
Transcriptome Analysis of an Aedes albopictus Cell Line Single- and Dual-Infected with Lammi Virus and WNV


 , , and
, , and 
Abstract
:1. Introduction
2. Results
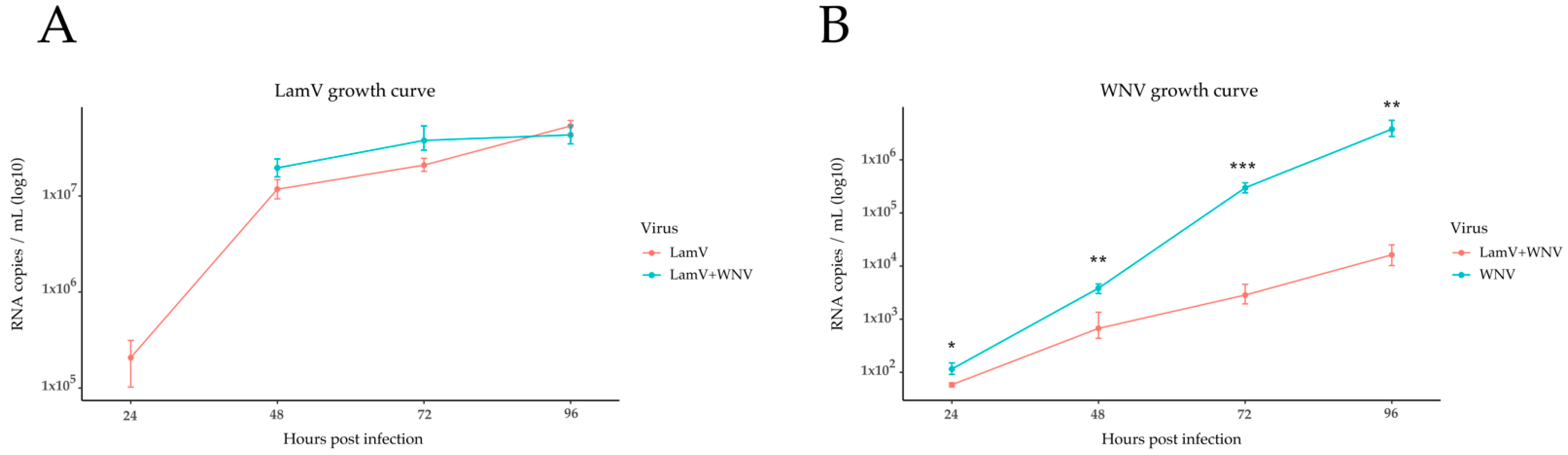
2.1. qPCR Results of Supernatant
2.2. Transcripts Differentially Expressed (DE) in LamV-Infected U4.4 Cells
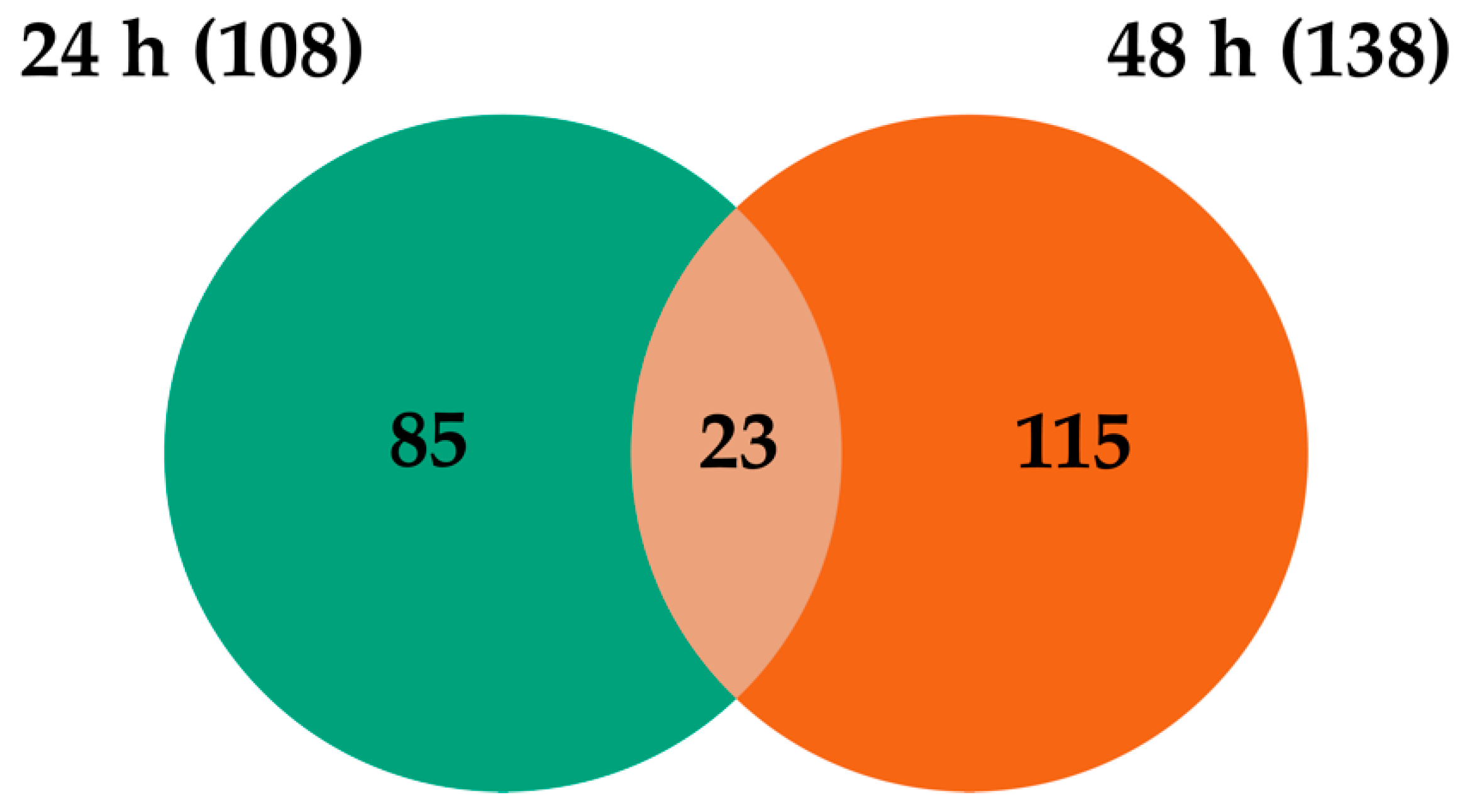
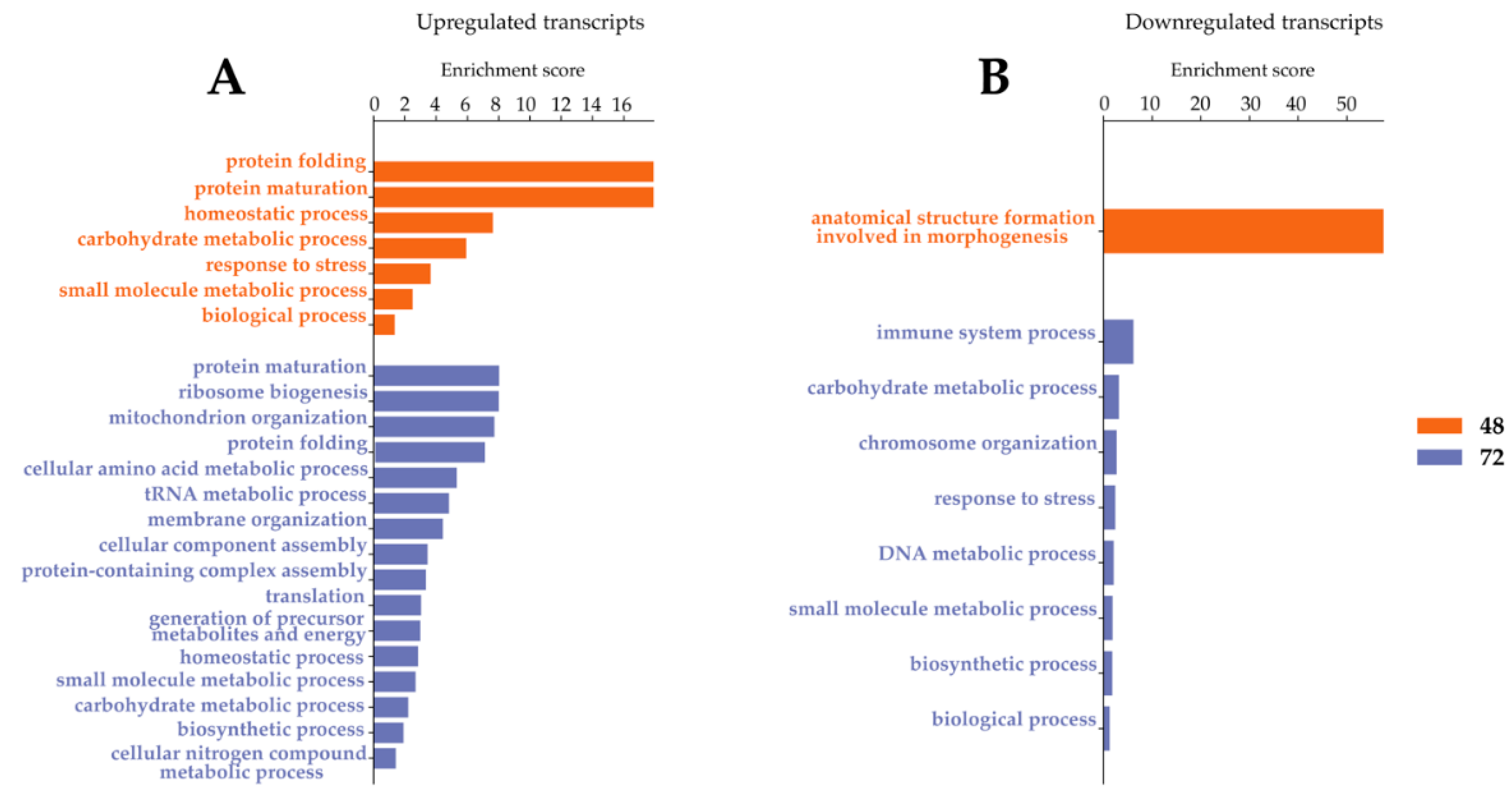
2.3. Transcripts Differentially Expressed in WNV-Infected U4.4 Cells
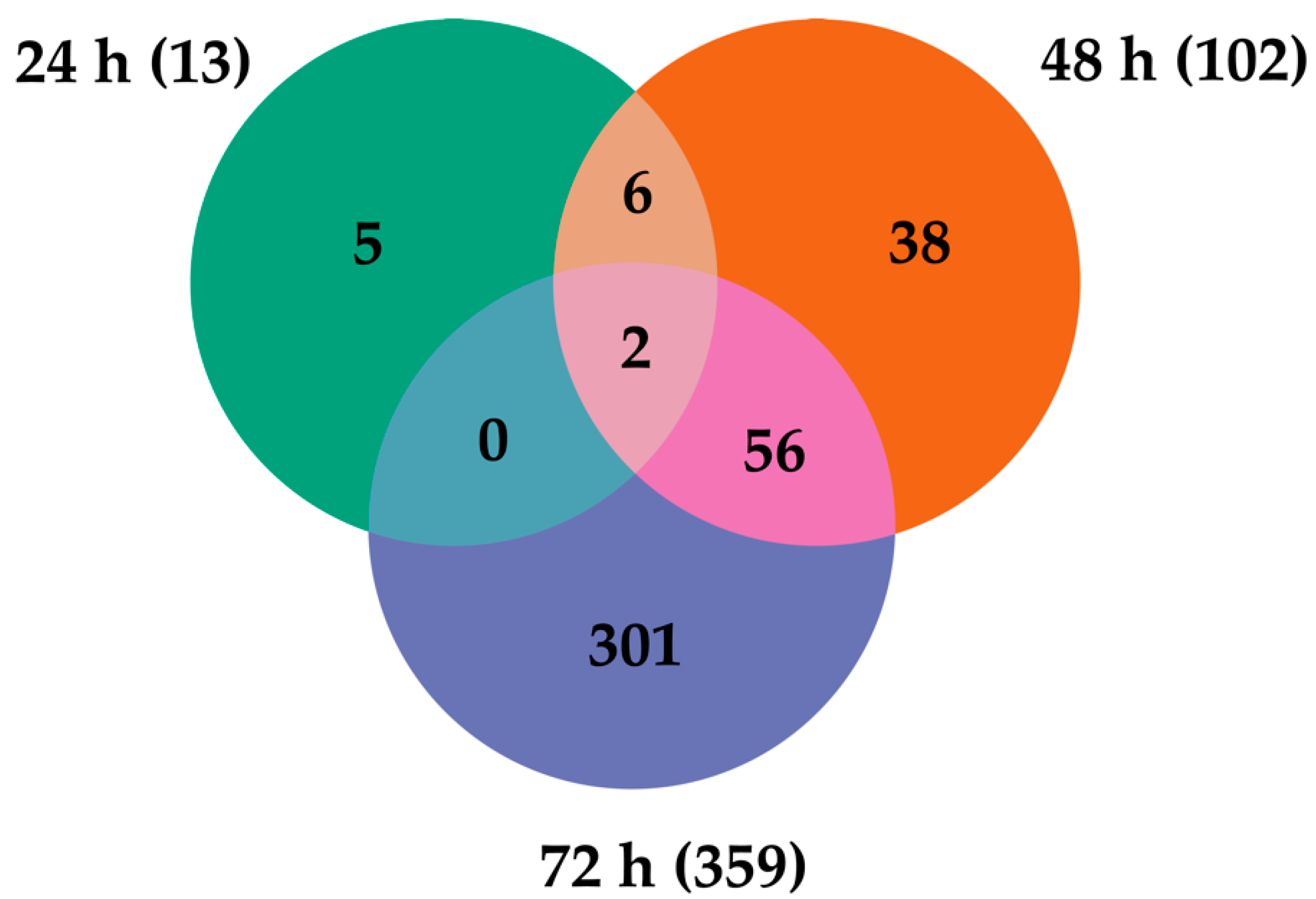
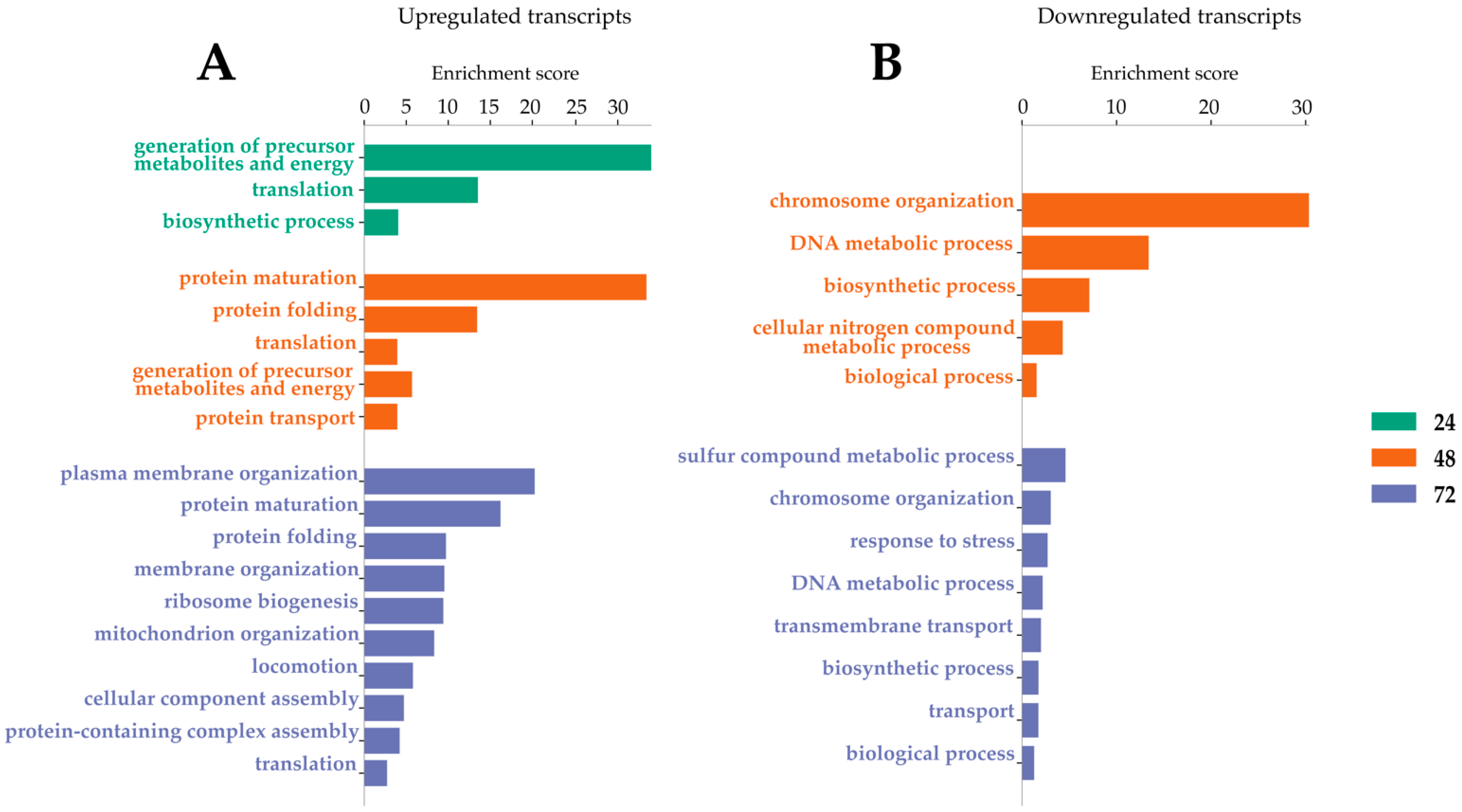
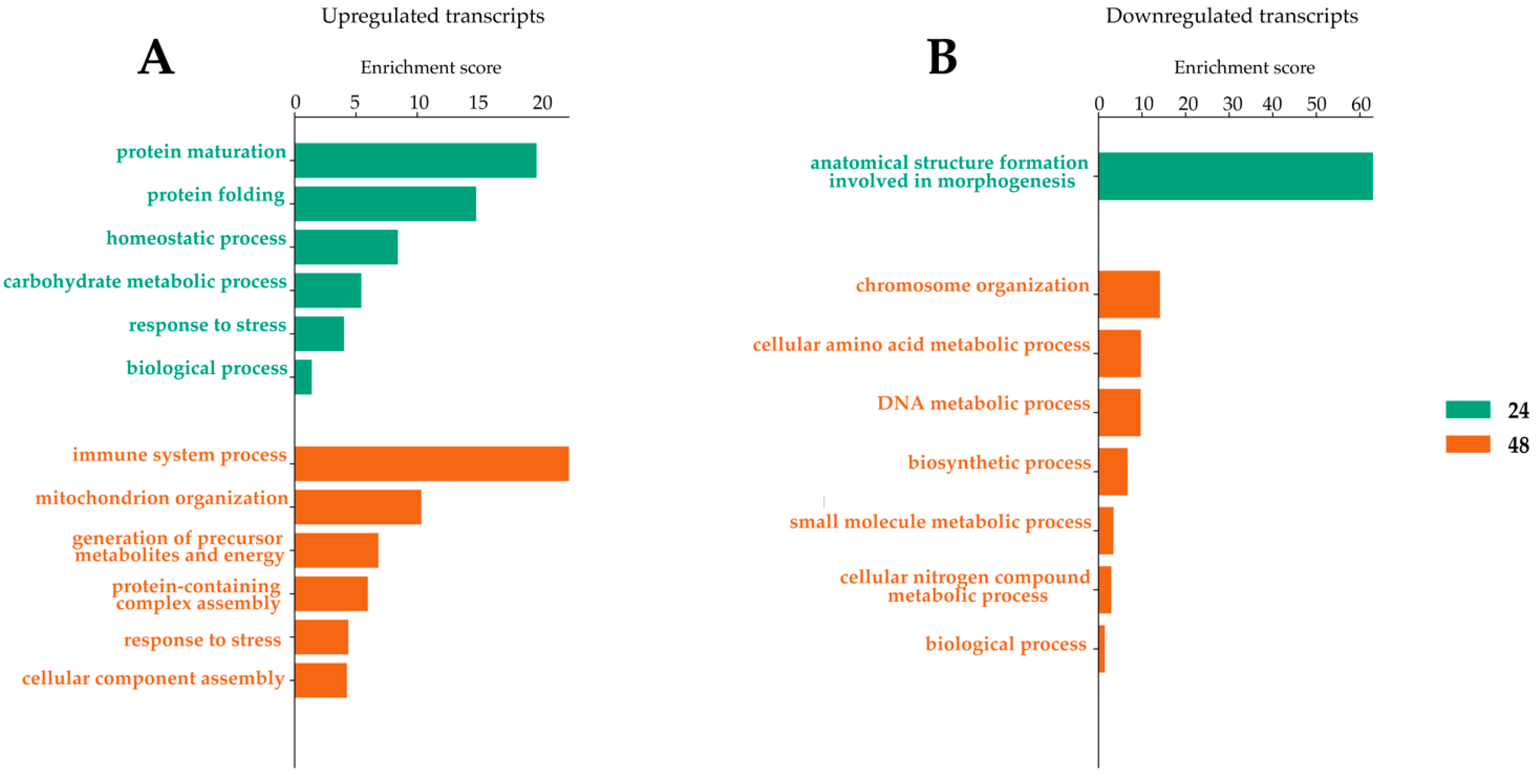
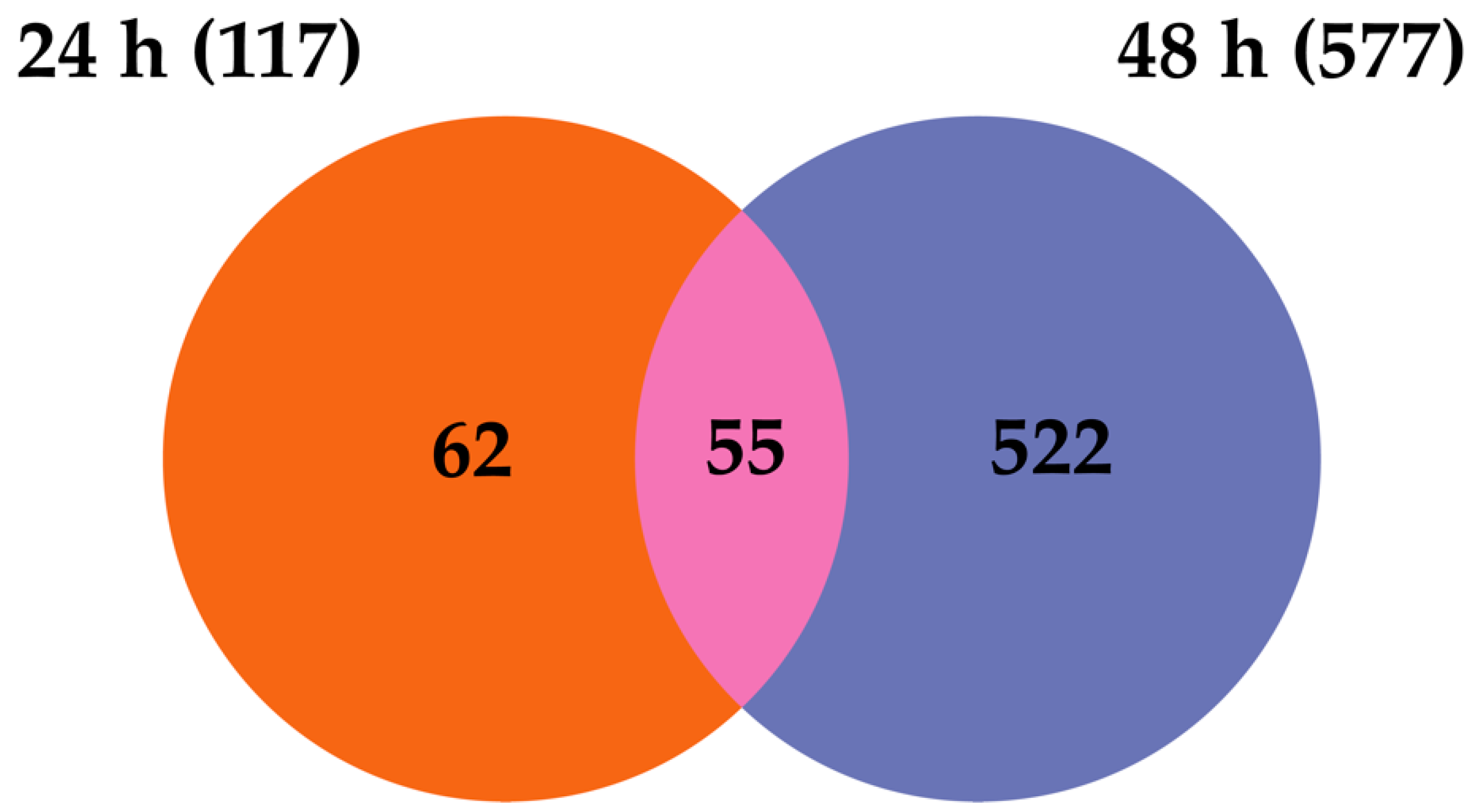
2.4. Transcripts Differentially Expressed in Dual-Infected U4.4 Cells
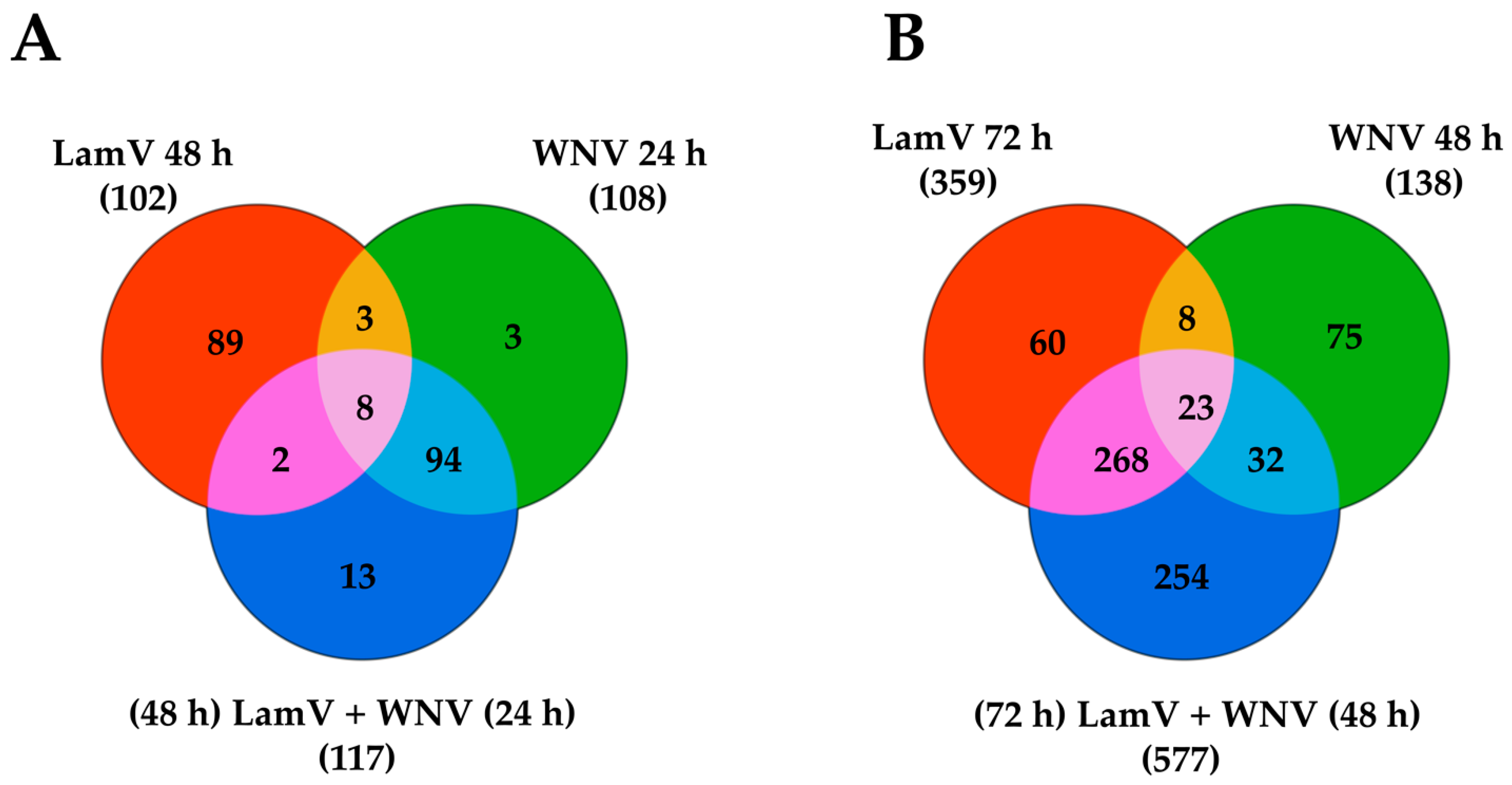
2.5. Transcripts Differentially Expressed among All Infection Groups
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Virus Stocks and Virus Titration
4.3. In Vitro Infection
4.4. Quantitative PCR
4.5. RNA Extractions
4.6. Sequencing of mRNA
4.7. RNA-Seq Data Analysis
4.8. Data Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- David, S.; Abraham, A.M. Epidemiological and clinical aspects on West Nile virus, a globally emerging pathogen. Infect. Dis. 2016, 48, 571–586. [Google Scholar] [CrossRef]
- Guo, C.; Zhou, Z.; Wen, Z.; Liu, Y.; Zeng, C.; Xiao, D.; Ou, M.; Han, Y.; Huang, S.; Liu, D.; et al. Global Epidemiology of Dengue Outbreaks in 1990-2015: A Systematic Review and Meta-Analysis. Front. Cell Infect. Microbiol. 2017, 7, 317. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, P.; Yssel, H.; Misse, D. Zika virus infection: An update. Microbes Infect. 2019, 21, 353–360. [Google Scholar] [CrossRef]
- WHO. Global Strategy for Dengue Prevention and Control, 2012–2020; WHO Report; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Luz, P.M.; Vanni, T.; Medlock, J.; Paltiel, A.D.; Galvani, A.P. Dengue vector control strategies in an urban setting: An economic modelling assessment. Lancet 2011, 377, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Sinkins, S.P.; Gould, F. Gene drive systems for insect disease vectors. Nat. Rev. Genet. 2006, 7, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Terenius, O.; Marinotti, O.; Sieglaff, D.; James, A.A. Molecular genetic manipulation of vector mosquitoes. Cell Host Microbe 2008, 4, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Yu, X.; Wang, P.; Cheng, G. Arbovirus lifecycle in mosquito: Acquisition, propagation and transmission. Expert Rev. Mol. Med. 2019, 21, e1. [Google Scholar] [CrossRef]
- Cheng, G.; Liu, Y.; Wang, P.; Xiao, X. Mosquito Defense Strategies against Viral Infection. Trends Parasitol. 2016, 32, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Srivastava, P.; Sirisena, P.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito Innate Immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.N.; Brackney, D.E.; Ebel, G.D. The role of innate immunity in conditioning mosquito susceptibility to West Nile virus. Viruses 2013, 5, 3142–3170. [Google Scholar] [CrossRef] [Green Version]
- Goic, B.; Stapleford, K.A.; Frangeul, L.; Doucet, A.J.; Gausson, V.; Blanc, H.; Schemmel-Jofre, N.; Cristofari, G.; Lambrechts, L.; Vignuzzi, M.; et al. Virus-derived DNA drives mosquito vector tolerance to arboviral infection. Nat. Commun. 2016, 7, 12410. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Webster, J.A.; Madzokere, E.T.; Stephenson, E.B.; Herrero, L.J. Mosquito antiviral defense mechanisms: A delicate balance between innate immunity and persistent viral infection. Parasit. Vectors 2019, 12, 165. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; You, M.; Rao, X.J.; Yu, X.Q. Insect C-type lectins in innate immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef]
- Dziarski, R. Peptidoglycan recognition proteins (PGRPs). Mol. Immunol. 2004, 40, 877–886. [Google Scholar] [CrossRef] [Green Version]
- Chalk, R.; Albuquerque, C.M.; Ham, P.J.; Townson, H. Full sequence and characterization of two insect defensins: Immune peptides from the mosquito Aedes aegypti. Proc. Biol. Sci. 1995, 261, 217–221. [Google Scholar] [CrossRef]
- Cho, W.L.; Fu, Y.C.; Chen, C.C.; Ho, C.M. Cloning and characterization of cDNAs encoding the antibacterial peptide, defensin A, from the mosquito, Aedes aegypti. Insect. Biochem. Mol. Biol. 1996, 26, 395–402. [Google Scholar] [CrossRef]
- Lowenberger, C.; Bulet, P.; Charlet, M.; Hetru, C.; Hodgeman, B.; Christensen, B.M.; Hoffmann, J.A. Insect immunity: Isolation of three novel inducible antibacterial defensins from the vector mosquito, Aedes aegypti. Insect. Biochem. Mol. Biol. 1995, 25, 867–873. [Google Scholar] [CrossRef]
- Xiao, X.; Liu, Y.; Zhang, X.; Wang, J.; Li, Z.; Pang, X.; Wang, P.; Cheng, G. Complement-related proteins control the flavivirus infection of Aedes aegypti by inducing antimicrobial peptides. PLoS Pathog. 2014, 10, e1004027. [Google Scholar] [CrossRef] [Green Version]
- Bartholomay, L.C.; Fuchs, J.F.; Cheng, L.L.; Beck, E.T.; Vizioli, J.; Lowenberger, C.; Christensen, B.M. Reassessing the role of defensin in the innate immune response of the mosquito, Aedes aegypti. Insect. Mol. Biol. 2004, 13, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Kanost, M.R.; Jiang, H. Clip-domain serine proteases as immune factors in insect hemolymph. Curr. Opin. Insect. Sci. 2015, 11, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.; Civana, A.; Acevedo, C.; Smartt, C.T. Transcriptomics of differential vector competence: West Nile virus infection in two populations of Culex pipiens quinquefasciatus linked to ovary development. BMC Genom. 2014, 15, 513. [Google Scholar] [CrossRef] [Green Version]
- Etebari, K.; Hegde, S.; Saldana, M.A.; Widen, S.G.; Wood, T.G.; Asgari, S.; Hughes, G.L. Global Transcriptome Analysis of Aedes aegypti Mosquitoes in Response to Zika Virus Infection. mSphere 2017, 2, e00456-17. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Xing, D.; Su, D.; Wang, D.; Gao, H.; Lan, C.; Gu, Z.; Zhao, T.; Li, C. Transcriptome Analysis of Responses to Dengue Virus 2 Infection in Aedes albopictus (Skuse) C6/36 Cells. Viruses 2021, 13, 343. [Google Scholar] [CrossRef] [PubMed]
- Li, M.J.; Lan, C.J.; Gao, H.T.; Xing, D.; Gu, Z.Y.; Su, D.; Zhao, T.Y.; Yang, H.Y.; Li, C.X. Transcriptome analysis of Aedes aegypti Aag2 cells in response to dengue virus-2 infection. Parasit. Vectors 2020, 13, 421. [Google Scholar] [CrossRef] [PubMed]
- Shrinet, J.; Srivastava, P.; Sunil, S. Transcriptome analysis of Aedes aegypti in response to mono-infections and co-infections of dengue virus-2 and chikungunya virus. Biochem. Biophys. Res. Commun. 2017, 492, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Behura, S.K.; Franz, A.W.E. The midgut transcriptome of Aedes aegypti fed with saline or protein meals containing chikungunya virus reveals genes potentially involved in viral midgut escape. BMC Genom. 2017, 18, 382. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Alto, B.W.; Jiang, Y.; Yu, F.; Zhang, Y. Transcriptomic Analysis of Aedes aegypti Innate Immune System in Response to Ingestion of Chikungunya Virus. Int. J. Mol. Sci. 2019, 20, 3133. [Google Scholar] [CrossRef] [Green Version]
- Ohlund, P.; Lunden, H.; Blomstrom, A.L. Insect-specific virus evolution and potential effects on vector competence. Virus Genes 2019, 55, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Nasar, F.; Gorchakov, R.V.; Tesh, R.B.; Weaver, S.C. Eilat virus host range restriction is present at multiple levels of the virus life cycle. J. Virol. 2015, 89, 1404–1418. [Google Scholar] [CrossRef] [Green Version]
- Saiyasombat, R.; Bolling, B.G.; Brault, A.C.; Bartholomay, L.C.; Blitvich, B.J. Evidence of efficient transovarial transmission of Culex flavivirus by Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2011, 48, 1031–1038. [Google Scholar] [CrossRef]
- Bolling, B.G.; Eisen, L.; Moore, C.G.; Blair, C.D. Insect-specific flaviviruses from Culex mosquitoes in Colorado, with evidence of vertical transmission. Am. J. Trop. Med. Hyg. 2011, 85, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddow, A.D.; Guzman, H.; Popov, V.L.; Wood, T.G.; Widen, S.G.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. First isolation of Aedes flavivirus in the Western Hemisphere and evidence of vertical transmission in the mosquito Aedes (Stegomyia) albopictus (Diptera: Culicidae). Virology 2013, 440, 134–139. [Google Scholar] [CrossRef] [Green Version]
- Marklewitz, M.; Zirkel, F.; Kurth, A.; Drosten, C.; Junglen, S. Evolutionary and phenotypic analysis of live virus isolates suggests arthropod origin of a pathogenic RNA virus family. Proc. Natl. Acad. Sci. USA 2015, 112, 7536–7541. [Google Scholar] [CrossRef] [Green Version]
- Hobson-Peters, J.; Yam, A.W.; Lu, J.W.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; et al. A new insect-specific flavivirus from northern Australia suppresses replication of West Nile virus and Murray Valley encephalitis virus in co-infected mosquito cells. PLoS ONE 2013, 8, e56534. [Google Scholar] [CrossRef]
- Hall-Mendelin, S.; McLean, B.J.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Hall, R.A.; van den Hurk, A.F. The insect-specific Palm Creek virus modulates West Nile virus infection in and transmission by Australian mosquitoes. Parasit. Vectors 2016, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Kenney, J.L.; Solberg, O.D.; Langevin, S.A.; Brault, A.C. Characterization of a novel insect-specific flavivirus from Brazil: Potential for inhibition of infection of arthropod cells with medically important flaviviruses. J. Gen. Virol. 2014, 95, 2796–2808. [Google Scholar] [CrossRef]
- Goenaga, S.; Kenney, J.L.; Duggal, N.K.; Delorey, M.; Ebel, G.D.; Zhang, B.; Levis, S.C.; Enria, D.A.; Brault, A.C. Potential for Co-Infection of a Mosquito-Specific Flavivirus, Nhumirim Virus, to Block West Nile Virus Transmission in Mosquitoes. Viruses 2015, 7, 5801–5812. [Google Scholar] [CrossRef] [Green Version]
- Huhtamo, E.; Putkuri, N.; Kurkela, S.; Manni, T.; Vaheri, A.; Vapalahti, O.; Uzcategui, N.Y. Characterization of a novel flavivirus from mosquitoes in northern europe that is related to mosquito-borne flaviviruses of the tropics. J. Virol. 2009, 83, 9532–9540. [Google Scholar] [CrossRef] [Green Version]
- Blitvich, B.J.; Firth, A.E. Insect-specific flaviviruses: A systematic review of their discovery, host range, mode of transmission, superinfection exclusion potential and genomic organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlund, P.; Hayer, J.; Hesson, J.C.; Blomstrom, A.L. Small RNA Response to Infection of the Insect-Specific Lammi Virus and Hanko Virus in an Aedes albopictus Cell Line. Viruses 2021, 13, 2181. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Pridgeon, J.W.; Becnel, J.J.; Clark, G.G.; Linthicum, K.J. Identification of genes differentially expressed during heat shock treatment in Aedes aegypti. J. Med. Entomol. 2009, 46, 490–495. [Google Scholar] [CrossRef]
- Randow, F.; Seed, B. Endoplasmic reticulum chaperone gp96 is required for innate immunity but not cell viability. Nat. Cell Biol. 2001, 3, 891–896. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.; Dai, J.; Srivastava, P.K.; Zammit, D.J.; Lefrancois, L.; Li, Z. Heat shock protein gp96 is a master chaperone for toll-like receptors and is important in the innate function of macrophages. Immunity 2007, 26, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Runtuwene, L.R.; Kawashima, S.; Pijoh, V.D.; Tuda, J.S.B.; Hayashida, K.; Yamagishi, J.; Sugimoto, C.; Nishiyama, S.; Sasaki, M.; Orba, Y.; et al. The Lethal(2)-Essential-for-Life [L(2)EFL] Gene Family Modulates Dengue Virus Infection in Aedes aegypti. Int. J. Mol. Sci. 2020, 21, 7520. [Google Scholar] [CrossRef]
- Colpitts, T.M.; Barthel, S.; Wang, P.; Fikrig, E. Dengue virus capsid protein binds core histones and inhibits nucleosome formation in human liver cells. PLoS ONE 2011, 6, e24365. [Google Scholar] [CrossRef] [Green Version]
- Horton, A.A.; Wang, B.; Camp, L.; Price, M.S.; Arshi, A.; Nagy, M.; Nadler, S.A.; Faeder, J.R.; Luckhart, S. The mitogen-activated protein kinome from Anopheles gambiae: Identification, phylogeny and functional characterization of the ERK, JNK and p38 MAP kinases. BMC Genom. 2011, 12, 574. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Miner, J.J.; Gorman, M.J.; Rausch, K.; Ramage, H.; White, J.P.; Zuiani, A.; Zhang, P.; Fernandez, E.; Zhang, Q.; et al. A CRISPR screen defines a signal peptide processing pathway required by flaviviruses. Nature 2016, 535, 164–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brien, J.D.; Lazear, H.M.; Diamond, M.S. Propagation, quantification, detection, and storage of West Nile virus. Curr. Protoc. Microbiol. 2013, 31, 15D-3. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [CrossRef] [Green Version]
- Ewels, P.A.; Peltzer, A.; Fillinger, S.; Patel, H.; Alneberg, J.; Wilm, A.; Garcia, M.U.; Di Tommaso, P.; Nahnsen, S. The nf-core framework for community-curated bioinformatics pipelines. Nat. Biotechnol. 2020, 38, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Babraham Bioinformatics. FastQC. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 8 December 2021).
- Babraham Bioinformatics. Trim Galore. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 8 December 2021).
- Kopylova, E.; Noe, L.; Touzet, H. SortMeRNA: Fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo-Calderon, G.I.; Emrich, S.J.; MacCallum, R.M.; Maslen, G.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; VectorBase, C.; Madey, G.; et al. VectorBase: An updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 2015, 43, D707–D713. [Google Scholar] [CrossRef] [PubMed]
- Schurch, N.J.; Schofield, P.; Gierlinski, M.; Cole, C.; Sherstnev, A.; Singh, V.; Wrobel, N.; Gharbi, K.; Simpson, G.G.; Owen-Hughes, T.; et al. How many biological replicates are needed in an RNA-seq experiment and which differential expression tool should you use? RNA 2016, 22, 839–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript ID | Functional Categories | Gene Description | Fold Change 24 hpi | Fold Change 48 hpi |
|---|---|---|---|---|
| AALF016234 | Immune response | C-type lectin | 82.89 | 17.75 |
| AALF013298 | Unspecified product | 69.1 | 14.16 | |
| AALF003774 | Immune response | Fibrinogen and fibronectin | 52.23 | 18.48 |
| AALF019859 | Immune response | Clip-Domain Serine Protease family B | 49.51 | 9.26 |
| AALF008821 | Immune response | Defensin antimicrobial peptide | 45.85 | 57.72 |
| AALF026731 | Unspecified product | 22.22 | 98.88 | |
| AALF008229 | Unspecified product | 19.82 | 7.04 | |
| AALF025212 | Ion homeostasis | Transferrin | 15.95 | 20.69 |
| AALF020799 | Immune response | Peptidoglycan-Recognition Protein | 14.48 | 10.23 |
| AALF001195 | Unspecified product | 12.68 | 9.79 | |
| AALF008452 | Unspecified product | 12.07 | 10.83 | |
| AALF002418 | Metabolic process | Imaginal disc growth factor | 10.45 | 10.37 |
| AALF019963 | Unspecified product | 9.01 | 4.88 | |
| AALF009200 | Unspecified product | 8.65 | 5.95 | |
| AALF005588 | Metabolic process | L-lactate dehydrogenase | 8.11 | 7.45 |
| AALF004727 | Metabolic process | Lipase | 7.49 | 5.5 |
| AALF012590 | Unspecified product | 7.38 | 4.56 | |
| AALF015711 | DNA replication | DNA helicase | 6.43 | −12.12 |
| AALF022735 | DNA replication | DNA helicase | 5.48 | −13.63 |
| AALF009419 | Unspecified product | 5.14 | 4.47 | |
| AALF028505 | Unspecified product | −13.71 | −12.83 | |
| AALF008879 | Metabolic process | Type IV inositol 5-phosphatase | −16.64 | −10.21 |
| AALF015015 | Unspecified product | −230.7 | 22.72 |
| Transcript ID | Functional Categories | Gene Description | Fold Change 24 hpi | Fold Change 48 hpi |
|---|---|---|---|---|
| AALF016234 | Immune response | C-type lectin | 82.76 | 22.12 |
| AALF013298 | Unspecified product | 69.07 | 23.19 | |
| AALF003774 | Immune response | Fibrinogen and fibronectin | 52.19 | 9.14 |
| AALF019859 | Immune response | Clip-Domain Serine Protease family B | 49.47 | 7.65 |
| AALF008821 | Immune response | Defensin antimicrobial peptide | 45.81 | 16.18 |
| AALF026731 | Unspecified product | 22.2 | 12.95 | |
| AALF020197 | Immune response | Clip-Domain Serine Protease family B | 15.24 | −6.49 |
| AALF020799 | Immune response | Peptidoglycan-Recognition Protein | 14.47 | 8.03 |
| AALF007525 | Unspecified product | 12.61 | 23.73 | |
| AALF016365 | Granules fusion | Munc13-4 | 10.03 | 6.05 |
| AALF004571 | Unspecified product | 9.85 | 122.64 | |
| AALF012716 | Immune response | Prophenoloxidase | 9.77 | −5.54 |
| AALF006247 | Signal peptide processing | Putative microsomal signal peptidase 25 kDa subunit | 8.72 | 20.65 |
| AALF021835 | Response to stress | BiP/GRP78 | 8.37 | 31.7 |
| AALF005588 | Metabolic process | L-lactate dehydrogenase | 8.1 | 28.92 |
| AALF019952 | Unspecified product | 7.65 | 6.1 | |
| AALF003990 | Metabolic process | Mannosyltransferase | 7.13 | 88.65 |
| AALF027716 | Metabolic process | Cytochrome P450 | 6.64 | 9.16 |
| AALF003192 | Signal peptide processing | Signal peptidase complex subunit 3 | 6.58 | 20.26 |
| AALF012257 | Peptidylprolyl isomerase | 6.55 | 7.31 | |
| AALF019397 | Putative reticulocalbin calumenin DNA supercoiling factor | 5.97 | −4.99 | |
| AALF022020 | Response to stress | Chaperonin-60kD | 5.38 | 6.94 |
| AALF002466 | Response to stress | Protein disulfide-isomerase A6 precursor | 5.2 | 11.04 |
| AALF011939 | Response to stress | Endoplasmin | 5.12 | 12.98 |
| AALF002534 | Response to stress | Putative heat-shock protein | 5.08 | 8.67 |
| AALF020666 | Protein degradation | Ubiquitin | −4.79 | −4.99 |
| AALF026911 | GPCR Orphan/Putative Class B Family | −7.02 | −25.17 | |
| AALF016818 | Unspecified product | −8.1 | −5.85 | |
| AALF025231 | Unspecified product | −9.13 | −20.68 | |
| AALF028121 | Unspecified product | −9.15 | −7.13 | |
| AALF015500 | DNA/RNA binding | Zinc finger protein | −9.41 | −7.58 |
| AALF008709 | Transport | Mfs transporter | −10.8 | −7.4 |
| AALF011706 | Unspecified product | −10.79 | −26.44 | |
| AALF028505 | Unspecified product | −13.72 | −65.33 | |
| AALF020168 | Unspecified product | −13.93 | −22.6 | |
| AALF008099 | Endothelin-converting enzyme | −15.39 | −66.8 | |
| AALF000742 | Unspecified product | −15.81 | −54.52 | |
| AALF008879 | Metabolic process | Type IV inositol 5-phosphatase | −16.65 | −60.51 |
| AALF004114 | No-mechanoreceptor potential a | −24.69 | −45.32 | |
| AALF011390 | Putative ecdysone-induced protein | −26.61 | −21.06 | |
| AALF020798 | Unspecified product | −29.45 | −10.58 | |
| AALF009909 | Unspecified product | −32.33 | −140.61 | |
| AALF001259 | Unspecified product | −36.13 | −36.84 | |
| AALF012770 | Metabolic process | Aldehyde oxidase | −37.85 | −39.12 |
| AALF001105 | Unspecified product | −40.91 | −26.97 | |
| AALF002857 | Unspecified product | −42.81 | −133.32 | |
| AALF002636 | Unspecified product | −43.27 | −11.27 | |
| AALF025810 | Unspecified product | −51.86 | −28.39 | |
| AALF013937 | Immune-related | Serine protease | −52.04 | −60.31 |
| AALF014375 | Unspecified product | −57.5 | −72.13 | |
| AALF013936 | Unspecified product | −78.5 | −156.98 | |
| AALF006472 | Unspecified product | −86.08 | −193.34 | |
| AALF015014 | Immune-related | Clip-Domain Serine Protease family D | −245.49 | −75.47 |
| AALF016295 | Unspecified product | −249.88 | −269.15 | |
| AALF014395 | Unspecified product | −605.75 | −300.78 |
| Transcript ID | Functional Categories | Gene Description | Fold Change 48 hpi with LamV | Fold Change 24 hpi with WNV | Fold Change Dual Infection 24 hpi with WNV |
|---|---|---|---|---|---|
| AALF004571 | DUF3456 domain-containing protein | 55.17 | 9.85 | 9.86 | |
| AALF021835 | Response to stress | BiP/GRP78 | 21.95 | 8.37 | 8.37 |
| AALF006247 | Signal peptide processing | Putative microsomal signal peptidase 25 kDa subunit | 19.29 | 8.73 | 8.72 |
| AALF003192 | Signal peptide processing | Signal peptidase complex subunit 3 | 15.78 | 6.58 | 6.58 |
| AALF011939 | Response to stress | Endoplasmin | 11.87 | 5.12 | 5.12 |
| AALF002466 | Response to stress | protein disulfide-isomerase A6 precursor | 8.33 | 5.2 | 5.2 |
| AALF009419 | Unspecified product | −4.9 | 5.14 | 5.13 | |
| AALF012716 | Immune response | Prophenoloxidase | −12.04 | 9.78 | 9.77 |
| Transcript ID | Functional Categories | Gene Description | Fold Change 72 hpi with LamV | Fold Change 48 hpi with WNV | Fold Change Dual Infection 48 hpi with WNV |
|---|---|---|---|---|---|
| AALF010887 | Unspecified product | 9.29 | 23.96 | 13.02 | |
| AALF020693 | Unspecified product | 27.96 | 16.03 | 40.21 | |
| AALF019136 | Unspecified product | 6.81 | 14.5 | 10.28 | |
| AALF014826 | Unspecified product | 12.18 | 11.12 | 22.82 | |
| AALF007397 | Unspecified product | 8.27 | 7.88 | 9.94 | |
| AALF022151 | Immune response | C-type lectin domain-containing protein | 7.5 | 7.47 | 7.91 |
| AALF005588 | Metabolic process | L-lactate dehydrogenase | 8.74 | 7.45 | 28.92 |
| AALF017030 | Unspecified product | 9.79 | 6.86 | 14.3 | |
| AALF007472 | Unspecified product | 7.8 | 6.68 | 13.9 | |
| AALF004337 | Unspecified product | 5.06 | 5.93 | 6.77 | |
| AALF003871 | Metabolic process | Pyrroline-5-carboxylate reductase | 14.48 | 5.21 | 24.79 |
| AALF010275 | Fibrinogen C-terminal domain-containing protein | −6.44 | 5.05 | −6.27 | |
| AALF015929 | DNA replication | Ribonucleoside-diphosphate reductase | −7.38 | −6.52 | −9.17 |
| AALF024232 | DNA replication | Ribonucleoside-diphosphate reductase | −7.4 | −7.42 | −9.81 |
| AALF015599 | DNA replication | DNA helicase | −6.85 | −10.3 | −8.42 |
| AALF020651 | DNA replication | DNA helicase | −5.72 | −10.51 | −8.23 |
| AALF000130 | dNK domain-containing protein | −12.04 | −12.54 | −16.06 | |
| AALF019880 | Oxidationreduction process | Dihydropyrimidine dehydrogenase NADP(+) | −14.35 | −12.85 | −35.91 |
| AALF013610 | Oxidationreduction process | Dihydropyrimidine dehydrogenase NADP(+) | −16.22 | −13.14 | −42.04 |
| AALF022735 | DNA replication | DNA helicase | −5.24 | −13.63 | −8.08 |
| AALF013129 | Metabolic process | Trehalose-6-phosphate synthase | −15.42 | −14.88 | −23.66 |
| AALF006549 | DNA replication | DNA replication licensing factor MCM7 | −6.56 | −15.21 | −9.89 |
| AALF000129 | Reverse transcription | Reverse transcriptase domain-containing protein | −21.58 | −20 | −15.36 |
| Primers | Binding Site | Sequence (5′ → 3′) | Ref |
|---|---|---|---|
| qLamV-F | 4659–4678 | TGGGTGTTACCGGGTTATGT | FJ606789 |
| qLamV-R | 4845–4864 | ACGTTCCATTCAGTTTCCAT | |
| qWNV-F | 10490–10508 | GAAGTCAGGCCGGAAAGTT | AF260968 |
| qWNV-R | 10668–10689 | TCTCCGCAGAGTGGCACGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Öhlund, P.; Delhomme, N.; Hayer, J.; Hesson, J.C.; Blomström, A.-L. Transcriptome Analysis of an Aedes albopictus Cell Line Single- and Dual-Infected with Lammi Virus and WNV. Int. J. Mol. Sci. 2022, 23, 875. https://doi.org/10.3390/ijms23020875
Öhlund P, Delhomme N, Hayer J, Hesson JC, Blomström A-L. Transcriptome Analysis of an Aedes albopictus Cell Line Single- and Dual-Infected with Lammi Virus and WNV. International Journal of Molecular Sciences. 2022; 23(2):875. https://doi.org/10.3390/ijms23020875
Chicago/Turabian StyleÖhlund, Pontus, Nicolas Delhomme, Juliette Hayer, Jenny C. Hesson, and Anne-Lie Blomström. 2022. "Transcriptome Analysis of an Aedes albopictus Cell Line Single- and Dual-Infected with Lammi Virus and WNV" International Journal of Molecular Sciences 23, no. 2: 875. https://doi.org/10.3390/ijms23020875