
Time-Dependent Changes in Protein Composition of Medial Prefrontal Cortex in Rats with Neuropathic Pain

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
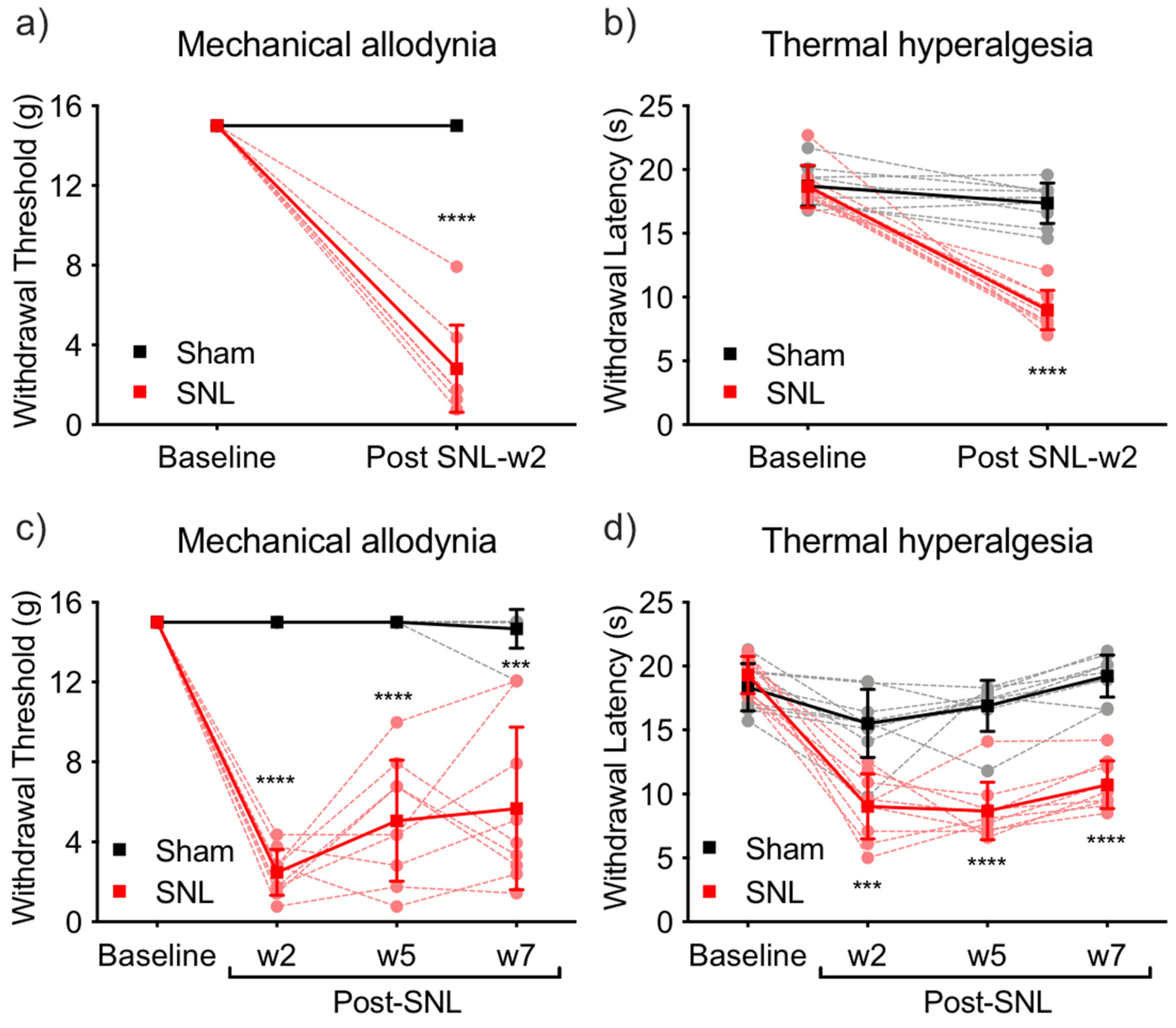
2.1. Pain Behavior
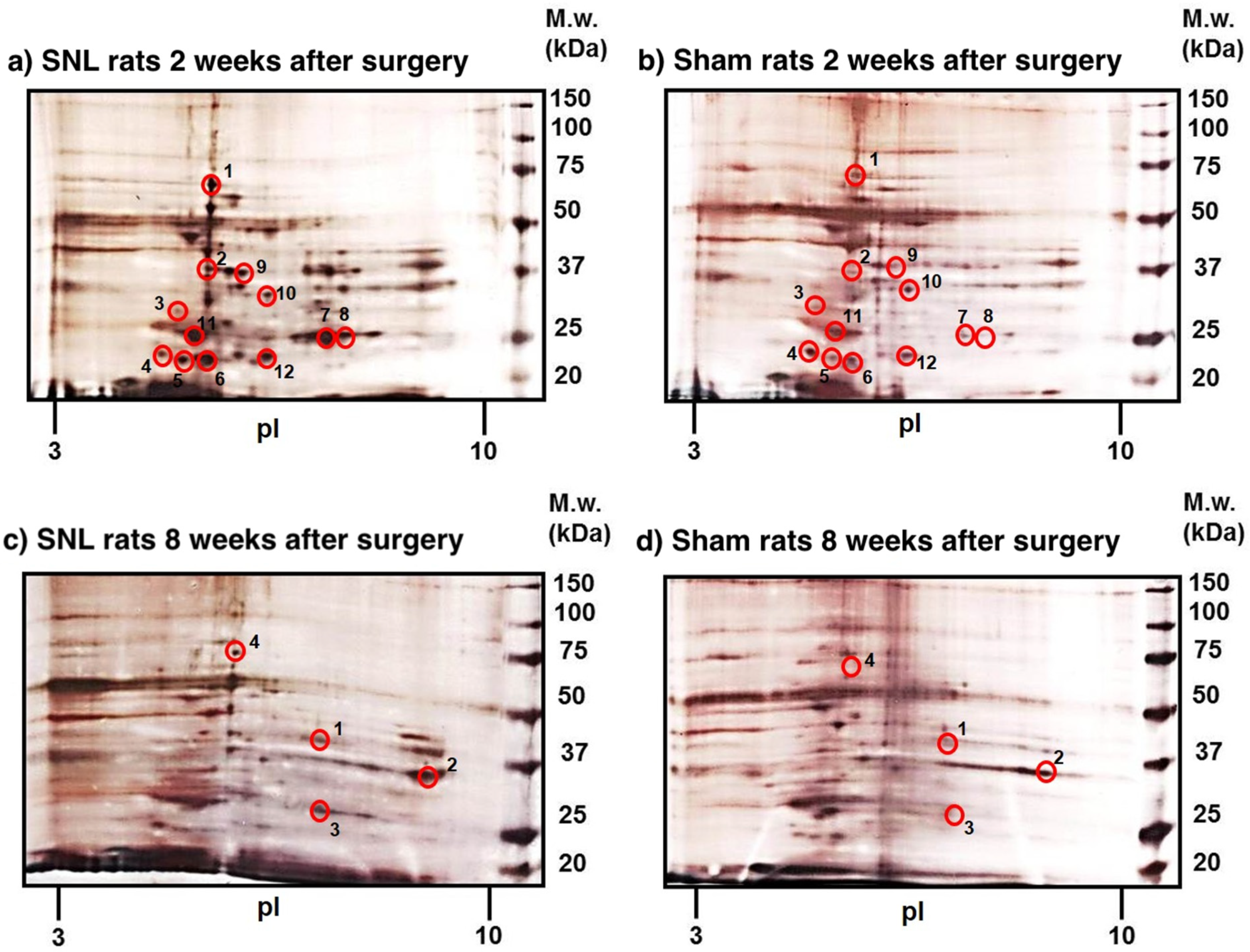
2.2. Identification of Differently Expressed Proteins
2.2.1. Differently Expressed Proteins Identified at 2 Weeks Post SNL
2.2.2. Differently Expressed Proteins Identified at 8 Weeks Post SNL
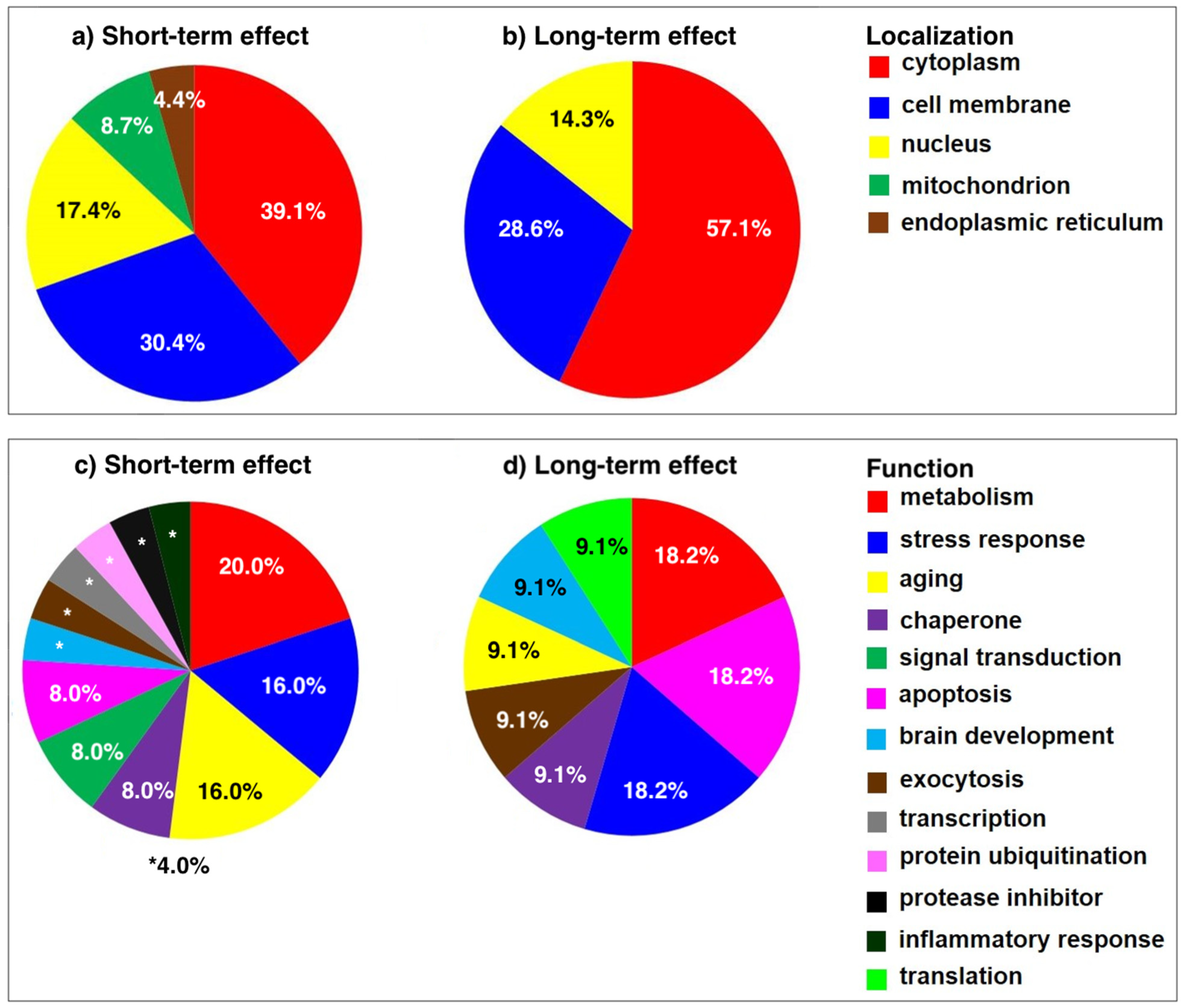
2.3. Functional Significance of Altered Proteins
2.3.1. Functional Significance of Proteins Altered at 2 Weeks Post SNL
2.3.2. Functional Significance of Proteins Altered at 8 Weeks Post SNL
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Surgeries
4.4. Behavioral Tests
4.4.1. Von Frey Tests
4.4.2. Hargreaves Tests
4.4.3. Test Groups
4.4.4. Statistical Analysis
4.5. Preparation of PNS Fraction from mPFC Tissues of SNL/Sham-Operated Rats
4.6. 2D-ELFO
4.6.1. Sample Preparation for Isoelectric Focusing (IEF)
4.6.2. Equilibration of IPG Strips and SDS-PAGE
4.7. Silver Staining
4.8. Image Analysis
4.9. Identification of Proteins
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Emery, E.C.; Luiz, A.P.; Wood, J.N. Nav1.7 and other voltage-gated sodium channels as drug targets for pain relief. Expert Opin. Ther. Targets 2016, 20, 975–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapplebeck, J.C.S.; Lorenzo, L.E.; Lee, K.Y.; Gauthier, C.; Muley, M.M.; De Koninck, Y.; Prescott, S.A.; Salter, M.W. Chloride Dysregulation through Downregulation of KCC2 Mediates Neuropathic Pain in Both Sexes. Cell Rep. 2019, 28, 590–596.e4. [Google Scholar] [CrossRef]
- Coull, J.A.; Boudreau, D.; Bachand, K.; Prescott, S.A.; Nault, F.; Sik, A.; De Koninck, P.; De Koninck, Y. Trans-synaptic shift in anion gradient in spinal lamina I neurons as a mechanism of neuropathic pain. Nature 2003, 424, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Farmer, M.A.; Baliki, M.N.; Apkarian, A.V. A dynamic network perspective of chronic pain. Neurosci. Lett. 2012, 520, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Baliki, M.N.; Petre, B.; Torbey, S.; Herrmann, K.M.; Huang, L.; Schnitzer, T.J.; Fields, H.L.; Apkarian, A.V. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat. Neurosci. 2012, 15, 1117–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apkarian, A.V.; Baliki, M.N.; Farmer, M.A. Predicting transition to chronic pain. Curr. Opin. Neurol. 2013, 26, 360–367. [Google Scholar] [CrossRef]
- Tracey, I.; Bushnell, M.C. How neuroimaging studies have challenged us to rethink: Is chronic pain a disease? J. Pain Off. J. Am. Pain Soc. 2009, 10, 1113–1120. [Google Scholar] [CrossRef]
- Harris, R.E.; Napadow, V.; Huggins, J.P.; Pauer, L.; Kim, J.; Hampson, J.; Sundgren, P.C.; Foerster, B.; Petrou, M.; Schmidt-Wilcke, T.; et al. Pregabalin rectifies aberrant brain chemistry, connectivity, and functional response in chronic pain patients. Anesthesiology 2013, 119, 1453–1464. [Google Scholar] [CrossRef] [Green Version]
- Iannetti, G.D.; Zambreanu, L.; Wise, R.G.; Buchanan, T.J.; Huggins, J.P.; Smart, T.S.; Vennart, W.; Tracey, I. Pharmacological modulation of pain-related brain activity during normal and central sensitization states in humans. Proc. Natl. Acad. Sci. USA 2005, 102, 18195–18200. [Google Scholar] [CrossRef] [Green Version]
- Oertel, B.G.; Preibisch, C.; Wallenhorst, T.; Hummel, T.; Geisslinger, G.; Lanfermann, H.; Lotsch, J. Differential opioid action on sensory and affective cerebral pain processing. Clin. Pharmacol. Ther. 2008, 83, 577–588. [Google Scholar] [CrossRef]
- Rauck, R.; Coffey, R.J.; Schultz, D.M.; Wallace, M.S.; Webster, L.R.; McCarville, S.E.; Grigsby, E.J.; Page, L.M. Intrathecal gabapentin to treat chronic intractable noncancer pain. Anesthesiology 2013, 119, 675–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hama, A.; Natsume, T.; Ogawa, S.Y.; Awaga, Y.; Hayashi, I.; Matsuda, A.; Takamatsu, H. Pain-Related Behavior and Brain Activation in a Cynomolgus Macaque Model of Postoperative Pain. CNS Neurol. Disord. Drug Targets 2018, 17, 348–360. [Google Scholar] [CrossRef]
- Bannister, K.; Qu, C.; Navratilova, E.; Oyarzo, J.; Xie, J.Y.; King, T.; Dickenson, A.H.; Porreca, F. Multiple sites and actions of gabapentin-induced relief of ongoing experimental neuropathic pain. Pain 2017, 158, 2386–2395. [Google Scholar] [CrossRef]
- Navratilova, E.; Xie, J.Y.; Meske, D.; Qu, C.; Morimura, K.; Okun, A.; Arakawa, N.; Ossipov, M.; Fields, H.L.; Porreca, F. Endogenous opioid activity in the anterior cingulate cortex is required for relief of pain. J. Neurosci. 2015, 35, 7264–7271. [Google Scholar] [CrossRef] [Green Version]
- Johansen, J.P.; Fields, H.L.; Manning, B.H. The affective component of pain in rodents: Direct evidence for a contribution of the anterior cingulate cortex. Proc. Natl. Acad. Sci. USA 2001, 98, 8077–8082. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, M.C.; Ceko, M.; Low, L.A. Cognitive and emotional control of pain and its disruption in chronic pain. Nat. Rev. Neurosci. 2013, 14, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Villemure, C.; Bushnell, M.C. Mood influences supraspinal pain processing separately from attention. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 705–715. [Google Scholar] [CrossRef]
- Wager, T.D.; Rilling, J.K.; Smith, E.E.; Sokolik, A.; Casey, K.L.; Davidson, R.J.; Kosslyn, S.M.; Rose, R.M.; Cohen, J.D. Placebo-induced changes in FMRI in the anticipation and experience of pain. Science 2004, 303, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.C.; Belfrage, M.; Stone-Elander, S.; Hansson, P.; Ingvar, M. Central representation of chronic ongoing neuropathic pain studied by positron emission tomography. Pain 1995, 63, 225–236. [Google Scholar] [CrossRef]
- Willoch, F.; Gamringer, U.; Medele, R.; Steude, U.; Tolle, T.R. Analgesia by electrostimulation of the trigeminal ganglion in patients with trigeminopathic pain: A PET activation study. Pain 2003, 103, 119–130. [Google Scholar] [CrossRef]
- Becerra, L.; Navratilova, E.; Porreca, F.; Borsook, D. Analogous Responses in the Nucleus Accumbens and Cingulate Cortex to Pain Onset (Aversion) and Offset (Relief) in Rats and Humans. J. Neurophysiol. 2013, 110, 1221–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Gadotti, V.M.; Chen, L.; Souza, I.A.; Stemkowski, P.L.; Zamponi, G.W. Role of Prelimbic GABAergic Circuits in Sensory and Emotional Aspects of Neuropathic Pain. Cell Rep. 2015, 12, 752–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Gadotti, V.M.; Chen, L.; Souza, I.A.; Huang, S.; Wang, D.; Ramakrishnan, C.; Deisseroth, K.; Zhang, Z.; Zamponi, G.W. A neuronal circuit for activating descending modulation of neuropathic pain. Nat. Neurosci. 2019, 22, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Price, T.J.; Ray, P.R. Recent advances toward understanding the mysteries of the acute to chronic pain transition. Curr. Opin. Physiol. 2019, 11, 42–50. [Google Scholar] [CrossRef]
- Zubieta, J.K.; Stohler, C.S. Neurobiological mechanisms of placebo responses. Ann. N. Y. Acad. Sci. 2009, 1156, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Johansen, J.P.; Fields, H.L. Glutamatergic activation of anterior cingulate cortex produces an aversive teaching signal. Nat. Neurosci. 2004, 7, 398–403. [Google Scholar] [CrossRef]
- LaGraize, S.C.; Labuda, C.J.; Rutledge, M.A.; Jackson, R.L.; Fuchs, P.N. Differential effect of anterior cingulate cortex lesion on mechanical hypersensitivity and escape/avoidance behavior in an animal model of neuropathic pain. Exp. Neurol. 2004, 188, 139–148. [Google Scholar] [CrossRef]
- Qu, C.; King, T.; Okun, A.; Lai, J.; Fields, H.L.; Porreca, F. Lesion of the rostral anterior cingulate cortex eliminates the aversiveness of spontaneous neuropathic pain following partial or complete axotomy. Pain 2011, 152, 1641–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashmi, J.A.; Baliki, M.N.; Huang, L.; Baria, A.T.; Torbey, S.; Hermann, K.M.; Schnitzer, T.J.; Apkarian, A.V. Shape shifting pain: Chronification of back pain shifts brain representation from nociceptive to emotional circuits. Brain 2013, 136 Pt 9, 2751–2768. [Google Scholar] [CrossRef] [Green Version]
- Seminowicz, D.A.; Laferriere, A.L.; Millecamps, M.; Yu, J.S.C.; Coderre, T.J.; Bushnell, M.C. MRI structural brain changes associated with sensory and emotional function in a rat model of long-term neuropathic pain. NeuroImage 2009, 47, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Ajit, S.K.; Ramineni, S.; Edris, W.; Hunt, R.A.; Hum, W.T.; Hepler, J.R.; Young, K.H. RGSZ1 interacts with protein kinase C interacting protein PKCI-1 and modulates mu opioid receptor signaling. Cell. Signal. 2007, 19, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Blazquez, P.; Rodriguez-Munoz, M.; Montero, C.; de la Torre-Madrid, E.; Garzon, J. Calcium/calmodulin-dependent protein kinase II supports morphine antinociceptive tolerance by phosphorylation of glycosylated phosducin-like protein. Neuropharmacology 2008, 54, 319–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, P.; Watanabe, H.; Ossipov, M.H.; Bakalkin, G.; Artemenko, K.; Bergquist, J. Proteomics of neuropathic pain: Proteins and signaling pathways affected in a rat model. J. Proteome Res. 2014, 13, 3957–3965. [Google Scholar] [CrossRef]
- Milligan, G.; Kostenis, E. Heterotrimeric G-proteins: A short history. Br. J. Pharmacol. 2006, 147 (Suppl 1), S46–S55. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Bajpayee, N.S. Molecular mechanisms of go signaling. Neurosignals 2009, 17, 23–41. [Google Scholar] [CrossRef] [Green Version]
- Singh, O.V.; Yaster, M.; Xu, J.T.; Guan, Y.; Guan, X.; Dharmarajan, A.M.; Raja, S.N.; Zeitlin, P.L.; Tao, Y.X. Proteome of synaptosome-associated proteins in spinal cord dorsal horn after peripheral nerve injury. Proteomics 2009, 9, 1241–1253. [Google Scholar] [CrossRef] [Green Version]
- Alzate, O.; Hussain, S.R.; Goettl, V.M.; Tewari, A.K.; Madiai, F.; Stephens, R.L., Jr.; Hackshaw, K.V. Proteomic identification of brainstem cytosolic proteins in a neuropathic pain model. Brain Res. Mol. Brain Res. 2004, 128, 193–200. [Google Scholar] [CrossRef]
- Karu, K.; Swanwick, R.S.; Novejarque-Gadea, A.; Antunes-Martins, A.; Thomas, B.; Yoshimi, E.; Foster, W.; Fang, M.; McMahon, S.B.; Bennett, D.L.H.; et al. Quantitative Proteomic Analysis of the Central Amygdala in Neuropathic Pain Model Rats. J. Proteome Res. 2020, 19, 1592–1619. [Google Scholar] [CrossRef]
- Persoon, C.M.; Hoogstraaten, R.I.; Nassal, J.P.; van Weering, J.R.T.; Kaeser, P.S.; Toonen, R.F.; Verhage, M. The RAB3-RIM Pathway Is Essential for the Release of Neuromodulators. Neuron 2019, 104, 1065–1080.e12. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Zhan, X.; Li, M.; Song, Z.; Liu, C.; Peng, F.; Guo, Q. Identification of differentially expressed proteins in the spinal cord of neuropathic pain models with PKCgamma silence by proteomic analysis. Brain Res. 2012, 1440, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Scapagnini, G.; Ravagna, A.; Giuffrida Stella, A.M.; Butterfield, D.A. Molecular chaperones and their roles in neural cell differentiation. Dev. Neurosci. 2002, 24, 1–13. [Google Scholar] [CrossRef]
- Zou, W.; Xu, W.; Song, Z.; Zhong, T.; Weng, Y.; Huang, C.; Li, M.; Zhang, C.; Zhan, X.; Guo, Q. Proteomic Identification of an Upregulated Isoform of Annexin A3 in the Spinal Cords of Rats in a Neuropathic Pain Model. Front. Neurosci. 2017, 11, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komori, N.; Takemori, N.; Kim, H.K.; Singh, A.; Hwang, S.H.; Foreman, R.D.; Chung, K.; Chung, J.M.; Matsumoto, H. Proteomics study of neuropathic and nonneuropathic dorsal root ganglia: Altered protein regulation following segmental spinal nerve ligation injury. Physiol. Genom. 2007, 29, 215–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederberger, E.; Geisslinger, G. Proteomics in neuropathic pain research. Anesthesiology 2008, 108, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, H.; Horinaka, M.; Yoshida, T.; Yano, K.; Goda, A.E.; Yasuda, S.; Wakada, M.; Sakai, T. Targeting the glyoxalase pathway enhances TRAIL efficacy in cancer cells by downregulating the expression of antiapoptotic molecules. Mol. Cancer Ther. 2012, 11, 2294–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolgacheva, L.P.; Berezhnov, A.V.; Fedotova, E.I.; Zinchenko, V.P.; Abramov, A.Y. Role of DJ-1 in the mechanism of pathogenesis of Parkinson’s disease. J. Bioenerg. Biomembr. 2019, 51, 175–188. [Google Scholar] [CrossRef] [Green Version]
- Dunyak, B.M.; Gestwicki, J.E. Peptidyl-Proline Isomerases (PPIases): Targets for Natural Products and Natural Product-Inspired Compounds. J. Med. Chem. 2016, 59, 9622–9644. [Google Scholar] [CrossRef] [Green Version]
- Chuang, D.M.; Hough, C.; Senatorov, V.V. Glyceraldehyde-3-phosphate dehydrogenase, apoptosis, and neurodegenerative diseases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 269–290. [Google Scholar] [CrossRef]
- Indraswari, F.; Wong, P.T.; Yap, E.; Ng, Y.K.; Dheen, S.T. Upregulation of Dpysl2 and Spna2 gene expression in the rat brain after ischemic stroke. Neurochem. Int. 2009, 55, 235–242. [Google Scholar] [CrossRef]
- Lei, L.G.; Zhang, Y.Q.; Zhao, Z.Q. Pain-related aversion and Fos expression in the central nervous system in rats. Neuroreport 2004, 15, 67–71. [Google Scholar] [CrossRef]
- Bliss, T.V.; Collingridge, G.L.; Kaang, B.K.; Zhuo, M. Synaptic plasticity in the anterior cingulate cortex in acute and chronic pain. Nat. Rev. Neurosci. 2016, 17, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Ujcikova, H.; Vosahlikova, M.; Roubalova, L.; Svoboda, P. Proteomic analysis of protein composition of rat forebrain cortex exposed to morphine for 10days; comparison with animals exposed to morphine and subsequently nurtured for 20days in the absence of this drug. J. Proteom. 2016, 145, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Baggerman, G.; Vierstraete, E.; De Loof, A.; Schoofs, L. Gel-based versus gel-free proteomics: A review. Comb. Chem. High Throughput Screen. 2005, 8, 669–677. [Google Scholar] [CrossRef]
- Gauci, V.J.; Wright, E.P.; Coorssen, J.R. Quantitative proteomics: Assessing the spectrum of in-gel protein detection methods. J. Chem. Biol. 2011, 4, 3–29. [Google Scholar] [CrossRef] [Green Version]
- Gygi, S.P.; Rist, B.; Gerber, S.A.; Turecek, F.; Gelb, M.H.; Aebersold, R. Quantitative analysis of complex protein mixtures using isotope-coded affinity tags. Nat. Biotechnol. 1999, 17, 994–999. [Google Scholar] [CrossRef] [PubMed]
- De Felice, M.; Sanoja, R.; Wang, R.; Vera-Portocarrero, L.; Oyarzo, J.; King, T.; Ossipov, M.H.; Vanderah, T.W.; Lai, J.; Dussor, G.O.; et al. Engagement of descending inhibition from the rostral ventromedial medulla protects against chronic neuropathic pain. Pain 2011, 152, 2701–2709. [Google Scholar] [CrossRef] [Green Version]
- Yarnitsky, D. Role of endogenous pain modulation in chronic pain mechanisms and treatment. Pain 2015, 156 (Suppl 1), S24–S31. [Google Scholar] [CrossRef]
- Yarnitsky, D. Conditioned pain modulation (the diffuse noxious inhibitory control-like effect): Its relevance for acute and chronic pain states. Curr. Opin. Anaesthesiol. 2010, 23, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Yarnitsky, D.; Crispel, Y.; Eisenberg, E.; Granovsky, Y.; Ben-Nun, A.; Sprecher, E.; Best, L.A.; Granot, M. Prediction of chronic post-operative pain: Pre-operative DNIC testing identifies patients at risk. Pain 2008, 138, 22–28. [Google Scholar] [CrossRef]
- Moutal, A.; White, K.A.; Chefdeville, A.; Laufmann, R.N.; Vitiello, P.F.; Feinstein, D.; Weimer, J.M.; Khanna, R. Dysregulation of CRMP2 Post-Translational Modifications Drive Its Pathological Functions. Mol. Neurobiol. 2019, 56, 6736–6755. [Google Scholar] [CrossRef] [PubMed]
- Moutal, A.; Yang, X.; Li, W.; Gilbraith, K.B.; Luo, S.; Cai, S.; Francois-Moutal, L.; Chew, L.A.; Yeon, S.K.; Bellampalli, S.S.; et al. CRISPR/Cas9 editing of Nf1 gene identifies CRMP2 as a therapeutic target in neurofibromatosis type 1-related pain that is reversed by (S)-Lacosamide. Pain 2017, 158, 2301–2319. [Google Scholar] [CrossRef]
- Buchta, W.C.; Moutal, A.; Hines, B.; Garcia-Keller, C.; Smith, A.C.W.; Kalivas, P.; Khanna, R.; Riegel, A.C. Dynamic CRMP2 Regulation of CaV2.2 in the Prefrontal Cortex Contributes to the Reinstatement of Cocaine Seeking. Mol. Neurobiol. 2020, 57, 346–357. [Google Scholar] [CrossRef]
- Metz, A.E.; Yau, H.J.; Centeno, M.V.; Apkarian, A.V.; Martina, M. Morphological and functional reorganization of rat medial prefrontal cortex in neuropathic pain. Proc. Natl. Acad. Sci. USA 2009, 106, 2423–2428. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, C.S.; Khan, S.A.; Xu, S.; Cha, M.; Masri, R.; Seminowicz, D.A. Behavioral, metabolic and functional brain changes in a rat model of chronic neuropathic pain: A longitudinal MRI study. Neuroimage 2015, 107, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Dixon, W.J. Efficient analysis of experimental observations. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 441–462. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef]
- Rabilloud, T.; Vuillard, L.; Gilly, C.; Lawrence, J.J. Silver-staining of proteins in polyacrylamide gels: A general overview. Cell Mol. Biol. 1994, 40, 57–75. [Google Scholar] [PubMed]
- Gharahdaghi, F.; Weinberg, C.R.; Meagher, D.A.; Imai, B.S.; Mische, S.M. Mass spectrometric identification of proteins from silver-stained polyacrylamide gel: A method for the removal of silver ions to enhance sensitivity. Electrophoresis 1999, 20, 601–605. [Google Scholar] [CrossRef]
- Andon, N.L.; Hollingworth, S.; Koller, A.; Greenland, A.J.; Yates, J.R., 3rd; Haynes, P.A. Proteomic characterization of wheat amyloplasts using identification of proteins by tandem mass spectrometry. Proteomics 2002, 2, 1156–1168. [Google Scholar] [CrossRef]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.J.; Liu, T.; Monroe, M.E.; Strittmatter, E.F.; Jacobs, J.M.; Kangas, L.J.; Petritis, K.; Camp, D.G., 2nd; Smith, R.D. Probability-based evaluation of peptide and protein identifications from tandem mass spectrometry and SEQUEST analysis: The human proteome. J. Proteome Res. 2005, 4, 53–62. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Spot | Accession Number | Protein | Exclusive Unique Peptides | SC a (%) | MW (kDa) | pI b | Change c (Fold) | p-Value |
|---|---|---|---|---|---|---|---|---|
| 1 | NP_077327.1 | Heat-shock cognate 71 kDa protein | 23 | 69 | 71 | 5.47 | ↑2.7 | 0.0398 |
| 2 | NP_059023.1 | Guanine nucleotide-binding protein G(o) subunit alpha | 16 | 58 | 40 | 5.48 | ↑2.4 | 0.0054 |
| 3 | P62260.1 | 14-3-3 protein epsilon | 18 | 84 | 29 | 4.86 | ↑3.6 | 0.0204 |
| 4 | NP_037150.2 | Ras-related protein Rab-3A | 6 | 49 | 25 | 4.73 | ↑1.5 | 0.0277 |
| 5 | NP_997477.1 | Lactoylglutathione lyase | 10 | 56 | 21 | 5.13 | ↑1.9 | 0.0191 |
| 6 | NP_058932.1 | Phosphatidylethanolamine-binding protein 1 | 8 | 62 | 21 | 5.47 | ↑1.8 | 0.0340 |
| 7 | NP_075211.2 | Triosephosphate isomerase | 14 | 71 | 27 | 7.37 | ↑2.3 | 0.0297 |
| 8 | NP_075211.2 | Triosephosphate isomerase | 13 | 64 | 27 | 7.59 | ↑3.1 | 0.0044 |
| 9 | NP_446090.1 | Isocitrate dehydrogenase (NAD) subunit alpha, mitochondrial precursor | 15 | 46 | 40 | 6.13 | ↑1.6 | 0.0116 |
| 10 | NP_150238.1 | Malate dehydrogenase, cytoplasmic isoform MDH1 | 10 | 50 | 37 | 6.41 | ↑2.5 | 0.0326 |
| 11 | NP_058933.2 | Ubiquitin carboxyl-terminal hydrolase isozyme L1 | 15 | 87 | 25 | 5.24 | ↑2.8 | 0.0110 |
| 12 | NP_476484.1 | Parkinson disease protein 7 homolog isoform 2 | 9 | 72 | 20 | 6.73 | ↑3.2 | 0.0352 |
| Accession Number | Protein | Subcellular Localization | Molecular Functions and Biological Processes |
|---|---|---|---|
| NP_077327.1 | Heat-shock cognate 71 kDa protein | nucleus, cell membrane, cytoplasm | chaperone, stress response, aging, transcription |
| NP_059023.1 | Guanine nucleotide-binding protein G(o) subunit alpha | cell membrane | transducer, AC-modulating G-protein-coupled receptor, signaling pathway, aging |
| P62260.1 | 14-3-3 protein epsilon | nucleus, cytoplasm | regulatory factor, signal transduction, brain development, MAPK cascade |
| NP_037150.2 | Ras-related protein Rab-3A | cell membrane | exocytosis, axonogenesis, protein transport |
| NP_997477.1 | Lactoylglutathione lyase | cytoplasm, nucleus, cell membrane | pyruvate metabolism, negative regulation of apoptosis |
| NP_058932.1 | Phosphatidylethanolamine-binding protein 1 | cytoplasm, cell membrane | protease inhibitor, aging, MAPK cascade, response to wounding, response to oxidative stress |
| NP_075211.2 | Triosephosphate isomerase | cytoplasm | glycolysis, gluconeogenesis |
| NP_446090.1 | Isocitrate dehydrogenase (NAD) subunit alpha, mitochondrial precursor | mitochondrion | Krebs cycle |
| NP_150238.1 | Malate dehydrogenase, cytoplasmic isoform MDH1 | cytoplasm | Krebs cycle |
| NP_058933.2 | Ubiquitin carboxyl-terminal hydrolase isozyme L1 | cytoplasm, endoplasmic reticulum, cell membrane | ubiquitin conjugation pathway, possible role in protecting cells under stress condition |
| NP_476484.1 | Parkinson disease protein 7 homolog isoform 2 | cell membrane, cytoplasm, nucleus, mitochondrion | chaperone, negative regulation of apoptosis, aging, response to oxidative stress, inflammatory response |
| Spot | Accession Number | Protein | Exclusive Unique Peptides | SC a (%) | MW (kDa) | pI b | Change c (Fold) | p-Value |
|---|---|---|---|---|---|---|---|---|
| 1 | NP_001004279.1 | Peptidyl-prolyl cis–trans isomerase D | 17 | 49 | 41 | 7.27 | ↑2.1 | 0.0267 |
| 2 | NP_058704.1 | Glyceraldehyde-3-phosphate dehydrogenase | 16 | 70 | 36 | 8.87 | ↑2.1 | 0.0386 |
| 3 | NP_075211.2 | Triosephosphate isomerase | 14 | 68 | 27 | 7.20 | ↑2.0 | 0.0370 |
| 4 | XP_006252159.1 | Dihydropyrimidinase-related protein 2 isoform X1 | 17 | 52 | 73 | 5.87 | ↑2.2 | 0.0216 |
| Accession Number | Protein | Subcellular Localization | Molecular Functions and Biological Processes |
|---|---|---|---|
| NP_001004279.1 | Peptidyl-prolyl cis–trans isomerase D | nucleus, cytoplasm | chaperone, negative regulation of apoptosis, protein transport, response to oxidative stress |
| NP_058704.1 | Glyceraldehyde-3-phosphate dehydrogenase | cytoplasm, nucleus | glycolysis, response to oxidative stress, apoptosis, regulation of translation |
| NP_075211.2 | Triosephosphate isomerase | cytoplasm | glycolysis, gluconeogenesis |
| XP_006252159.1 | Dihydropyrimidinase-related protein 2 isoform X1 | cytoplasm, cell membrane | developmental protein, remodeling of the cytoskeleton after injury, differentiation, neurogenesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ujcikova, H.; Robles, D.; Yue, X.; Svoboda, P.; Lee, Y.S.; Navratilova, E. Time-Dependent Changes in Protein Composition of Medial Prefrontal Cortex in Rats with Neuropathic Pain. Int. J. Mol. Sci. 2022, 23, 955. https://doi.org/10.3390/ijms23020955
Ujcikova H, Robles D, Yue X, Svoboda P, Lee YS, Navratilova E. Time-Dependent Changes in Protein Composition of Medial Prefrontal Cortex in Rats with Neuropathic Pain. International Journal of Molecular Sciences. 2022; 23(2):955. https://doi.org/10.3390/ijms23020955
Chicago/Turabian StyleUjcikova, Hana, Dagoberto Robles, Xu Yue, Petr Svoboda, Yeon Sun Lee, and Edita Navratilova. 2022. "Time-Dependent Changes in Protein Composition of Medial Prefrontal Cortex in Rats with Neuropathic Pain" International Journal of Molecular Sciences 23, no. 2: 955. https://doi.org/10.3390/ijms23020955