
Role of Dorsomedial Hypothalamus GABAergic Neurons in Sleep–Wake States in Response to Changes in Ambient Temperature in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
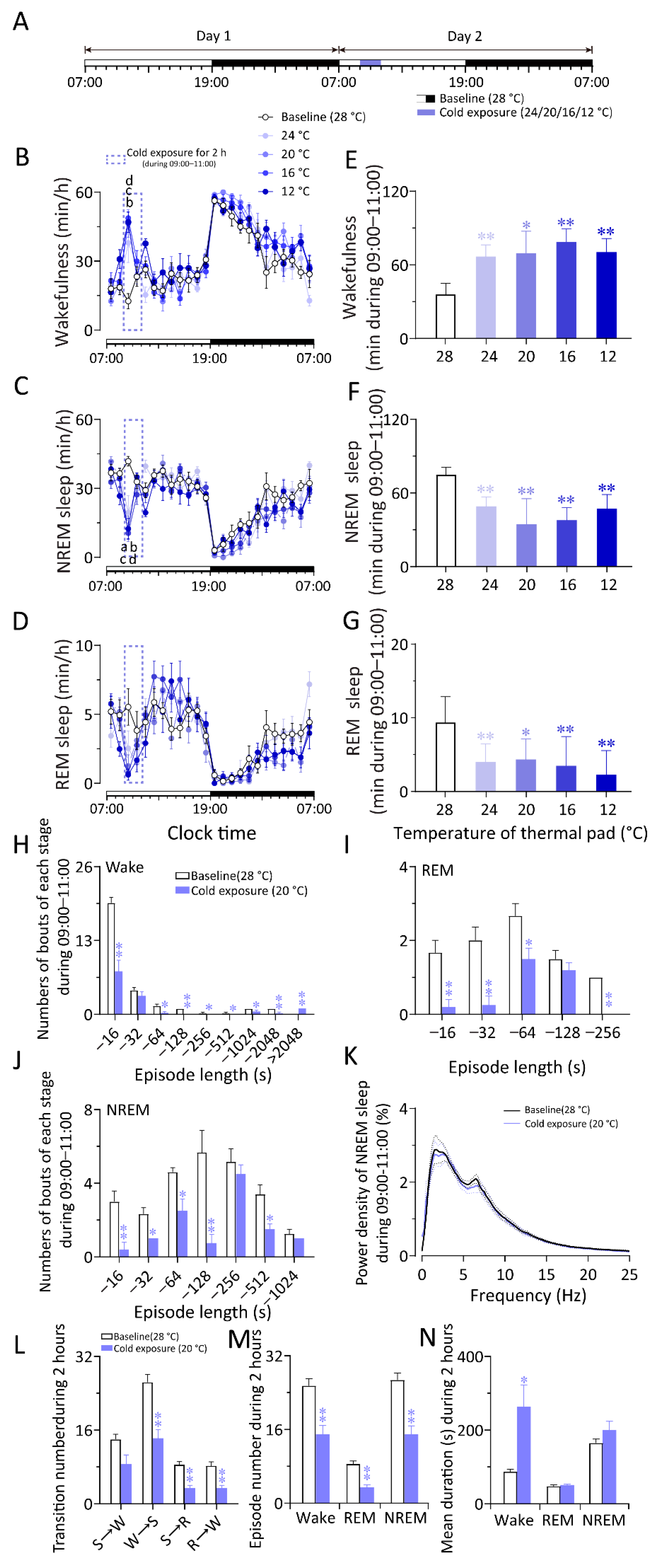
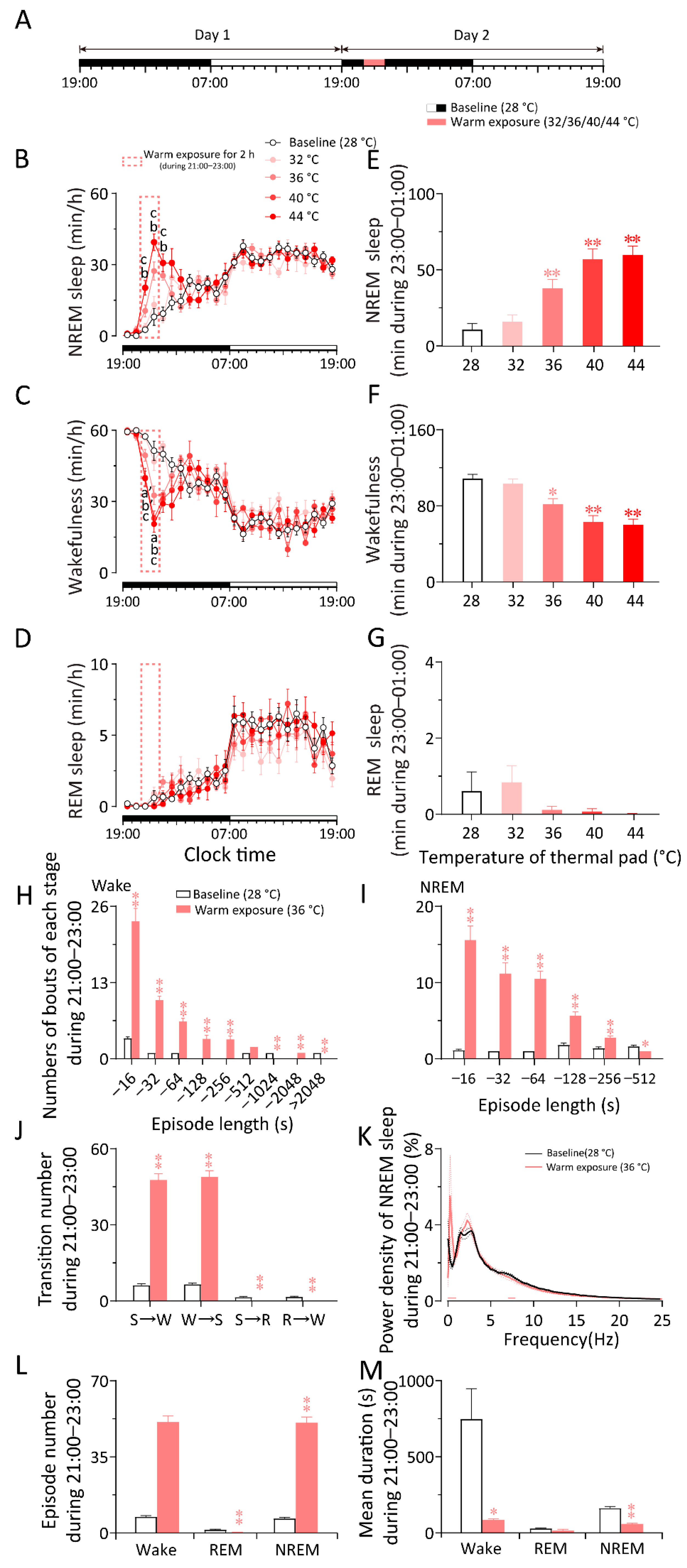
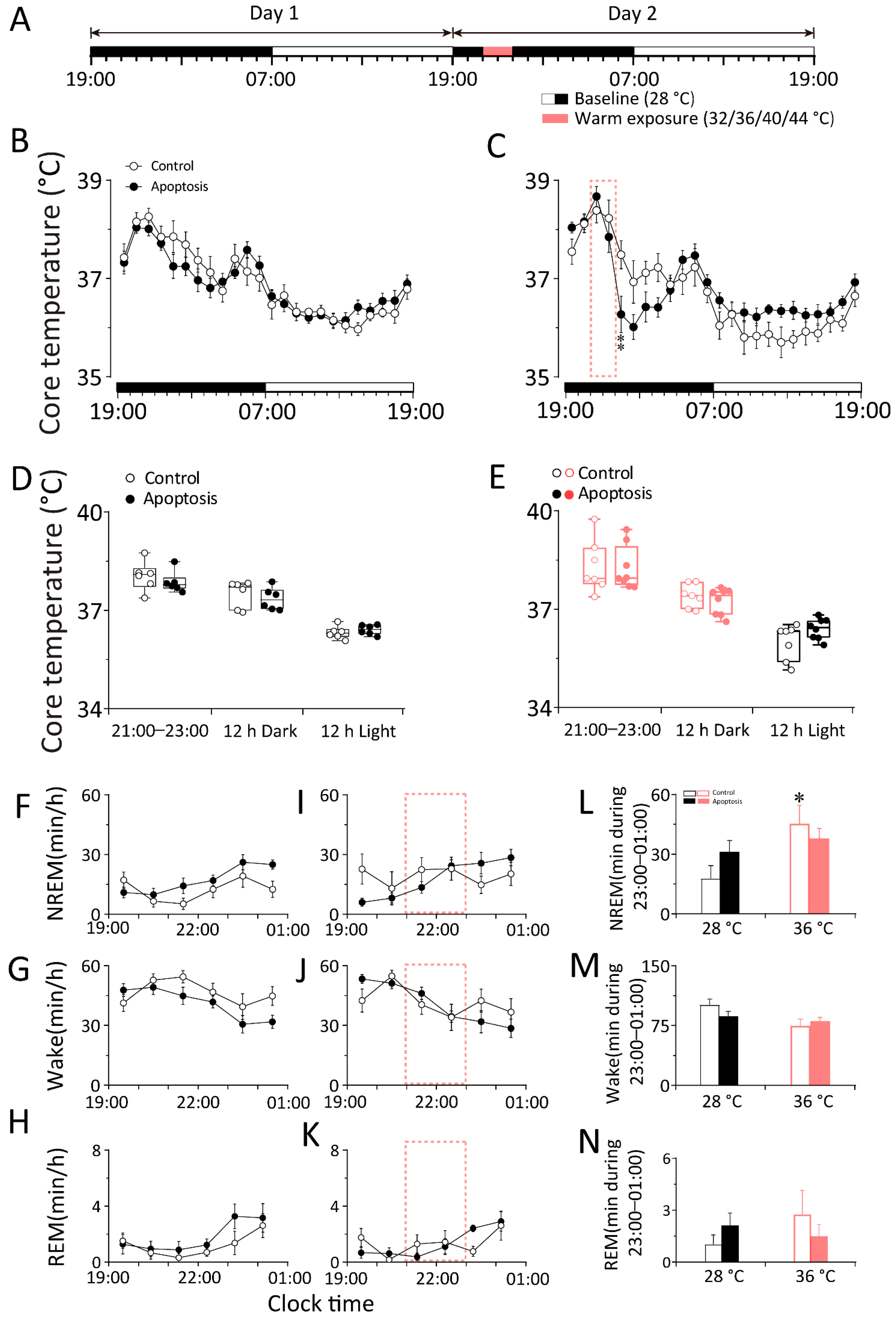
2.1. Cold Exposure Increased Wakefulness While Warm Exposure Increased NREM Sleep in WT Mice
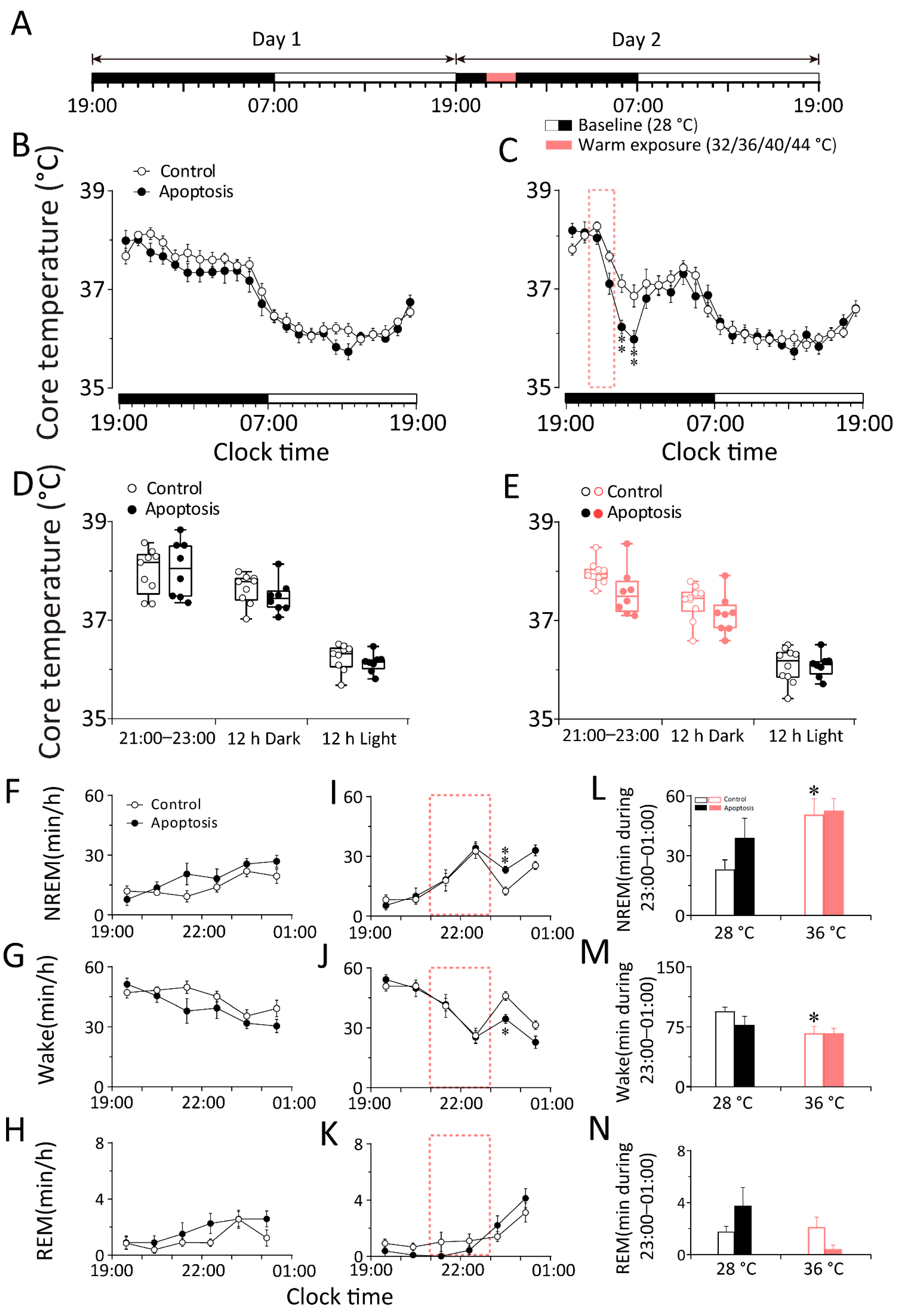
2.2. Whole DMH Neurons Are Necessary to Respond to Changes in Ambient Temperature during Sleep–Wake Behavior in Mice
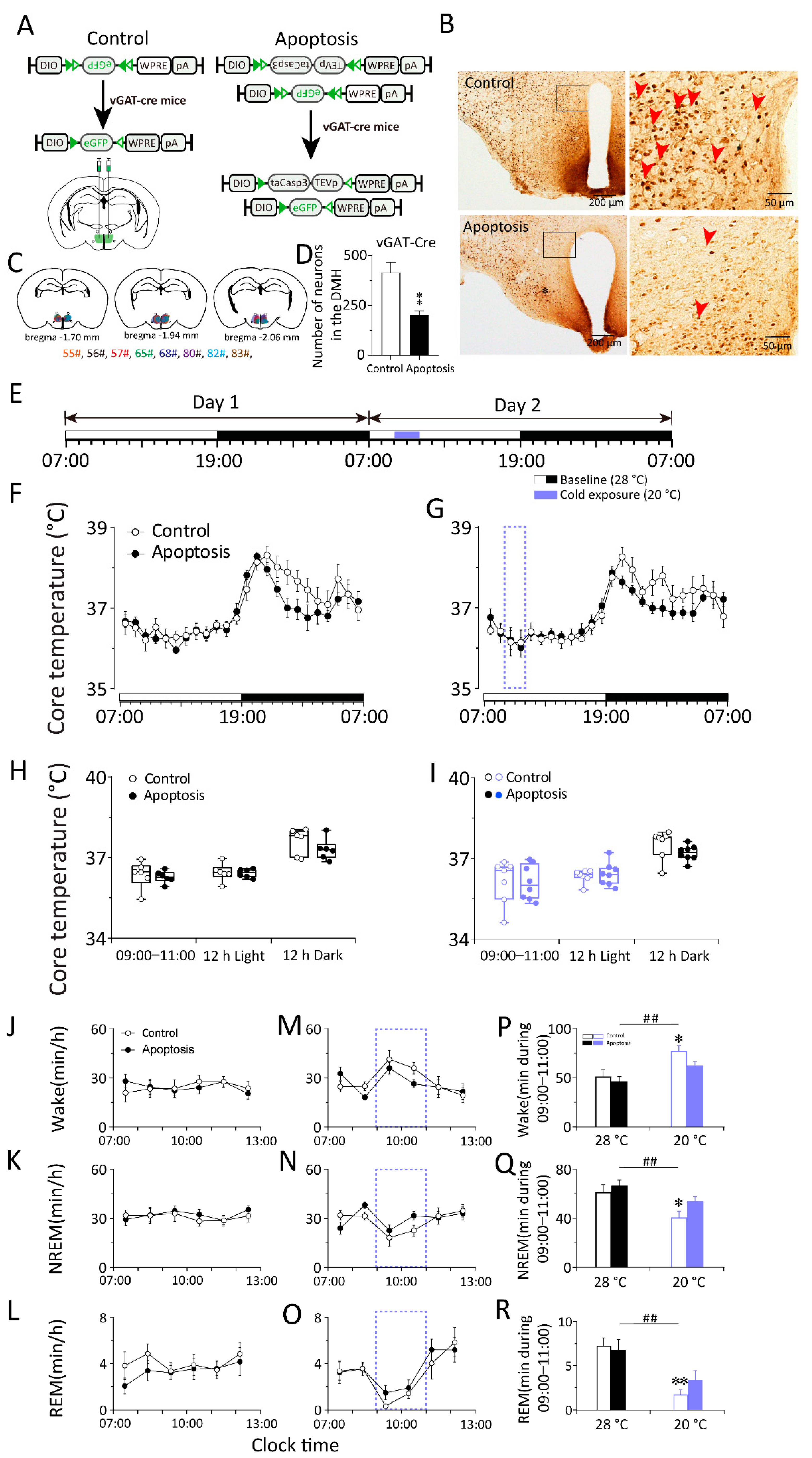
2.3. Response to Changes in Ambient Temperature during Sleep–Wake Behavior in Mice Requires DMH GABAergic Neurons
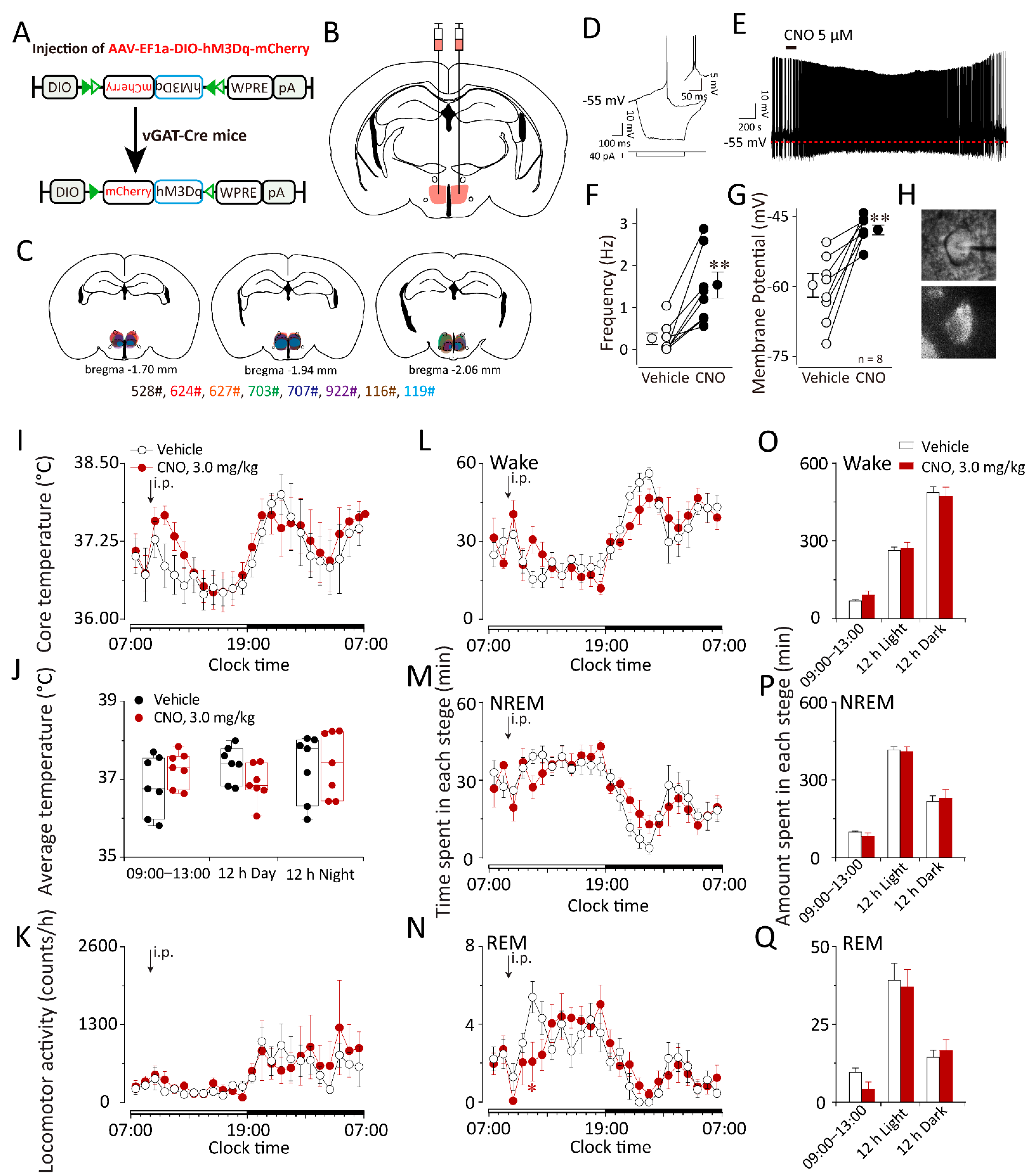
2.4. Chemogenetic Activation of GABAergic Neurons in the DMH Increased Body Temperature during the Inactive Period but Did Not Influence Sleep–Wake Behaviors in vGAT–Cre Mice
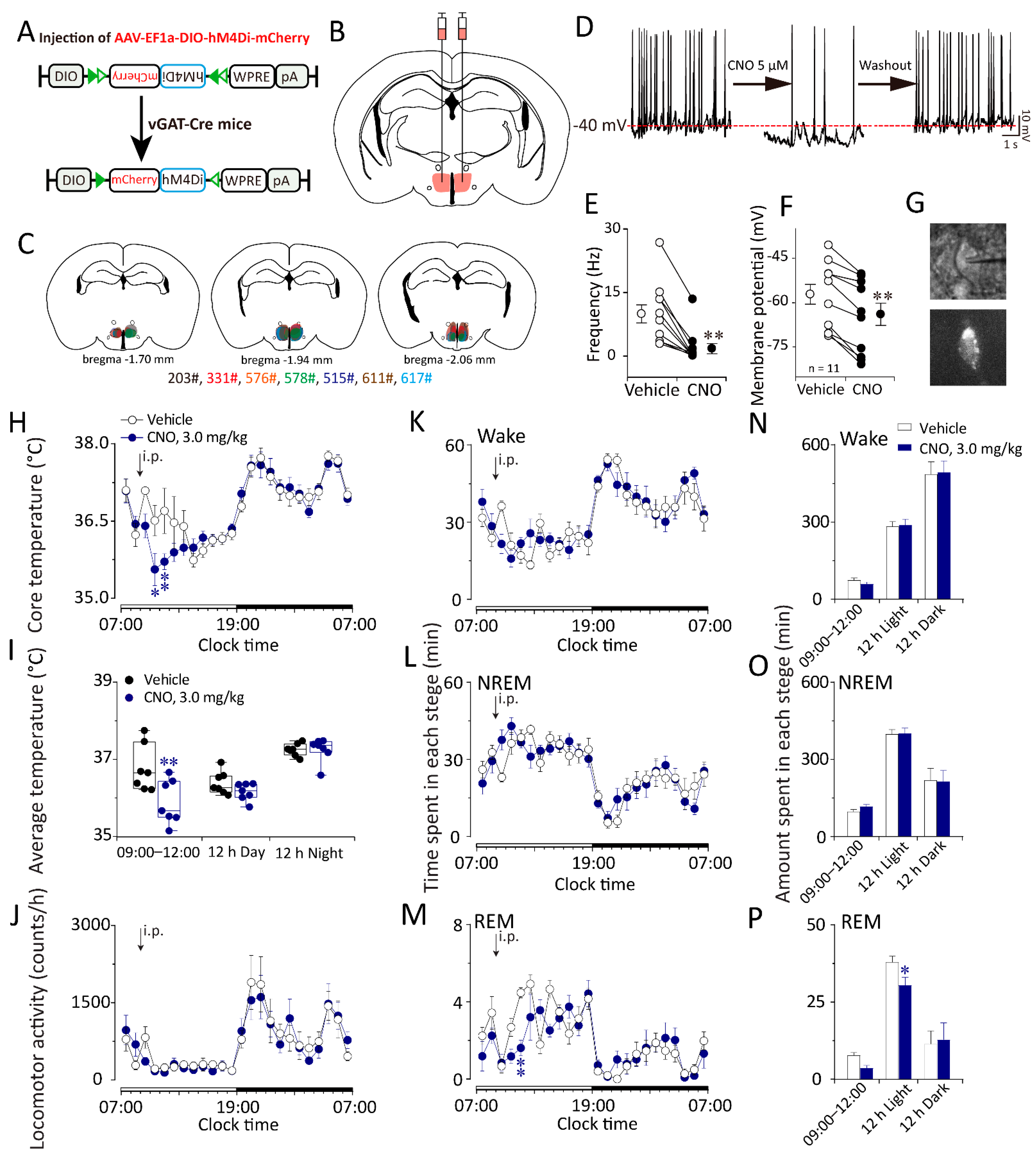
2.5. Chemogenetic Inhibition of GABAergic Neurons in the DMH Decreased Body Temperature and Increased NREM Sleep in vGAT–Cre Mice during the Inactive Period
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. AAV Vectors
4.3. Stereotaxic Surgery
4.4. The Intraperitoneal Implantation of DSI Telemetry Implant in Mice and the Control of the Constant Temperature Hot–Plate
4.5. Polygraphic Recordings and Analysis
4.6. Locomotor Activity and Core Temperature Recordings
4.7. In Vitro Electrophysiology
4.8. Immunohistochemistry
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tan, C.L.; Knight, Z.A. Regulation of Body Temperature by the Nervous System. Neuron 2018, 98, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K. Central circuitries for body temperature regulation and fever. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 301, R1207–R1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahapatra, A.P.K.; Mallick, H.N.; Kumar, V.M. Changes in sleep on chronic exposure to warm and cold ambient temperatures. Physiol. Behav. 2005, 84, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-R.; Zhong, Y.-H.; Jiang, S.; Xu, W.; Xiao, L.; Wang, Z.; Qu, W.-M.; Huang, Z.-L. Dysfunctions of the paraventricular hypothalamic nucleus induce hypersomnia in mice. ELife 2021, 10, e69909. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Li, J.; Tang, K. The Paraventricular Nucleus of the Hypothalamus: Development, Function, and Human Diseases. Endocrinology 2018, 159, 3458–3472. [Google Scholar] [CrossRef]
- Franklin, G.P.; Keith, B.J. The Mouse Brainin Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA, 2001; pp. 1–350. [Google Scholar]
- Shiromani, D.G.; Priyattam, J. Different Neuronal Phenotypes in the Lateral Hypothalamus and Their Role in Sleep and Wakefulness. Mol. Neurobiol. 2004, 29, 41–59. [Google Scholar]
- Ono, D.; Yamanaka, A. Hypothalamic regulation of the sleep/wake cycle. Neurosci. Res. 2017, 118, 74–81. [Google Scholar] [CrossRef]
- Chien, L.; Peilin, C.; Smith, M.S. Neuropeptide Y (Npy) Neurons in the Arcuate Nucleus (Arh) and Dorsomedial Nucleus (Dmh), Areas Activated During Lactation, Project to the Paraventricular Nucleus of the Hypothalamus (Pvh). Regul. Pept. 1998, 75–76, 93–100. [Google Scholar]
- DiMicco, J.A.; Zaretsky, D. The dorsomedial hypothalamus: A new player in thermoregulation. Am. J. Physiol. Integr. Comp. Physiol. 2007, 292, R47–R63. [Google Scholar] [CrossRef]
- Gall, A.; Todd, W.D.; Blumberg, M.S. Development of SCN Connectivity and the Circadian Control of Arousal: A Diminishing Role for Humoral Factors? PLoS ONE 2012, 7, e45338. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Palm, I.F.; La Fleur, S.E.; Scheer, F.; Perreau-Lenz, S.; Ruiter, M.; Kreier, F.; Cailotto, C.; Buijs, R.M. SCN Outputs and the Hypothalamic Balance of Life. J. Biol. Rhythm. 2006, 21, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aston-Jones, G. Brain structures and receptors involved in alertness. Sleep Med. 2005, 6, S3–S7. [Google Scholar] [CrossRef]
- Acosta-Galvan, G.; Yi, C.-X.; van der Vliet, J.; Jhamandas, J.H.; Panula, P.; Angeles-Castellanos, M.; Del Basualdo, M.C.; Escobar, C.; Buijs, R.M. Interaction between hypothalamic dorsomedial nucleus and the suprachiasmatic nucleus determines intensity of food anticipatory behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 5813–5818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.-S.; Wei, H.-H.; Xu, W.; Wang, L.; Qu, W.-M.; Li, R.-X.; Huang, Z.-L. Whole-Brain Monosynaptic Afferent Projections to the Cholecystokinin Neurons of the Suprachiasmatic Nucleus. Front. Neurosci. 2018, 12, 807. [Google Scholar] [CrossRef]
- Wei, H.-H.; Yuan, X.-S.; Chen, Z.-K.; Chen, P.-P.; Xiang, Z.; Qu, W.-M.; Li, R.-X.; Zhou, G.-M.; Huang, Z.-L. Presynaptic inputs to vasopressin neurons in the hypothalamic supraoptic nucleus and paraventricular nucleus in mice. Exp. Neurol. 2021, 343, 113784. [Google Scholar] [CrossRef]
- Jeong, J.H.; Lee, D.K.; Jo, Y.-H. Cholinergic neurons in the dorsomedial hypothalamus regulate food intake. Mol. Metab. 2017, 6, 306–312. [Google Scholar] [CrossRef]
- Jhaveri, K.; Trammell, R.; Toth, L. Effect of environmental temperature on sleep, locomotor activity, core body temperature and immune responses of C57BL/6J mice. Brain, Behav. Immun. 2007, 21, 975–987. [Google Scholar] [CrossRef] [Green Version]
- Joseph, A.; DiMicco, B.; Samuels, C.; Maria, V.; Zaretskaia, D.; Zaretsky, V. The Dorsomedial Hypothalamus and the Response to Stress Part Renaissance, Part Revolution. Pharmacology. Biochem. Behav. 2002, 71, 469–480. [Google Scholar]
- Ajwad, A.; Yaghouby, F.; Huffman, D.; O’Hara, B.; Sunderam, S. Effect of temperature on sleep regulation in an animal epilepsy model. In Proceedings of the 2016 38th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Orlando, FL, USA, 16–20 August 2016; IEEE: New York, NY, USA, 2016; Volume 2016, pp. 1644–1647. [Google Scholar]
- Obál, F., Jr.; Tobler, I.; Borbély, A.A. Effect of Ambient Temperature on the 24-Hour Sleep-Wake Cycle in Normal and Capsaicin-Treated Rats. Physiol. Behav. 1983, 30, 425–430. [Google Scholar] [CrossRef]
- Kräuchi, K.; Cajochen, C.; Werth, E.; Wirz-Justice, A. Warm feet promote the rapid onset of sleep. Nat. Cell Biol. 1999, 401, 36–37. [Google Scholar] [CrossRef]
- Chen, K.-S.; Xu, M.; Zhang, Z.; Chang, W.-C.; Gaj, T.; Schaffer, D.V.; Dan, Y. A Hypothalamic Switch for REM and Non-REM Sleep. Neuron 2018, 97, 1168–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Yang, W.Z.; Gao, C.; Fu, X.; Zhang, W.; Zhou, Q.; Chen, W.; Ni, X.; Lin, J.-K.; Yang, J.; et al. A hypothalamic circuit that controls body temperature. Proc. Natl. Acad. Sci. USA 2017, 114, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon, D.D.; Perrier, L.; Dorman, S.C.; Oddson, B.; Larivière, C.; Serresse, O. Ambient temperature influences metabolic substrate oxidation curves during running and cycling in healthy men. Eur. J. Sport Sci. 2019, 20, 90–99. [Google Scholar] [CrossRef]
- Carpenter, K.C.; Zhou, Y.; Hakenjos, J.M.; Fry, C.D.; Nemzek, J.A. Thermoneutral Housing Temperature Improves Survival in a Murine Model of Polymicrobial Peritonitis. Shock 2020, 54, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Poole, E.I.; Rust, V.A.; Crosby, K.M. Nitric Oxide Acts in the Rat Dorsomedial Hypothalamus to Increase High Fat Food Intake and Glutamate Transmission. Neuroscience 2020, 440, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Houtz, J.; Liao, G.Y.; An, J.J.; Xu, B. Discrete Trkb-Expressing Neurons of the Dorsomedial Hypothalamus Regulate Feeding and Thermogenesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2017218118. [Google Scholar] [CrossRef]
- Faber, C.L.; Deem, J.D.; Phan, B.A.; Doan, T.P.; Ogimoto, K.; Mirzadeh, Z.; Schwartz, M.W.; Morton, G.J. Leptin receptor neurons in the dorsomedial hypothalamus regulate diurnal patterns of feeding, locomotion, and metabolism. ELife 2021, 10, e63671. [Google Scholar] [CrossRef] [PubMed]
- Merkestein, M.; Van Gestel, M.A.; Van Der Zwaal, E.M.; Brans, M.A.; Luijendijk, M.C.; Van Rozen, A.J.; Hendriks, J.; Garner, K.M.; Boender, A.; Pandit, R.; et al. GHS-R1a signaling in the DMH and VMH contributes to food anticipatory activity. Int. J. Obes. 2013, 38, 610–618. [Google Scholar] [CrossRef]
- Stamper, C.E.; Hassell, J.E.; Kapitz, A.J.; Renner, K.J.; Orchinik, M.; Lowry, C.A. Activation of 5-HT1A receptors in the rat dorsomedial hypothalamus inhibits stress-induced activation of the hypothalamic–pituitary–adrenal axis. Stress 2017, 20, 223–230. [Google Scholar] [CrossRef]
- Quaresma, P.G.; Teixeira, P.D.; Wasinski, F.; Campos, A.M.; List, E.O.; Kopchick, J.J.; Donato, J. Cholinergic neurons in the hypothalamus and dorsal motor nucleus of the vagus are directly responsive to growth hormone. Life Sci. 2020, 259, 118229. [Google Scholar] [CrossRef]
- Picard, A.; Metref, S.; Tarussio, D.; Dolci, W.; Berney, X.; Croizier, S.; Labouebe, G.; Thorens, B. Fgf15 Neurons of the Dorsomedial Hypothalamus Control Glucagon Secretion and Hepatic Gluconeogenesis. Diabetes 2021, 70, 1443–1457. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.O.; Franken, P.; Tobler, I.; Borbely, A.A. Effect of elevated ambient temperature on sleep, EEG spectra, and brain temperature in the rat. Am. J. Physiol. Integr. Comp. Physiol. 1995, 268, R1365–R1373. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Bonillas, A.C.; Wong, J.; Padilla, S.L.; Brooks, V.L. Neuropeptide Y suppresses thermogenic and cardiovascular sympathetic nerve activity via Y1 receptors in the paraventricular nucleus and dorsomedial hypothalamus. J. Neuroendocr. 2021, 33, e13006. [Google Scholar] [CrossRef] [PubMed]
- Baimel, C.; E Bartlett, S.; Chiou, L.-C.; Lawrence, A.J.; Muschamp, J.W.; Patkar, O.; Tung, L.-W.; Borgland, S.L. Orexin/hypocretin role in reward: Implications for opioid and other addictions. J. Cereb. Blood Flow Metab. 2014, 172, 334–348. [Google Scholar] [CrossRef]
- Gordon, C.J. Simultaneous measurement of preferred ambient temperature and metabolism in rats. Am. J. Physiol. Content 1988, 254, 229–234. [Google Scholar] [CrossRef]
- Mortola, J.P.; Maskrey, M. Metabolism, Temperature, and Ventilation. Comprehensive Physiology 2011, 1, 1679–1709. [Google Scholar] [CrossRef]
- Krystal, A.D.; Schopler, B.; Kobbe, S.; Williams, C.; Rakatondrainibe, H.; Yoder, A.D.; Klopfer, P. The Relationship of Sleep with Temperature and Metabolic Rate in a Hibernating Primate. PLoS ONE 2013, 8, e69914. [Google Scholar] [CrossRef]
- Van Someren, E.J. More Than a Marker: Interaction between the Circadian Regulation of Temperature and Sleep, Age-Related Changes, and Treatment Possibilities. Chronobiol. Int. 2000, 17, 313–354. [Google Scholar] [CrossRef]
- Chou, T.C.; Scammell, T.E.; Gooley, J.J.; Gaus, S.E.; Saper, C.B.; Lu, J. Critical Role of Dorsomedial Hypothalamic Nucleus in a Wide Range of Behavioral Circadian Rhythms. J. Neurosci. 2003, 23, 10691–10702. [Google Scholar] [CrossRef]
- Refinetti, R.; Menaker, M. The Circadian Rhythm of Body Temperature. Psysiol. Behav. 1992, 51, 631–637. [Google Scholar] [CrossRef]
- Farsi, H.; Achaâban, M.R.; Piro, M.; Bothorel, B.; Ouassat, M.; Challet, E.; Pévet, P.; El Allali, K. Entrainment of circadian rhythms of locomotor activity by ambient temperature cycles in the dromedary camel. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Refinetti, R. Entrainment of Circadian Rhythm by Ambient Temperature Cycles in Mice. J. Biol. Rhythm. 2010, 25, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Huhman, A.; Hennessey, C.; Elliott Albers, H. Rhythms of Glutamic Acid Decarboxylase Mrna in the Suprachiasmatic Nucleus. J. Biol. Rhythm. 1996, 11, 311–316. [Google Scholar]
- Muzet, A.; Libert, J.-P.; Candas, V. Ambient temperature and human sleep. Cell. Mol. Life Sci. 1984, 40, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Dispersyn, G.; Sauvet, F.; Gomez-Merino, D.; Ciret, S.; Drogou, C.; Leger, D.; Gallopin, T.; Chennaoui, M. The homeostatic and circadian sleep recovery responses after total sleep deprivation in mice. J. Sleep Res. 2017, 26, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.-L.; Qu, W.-M.; Li, W.-D.; Mochizuki, T.; Eguchi, N.; Watanabe, T.; Urade, Y.; Hayaishi, O. Arousal effect of orexin A depends on activation of the histaminergic system. Proc. Natl. Acad. Sci. USA 2001, 98, 9965–9970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.-L.; Qu, W.-M.; Eguchi, N.; Chen, J.-F.; A Schwarzschild, M.; Fredholm, B.B.; Urade, Y.; Hayaishi, O. Adenosine A2A, but not A1, receptors mediate the arousal effect of caffeine. Nat. Neurosci. 2005, 8, 858–859. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.-S.; Wang, L.; Dong, H.; Qu, W.-M.; Yang, S.-R.; Cherasse, Y.; Lazarus, M.; Schiffmann, S.N.; D’Exaerde, A.D.K.; Li, R.-X.; et al. Striatal adenosine A2A receptor neurons control active-period sleep via parvalbumin neurons in external globus pallidus. ELife 2017, 6, e29055. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Zhang, M.-Q.; Sun, X.; Liu, W.-Y.; Huang, Z.-L.; Wang, Y.-Q. Role of Dorsomedial Hypothalamus GABAergic Neurons in Sleep–Wake States in Response to Changes in Ambient Temperature in Mice. Int. J. Mol. Sci. 2022, 23, 1270. https://doi.org/10.3390/ijms23031270
Li L, Zhang M-Q, Sun X, Liu W-Y, Huang Z-L, Wang Y-Q. Role of Dorsomedial Hypothalamus GABAergic Neurons in Sleep–Wake States in Response to Changes in Ambient Temperature in Mice. International Journal of Molecular Sciences. 2022; 23(3):1270. https://doi.org/10.3390/ijms23031270
Chicago/Turabian StyleLi, Lei, Meng-Qi Zhang, Xiao Sun, Wen-Ying Liu, Zhi-Li Huang, and Yi-Qun Wang. 2022. "Role of Dorsomedial Hypothalamus GABAergic Neurons in Sleep–Wake States in Response to Changes in Ambient Temperature in Mice" International Journal of Molecular Sciences 23, no. 3: 1270. https://doi.org/10.3390/ijms23031270