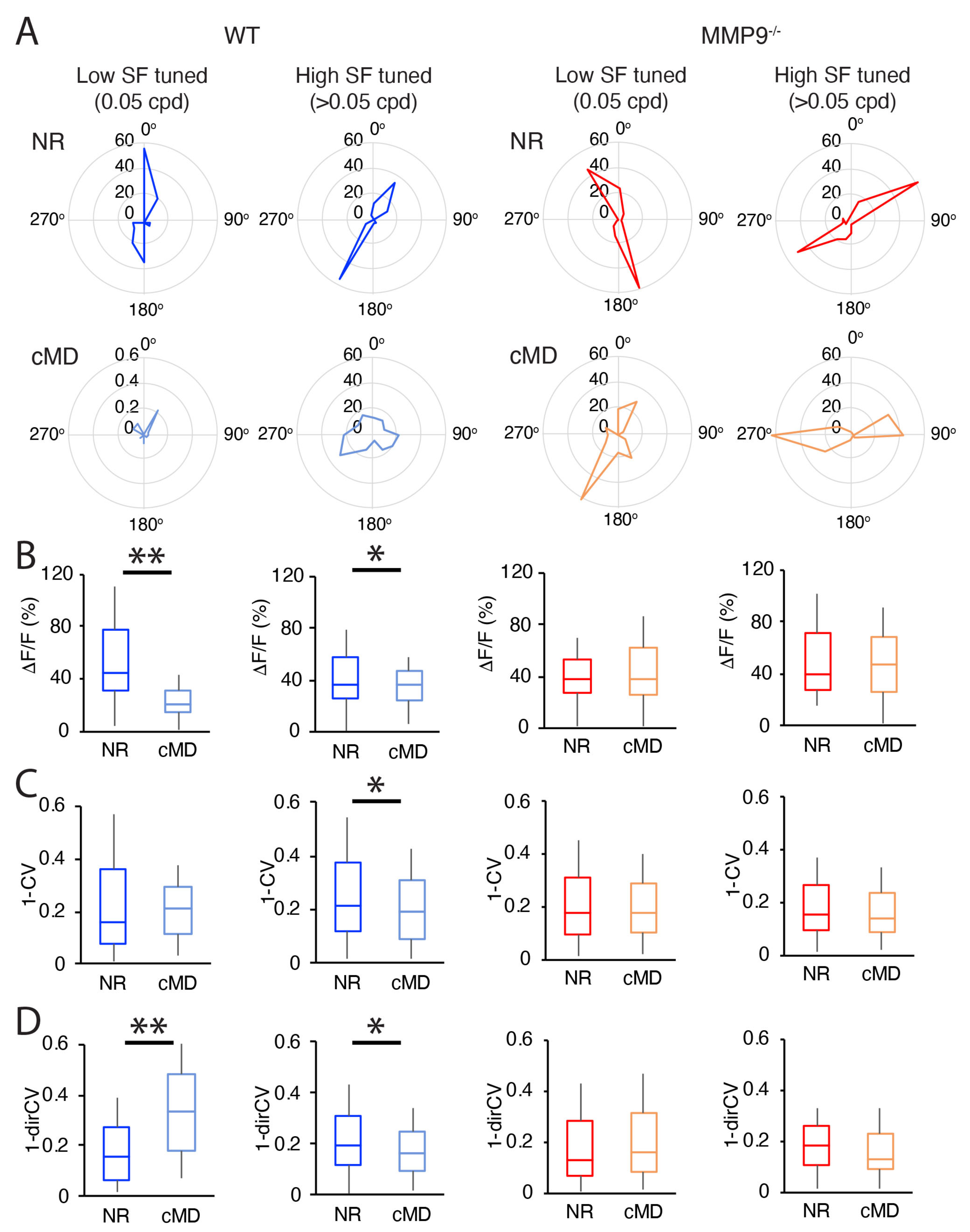
2.3. cMD Impacts the Strength and Selectivity to Visual Stimulus Orientation and Direction in WT but Not MMP9−/− Mice
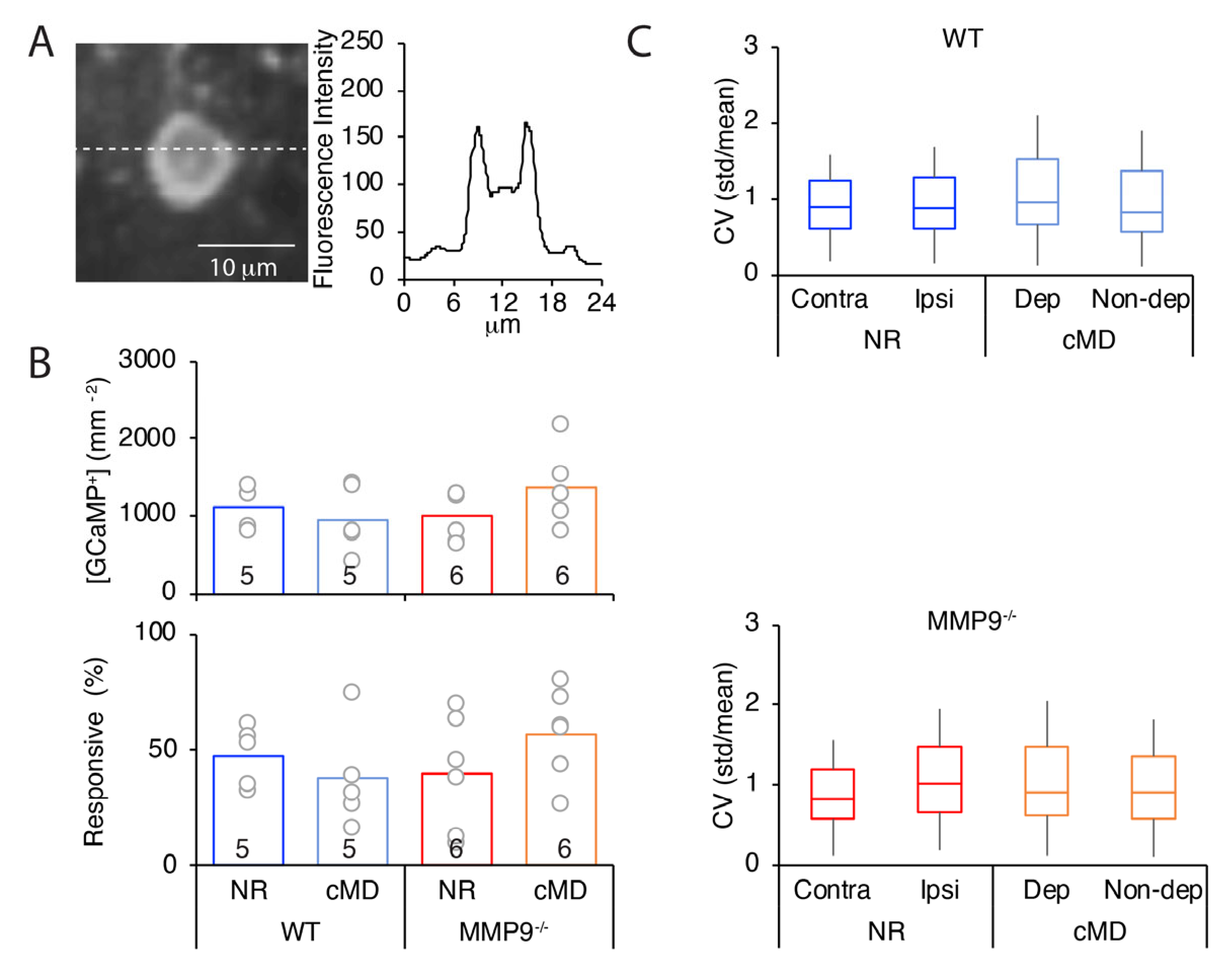
2.4. Anatomical Correlates of cMD in WT and MMP9−/− Mice
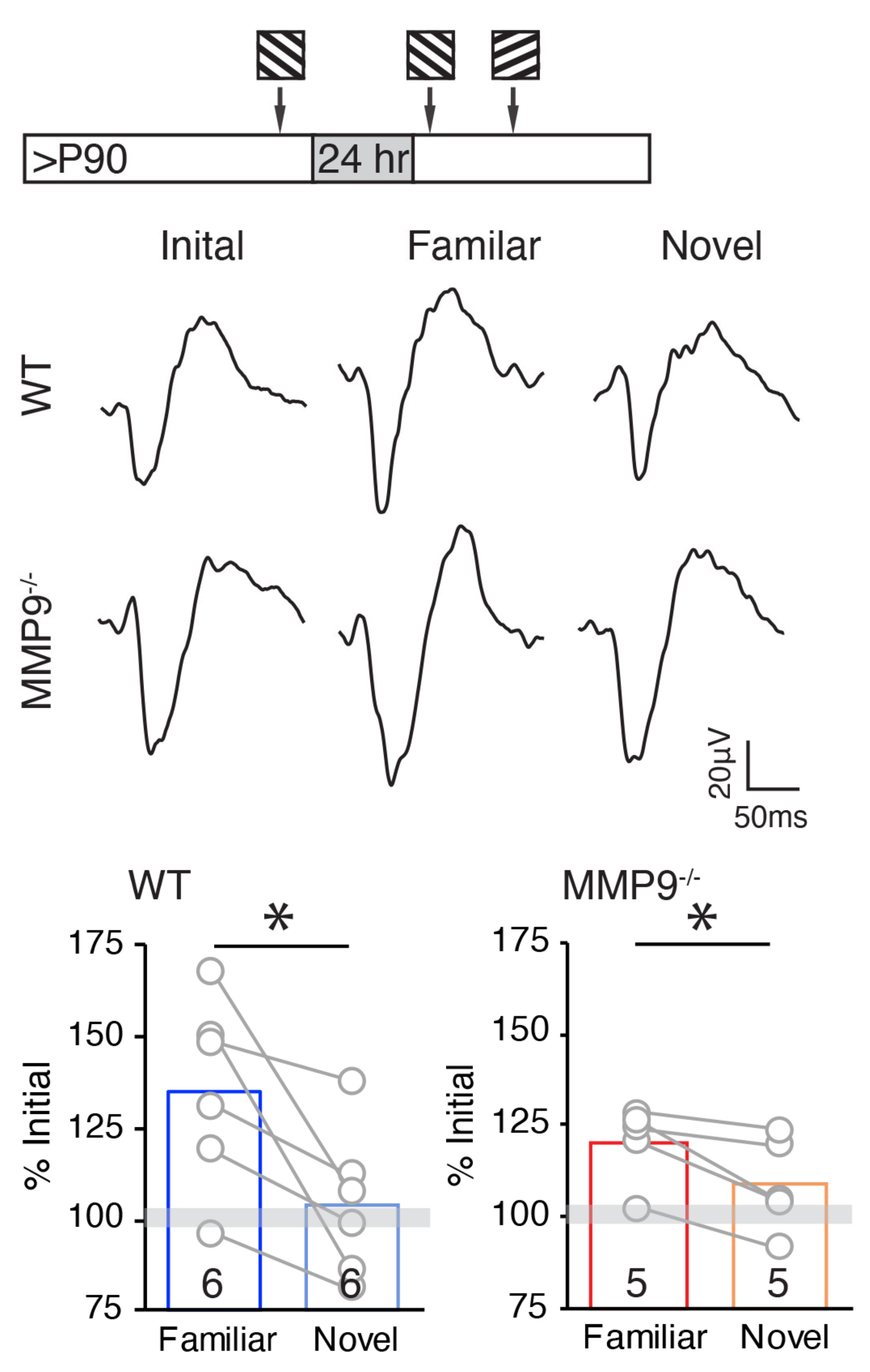
2.5. Persistence of Stimulus-Selective Response Potentiation (SRP) in Adult MMP9−/− Mice
3. Discussion
3.1. cMD Regulates Visual Response Strength and Selectivity in WT but Not MMP9−/− Mice
3.2. cMD and MD during the Critical Period Recruit Distinct Forms of Plasticity
3.3. Assessment of Binocularity in Mouse V1b
3.4. Structural and Functional Plasticity Induced by MMP9 Activity
3.5. Developmental Expression of MMP9
4. Materials and Methods
4.1. Subjects
4.2. Chronic Monocular Deprivation
4.3. Immunohistochemistry
4.4. Golgi staining and Dendritic Spine Density Analysis
4.5. Intraocular Injections of Anterograde Tracer
4.6. Virus Injection and Cranial Window Implantation
4.7. Two-Photon Imaging
4.8. Ocular Dominance Analysis
4.9. Orientation and Direction Selectivity Analysis
4.10. Chronic In Vivo Recordings for Stimulus-Selective Response Potentiation (SRP)
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuhlman, S.J.; Olivas, N.D.; Tring, E.; Ikrar, T.; Xu, X.; Trachtenberg, J.T. A disinhibitory microcircuit initiates critical-period plasticity in the visual cortex. Nature 2013, 501, 543–546. Available online: http://www.Nature.com/articles/Nature12485 (accessed on 23 January 2019). [CrossRef] [PubMed] [Green Version]
- Frenkel, M.Y.; Bear, M.F. How monocular deprivation shifts ocular dominance in visual cortex of young mice. Neuron 2004, 44, 917–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mataga, N.; Mizuguchi, Y.; Hensch, T.K. Experience-dependent pruning of dendritic spines in visual cortex by tissue plasminogen activator. Neuron 2004, 44, 1031–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Lai, B.; Gan, W.B. Monocular deprivation induces dendritic spine elimination in the developing mouse visual cortex. Sci. Rep. 2017, 7, 4977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawtell, N.B.B.; Frenkel, M.Y.Y.; Philpot, B.D.D.; Nakazawa, K.; Tonegawa, S.; Bear, M.F.F. NMDA receptor-dependent ocular dominance plasticity in adult visual cortex. Neuron 2003, 38, 977–985. Available online: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=12818182%5Cnhttp://www.sciencedirect.com/science/article/pii/s0896627303003234 (accessed on 31 December 2021). [CrossRef] [Green Version]
- Giffin, F.; Mitchell, D.E. The rate of recovery of vision after early monocular deprivation in kittens. J. Physiol. 1978, 274, 511–537. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, D.E.; Gingras, G. Visual recovery after monocular deprivation is driven by absolute, rather than relative, visually evoked activity levels. Curr. Biol. 1998, 8, 1179–1182. [Google Scholar] [CrossRef] [Green Version]
- Liao, D.S.; Mower, A.F.; Neve, R.L.; Sato-Bigbee, C.; Ramoa, A.S. Different mechanisms for loss and recovery of binocularity in the visual cortex. J. Neurosci. 2002, 22, 9015–9023. [Google Scholar] [CrossRef] [Green Version]
- He, H.-Y.; Ray, B.; Dennis, K.; Quinlan, E.M. Experience-dependent recovery of vision following chronic deprivation amblyopia. Nat. Neurosci. 2007, 10, 1134–1136. [Google Scholar] [CrossRef]
- Montey, K.L.; Quinlan, E.M. Recovery from chronic monocular deprivation following reactivation of thalamocortical plasticity by dark exposure. Nat. Commun. 2011, 2, 317. [Google Scholar] [CrossRef] [Green Version]
- Eaton, N.C.; Sheehan, H.M.; Quinlan, E.M. Optimization of visual training for full recovery from severe amblyopia in adults. Learn. Mem. 2016, 23, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, E.J.; Moon, L.D.F.; Popat, R.J.; King, V.R.; Bennett, G.S.; Patel, P.N.; Fawcett, J.W.; McMahon, S.B. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature 2002, 416, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Gogolla, N.; Caroni, P.; Luethi, A.; Herry, C. Perineuronal Nets Protect Fear Memories from Erasure. Science 2009, 325, 1258–1261. Available online: http://www.sciencemag.org/cgi/doi/10.1126/science.1174146%5Cnpapers3://publication/doi/10.1126/science.1174146 (accessed on 31 December 2021). [CrossRef] [PubMed] [Green Version]
- Romberg, C.; Yang, S.; Melani, R.; Andrews, M.R.; Horner, A.E.; Spillantini, M.G.; Bussey, T.J.; Fawcett, J.W.; Pizzorusso, T.; Saksida, L.M. Depletion of Perineuronalnets enhances recognition memory and long-term depression in the perirhinal cortex. J. Neurosci. 2013, 33, 7057–7065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzorusso, T.; Medini, P.; Berardi, N.; Chierzi, S.; Fawcett, J.; Maffei, L. Reactivation of Ocular Dominance Plasticity in the Adult Visual Cortex. Science 2002, 298, 1248–1251. Available online: http://www.ncbi.nlm.nih.gov/pubmed/12424383 (accessed on 31 December 2021). [CrossRef] [PubMed] [Green Version]
- Pizzorusso, T.; Medini, P.; Landi, S.; Baldini, S.; Berardi, N.; Maffei, L. Structural and functional recovery from early monocular deprivation in adult rats. Proc. Natl. Acad. Sci. USA 2006, 103, 8517–8522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frischknecht, R.; Heine, M.; Perrais, D.; Seidenbecher, C.I.; Choquet, D.; Gundelfinger, E.D. Brain extracellular matrix affects AMPA receptor lateral mobility and short-term synaptic plasticity. Nat. Neurosci. 2009, 12, 897–904. [Google Scholar] [CrossRef]
- Murase, S.; Lantz, C.L.; Quinlan, E.M. Light reintroduction after dark exposure reactivates plasticity in adults via perisynaptic activation of MMP-9. eLife 2017, 6, e27345. [Google Scholar] [CrossRef]
- Carulli, D.; Pizzorusso, T.; Kwok, J.C.F.; Putignano, E.; Poli, A.; Forostyak, S.; Andrews, M.R.; Deepa, S.S.; Glant, T.T.; Fawcett, J.W.; et al. Animals lacking link protein have attenuated Perineuronal nets and persistent plasticity. Brain 2010, 133, 2331–2347. Available online: https://academic.oup.com/brain/article/133/8/2331/386728 (accessed on 31 December 2021). [CrossRef] [Green Version]
- Nagy, V.; Bozdagi, O.; Matynia, A.; Balcerzyk, M.; Okulski, P.; Dzwonek, J.; Costa, R.; Silva, A.; Kaczmarek, L.; Huntley, G.W. Matrix Metalloproteinase-9 Is Required for Hippocampal Late-Phase Long-Term Potentiation and Memory. J. Neurosci. 2006, 26, 1923–1934. Available online: https://doi.org/10.1523/jneurosci.4359-05.2006 (accessed on 31 December 2021). [CrossRef]
- Meighan, S.E.; Meighan, P.C.; Choudhury, P.; Davis, C.J.; Olson, M.L.; Zornes, P.A.; Wright, J.W.; Harding, J.W. Effects of extracellular matrix-degrading proteases matrix metalloproteinases 3 and 9 on spatial learning and synaptic plasticity. J. Neurochem. 2006, 96, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.E.; Forquer, M.R.; Cocking, D.L.; Jansen, H.T.; Harding, J.W.; Sorg, B.A. Role of matrix metalloproteinases in the acquisition and reconsolidation of cocaine-induced conditioned place preference. Learn. Mem. 2007, 14, 214–223. Available online: http://learnmem.cshlp.org/content/14/3/214.short (accessed on 31 December 2021). [CrossRef] [PubMed] [Green Version]
- Smith, A.C.; Kupchik, Y.M.; Scofield, M.D.; Gipson, C.D.; Wiggins, A.; Thomas, C.A.; Kalivas, P.W. Synaptic plasticity mediating cocaine relapse requires matrix metalloproteinases. Nat. Neurosci. 2014, 17, 1655–1657. Available online: http://www.ncbi.nlm.nih.gov/pubmed/25326689 (accessed on 31 December 2021). [CrossRef] [PubMed] [Green Version]
- Szepesi, Z.; Hosy, E.; Ruszczycki, B.; Bijata, M.; Pyskaty, M.; Bikbaev, A.; Heine, M.; Choquet, D.; Kaczmarek, L.; Wlodarczyk, J. Synaptically released matrix metalloproteinase activity in control of structural plasticity and the cell surface distribution of GluA1-AMPA receptors. PLoS ONE 2014, 9, e98274. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Bozdagi, O.; Nikitczuk, J.S.; Zhai, Z.W.; Zhou, Q.; Huntley, G.W. Extracellular proteolysis by matrix metalloproteinase-9 drives dendritic spine enlargement and long-term potentiation coordinately. Proc. Natl. Acad. Sci. USA 2008, 105, 19520–19525. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19047646%5Cnhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC2614793 (accessed on 31 December 2021). [CrossRef] [Green Version]
- Carstens, K.E.; Phillips, M.L.; Pozzo-Miller, L.; Weinberg, R.J.; Dudek, S.M. Perineuronal Nets Suppress Plasticity of Excitatory Synapses on CA2 Pyramidal Neurons. J. Neurosci. 2016, 36, 6312–6320. [Google Scholar] [CrossRef] [Green Version]
- Alaiyed, S.; McCann, M.; Mahajan, G.; Rajkowska, G.; Stockmeier, C.A.; Kellar, K.J.; Wu, J.Y.; Conant, K. Venlafaxine stimulates an MMP-9-dependent increase in excitatory/inhibitory balance in a stress model of depression. J. Neurosci. 2022, 40, 4418–4431. [Google Scholar] [CrossRef]
- Peixoto, R.T.; Kunz, P.A.; Kwon, H.; Mabb, A.; Sabatini, B.L.; Philpot, B.D.; Ehlers, M.D. Transsynaptic Signaling by Activity-Dependent Cleavage of Neuroligin-1. Neuron 2012, 76, 396–409. [Google Scholar] [CrossRef] [Green Version]
- Bemben, M.A.; Nguyen, T.A.; Li, Y.; Wang, T.; Nicoll, R.A.; Roche, K.W. Isoform-specific cleavage of neuroligin-3 reduces synapse strength. Mol. Psychiatry 2018, 24, 145–160. [Google Scholar] [CrossRef]
- Murase, S.; Winkowski, D.E.; Liu, J.; Kanold, P.O.; Quinlan, E.M. Homeostatic regulation of perisynaptic MMP9 activity in the amblyopic visual cortex. eLife 2019, 8, e52503. [Google Scholar] [CrossRef]
- Chen, T.-W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging Neuronal activity. Nature 2013, 499, 295–300. Available online: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3777791&tool=pmcentrez&rendertype=abstract (accessed on 31 December 2021). [CrossRef] [PubMed] [Green Version]
- Lantz, C.L.; Quinlan, E.M. High-Frequency Visual Stimulation Primes Gamma Oscillations for Visually Evoked Phase Reset and Enhances Spatial Acuity. Cereb. Cortex Commun. 2021, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Grieco, S.F.; Qiao, X.; Zheng, X.; Liu, Y.; Chen, L.; Zhang, H.; Yu, Z.; Gavornik, J.P.; Lai, C.; Gandhi, S.P.; et al. Subanesthetic Ketamine Reactivates Adult Cortical Plasticity to Restore Vision from Amblyopia. Curr. Biol. 2020, 30, 3591–3603.e8. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.F.; Komorowski, R.W.; Kaplan, E.S.; Gavornik, J.P.; Bear, M.F. Visual recognition memory, manifested as long-term habituation, requires synaptic plasticity in V1. Nat. Neurosci. 2015, 18, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Cooke, S.F.; Bear, M.F. How the mechanisms of long-term synaptic potentiation and depression serve experience-dependent plasticity in primary visual cortex. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130284. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, N.; He, Y.; Liu, Y.; Ge, L.; Zou, L.; Song, S.; Xiong, W.; Liu, X. Improved calcium sensor GCaMP-X overcomes the calcium channel perturbations induced by the calmodulin in GCaMP. Nat. Commun. 2018, 9, 1504. [Google Scholar] [CrossRef]
- Salinas, K.J.; Velez, D.X.F.; Zeitoun, J.H.; Kim, H.; Gandhi, S.P. Contralateral bias of high spatial frequency tuning and cardinal direction selectivity in mouse visual cortex. J. Neurosci. 2017, 37, 10125–10138. [Google Scholar] [CrossRef] [Green Version]
- Jaubert-Miazza, L.; Green, E.; Lo, F.S.; Bui, K.; Mills, J.; Guido, W. Structural and functional composition of the developing retinogeniculate pathway in the mouse. Vis. Neurosci. 2005, 22, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Petros, T.J.; Rebsam, A.; Mason, C.A. Retinal Axon Growth at the Optic Chiasm: To Cross or Not to Cross. Annu. Rev. Neurosci. 2008, 31, 295–315. [Google Scholar] [CrossRef] [Green Version]
- Dhande, O.S.; Huberman, A.D. Retinal ganglion cell maps in the brain: Implications for visual processing. Curr. Opin. Neurobiol. 2014, 24, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Reese, B.E.; Jeffery, G. Crossed and uncrossed visual topography in dorsal lateral geniculate nucleus of the pigmented rat. J. Neurophysiol. 1983, 49, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Reese, B.E. Hidden lamination in the dorsal lateral geniculate nucleus: The functional organization of this thalamic region in the rat. Brain Res. Rev. 1988, 13, 119–137. [Google Scholar] [CrossRef]
- Coleman, J.E.; Nahmani, M.; Gavornik, J.P.; Haslinger, R.; Heynen, A.J.; Erisir, A.; Bear, M.F. Rapid Structural Remodeling of Thalamocortical Synapses Parallels Experience-Dependent Functional Plasticity in Mouse Primary Visual Cortex. J. Neurosci. 2010, 30, 9670–9682. Available online: https://pubmed.ncbi.nlm.nih.gov/20660250/ (accessed on 25 January 2021). [CrossRef] [PubMed] [Green Version]
- Nahmani, M.; Erisir, A. VGluT2 immunochemistry identifies thalamocortical terminals in layer 4 of adult and developing visual cortex. J. Comp. Neurol. 2005, 484, 458–473. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.J.; Blundon, J.A.; Bayazitov, I.T.; Zakharenko, S.S. Connectivity patterns revealed by mapping of active inputs on dendrites of thalamorecipient Neurons in the auditory cortex. J. Neurosci. 2009, 29, 6406–6417. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Martín, A.; El-Danaf, R.N.; Osakada, F.; Sriram, B.; Dhande, O.S.; Nguyen, P.L.; Callaway, E.M.; Ghosh, A.; Huberman, A.D. A dedicated circuit links direction-selective retinal ganglion cells to the primary visual cortex. Nature 2014, 507, 358–361. [Google Scholar] [CrossRef] [Green Version]
- Kelly, E.A.; Russo, A.S.; Jackson, C.D.; Lamantia, C.E.; Majewska, A.K. Proteolytic regulation of synaptic plasticity in the mouse primary visual cortex: Analysis of matrix metalloproteinase 9 deficient mice. Front Cell Neurosci. 2015, 9, 369. Available online: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=4585116&tool=pmcentrez&rendertype=abstract (accessed on 31 December 2021). [CrossRef] [Green Version]
- Frenkel, M.Y.; Sawtell, N.B.; Diogo, A.C.M.; Yoon, B.; Neve, R.L.; Bear, M.F. Instructive Effect of Visual Experience in Mouse Visual Cortex. Neuron 2006, 51, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, E.S.; Cooke, S.F.; Komorowski, R.W.; Chubykin, A.A.; Thomazeau, A.; Khibnik, L.A.; Gavornik, J.P.; Bear, M.F. Contrasting roles for parvalbumin-expressing inhibitory Neurons in two forms of adult visual cortical plasticity. eLife 2016, 5, e11450. [Google Scholar] [CrossRef]
- Spolidoro, M.S.; Putignano, E.; Munaf, C.; Maffei, L.; Pizzorusso, T. Inhibition of matrix metalloproteinases prevents the potentiation of nondeprived-eye responses after monocular deprivation in juvenile rats. Cereb. Cortex 2012, 22, 725–734. Available online: https://pubmed.ncbi.nlm.nih.gov/21685398/ (accessed on 23 September 2020). [CrossRef]
- Salinas, K.J.; Huh, C.Y.L.; Zeitoun, J.H.; Gandhi, S.P. Functional differentiation of mouse visual cortical areas depends upon early binocular experience. J. Neurosci. 2021, 41, 1470–1488. [Google Scholar] [CrossRef]
- Montey, K.L.; Eaton, N.C.; Quinlan, E.M. Repetitive visual stimulation enhances recovery from severe amblyopia. Learn. Mem. 2013, 20, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, H.; Zhang, B.; Tao, X.; Harwerth, R.S.; Smith, E.L.; Chino, Y.M. Neuronal responses in visual area V2 (V2) of macaque monkeys with strabismic amblyopia. Cereb. Cortex 2011, 21, 2033–2045. [Google Scholar] [CrossRef] [PubMed]
- Kiorpes, L.; Kiper, D.C.; O’Keefe, L.P.; Cavanaugh, J.R.; Movshon, J.A. Neuronal correlates of amblyopia in the visual cortex of macaque monkeys with experimental strabismus and anisometropia. J. Neurosci. 1998, 18, 6411–6424. [Google Scholar] [CrossRef]
- Shooner, C.; Hallum, L.; Kumbhani, R.D.; Ziemba, C.; Garcia-Marin, V.; Kelly, J.G.; Majaj, N.J.; Movshon, J.A.; Kiorpes, L. Population representation of visual information in areas V1 and V2 of amblyopic macaques. Vis. Res. 2015, 114, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Niell, C.M.; Stryker, M.P. Highly selective receptive fields in mouse visual cortex. J. Neurosci. 2008, 28, 7520–7536. Available online: http://www.ncbi.nlm.nih.gov/pubmed/18650330%5Cnhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3040721&tool=pmcentrez&rendertype=abstract (accessed on 31 December 2021). [CrossRef] [PubMed] [Green Version]
- Ledgeway, T.; Hess, R.F. The spatial frequency and orientation selectivity of the mechanisms that extract motion-defined contours. Vision Res. 2006, 46, 568–578. [Google Scholar] [CrossRef] [Green Version]
- Jeon, B.B.; Swain, A.D.; Good, J.T.; Chase, S.M.; Kuhlman, S.J. Feature selectivity is stable in primary visual cortex across a range of spatial frequencies. Sci. Rep. 2018, 8, 15288. Available online: http://www.Nature.com/articles/s41598-018-33633-2 (accessed on 17 January 2019). [CrossRef] [Green Version]
- Scholl, B.; Pattadkal, J.J.; Priebe, N.J. Binocular disparity selectivity weakened after monocular deprivation in mouse V1. J. Neurosci. 2017, 37, 6517–6526. [Google Scholar] [CrossRef] [Green Version]
- Huh, C.Y.L.; Abdelaal, K.; Salinas, K.J.; Gu, D.; Zeitoun, J.; Figueroa Velez, D.X.; Peach, J.P.; Fowlkes, C.C.; Gandhi, S.P. Long-term Monocular Deprivation during Juvenile Critical Period Disrupts Binocular Integration in Mouse Visual Thalamus. J. Neurosci. 2020, 40, 585–604. Available online: https://www.jneurosci.org/content/40/3/585 (accessed on 26 June 2020). [CrossRef]
- Jenks, K.R.; Shepherd, J.D. Experience-Dependent Development and Maintenance of Binocular Neurons in the Mouse Visual Cortex. Cell Rep. 2020, 30, 1982–1994.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.; Tring, E.; Ringach, D.L.; Zipursky, S.L.; Trachtenberg, J.T. Vision Changes the Cellular Composition of Binocular Circuitry during the Critical Period. Neuron 2020, 108, 735–747.e6. [Google Scholar] [CrossRef]
- Gordon, J.A.; Stryker, M.P. Experience-dependent plasticity of binocular responses in the primary visual cortex of the mouse. J. Neurosci. 1996, 16, 3274–3286. [Google Scholar] [CrossRef] [PubMed]
- Dräger, U.C. Receptive fields of single cells and topography in mouse visual cortex. J. Comp. Neurol. 1975, 160, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Wagor, E.; Mangini, N.J.; Pearlman, A.L. Retinotopic organization of striate and extrastriate visual cortex in the mouse. J. Comp. Neurol. 1980, 193, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Hanover, J.L.; Huang, Z.J.; Tonegawa, S.; Stryker, M.P. Brain-derived neurotrophic factor overexpression induces precocious critical period in mouse visual cortex. J. Neurosci. 1999, 19, 1–5. [Google Scholar] [CrossRef]
- Tagawa, Y.; Kanold, P.O.; Majdan, M.; Shatz, C.J. Multiple periods of functional ocular dominance plasticity in mouse visual cortex. Nat. Neurosci. 2005, 8, 380–388. Available online: http://www.Nature.com/articles/nn1410 (accessed on 15 January 2019). [CrossRef]
- Hofer, S.B.; Mrsic-Flogel, T.D.; Bonhoeffer, T.; Hübener, M. Experience leaves a lasting structural trace in cortical circuits. Nature 2009, 457, 313–317. [Google Scholar] [CrossRef]
- Majewska, A.; Sur, M. Motility of dendritic spines in visual cortex in vivo: Changes during the critical period and effects of visual deprivation. Proc. Natl. Acad. Sci. USA 2003, 100, 16024–16029. [Google Scholar] [CrossRef] [Green Version]
- Gorlewicz, A.; Kaczmarek, L. Pathophysiology of trans-synaptic adhesion molecules: Implications for epilepsy. Front. Cell Dev. Biol. 2018, 6. Available online: https://www.frontiersin.org (accessed on 28 September 2020). [CrossRef]
- Conant, K.; Wang, Y.; Szklarczyk, A.; Dudak, A.; Mattson, M.P.; Lim, S.T. Matrix metalloproteinase-dependent shedding of intercellular adhesion molecule-5 occurs with long-term potentiation. Neuroscience 2010, 166, 508–521. Available online: https://www.sciencedirect.com/science/article/pii/S0306452209021496?via%3Dihub (accessed on 28 February 2019). [CrossRef] [PubMed] [Green Version]
- Yoshihara, Y.; Oka, S.; Nemoto, Y. An I CAM- Related Neuronal Glycoprotein, Telencephalin, with Brain Segment. Specif. Express. 1994, 12, 541–553. [Google Scholar]
- Matsuno, H.; Okabe, S.; Mishina, M.; Yanagida, T.; Mori, K.; Yoshihara, Y. Development/Plasticity/Repair Telencephalin Slows Spine Maturation. J. Neurosci. 2006, 26, 1776–1786. Available online: https://www.jneurosci.org/content/26/6/1776.short (accessed on 3 April 2019). [CrossRef] [PubMed] [Green Version]
- Kelly, E.A.; Tremblay, M.E.; Gahmberg, C.G.; Tian, L.; Majewska, A.K. Subcellular localization of intercellular adhesion molecule-5 (telencephalin) in the visual cortex is not developmentally regulated in the absence of matrix metalloproteinase-9. J. Comp. Neurol. 2014, 522, 676–688. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Stefanidakis, M.; Ning, L.; Van Lint, P.; Nyman-Huttunen, H.; Libert, C.; Itohara, S.; Mishina, M.; Rauvala, H.; Gahmberg, C.G. Activation of NMDA receptors promotes dendritic spine development through MMP-mediated ICAM-5 cleavage. J. Cell Biol. 2007, 178, 687–700. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, H.; Dansie, L.E.; Hickmott, P.W.; Ethell, D.W.; Ethell, I.M. Genetic removal of matrix metalloproteinase 9 rescues the symptoms of fragile X syndrome in a mouse model. J. Neurosci. 2014, 34, 9867–9879. Available online: http://www.ncbi.nlm.nih.gov/pubmed/25057190%5Cnhttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC4107404 (accessed on 31 December 2021). [CrossRef] [Green Version]
- Janusz, A.; Milek, J.; Perycz, M.; Pacini, L.; Bagni, C.; Kaczmarek, L.; Dziembowska, M. The Fragile X mental retardation protein regulates matrix metalloproteinase 9 mRNA at synapses. J. Neurosci. 2013, 33, 18234–18241. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24227732 (accessed on 31 December 2021). [CrossRef] [Green Version]
- Mercuri, F.A.; Maciewicz, R.A.; Tart, J.; Last, K.; Fosang, A.J. Mutations in the interglobular domain of aggrecan alter matrix metalloproteinase and aggrecanase cleavage patterns: Evidence that matrix metalloproteinase cleavage interferes with aggrecanase activity. J. Biol. Chem. 2000, 275, 33038–33045. [Google Scholar] [CrossRef] [Green Version]
- Bednarek, N.; Clément, Y.; Lelièvre, V.; Olivier, P.; Loron, G.; Garnotel, R.; Gressens, P. Ontogeny of MMPs and TIMPs in the murine neocortex. Pediatr. Res. 2009, 65, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Aujla, P.K.; Huntley, G.W. Early postnatal expression and localization of matrix metalloproteinases-2 and -9 during establishment of rat hippocampal synaptic circuitry. J. Comp. Neurol. 2014, 522, 1249–1263. [Google Scholar] [CrossRef] [Green Version]
- Demas, J.; Sagdullaev, B.T.; Green, E.; Jaubert-Miazza, L.; McCall, M.A.; Gregg, R.G.; Wong, R.O.L.; Guido, W. Failure to Maintain Eye-Specific Segregation in nob, a Mutant with Abnormally Patterned Retinal Activity. Neuron 2006, 50, 247–259. Available online: https://www.sciencedirect.com/science/article/pii/S0896627306002303 (accessed on 26 February 2019). [CrossRef] [PubMed] [Green Version]
- Hayakawa, I.; Kawasaki, H. Rearrangement of Retinogeniculate Projection Patterns after Eye-Specific Segregation in Mice. PLoS ONE 2010, 5, e11001. [Google Scholar] [CrossRef]
- Kang, E.; Durand, S.; LeBlanc, J.J.; Hensch, T.K.; Chen, C.; Fagiolini, M. Visual acuity development and plasticity in the absence of sensory experience. J. Neurosci. 2013, 33, 17789–17796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephany, C.; Chan, L.L.; Parivash, S.N.; Dorton, H.M.; Piechowicz, M.; Qiu, S.; McGee, A.W. Plasticity of Binocularity and Visual Acuity Are Differentially Limited by Nogo Receptor. J. Neurosci. 2014, 34, 11631–11640. [Google Scholar] [CrossRef] [Green Version]
- Stephany, C.É.; Ma, X.; Dorton, H.M.; Wu, J.; Solomon, A.M.; Frantz, M.G.; Qiu, S.; McGee, A.W. Distinct Circuits for Recovery of Eye Dominance and Acuity in Murine Amblyopia. Curr. Biol. 2018, 28, 1914–1923.e5. [Google Scholar] [CrossRef] [PubMed]
- Durkin, J.; Suresh, A.K.; Colbath, J.; Broussard, C.; Wu, J.; Zochowski, M.; Aton, S.J. Cortically coordinated NREM thalamocortical oscillations play an essential, instructive role in visual system plasticity. Proc. Natl. Acad. Sci. USA 2017, 114, 10485–10490. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28893999 (accessed on 3 April 2019). [CrossRef] [PubMed] [Green Version]
- Thomas, A.C.; Newby, A.C. Effect of matrix metalloproteinase-9 knockout on vein graft remodelling in mice. J. Vasc. Res. 2010, 47, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, M.; Stellwagen, D.; Malenka, R.C.; Stryker, M.P. Tumor Necrosis Factor-α Mediates One Component of Competitive, Experience-Dependent Plasticity in Developing Visual Cortex. Neuron 2008, 58, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Kauppinen, T.M.; Swanson, R.A. Poly(ADP-Ribose) Polymerase-1 Promotes Microglial Activation, Proliferation, and Matrix Metalloproteinase-9-Mediated Neuron Death. J. Immunol. 2005, 174, 2288–2296. [Google Scholar] [CrossRef] [Green Version]
- Pielecka-Fortuna, J.; Kalogeraki, E.; Fortuna, M.G.; Löwel, S. Optimal level activity of matrix metalloproteinases is critical for adult visual plasticity in the healthy and stroke-affected brain. eLife 2015, 4, e11290. [Google Scholar] [CrossRef]
- Seabrook, T.A.; El-Danaf, R.N.; Krahe, T.E.; Fox, M.A.; Guido, W. Retinal Input Regulates the Timing of Corticogeniculate Innervation. J. Neurosci. 2013, 33, 10085–10097. Available online: http://www.ncbi.nlm.nih.gov/pubmed/23761904 (accessed on 16 January 2019). [CrossRef] [PubMed] [Green Version]
- Goldey, G.J.; Roumis, D.K.; Glickfeld, L.L.; Kerlin, A.M.; Reid, R.C.; Bonin, V.; Schafer, D.P.; Andermann, M.L. Removable cranial windows for long-term imaging in awake mice. Nat. Protoc. 2014, 9, 2515–2538. [Google Scholar] [CrossRef] [PubMed]
- Pachitariu, M.; Stringer, C.; Dipoppa, M.; Schröder, S.; Rossi, L.F.; Dalgleish, H.; Carandini, M.; Harris, K. Suite2p: Beyond 10,000 Neurons with standard two-photon microscopy. bioRxiv 2016. bioRxiv:061507. [Google Scholar]
- Mazurek, M.; Kager, M.; Van Hooser, S.D. Robust quantification of orientation selectivity and direction selectivity. Front Neural. Circ. 2014, 8, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brainard, D.H. The Psychophysics Toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelli, D.G. The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spat. Vis. 1997, 10, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murase, S.; Robertson, S.E.; Lantz, C.L.; Liu, J.; Winkowski, D.E.; Quinlan, E.M. Chronic Monocular Deprivation Reveals MMP9-Dependent and -Independent Aspects of Murine Visual System Plasticity. Int. J. Mol. Sci. 2022, 23, 2438. https://doi.org/10.3390/ijms23052438
Murase S, Robertson SE, Lantz CL, Liu J, Winkowski DE, Quinlan EM. Chronic Monocular Deprivation Reveals MMP9-Dependent and -Independent Aspects of Murine Visual System Plasticity. International Journal of Molecular Sciences. 2022; 23(5):2438. https://doi.org/10.3390/ijms23052438
Chicago/Turabian StyleMurase, Sachiko, Sarah E. Robertson, Crystal L. Lantz, Ji Liu, Daniel E. Winkowski, and Elizabeth M. Quinlan. 2022. "Chronic Monocular Deprivation Reveals MMP9-Dependent and -Independent Aspects of Murine Visual System Plasticity" International Journal of Molecular Sciences 23, no. 5: 2438. https://doi.org/10.3390/ijms23052438