
Indigo Pulverata Levis (Chung-Dae, Persicaria tinctoria) Alleviates Atopic Dermatitis-like Inflammatory Responses In Vivo and In Vitro



Abstract
:1. Introduction
2. Results
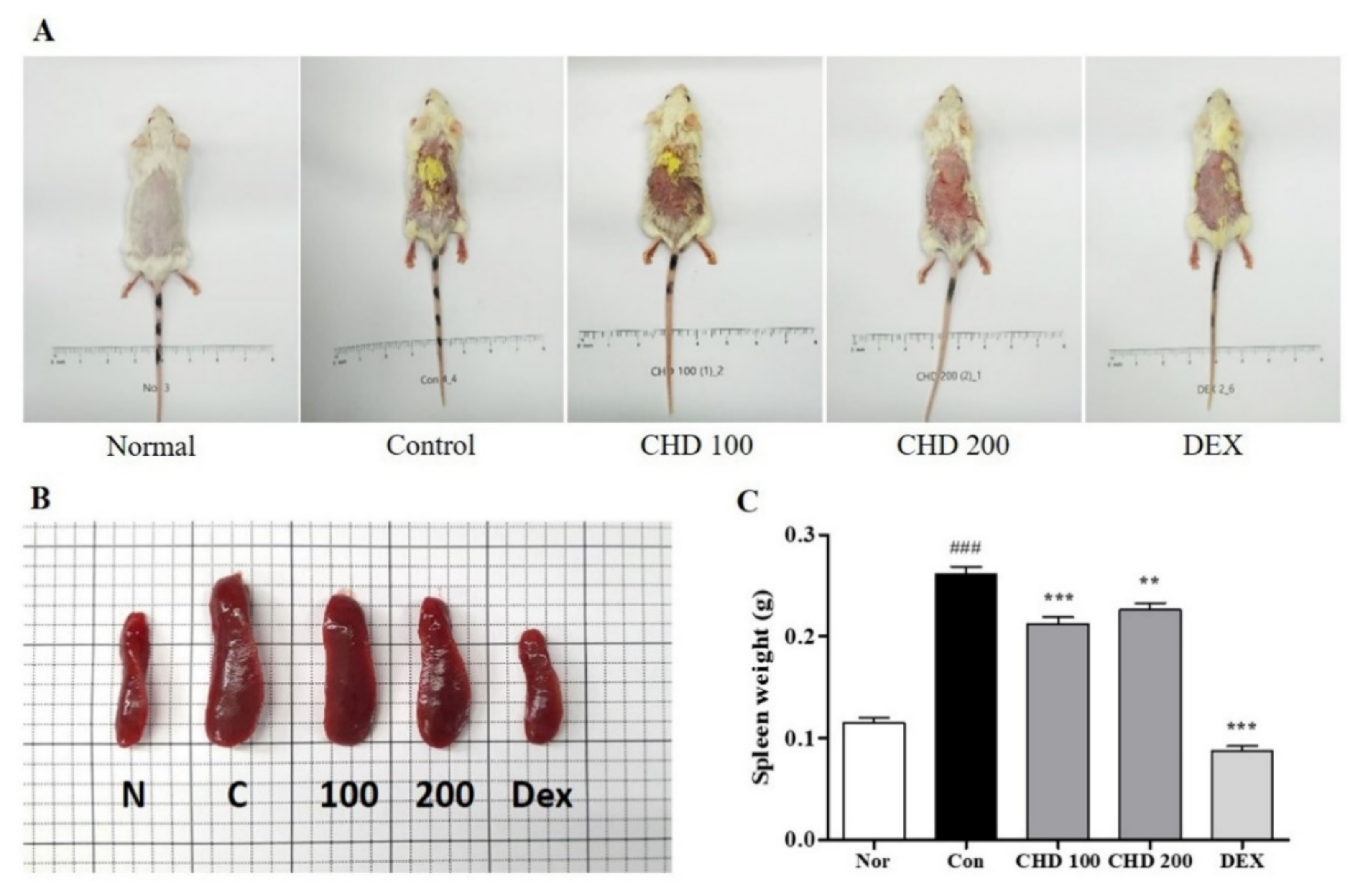
2.1. Effects of CHD on AD Symptoms in Mice
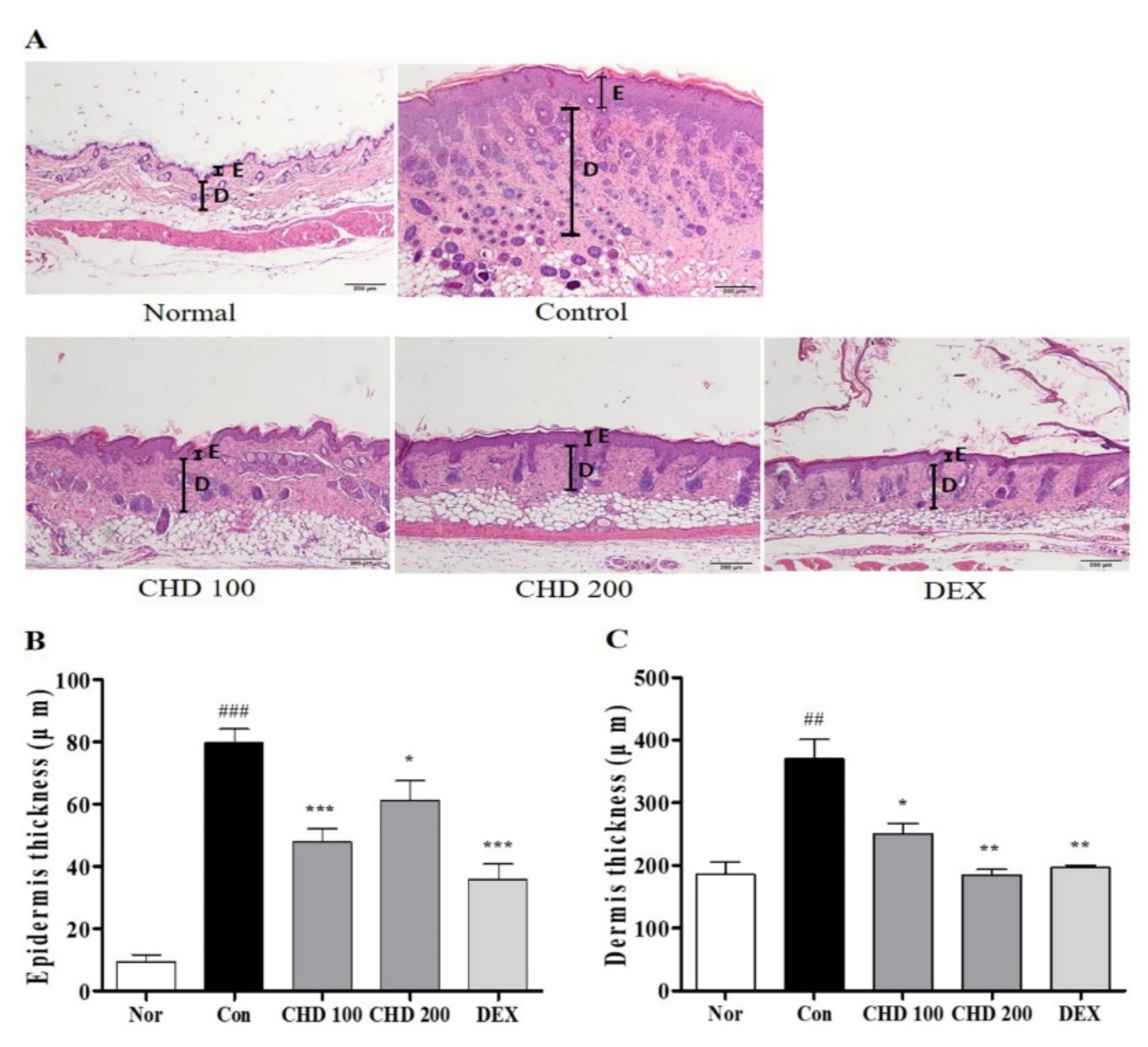
2.2. Effects of CHD on Epidermis and Dermis Thicknesses in AD Mice
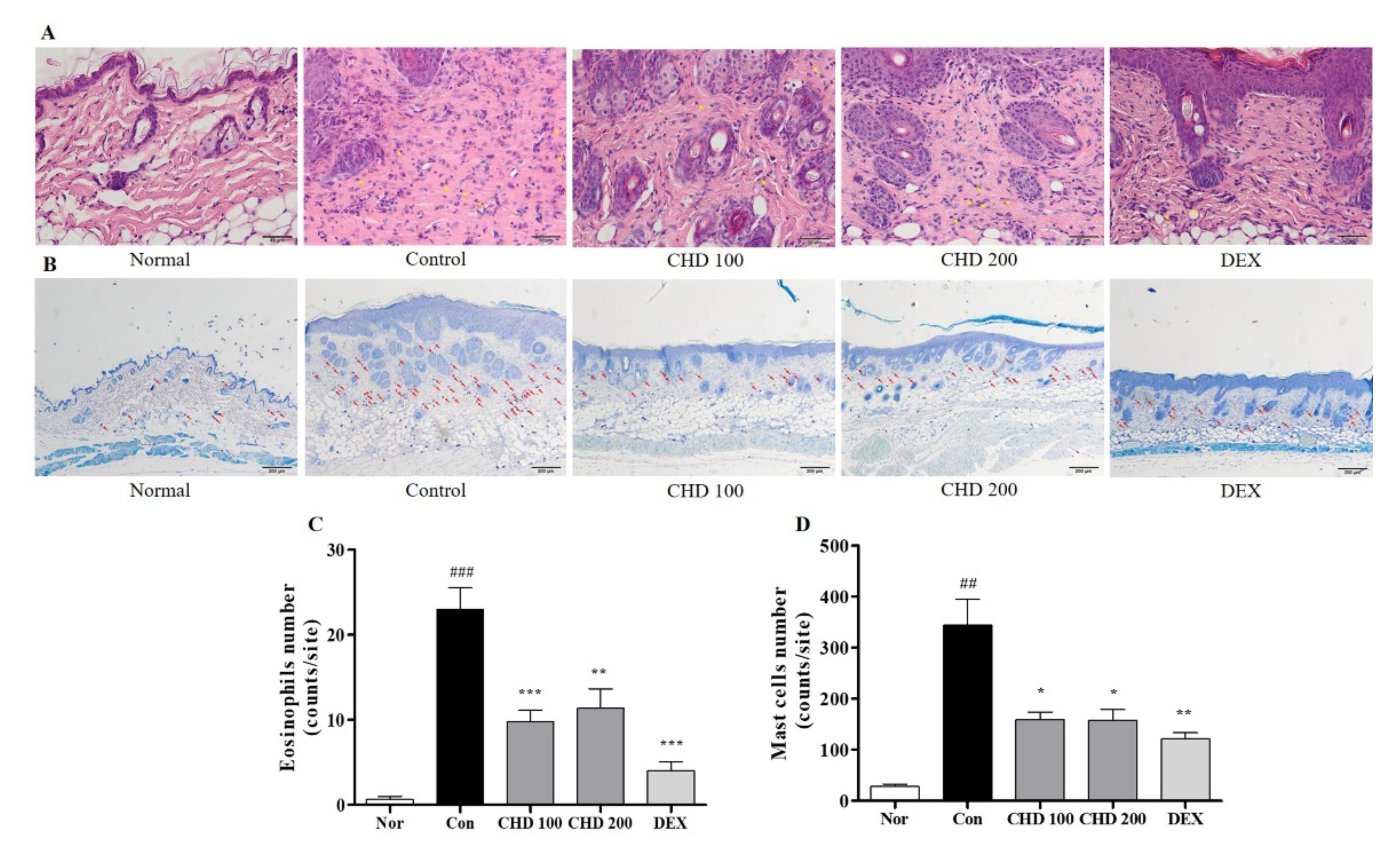
2.3. Effects of CHD on Immune Cell Infiltration in AD Mice
2.4. Effects of CHD on the Levels of IgE and Pro-Inflammatory Cytokines in AD Mice
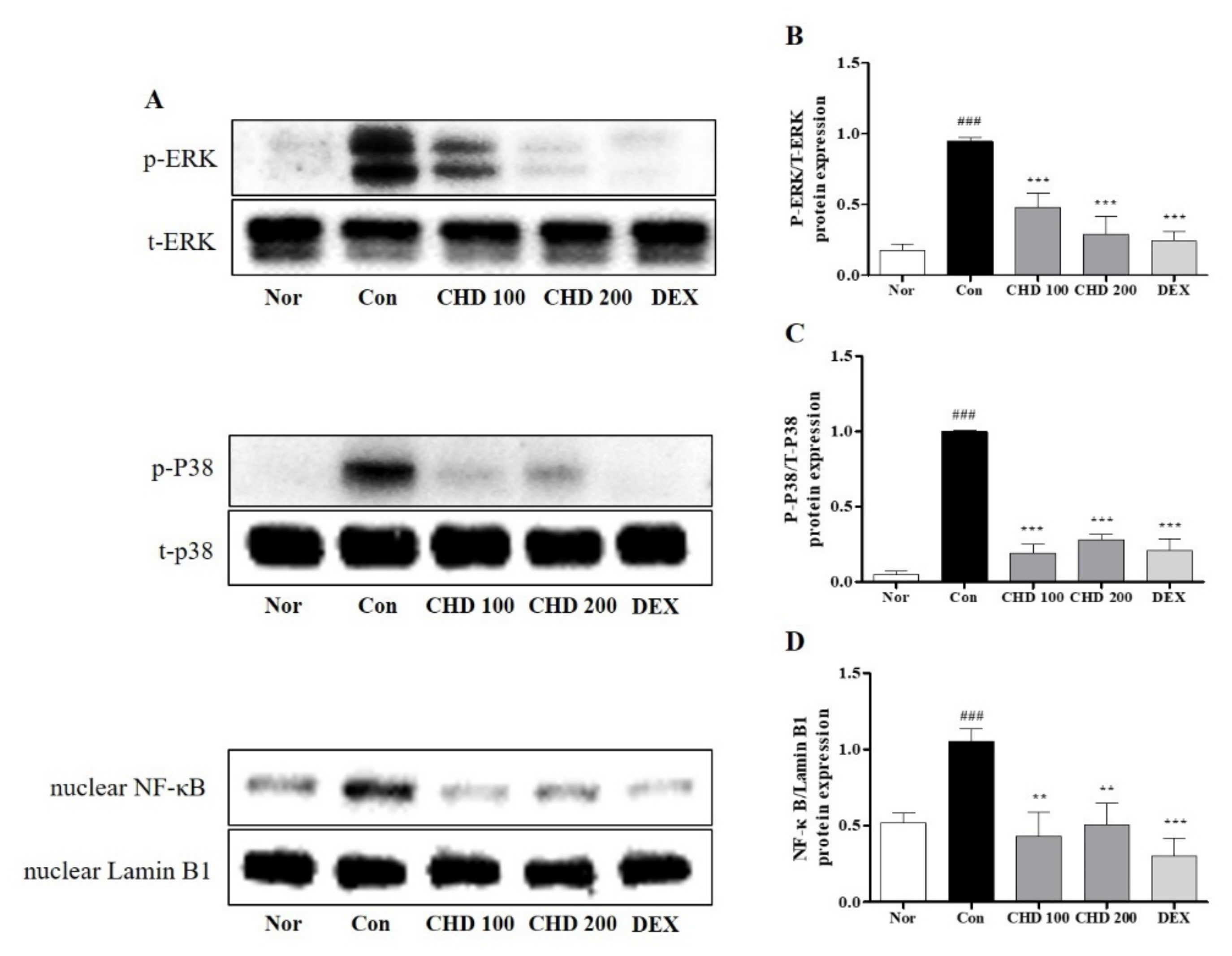
2.5. Effects of CHD on the MAPK and NF-κB Protein Levels in AD Mice
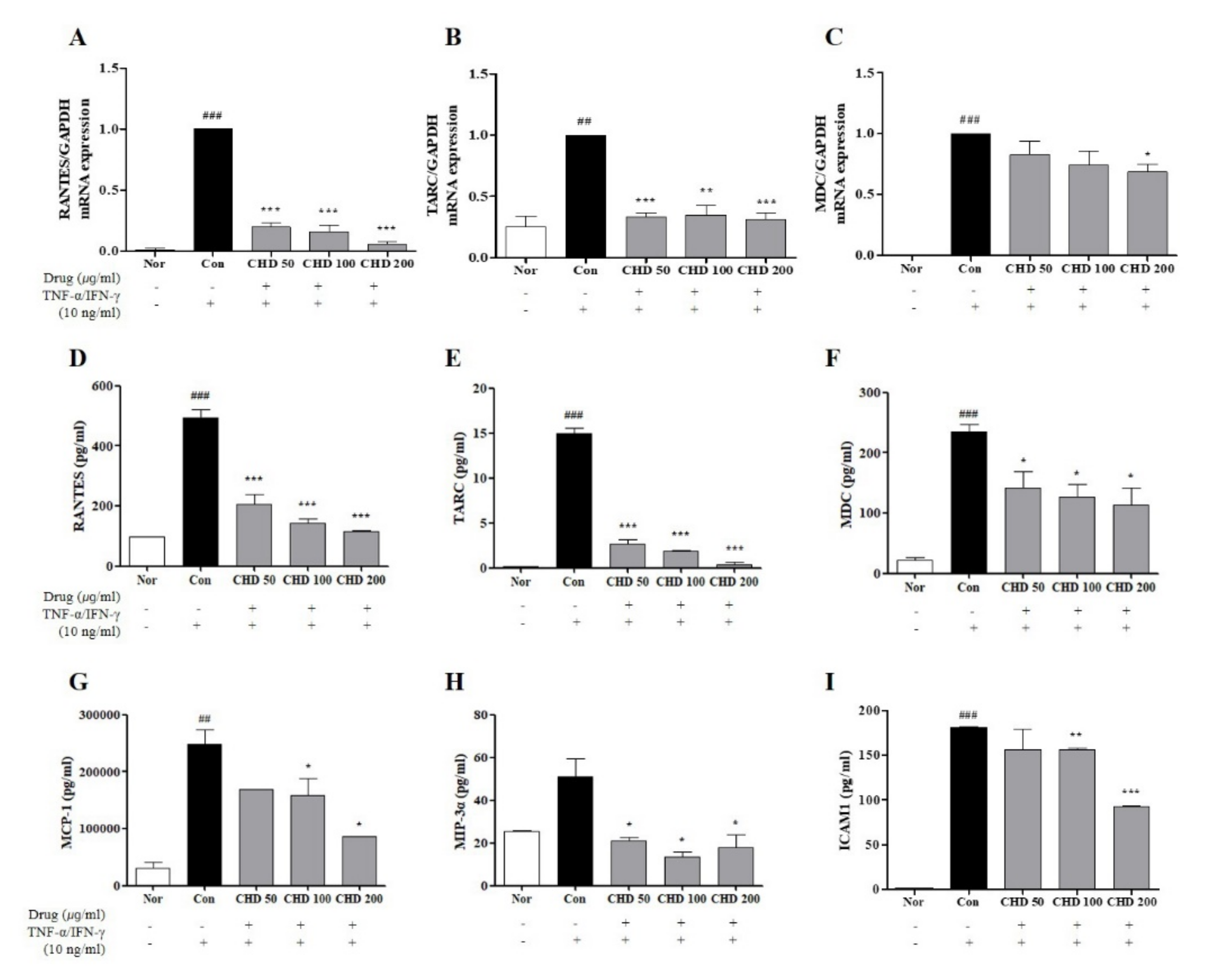
2.6. Effects of CHD on the Chemokine Expression Levels in TNF-α- and IFN-γ-Stimulated HaCaT Cells
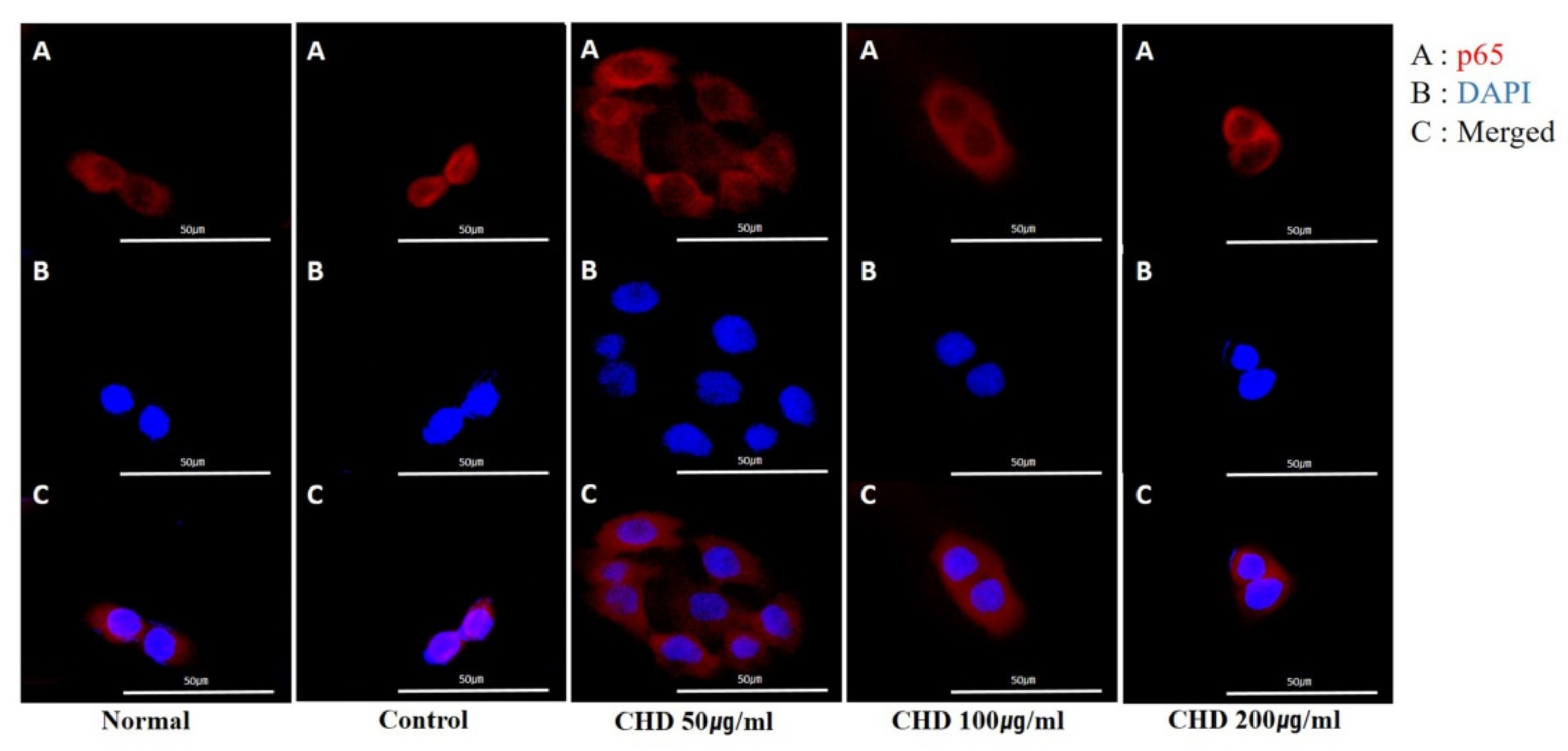
2.7. Effects of CHD on NF-κB p65 Translocation in TNF-α- and IFN-γ -Stimulated HaCaT Cells
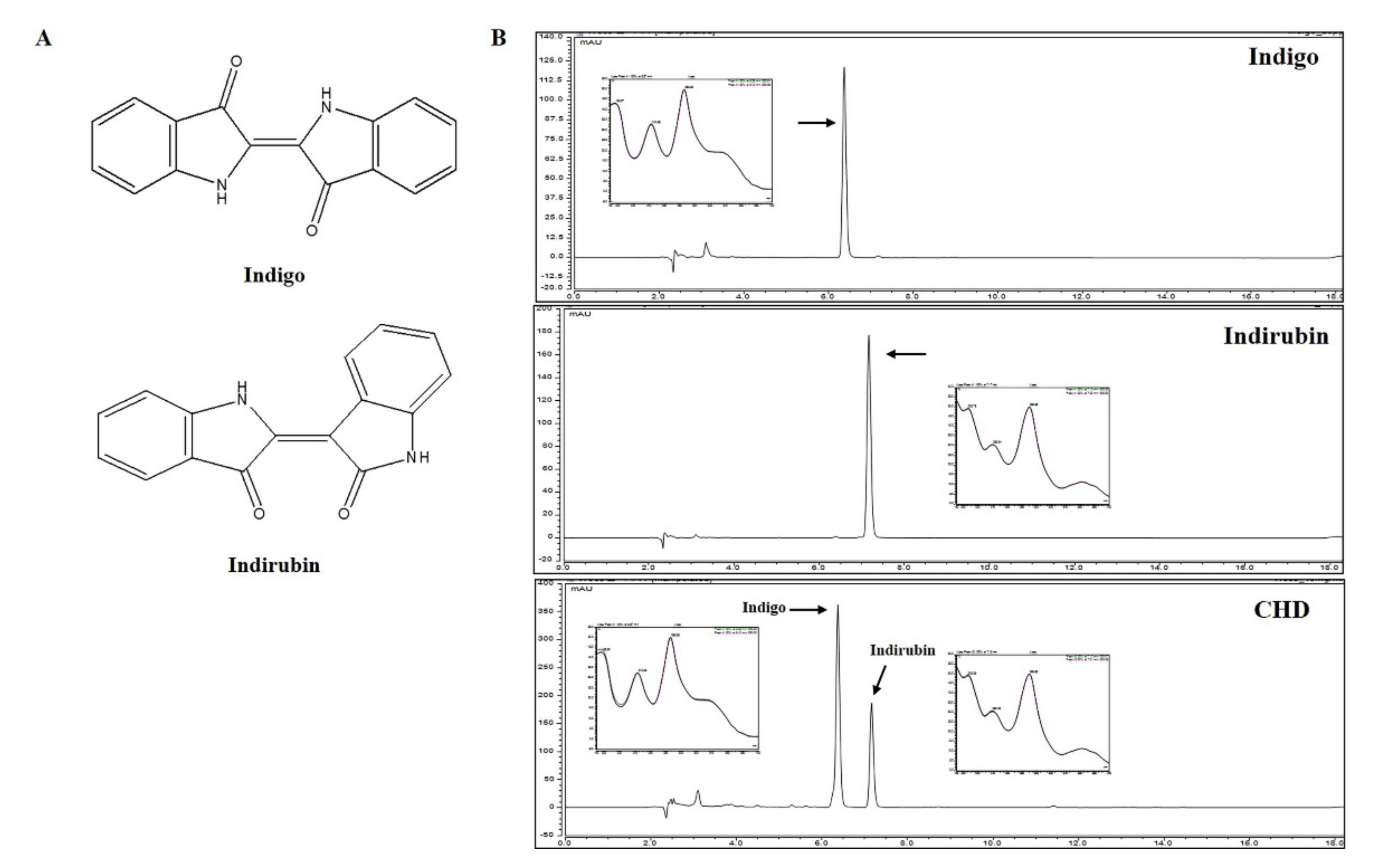
2.8. Identification and Quantification of Constituents of CHD
3. Discussion
4. Materials and Methods
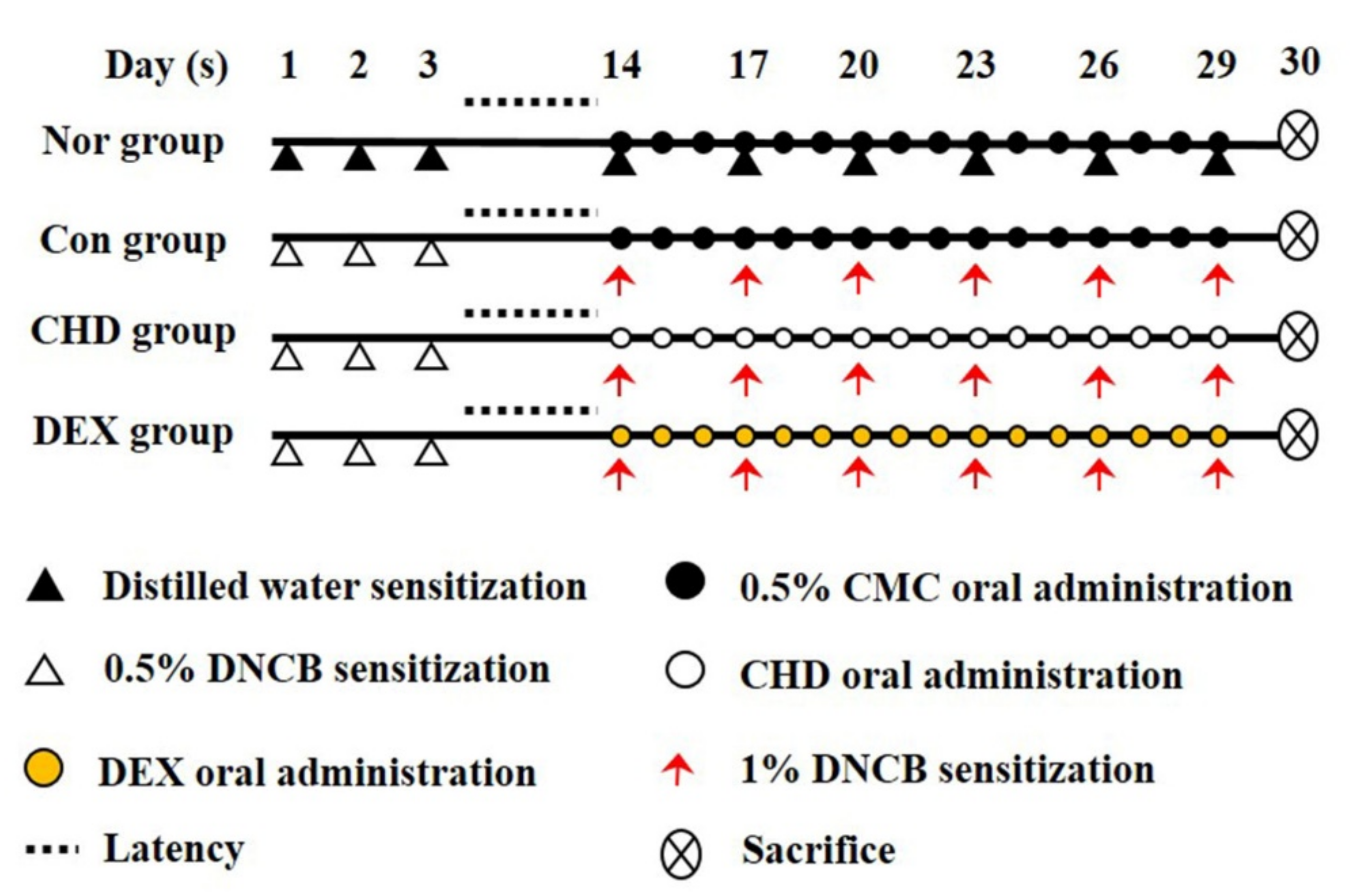
4.1. AD Mice Model and Drug Treatment
4.2. Histological Analysis
4.3. Preparation of CHD and Cell Culture
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
4.6. Western Blot Analysis
4.7. Immunocytochemical Analysis
4.8. CHD Sample Preparation and High-Performance Liquid Chromatography with Diode-Array Detection (HPLC-DAD)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Avena-Woods, C. Overview of atopic dermatitis. Am. J. Manag. Care 2017, 23, S115–S123. [Google Scholar]
- Guttman-Yassky, E.; Dhingra, N.; Leung, D.Y. New era of biologic therapeutics in atopic dermatitis. Expert Opin. Biol. Ther. 2013, 13, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Guttman-Yassky, E.; Nograles, K.E.; Krueger, J.G. Contrasting pathogenesis of atopic dermatitis and psoriasis—Part I: Clinical and pathologic concepts. J. Allergy Clin. Immunol. 2011, 127, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Kiebert, G.; Sorensen, S.V.; Revicki, D.; Fagan, S.C.; Doyle, J.J.; Cohen, J.; Fivenson, D. Atopic dermatitis is associated with a decrement in health-related quality of life. Int. J. Dermatol. 2002, 41, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Drucker, A.M.; Lebwohl, M.; Silverberg, J.I. A systematic review of the safety and efficacy of systemic corticosteroids in atopic dermatitis. J. Am. Acad. Dermatol. 2018, 78, 733–740.e11. [Google Scholar] [CrossRef] [PubMed]
- Saeki, H.; Nakahara, T.; Tanaka, A.; Kabashima, K.; Sugaya, M.; Murota, H.; Ebihara, T.; Kataoka, Y.; Aihara, M.; Etoh, T.; et al. Clinical Practice Guidelines for the Management of Atopic Dermatitis 2016. J. Dermatol. 2016, 43, 1117–1145. [Google Scholar] [CrossRef]
- Lu, C.-Y.; Chang, H.-H.; Sung, F.-C.; Chen, P.-C. Characteristics of Traditional Chinese Medicine Use in Pediatric Dislocations, Sprains and Strains. Int. J. Environ. Res. Public Health 2017, 14, 153. [Google Scholar] [CrossRef] [Green Version]
- Gutermuth, J.; Ollert, M.; Ring, J.; Behrendt, H.; Jakob, T. Mouse models of atopic eczema critically evaluated. Int. Arch. Allergy Immunol. 2004, 135, 262–276. [Google Scholar] [CrossRef]
- Brandt, E.B.; Sivaprasad, U. Th2 cytokines and atopic dermatitis. J. Clin. Cell. Immunol. 2011. [Google Scholar] [CrossRef]
- Huidrom, S. Therapeutic Approach of Probiotics in Children with Atopic Dermatitis. Antiinflamm. Antiallergy Agents Med. Chem. 2021, 20, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.-Y.; Ryu, M.-S.; Yang, H.-J.; Jeong, S.-Y.; Zhang, T.; Yang, H.J.; Kim, M.J.; Park, S. Pediococcus acidilactici intake decreases the clinical severity of atopic dermatitis along with increasing mucin production and improving the gut microbiome in Nc/Nga mice. Biomed. Pharmacother. 2020, 129, 110488. [Google Scholar] [CrossRef] [PubMed]
- Seitz, C.S.; Lin, Q.; Deng, H.; Khavari, P.A. Alterations in NF-κB function in transgenic epithelial tissue demonstrate a growth inhibitory role for NF-κB. Proc. Natl. Acad. Sci. USA 1998, 95, 2307–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smahi, A.; Courtois, G.; Rabia, S.H.; Döffinger, R.; Bodemer, C.; Munnich, A.; Casanova, J.-L.; Israël, A. The NF-κB signalling pathway in human diseases: From incontinentia pigmenti to ectodermal dysplasias and immune-deficiency syndromes. Hum. Mol. Genet. 2002, 11, 2371–2375. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-S.; Chun, S.-Y.; Lee, M.-G.; Kim, S.; Jang, T.-J.; Nam, K.-S. The prevention of TNF-α/IFN-γ mixture-induced inflammation in human keratinocyte and atopic dermatitis-like skin lesions in Nc/Nga mice by mineral-balanced deep sea water. Biomed. Pharmacother. 2018, 97, 1331–1340. [Google Scholar] [CrossRef]
- Park, D.-N.; Beak, S.-R.; Jeon, D.-W. A study on the effect of the changes of dyeing conditions on the dyeability of silk fabrics dyed with natural polygoum tinctoria. J. Fash. Bus. 2011, 15, 120–130. [Google Scholar]
- Tang, W.; Eisenbrand, G. Chinese drugs of plant origin. Chemistry, pharmacology, and use in traditional modern medicine. In Chinese Drugs of Plant Origin; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Kim, H.-M.; Hong, D.-R.; Lee, E.-H. Inhibition of Mast Cell-Dependent Anaphylactic Reactions by the Pigment of Polygonum tinctorium (Chung-Dae) in Rats. Gen. Pharmacol. Vasc. Syst. 1998, 31, 361–365. [Google Scholar] [CrossRef]
- Seo, H.-S. The experimental study on anti-inflammation and anti-oxidation of Indigo Naturalis and Rehmanniae Radix. J. Korean Med. Ophthalmol. Otolaryngol. Dermatol. 2008, 21, 104–110. [Google Scholar]
- Sun, Q.; Leng, J.; Tang, L.; Wang, L.; Fu, C. A Comprehensive Review of the Chemistry, Pharmacokinetics, Pharmacology, Clinical Applications, Adverse Events, and Quality Control of Indigo Naturalis. Front. Pharmacol. 2021, 12, 664022. [Google Scholar] [CrossRef]
- Han, N.-R.; Kang, S.W.; Moon, P.-D.; Jang, J.-B.; Kim, H.-M.; Jeong, H.-J. Genuine traditional Korean medicine, Naju Jjok (Chung-Dae, Polygonum tinctorium) improves 2,4-dinitrofluorobenzene-induced atopic dermatitis-like lesional skin. Phytomedicine 2014, 21, 453–460. [Google Scholar] [CrossRef]
- Choo, H.-N.; Park, E.-H.; Jeong, J.-K.; Paek, Y.-W.; Kim, J.-S. The Effect of the Polygonum tinctoria Niram on Atopic Dermatitis in DNCB-Induced Hairless Mice. Herb. Formula Sci. 2014, 22, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-K.; Leu, Y.-L.; Huang, T.-H.; Wu, Y.-H.; Chung, P.-J.; Pang, J.-H.S.; Hwang, T.-L. Anti-inflammatory effects of the extract of indigo naturalis in human neutrophils. J. Ethnopharmacol. 2009, 125, 51–58. [Google Scholar] [CrossRef]
- Watanabe, H.; Unger, M.; Tuvel, B.; Wang, B.; Sauder, D.N. Review: Contact Hypersensitivity: The Mechanism of Immune Responses and T Cell Balance. J. Interf. Cytokine Res. 2002, 22, 407–412. [Google Scholar] [CrossRef]
- Yarbrough, K.B.; Neuhaus, K.J.; Simpson, E.L. The effects of treatment on itch in atopic dermatitis. Dermatol. Ther. 2013, 26, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-S.; Jeong, E.-S.; Heo, S.-H.; Seo, J.-H.; Jeong, D.-G.; Choi, Y.-K. A Novel Model for Human Atopic Dermatitis: Application of Repeated DNCB Patch in BALB/c Mice, in Comparison with NC/Nga Mice. Lab. Anim. Res. 2010, 26, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Gao, S.; Wu, G.-Z.; Yang, N.; Zu, X.-P.; Li, W.-C.; Xie, N.; Zhang, R.-R.; Li, C.-W.; Hu, Z.-L.; et al. Total sesquiterpene lactones isolated from Inula helenium L. attenuates 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice. Phytomedicine 2018, 46, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Bieber, T. Atopic dermatitis. N. Engl. J. Med. 2008, 358, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Kim, J.S.; Cho, D.H.; Park, H.J. Molecular mechanisms of cutaneous inflammatory disorder: Atopic dermatitis. Int. J. Mol. Sci. 2016, 17, 1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleimer, R.P.; Sterbinsky, S.; Kaiser, J.; Bickel, A.C.; Klunk, D.; Tomioka, K.; Newman, W.; Luscinskas, F.W.; Gimbrone, M.; McIntyre, B.W. IL-4 induces adherence of human eosinophils and basophils but not neutrophils to endothelium. Association with expression of VCAM-1. J. Immunol. 1992, 148. [Google Scholar]
- Metcalfe, D.D.; Baram, D.; Mekori, Y.A. Mast cells. Physiol. Rev. 1997, 77, 1033–1079. [Google Scholar] [CrossRef]
- Hong, S.H.; Ku, J.M.; Kim, H.I.; Kim, T.Y.; Seo, H.S.; Shin, Y.C.; Ko, S.-G. Topical Application of KAJD Attenuates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis Symptoms Through Regulation of IgE and MAPK Pathways in BALB/C Mice and Several Immune Cell Types. Front. Pharmacol. 2019, 10, 1097. [Google Scholar] [CrossRef]
- Kawakami, T.; Ando, T.; Kimura, M.; Wilson, B.S.; Kawakami, Y. Mast cells in atopic dermatitis. Curr. Opin. Immunol. 2009, 21, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Marone, G.; Lichtenstein, L.M.; Galli, F.J. Mast Cells and Basophils; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Galli, S.J.; Maurer, M.; Lantz, C.S. Mast cells as sentinels of innate immunity. Curr. Opin. Immunol. 1999, 11, 53–59. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.-T.; Goodarzi, H.; Chen, H.-Y. IgE, mast cells, and eosinophils in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Schmelz, M.; Bíró, T.; Steinhoff, M. Frontiers in pruritus research: Scratching the brain for more effective itch therapy. J. Clin. Investig. 2006, 116, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- McGirt, L.Y.; Beck, L.A. Innate immune defects in atopic dermatitis. J. Allergy Clin. Immunol. 2006, 118, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Rollins, B.J.; Yoshimura, T.; Leonard, E.J.; Pober, J.S. Cytokine-activated human endothelial cells synthesize and secrete a monocyte chemoattractant, MCP-1/JE. Am. J. Pathol. 1990, 136, 1229–1233. [Google Scholar] [PubMed]
- Power, C.; Proudfoot, E.A. The chemokine system: Novel broad-spectrum therapeutic targets. Curr. Opin. Pharmacol. 2001, 1, 417–424. [Google Scholar] [CrossRef]
- Campbell, J.J.; Haraldsen, G.; Pan, J.; Rottman, J.; Qin, S.; Ponath, P.; Andrew, D.P.; Warnke, R.; Ruffing, N.; Kassam, N.; et al. The chemokine receptor CCR4 in vascular recognition by cutaneous but not intestinal memory T cells. Nature 1999, 400, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Song, T.W.; Kwon, B.C.; Choi, S.Y.; Shin, Y.H.; Lee, K.E.; Yang, H.S.; Kim, K.W.; Kim, E.S.; Sohn, M.H.; Kim, K.E. Increased serum thymus and activation-regulated chemokine (TARC) levels in children with atopic dermatitis. Pediatric Allergy Respir. Dis. 2005, 15, 250–256. [Google Scholar]
- Luster, A.D. The role of chemokines in linking innate and adaptive immunity. Curr. Opin. Immunol. 2002, 14, 129–135. [Google Scholar] [CrossRef]
- Bochner, B.S.; Luscinskas, F.W.; Gimbrone Jr, M.A.; Newman, W.; Sterbinsky, S.A.; Derse-Anthony, C.P.; Klunk, D.; Schleimer, R.P. Adhesion of human basophils, eosinophils, and neutrophils to interleukin 1-activated human vascular endothelial cells: Contributions of endothelial cell adhesion molecules. J. Exp. Med. 1991, 173, 1553–1557. [Google Scholar] [CrossRef] [Green Version]
- Gudbjörnsson, B.; Hällgren, R.; Nettelbladt, O.; Gustafsson, R.; Mattsson, A.; af Geijerstam, E.; Tötterman, T. Phenotypic and functional activation of alveolar macrophages, T lymphocytes and NK cells in patients with systemic sclerosis and primary Sjögren’s syndrome. Ann. Rheum. Dis. 1994, 53, 574–579. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Chen, Q.; Hoover, D.M.; Staley, P.; Tucker, K.D.; Lubkowski, J.; Oppenheim, J.J. Many chemokines including CCL20/MIP-3α display antimicrobial activity. J. Leukoc. Biol. 2003, 74, 448–455. [Google Scholar] [CrossRef]
- Dieu-Nosjean, M.-C.; Massacrier, C.; Homey, B.; Vanbervliet, B.; Pin, J.-J.; Vicari, A.; Lebecque, S.; Dezutter-Dambuyant, C.; Schmitt, D.; Zlotnik, A.; et al. Macrophage Inflammatory Protein 3α Is Expressed at Inflamed Epithelial Surfaces and Is the Most Potent Chemokine Known in Attracting Langerhans Cell Precursors. J. Exp. Med. 2000, 192, 705–718. [Google Scholar] [CrossRef]
- Hoover, D.M.; Boulegue, C.; Yang, D.; Oppenheim, J.J.; Tucker, K.; Lu, W.; Lubkowski, J. The structure of human macrophage inflammatory protein-3α/CCL20: Linking antimicrobial and cc chemokine receptor-6-binding activities with human β-defensins. J. Biol. Chem. 2002, 277, 37647–37654. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.R.; Lee, H.; Kim, J.-H.; Hong, I.-H.; Hwang, D.H.; Rho, I.R.; Kim, G.S.; Kim, E.; Kang, C. Therapeutic Effect of Rumex japonicus Houtt. on DNCB-Induced Atopic Dermatitis-Like Skin Lesions in Balb/c Mice and Human Keratinocyte HaCaT Cells. Nutrients 2019, 11, 573. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.; Shen, Z.; Chen, H.; Liu, J.; Jiang, K.; Fan, L.; Jia, L.; Shao, J. Metapristone suppresses non-small cell lung cancer proliferation and metastasis via modulating RAS/RAF/MEK/MAPK signaling pathway. Biomed. Pharmacother. 2017, 90, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.H.; Oh, Y.-C.; Cho, W.-K.; Lee, B.; Ma, J.Y. Anti-inflammatory effects of melandrii herba ethanol extract via inhibition of NF-κB and MAPK signaling pathways and induction of HO-1 in RAW 264. 7 cells and mouse primary macrophages. Molecules 2016, 21, 818. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Sun, Y.; Li, X.; Wang, J.; Yan, L.; Zhang, Z.; Wang, D.; Dai, J.; He, J.; Wang, S. Scutellarin inhibits RANKL-mediated osteoclastogenesis and titanium particle-induced osteolysis via suppression of NF-κB and MAPK signaling pathway. Int. Immunopharmacol. 2016, 40, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Kwon, O.-K.; Yuniato, P.; Marwoto, B.; Lee, J.; Oh, S.-R.; Kim, J.-H.; Ahn, K.-S. Amelioration of an LPS-induced inflammatory response using a methanolic extract of Lagerstroemia ovalifolia to suppress the activation of NF-κB in RAW264. 7 macrophages. Int. J. Mol. Med. 2016, 38, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandel, N.S.; Trzyna, W.C.; McClintock, D.S.; Schumacker, P.T. Role of oxidants in NF-κB activation and TNF-α gene transcription induced by hypoxia and endotoxin. J. Immunol. 2000, 165, 1013–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, Y.; Lee, W.-H.; Jeong, J.; Park, M.; Ko, J.-Y.; Kwon, O.W.; Lee, J.; Kim, Y.-J. Pyropia yezoensis extract suppresses IFN-gamma-and TNF-alpha-induced proinflammatory chemokine production in HaCaT cells via the down-regulation of NF-κB. Nutrients 2020, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Kawai, S.; Iijima, H.; Shinzaki, S.; Hiyama, S.; Yamaguchi, T.; Araki, M.; Iwatani, S.; Shiraishi, E.; Mukai, A.; Inoue, T.; et al. Indigo Naturalis ameliorates murine dextran sodium sulfate-induced colitis via aryl hydrocarbon receptor activation. J. Gastroenterol. 2016, 52, 904–919. [Google Scholar] [CrossRef]
- Lee, C.-L.; Wang, C.-M.; Hu, H.-C.; Yen, H.-R.; Song, Y.-C.; Yu, S.-J.; Chen, C.-J.; Li, W.-C.; Wu, Y.-C. Indole alkaloids indigodoles A–C from aerial parts of Strobilanthes cusia in the traditional Chinese medicine Qing Dai have anti-IL-17 properties. Phytochemistry 2019, 162, 39–46. [Google Scholar] [CrossRef]
- Yang, J.-H.; Do, H.J.; Lee, E.; Yim, N.-H.; Cho, W.-K.; Park, K.-I.; Ma, J.Y. Jageum-Jung improves 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice and suppresses pro-inflammatory chemokine production by inhibiting TNF-α/IFN-γ-induced STAT-1 and NFκB signaling in HaCaT cells. J. Ethnopharmacol. 2018, 221, 48–55. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Range (ug/mL, ppm) | Regression Equation | r2 | LOD (μg/mL) | LOQ (μg/mL) |
|---|---|---|---|---|---|
| Indigo | 10.0~80.0 | y = 0.3334x − 0.5198 | 0.9958 | 0.0164 | 0.0499 |
| Indirubin | 2.0~16.0 | y = 1.7955x + 0.2972 | 0.9997 | 0.0030 | 0.0091 |
| Name | Forward | Reverse |
|---|---|---|
| h-RANTES | 5′ GATGCCAAAG AGAGAGGGAC 3′ | 5′ AAATTTGTGT AAGTTCAGGT 3′ |
| h-TARC | 5′ CTGCACACAG AGACTCCCTC 3′ | 5′ CTGGTACCAC GTCTTCAGCT 3′ |
| h-MDC | 5′ GAAACACTTC TACTGGACCT 3′ | 5′ CAGGGAGGTA GGGCTCCTGA 3′ |
| h-GAPDH | 5′ TCAAGGCTGA GAACGGGAAG 3′ | 5′ TGGACTCCAC GACGTACTCA 3′ |
| m-TNF-α | 5′ ATGAGCACAG AAAGCATGAT 3′ | 5′ TACAGGCTTG TCACTCGAAT 3′ |
| m-IL-6 | 5′ TTCCATCCAG TTGCCTTCTT 3′ | 5′ ATTTCCACGA TTTCCCAGAG 3′ |
| m-IL-13 | 5′ CATCTCCAAT TGCAATGCCA 3′ | 5′ GCCCAGGGAT GGTCTCTCCT 3′ |
| m-GAPDH | 5′ AACGACCCCT TCATTGAC 3′ | 5′ TCCACGACAT ACTCAGCAC 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, G.-Y.; Kim, J.-H.; Kim, T.-I.; Cho, W.-K.; Yang, J.-H.; Ma, J.-Y. Indigo Pulverata Levis (Chung-Dae, Persicaria tinctoria) Alleviates Atopic Dermatitis-like Inflammatory Responses In Vivo and In Vitro. Int. J. Mol. Sci. 2022, 23, 553. https://doi.org/10.3390/ijms23010553
Min G-Y, Kim J-H, Kim T-I, Cho W-K, Yang J-H, Ma J-Y. Indigo Pulverata Levis (Chung-Dae, Persicaria tinctoria) Alleviates Atopic Dermatitis-like Inflammatory Responses In Vivo and In Vitro. International Journal of Molecular Sciences. 2022; 23(1):553. https://doi.org/10.3390/ijms23010553
Chicago/Turabian StyleMin, Ga-Yul, Ji-Hye Kim, Tae-In Kim, Won-Kyung Cho, Ju-Hye Yang, and Jin-Yeul Ma. 2022. "Indigo Pulverata Levis (Chung-Dae, Persicaria tinctoria) Alleviates Atopic Dermatitis-like Inflammatory Responses In Vivo and In Vitro" International Journal of Molecular Sciences 23, no. 1: 553. https://doi.org/10.3390/ijms23010553