
Genome-Wide Identification and Expression Analysis of Heat Shock Protein 70 (HSP70) Gene Family in Pumpkin (Cucurbita moschata) Rootstock under Drought Stress Suggested the Potential Role of these Chaperones in Stress Tolerance


Abstract
:1. Introduction
2. Results
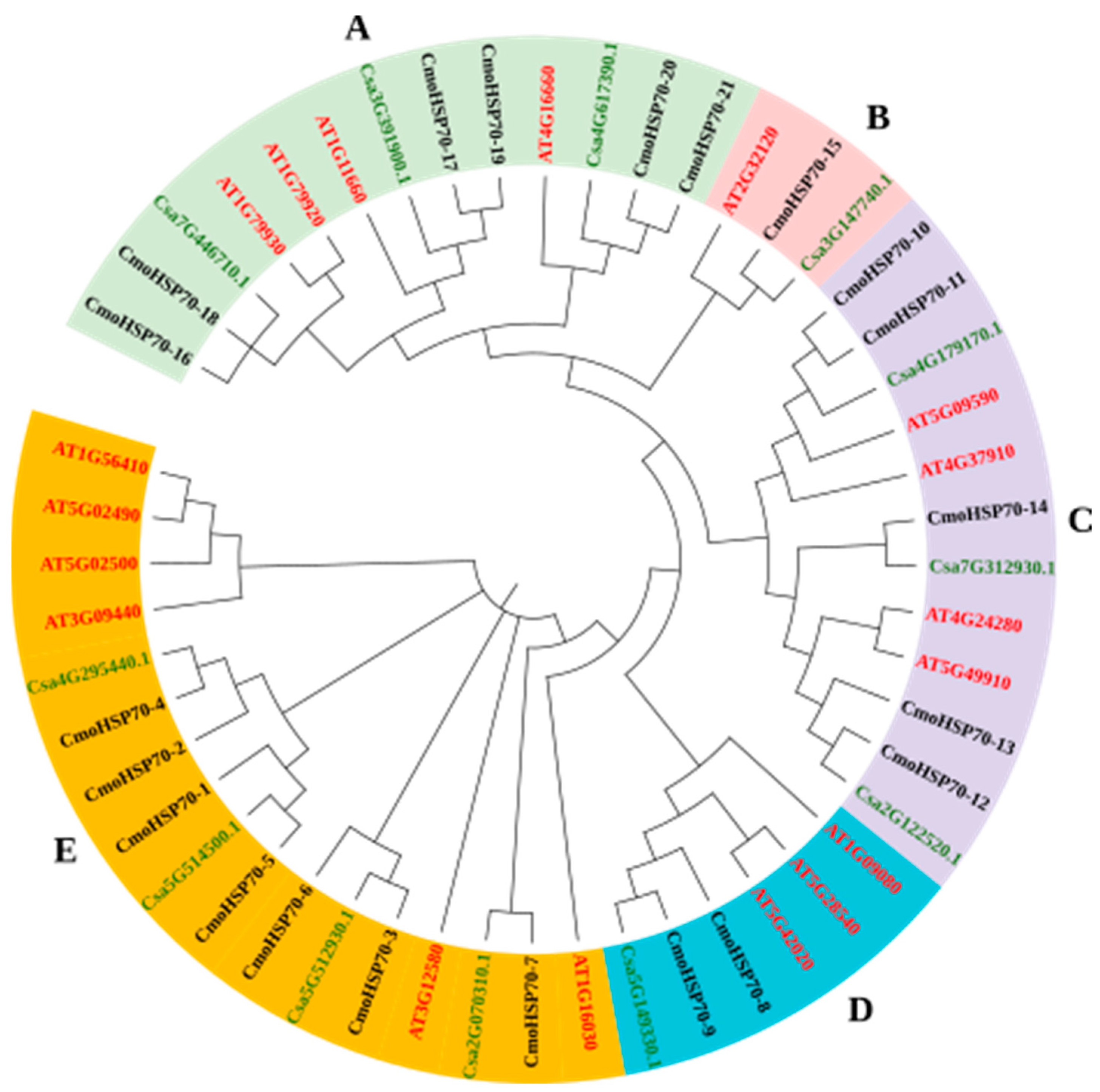
2.1. Identification of HSP70 Gene Family Members in Cucurbita moschata
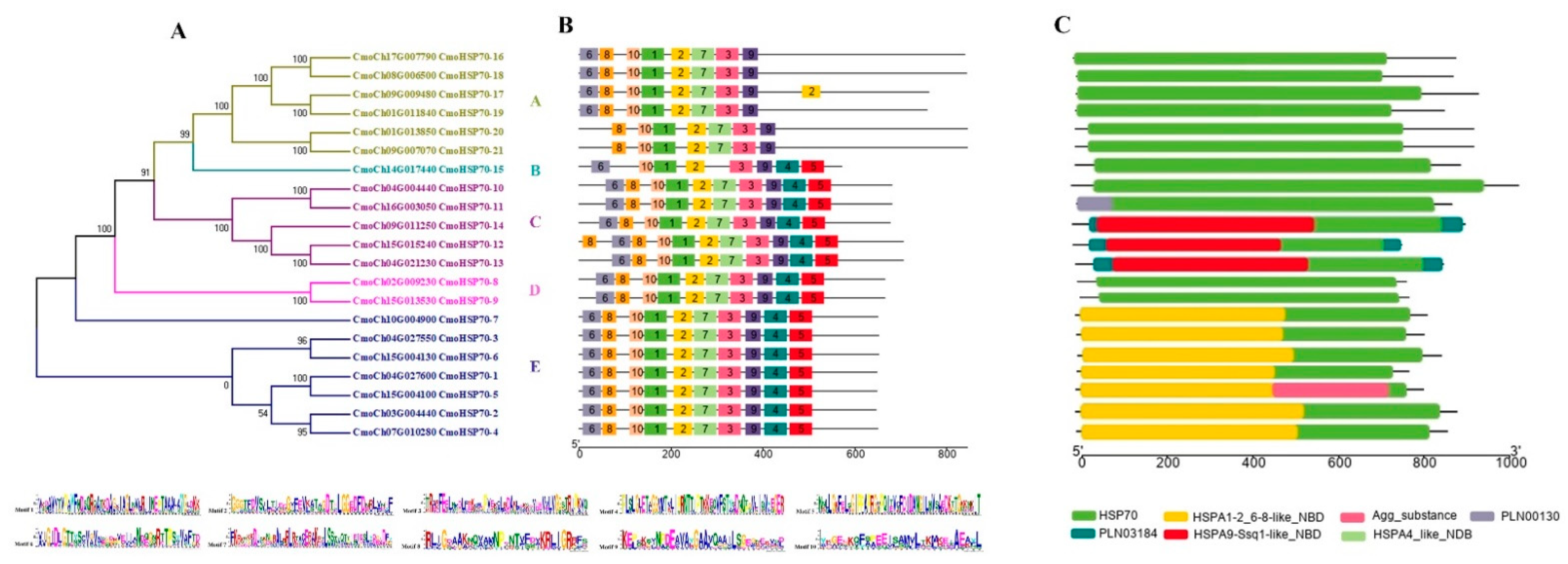
2.2. Evolutionary Relationship, Motif, Gene Structure, and Domain Analysis
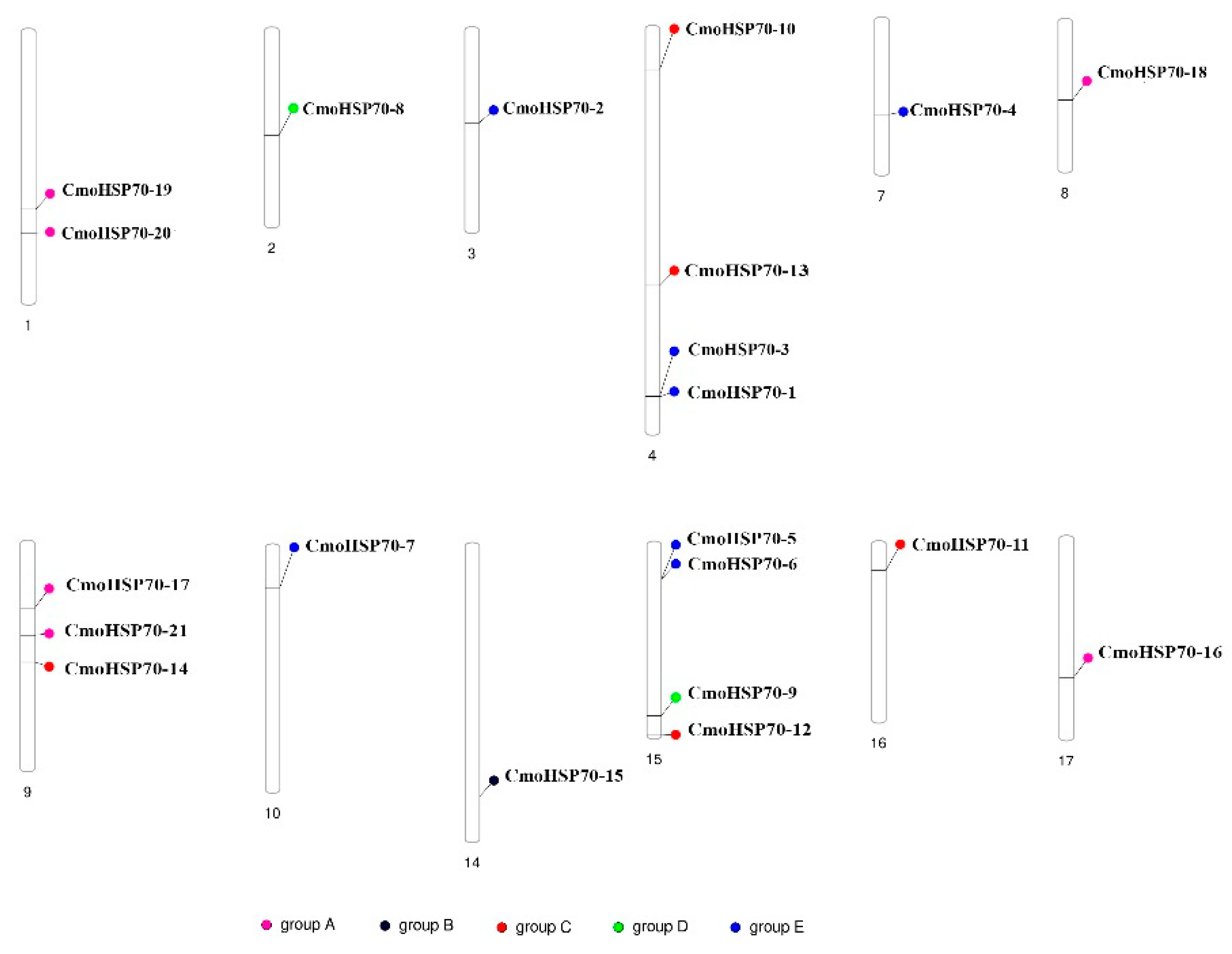
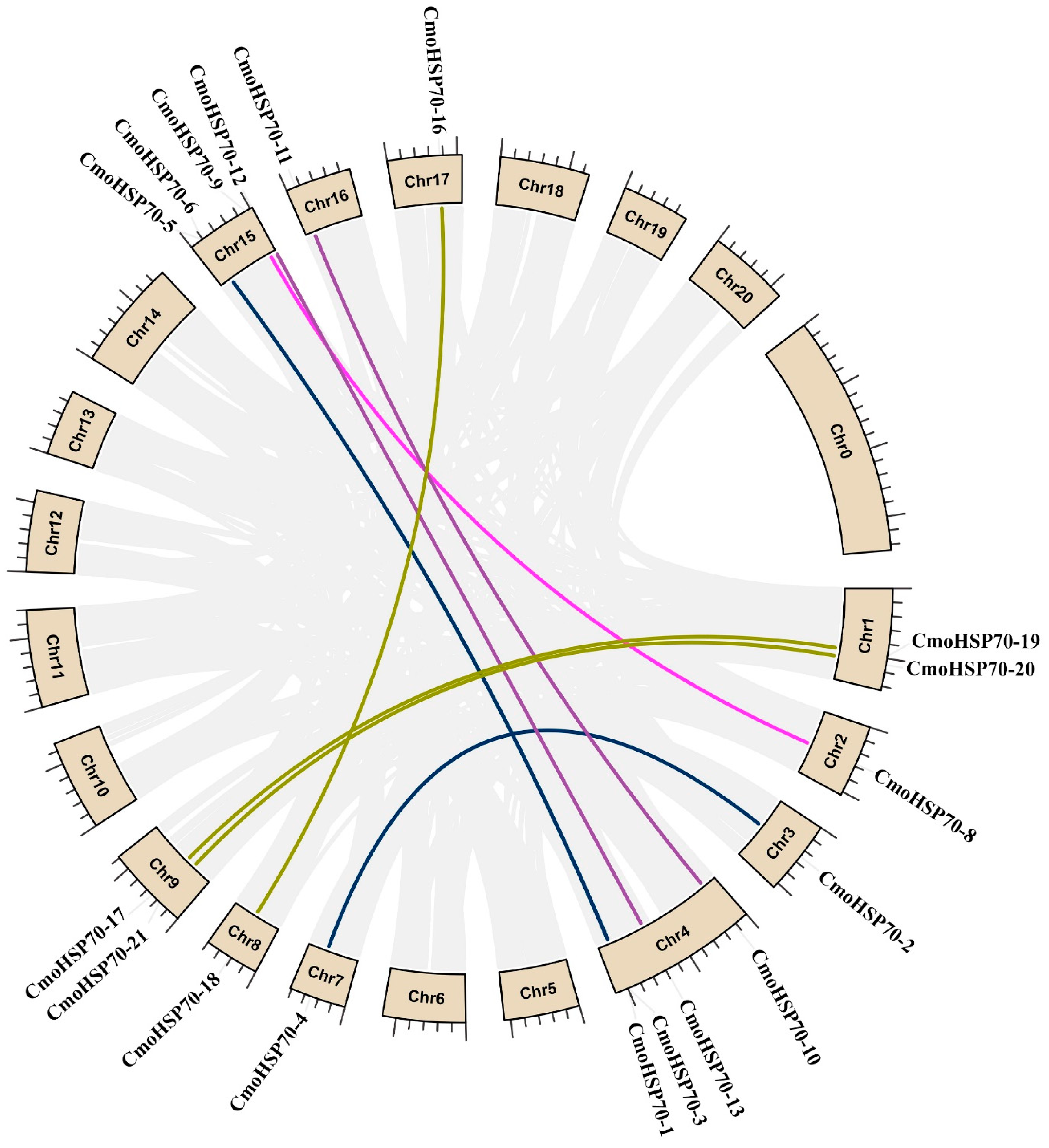
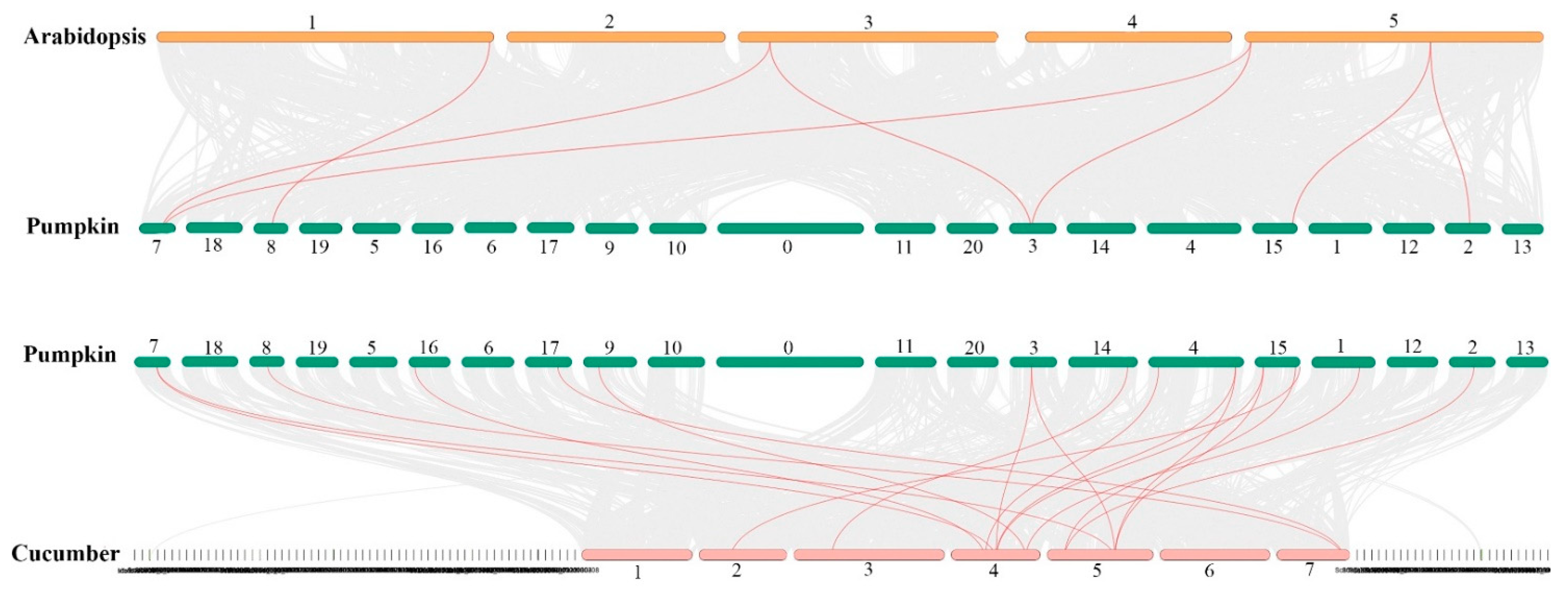
2.3. Chromosomal Locations and Gene Duplication of the CmoHsp70 Genes in Pumpkin
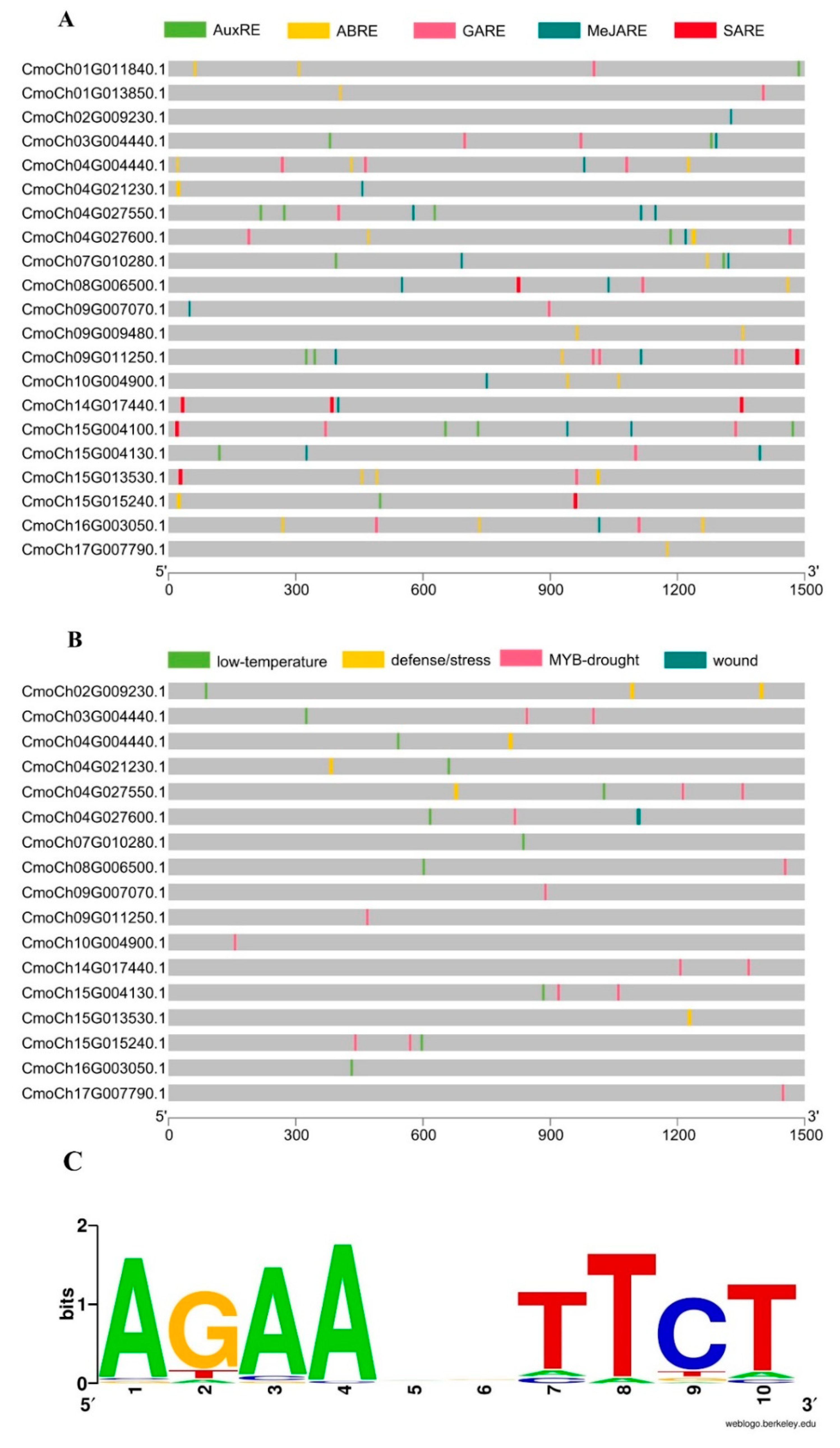
2.4. Promoter Region Analysis and Cis-acting Elements of the CmoHSP70 Genes
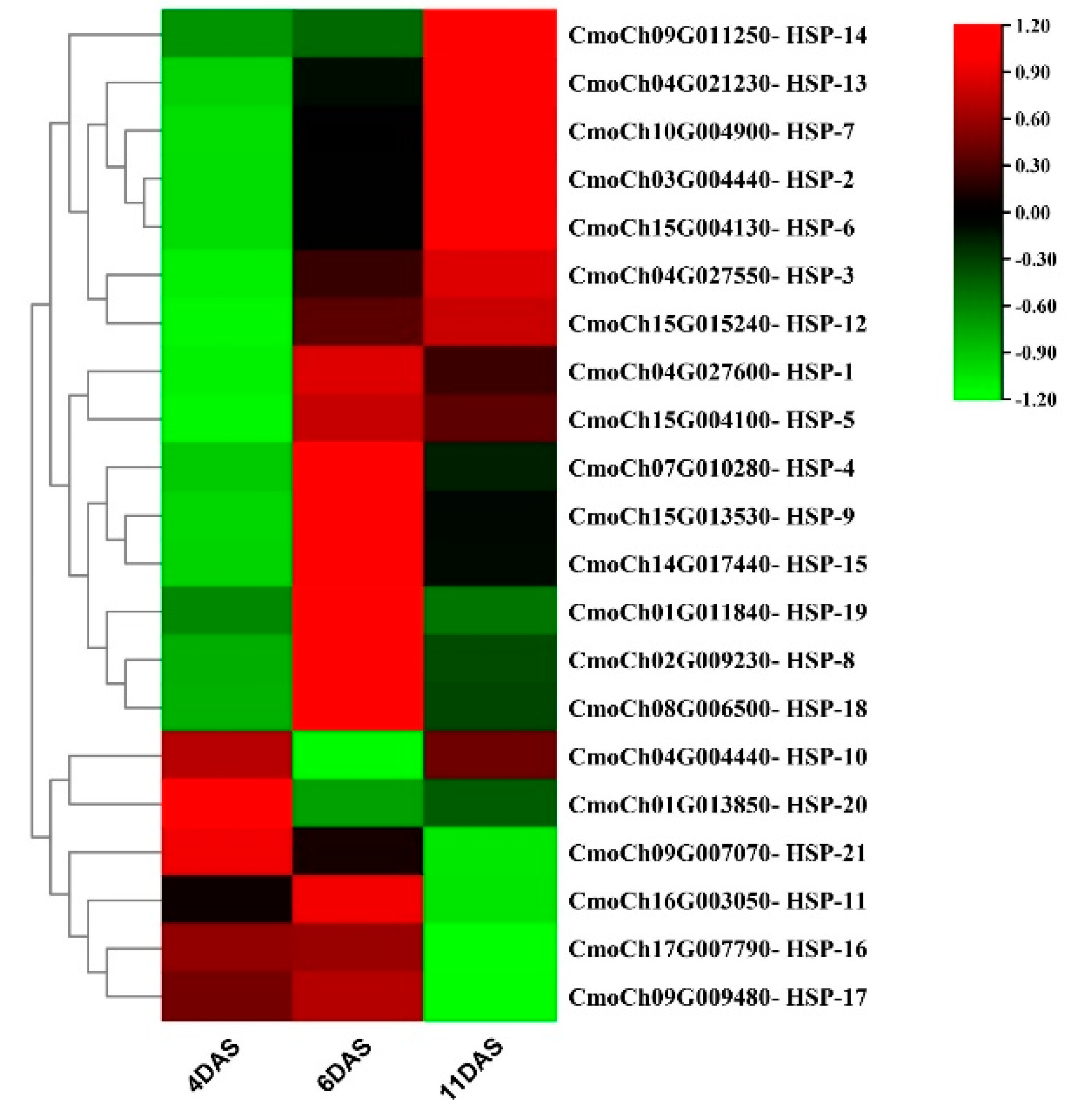
2.5. Expression Pattern of HSP70s in Response to Drought Stress
3. Discussion
4. Materials and Methods
4.1. CmoHSP70 Sequences Extraction from Cucurbit Database and Collection of their Physicochemical Properties
4.2. Construction of Phylogenetic Tree
4.3. Analysis of Gene Structures, Motifs, and Conserved Domains
4.4. Chromosomal Location, Gene Duplication, and Synteny Analysis
4.5. Promoter Analysis of the Identified HSP70 Genes of C. moschata
4.6. The Analysis of Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peck, S.; Mittler, R. Plant signaling in biotic and abiotic stress. J. Exp. Bot. 2020, 71, 1649–1651. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.Y.; Bassham, D.C. Combating stress: The interplay between hormone signaling and autophagy in plants. J. Exp. Bot. 2020, 71, 1723–1733. [Google Scholar] [CrossRef]
- Sterck, L.; Rombauts, S.; Vandepoele, K.; Rouzé, P.; Van de Peer, Y. How many genes are there in plants (... and why are they there)? Curr. Opin. Plant Biol. 2007, 10, 199–203. [Google Scholar] [CrossRef]
- Schlesinger, M.J. Heat shock proteins. J. Biol. Chem. 1990, 265, 12111–12114. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Lu, J.P.; Zhai, Y.F.; Wang, H.; Gong, Z.H.; Wang, S.B.; Lu, M.H. Genome-wide analysis of the CaHsp20 gene family in pepper: Comprehensive sequence and expression profile analysis under heat stress. Front. Plant Sci. 2015, 6, 806. [Google Scholar] [CrossRef] [Green Version]
- Cazale, A.C.; Clement, M.; Chiarenza, S.; Roncato, M.A.; Pochon, N.; Creff, A.; Marin, E.; Leonhardt, N.; Noel, L.D. Altered expression of cytosolic/nuclear HSC70-1 molecular chaperone affects development and abiotic stress tolerance in Arabidopsis thaliana. J. Exp. Bot. 2009, 60, 2653–2664. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Gong, W.; Zhang, H.; Perrett, S.; Jones, G.W. The same but different: The role of Hsp70 in heat shock response and prion propagation. Prion 2018, 12, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.X.; Zhang, H.X.; Wei, A.M.; Gong, Z.H. Heat Shock Proteins: Dynamic Biomolecules to Counter Plant Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef] [Green Version]
- Rehman, A.; Atif, R.M.; Qayyum, A.; Du, X.; Hinze, L.; Azhar, M.T. Genome-wide identification and characterization of HSP70 gene family in four species of cotton. Genomics 2020, 112, 4442–4453. [Google Scholar] [CrossRef]
- Song, Z.; Pan, F.; Lou, X.; Wang, D.; Yang, C.; Zhang, B.; Zhang, H. Genome-wide identification and characterization of Hsp70 gene family in Nicotiana tabacum. Mol. Biol. Rep. 2019, 46, 1941–1954. [Google Scholar] [CrossRef]
- Truman, A.W.; Bourboulia, D.; Mollapour, M. Decrypting the chaperone code. J. Biol. Chem. 2021, 296, 100293. [Google Scholar] [CrossRef]
- Backe, S.J.; Sager, R.A.; Woodford, M.R.; Makedon, A.M.; Mollapour, M. Post-translational modifications of Hsp90 and translating the chaperone code. J. Biol. Chem. 2020, 295, 11099–11117. [Google Scholar] [CrossRef]
- Lim, S.; Kim, D.G.; Kim, S. ERK-dependent phosphorylation of the linker and substrate-binding domain of HSP70 increases folding activity and cell proliferation. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Beltrao, P.; Albanese, V.; Kenner, L.R.; Swaney, D.L.; Burlingame, A.; Villen, J.; Lim, W.A.; Fraser, J.S.; Frydman, J.; Krogan, N.J. Systematic functional prioritization of protein posttranslational modifications. Cell 2012, 150, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Nitika; Porter, C.M.; Truman, A.W.; Truttmann, M.C. Post-translational modifications of Hsp70 family proteins: Expanding the chaperone code. J. Biol. Chem. 2020, 295, 10689–10708. [Google Scholar] [CrossRef]
- Sung, D.Y.; Vierling, E.; Guy, C.L. Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family. Plant Physiol. 2001, 126, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.H.; Gho, H.J.; Nguyen, M.X.; Kim, S.R.; An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genom. 2013, 13, 391–402. [Google Scholar] [CrossRef]
- Jungkunz, I.; Link, K.; Vogel, F.; Voll, L.M.; Sonnewald, S.; Sonnewald, U. AtHsp70-15-deficient Arabidopsis plants are characterized by reduced growth, a constitutive cytosolic protein response and enhanced resistance to TuMV. Plant J. 2011, 66, 983–995. [Google Scholar] [CrossRef]
- Chen, T.; Cao, X. Stress for maintaining memory: HSP70 as a mobile messenger for innate and adaptive immunity. Eur. J. Immunol. 2010, 40, 1541–1544. [Google Scholar] [CrossRef]
- Li, H.; Liu, S.S.; Yi, C.Y.; Wang, F.; Zhou, J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Hydrogen peroxide mediates abscisic acid-induced HSP70 accumulation and heat tolerance in grafted cucumber plants. Plant Cell Environ. 2014, 37, 2768–2780. [Google Scholar] [CrossRef]
- Liu, J.; Pang, X.; Cheng, Y.; Yin, Y.; Zhang, Q.; Su, W.; Hu, B.; Guo, Q.; Ha, S.; Zhang, J.; et al. The Hsp70 Gene Family in Solanum tuberosum: Genome-Wide Identification, Phylogeny, and Expression Patterns. Sci. Rep. 2018, 8, 16628. [Google Scholar] [CrossRef]
- Cho, E.K.; Choi, Y.J. A nuclear-localized HSP70 confers thermoprotective activity and drought-stress tolerance on plants. Biotechnol. Lett. 2009, 31, 597–606. [Google Scholar] [CrossRef]
- Kallamadi, P.R.; Dandu, K.; Kirti, P.B.; Rao, C.M.; Thakur, S.S.; Mulpuri, S. An Insight into Powdery Mildew–Infected, Susceptible, Resistant, and Immune Sunflower Genotypes. Proteomics 2018, 18, 1700418. [Google Scholar] [CrossRef]
- Devarajan, A.K.; Muthukrishanan, G.; Truu, J.; Truu, M.; Ostonen, I.; Kizhaeral, S.S.; Panneerselvam, P.; Kuttalingam Gopalasubramanian, S. The Foliar Application of Rice Phyllosphere Bacteria induces Drought-Stress Tolerance in Oryza sativa (L.). Plants 2021, 10, 387. [Google Scholar] [CrossRef]
- Pulido, P.; Llamas, E.; Rodriguez-Concepcion, M. Both Hsp70 chaperone and Clp protease plastidial systems are required for protection against oxidative stress. Plant Signal. Behav. 2017, 12, e1290039. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.K.; Hong, C.B. Over-expression of tobacco NtHSP70-1 contributes to drought-stress tolerance in plants. Plant Cell Rep. 2006, 25, 349–358. [Google Scholar] [CrossRef]
- Augustine, S.M.; Cherian, A.V.; Syamaladevi, D.P.; Subramonian, N. Erianthus arundinaceus HSP70 (EaHSP70) Acts as a Key Regulator in the Formation of Anisotropic Interdigitation in Sugarcane (Saccharum spp. hybrid) in Response to Drought Stress. Plant Cell Physiol. 2015, 56, 2368–2380. [Google Scholar] [CrossRef] [Green Version]
- Song, A.; Zhu, X.; Chen, F.; Gao, H.; Jiang, J.; Chen, S. A chrysanthemum heat shock protein confers tolerance to abiotic stress. Int. J. Mol. Sci. 2014, 15, 5063–5078. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Liu, J.H.; Ma, X.; Zhai, Y.F.; Gong, Z.H.; Lu, M.H. Genome-wide analysis of the Hsp70 family genes in pepper (Capsicum annuum L.) and functional identification of CaHsp70-2 involvement in heat stress. Plant Sci. 2016, 252, 246–256. [Google Scholar] [CrossRef]
- Hu, X.; Liu, R.; Li, Y.; Wang, W.; Tai, F.; Xue, R.; Li, C. Heat shock protein 70 regulates the abscisic acid-induced antioxidant response of maize to combined drought and heat stress. Plant Growth Regul. 2010, 60, 225–235. [Google Scholar] [CrossRef]
- Baloji, G.; Pasham, S.; Mahankali, V.; Garladinne, M.; Ankanagari, S. Insights from the molecular docking analysis of phytohormone reveal brassinolide interaction with HSC70 from Pennisetum glaucum. Bioinformation 2019, 15, 131–138. [Google Scholar] [CrossRef]
- Zhao, D.; Xia, X.; Su, J.; Wei, M.; Wu, Y.; Tao, J. Overexpression of herbaceous peony HSP70 confers high temperature tolerance. BMC Genom. 2019, 20, 70. [Google Scholar] [CrossRef]
- Masand, S.; Yadav, S.K. Overexpression of MuHSP70 gene from Macrotyloma uniflorum confers multiple abiotic stress tolerance in transgenic Arabidopsis thaliana. Mol. Biol. Rep. 2016, 43, 53–64. [Google Scholar] [CrossRef]
- Mokhtar, M.; Bouamar, S.; Di Lorenzo, A.; Temporini, C.; Daglia, M.; Riazi, A. The Influence of Ripeness on the Phenolic Content, Antioxidant and Antimicrobial Activities of Pumpkins (Cucurbita moschata Duchesne). Molecules 2021, 26, 3623. [Google Scholar] [CrossRef]
- Vinayashree, S.; Vasu, P. Biochemical, nutritional and functional properties of protein isolate and fractions from pumpkin (Cucurbita moschata var. Kashi Harit) seeds. Food Chem. 2021, 340, 128177. [Google Scholar] [CrossRef]
- Shen, C.; Yuan, J. Genome-wide characterization and expression analysis of the heat shock transcription factor family in pumpkin (Cucurbita moschata). BMC Plant Biol. 2020, 20, 471. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, J.; Yang, Y.; Luo, J.; Zheng, X.; Wen, C.; Xu, Y. Transcription factor CsWIN1 regulates pericarp wax biosynthesis in cucumber grafted on pumpkin. Front. Plant Sci. 2019, 10, 1564. [Google Scholar] [CrossRef]
- Li, L.; Shu, S.; Xu, Q.; An, Y.H.; Sun, J.; Guo, S.R. NO accumulation alleviates H2O2-dependent oxidative damage induced by Ca(NO3)2 stress in the leaves of pumpkin-grafted cucumber seedlings. Physiol. Plant 2017, 160, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Li, H.; Lv, X.; Ahammed, G.J.; Xia, X.; Zhou, J.; Shi, K.; Asami, T.; Yu, J.; Zhou, Y. Grafting cucumber onto luffa improves drought tolerance by increasing ABA biosynthesis and sensitivity. Sci. Rep. 2016, 6, 20212. [Google Scholar] [CrossRef] [Green Version]
- Davoudi, M.; Song, M.; Zhang, M.; Chen, J.; Lou, Q. Long-distance control of pumpkin rootstock over cucumber scion under drought stress as revealed by transcriptome sequencing and mobile mRNAs identifications. Hortic. Res. 2022. [Google Scholar] [CrossRef]
- Lin, B.L.; Wang, J.S.; Liu, H.C.; Chen, R.W.; Meyer, Y.; Barakat, A.; Delseny, M. Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 201–208. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, H.K.; Dong, Q.L.; Zhang, Y.Y.; Wang, Y.M.; Li, H.Y.; Xing, G.J.; Li, Q.Y.; Dong, Y.S. Genome-wide analysis and expression profiling under heat and drought treatments of HSP70 gene family in soybean (Glycine max L.). Front. Plant Sci. 2015, 6, 773. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Xing, M.; Liu, X.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Lv, H. Genome-wide analysis of HSP70 family genes in cabbage (Brassica oleracea var. capitata) reveals their involvement in floral development. BMC Genom. 2019, 20, 369. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Hu, W.; Qian, Y.; Ren, Q.; Zhang, J. Genome-wide identification, classification and expression analysis of the Hsf and Hsp70 gene families in maize. Gene 2021, 770, 145348. [Google Scholar] [CrossRef]
- Wang, X.R.; Wang, C.; Ban, F.X.; Zhu, D.T.; Liu, S.S.; Wang, X.W. Genome-wide identification and characterization of HSP gene superfamily in whitefly (Bemisia tabaci) and expression profiling analysis under temperature stress. Insect Sci. 2019, 26, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Semon, M.; Wolfe, K.H. Consequences of genome duplication. Curr. Opin. Genet. Dev. 2007, 17, 505–512. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Schlueter, J.A.; Lin, J.Y.; Schlueter, S.D.; Vasylenko-Sanders, I.F.; Deshpande, S.; Yi, J.; O’Bleness, M.; Roe, B.A.; Nelson, R.T.; Scheffler, B.E.; et al. Gene duplication and paleopolyploidy in soybean and the implications for whole genome sequencing. BMC Genom. 2007, 8, 330. [Google Scholar] [CrossRef] [Green Version]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-Regulating Factors (GRFs): A Small Transcription Factor Family with Important Functions in Plant Biology. Mol. Plant 2015, 8, 998–1010. [Google Scholar] [CrossRef] [Green Version]
- Cui, F.; Taier, G.; Wang, X.; Wang, K. Genome-Wide Analysis of the HSP20 Gene Family and Expression Patterns of HSP20 Genes in Response to Abiotic Stresses in Cynodon transvaalensis. Front. Genet. 2021, 12, 732812. [Google Scholar] [CrossRef]
- Chen, H.Y.; Hsieh, E.J.; Cheng, M.C.; Chen, C.Y.; Hwang, S.Y.; Lin, T.P. ORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element. New Phytol. 2016, 211, 599–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Song, Z.; Li, F.; Li, X.; Ji, H.; Yang, S. The specific MYB binding sites bound by TaMYB in the GAPCp2/3 promoters are involved in the drought stress response in wheat. BMC Plant Biol. 2019, 19, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef]
- Zhao, P.; Javed, S.; Shi, X.; Wu, B.; Zhang, D.; Xu, S.; Wang, X. Varying Architecture of Heat Shock Elements Contributes to Distinct Magnitudes of Target Gene Expression and Diverged Biological Pathways in Heat Stress Response of Bread Wheat. Front. Genet. 2020, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skaggs, H.S.; Xing, H.; Wilkerson, D.C.; Murphy, L.A.; Hong, Y.; Mayhew, C.N.; Sarge, K.D. HSF1-TPR interaction facilitates export of stress-induced HSP70 mRNA. J. Biol. Chem. 2007, 282, 33902–33907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, A.; Trotta, E.; Brandi, R.; Arisi, I.; Coccia, M.; Santoro, M.G. AIRAP, a new human heat shock gene regulated by heat shock factor 1. J. Biol. Chem. 2010, 285, 13607–13615. [Google Scholar] [CrossRef] [Green Version]
- Peffer, S.; Goncalves, D.; Morano, K.A. Regulation of the Hsf1-dependent transcriptome via conserved bipartite contacts with Hsp70 promotes survival in yeast. J. Biol. Chem. 2019, 294, 12191–12202. [Google Scholar] [CrossRef]
- Kim, B.H.; Schoffl, F. Interaction between Arabidopsis heat shock transcription factor 1 and 70 kDa heat shock proteins. J. Exp. Bot 2002, 53, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Baler, R.; Zou, J.; Voellmy, R. Evidence for a role of Hsp70 in the regulation of the heat shock response in mammalian cells. Cell Stress Chaperones 1996, 1, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Jan, A.; Ohama, N.; Kidokoro, S.; Soma, F.; Koizumi, S.; Mogami, J.; Todaka, D.; Mizoi, J.; Shinozaki, K.; et al. Cytosolic HSC70s repress heat stress tolerance and enhance seed germination under salt stress conditions. Plant Cell Environ. 2021, 44, 1788–1801. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript ID | Gene Name | Chr. | Location Start-End | CDS (bp) | Protein Length (A.A) | Protein Molecular Weight (kDa) | pI | GRAVY | NO. Intron/Exon | Subcellular Localization ‘Cello Life‘ |
|---|---|---|---|---|---|---|---|---|---|---|
| CmoCh04G027600.1 | CmoHSP70-1 | 4 | 19963206-19966159 | 1947 | 648 | 71.122 | 5.17 | −0.417 | 1:2 | Cytoplasmic |
| CmoCh03G004440.1 | CmoHSP70-2 | 3 | 5028811-5031809 | 1944 | 647 | 70.782 | 5.13 | −0.384 | 1:2 | Cytoplasmic |
| CmoCh04G027550.1 | CmoHSP70-3 | 4 | 19938575-19941497 | 1959 | 652 | 71.435 | 5.1 | −0.434 | 1:2 | Cytoplasmic |
| CmoCh07G010280.1 | CmoHSP70-4 | 7 | 5088728-5092059 | 1953 | 650 | 71.226 | 5.16 | −0.402 | 1:2 | Cytoplasmic |
| CmoCh15G004100.1 | CmoHSP70-5 | 15 | 1867166-1869729 | 1950 | 649 | 71.019 | 5.16 | −0.406 | 1:2 | Cytoplasmic |
| CmoCh15G004130.1 | CmoHSP70-6 | 15 | 1874588-1877079 | 1959 | 652 | 71.619 | 5.11 | −0.407 | 1:2 | Cytoplasmic |
| CmoCh10G004900.1 | CmoHSP70-7 | 10 | 2190331-2193020 | 2034 | 677 | 71.26 | 5.15 | −0.393 | 2:3 | Cytoplasmic |
| CmoCh02G009230.1 | CmoHSP70-8 | 2 | 5674154-5678674 | 2001 | 666 | 73.445 | 5.07 | −0.451 | 7:8 | E.R. |
| CmoCh15G013530.1 | CmoHSP70-9 | 15 | 9263256-9266895 | 1998 | 665 | 73.408 | 5.13 | −0.463 | 6:7 | E.R. |
| CmoCh04G004440.1 | CmoHSP70-10 | 4 | 2199041-2202895 | 2043 | 680 | 72.994 | 5.7 | −0.309 | 5:6 | Mitochondrial |
| CmoCh16G003050.1 | CmoHSP70-11 | 16 | 1399820-1403923 | 2043 | 680 | 73.047 | 5.7 | −0.32 | 5:6 | Mitochondrial |
| CmoCh15G015240.1 | CmoHSP70-12 | 15 | 10324523-10328113 | 2178 | 725 | 75.479 | 5.3 | −0.284 | 8:9 | Chloroplast |
| CmoCh04G021230.1 | CmoHSP70-13 | 4 | 13928946-13932753 | 2121 | 706 | 75.66 | 5.26 | −0.297 | 7:8 | Chloroplast |
| CmoCh09G011250.1 | CmoHSP70-14 | 9 | 6416327-6421038 | 2130 | 709 | 72.927 | 4.98 | −0.298 | 5:6 | Chloroplast |
| CmoCh14G017440.1 | CmoHSP70-15 | 14 | 13583561-13586514 | 1950 | 572 | 62.01 | 5.48 | 0.027 | 2:3 | Cytoplasmic |
| CmoCh17G007790.1 | CmoHSP70-16 | 17 | 7528949-7534538 | 2523 | 840 | 92.562 | 5.32 | −0.425 | 8:9 | Cytoplasmic |
| CmoCh09G009480.1 | CmoHSP70-17 | 9 | 4976312-4985160 | 2283 | 760 | 84.941 | 5.46 | −0.41 | 8:9 | Nuclear |
| CmoCh08G006500.1 | CmoHSP70-18 | 8 | 4233271-4239067 | 2532 | 843 | 92.659 | 5.39 | −0.42 | 8:9 | Cytoplasmic |
| CmoCh01G011840.1 | CmoHSP70-19 | 1 | 9626023-9630560 | 2277 | 758 | 84.845 | 5.62 | −0.411 | 8:9 | Nuclear |
| CmoCh01G013850.1 | CmoHSP70-20 | 1 | 10902171-10939329 | 9318 | 900 | 99.903 | 5.23 | −0.488 | 51:52 | Nuclear, ER |
| CmoCh09G007070.1 | CmoHSP70-21 | 9 | 3490439-3499658 | 2868 | 955 | 99.995 | 5.27 | −0.481 | 14:15 | ER, Nuclear, Cytoplasmic |
| Gene 1 | Gene_2 | Ka | Ks | Ka/Ks | Duplication Type | T (MYA) 1 |
|---|---|---|---|---|---|---|
| CmoCh17G007790.1 | CmoCh08G006500.1 | 0.02935 | 0.338926 | 0.086597 | SD 2 | 11.29753381 |
| CmoCh09G009480.1 | CmoCh01G011840.1 | 0.063151 | 0.467286 | 0.135145 | SD | 15.57619476 |
| CmoCh01G013850.1 | CmoCh09G007070.1 | 3.304909 | 3.049809 | 1.083645 | SD | 101.6603093 |
| CmoCh04G004440.1 | CmoCh16G003050.1 | 0.019189 | 0.364794 | 0.052602 | SD | 12.15981475 |
| CmoCh15G015240.1 | CmoCh04G021230.1 | 0.977111 | 1.342964 | 0.727578 | SD | 44.76545955 |
| CmoCh02G009230.1 | CmoCh15G013530.1 | 0.00843 | 0.420521 | 0.020047 | SD | 14.01737417 |
| CmoCh04G027550.1 | CmoCh15G004130.1 | 0.031706 | 0.633027 | 0.050087 | SD | 21.10089572 |
| CmoCh04G027600.1 | CmoCh15G004100.1 | 0.014152 | 0.492575 | 0.028731 | SD | 16.41915256 |
| CmoCh03G004440.1 | CmoCh07G010280.1 | 0.01195 | 0.521627 | 0.022909 | SD | 17.38757986 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davoudi, M.; Chen, J.; Lou, Q. Genome-Wide Identification and Expression Analysis of Heat Shock Protein 70 (HSP70) Gene Family in Pumpkin (Cucurbita moschata) Rootstock under Drought Stress Suggested the Potential Role of these Chaperones in Stress Tolerance. Int. J. Mol. Sci. 2022, 23, 1918. https://doi.org/10.3390/ijms23031918
Davoudi M, Chen J, Lou Q. Genome-Wide Identification and Expression Analysis of Heat Shock Protein 70 (HSP70) Gene Family in Pumpkin (Cucurbita moschata) Rootstock under Drought Stress Suggested the Potential Role of these Chaperones in Stress Tolerance. International Journal of Molecular Sciences. 2022; 23(3):1918. https://doi.org/10.3390/ijms23031918
Chicago/Turabian StyleDavoudi, Marzieh, Jinfeng Chen, and Qunfeng Lou. 2022. "Genome-Wide Identification and Expression Analysis of Heat Shock Protein 70 (HSP70) Gene Family in Pumpkin (Cucurbita moschata) Rootstock under Drought Stress Suggested the Potential Role of these Chaperones in Stress Tolerance" International Journal of Molecular Sciences 23, no. 3: 1918. https://doi.org/10.3390/ijms23031918