
Structural Features of Cytochrome b5–Cytochrome b5 Reductase Complex Formation and Implications for the Intramolecular Dynamics of Cytochrome b5 Reductase

 ,
,  , and
, and 
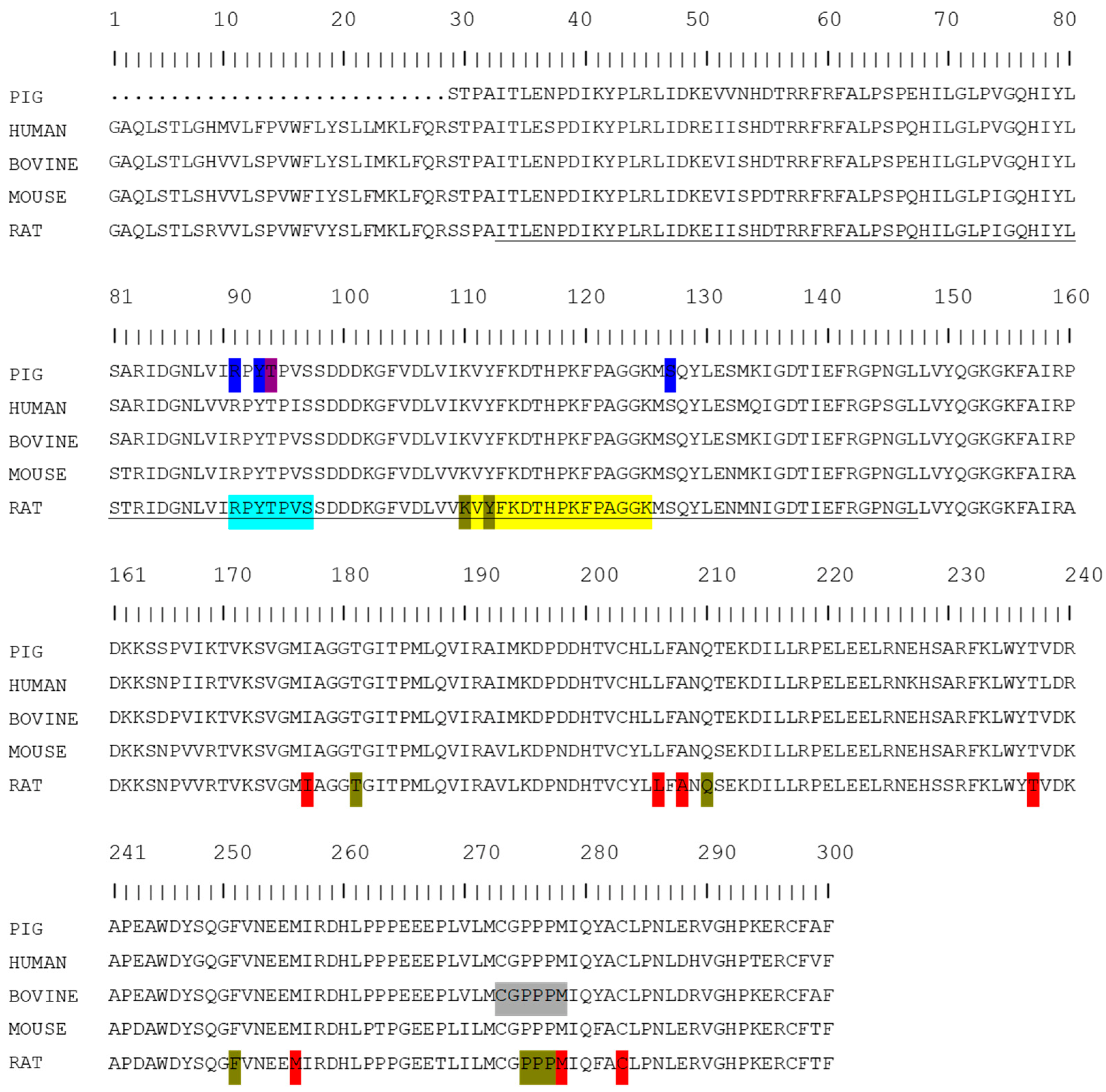{kind=link}
{kind=link}
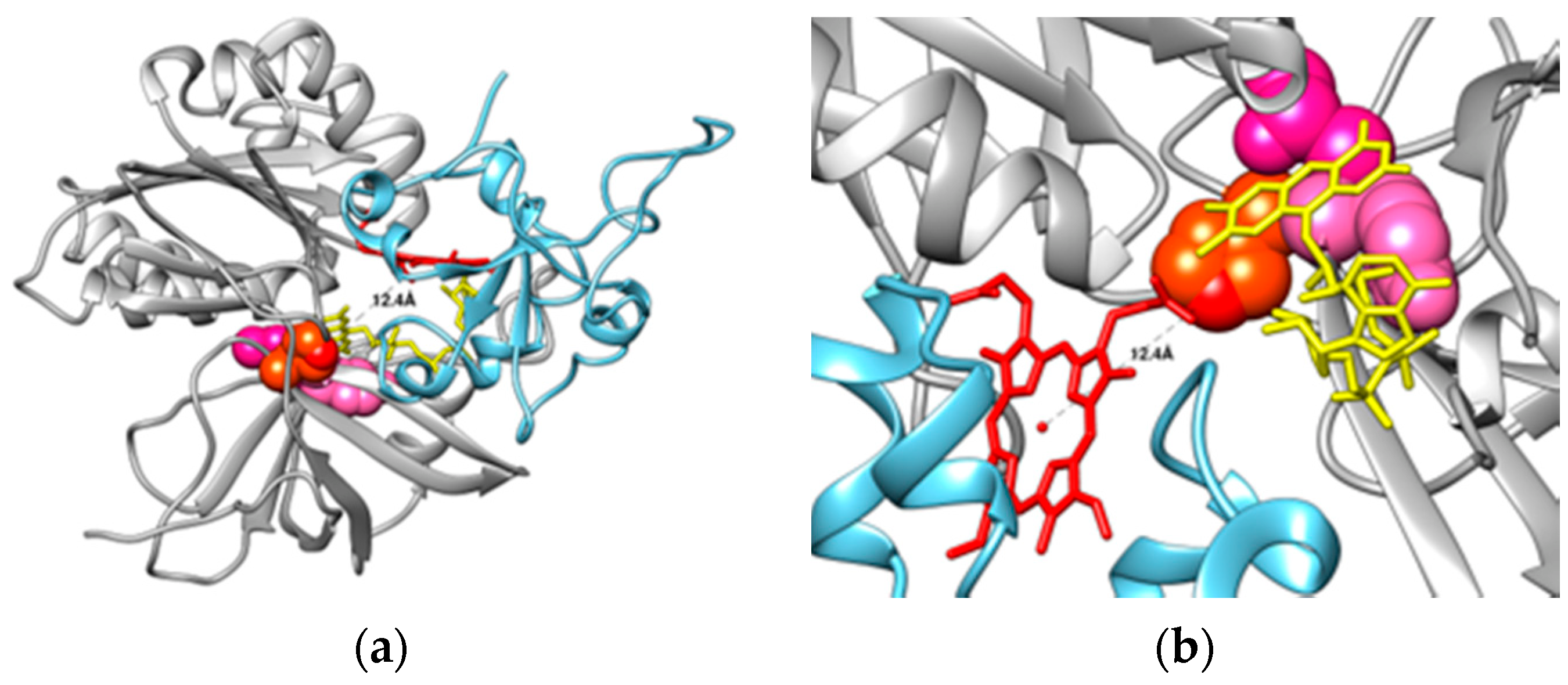{kind=link}
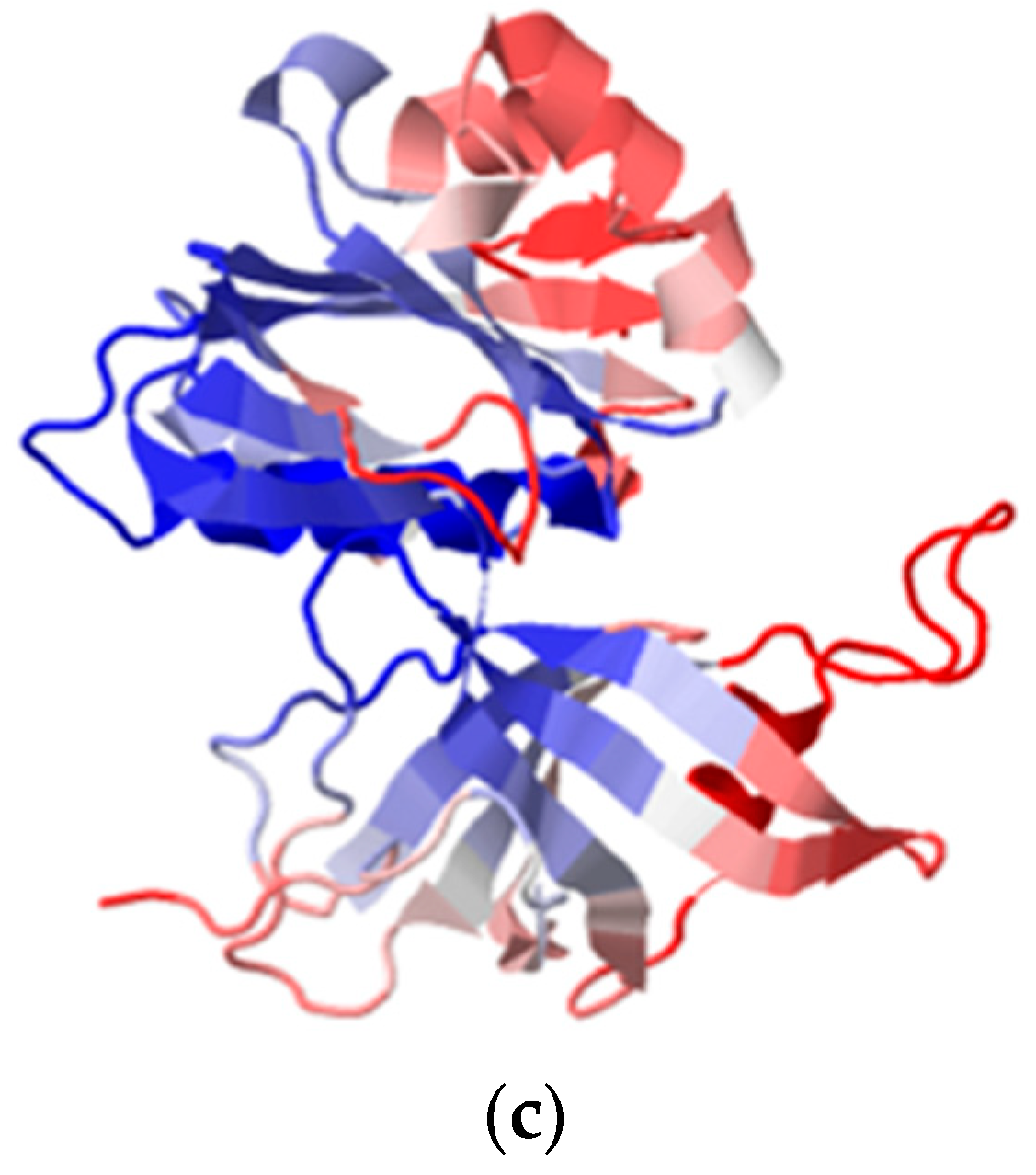{kind=link}
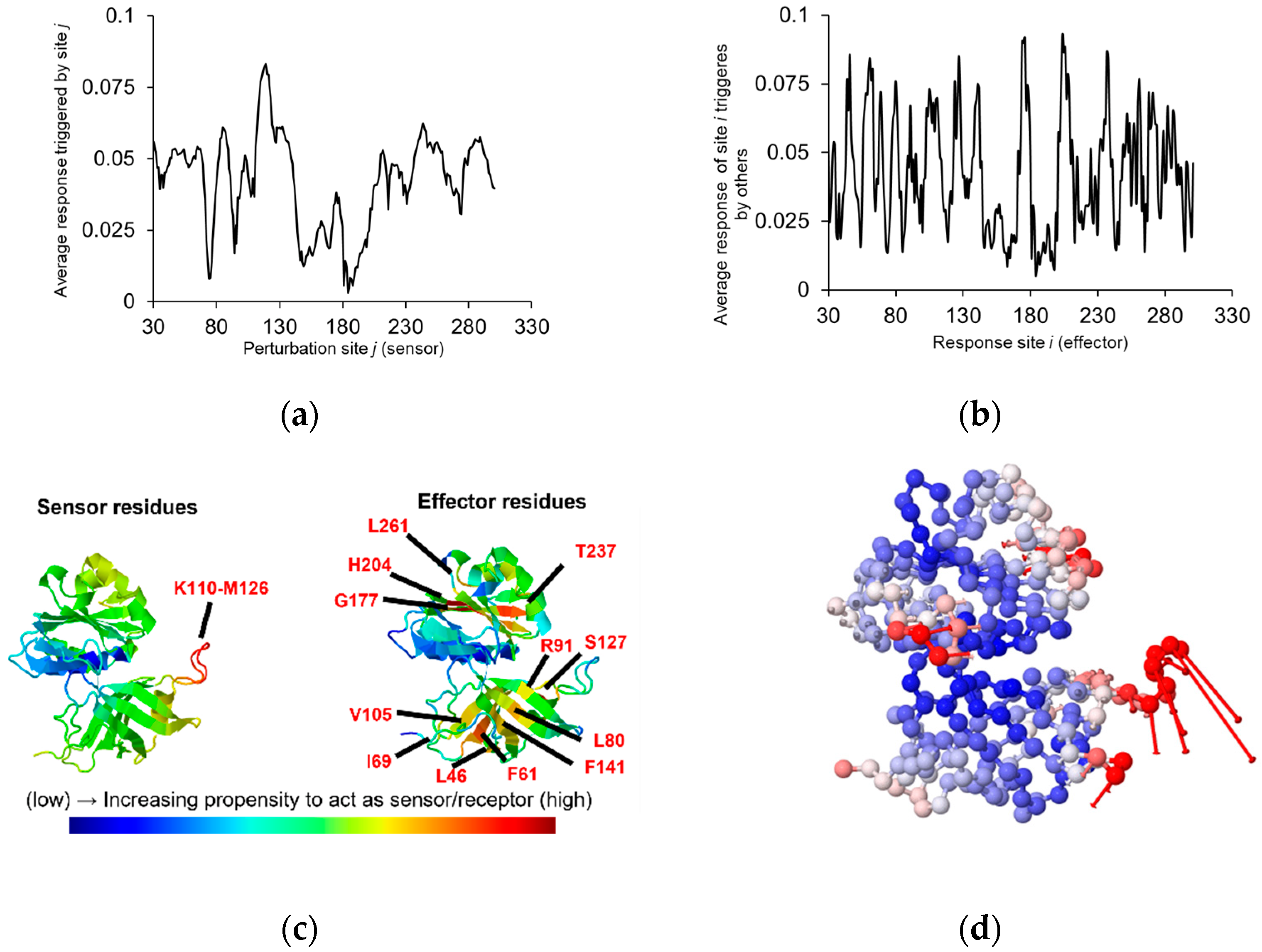{kind=link}
Abstract
:1. The Electron Transfer from the Cytochrome b5 Reductase to Multiple Acceptors and Implication in Reactive Oxygen Species Formation
1.1. The Endoplasmic Reticulum Cb5R
1.2. The MOM Cb5R
1.3. The Plasma Membrane Cb5R
1.4. Antioxidants Recycling Activity of Cb5R and the Importance in Cellular ROS Balance
2. Structural Features of Soluble Cb5R Helps to Rationalize the Electron Transfer Processes Using NADH as a Substrate
2.1. FAD-Binding Domain of Cb5R
2.2. The NADH-Binding Pocket and Hydride Transfer to the Flavin Group of the Reductase
3. Cb5-Interacting Domain of Cb5R
Electron Transfer from the Flavoprotein to Cb5
4. Flavin Fluorescence and Structural Alterations by Complexes Formation with Electron Acceptors
5. Intramolecular Dynamics of Cb5R
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, F. Flavin radicals: Chemistry and biochemistry. Free Radic. Biol. Med. 1987, 3, 215–230. [Google Scholar] [CrossRef]
- Strittmatter, P. The Reaction Sequence in Electron Transfer in the Reduced Nicotinamide Adenine Dinucleotide-Cytochrome B5 Reductase System. J. Biol. Chem. 1965, 240, 4481–4487. [Google Scholar] [CrossRef]
- Iyanagi, T. Redox Properties of Microsomal Reduced Nicotinamide Adenine Dinucleotide-Cytochrome B5 Reductase and Cytochrome B5. Biochemistry 1977, 16, 2725–2730. [Google Scholar] [CrossRef]
- Meyer, T.E.; Shirabe, K.; Yubisui, T.; Takeshita, M.; Bes, M.T.; Cusanovich, M.A.; Tollin, G. Transient Kinetics of Intracomplex Electron-Transfer in the Human Cytochrome B5 Reductase-Cytochrome B5 System: NAD+ Modulates Protein-Protein Binding and Electron Transfer. Arch. Biochem. Biophys. 1995, 318, 457–464. [Google Scholar] [CrossRef]
- Kobayashi, K.; Iyanagi, T.; Ohara, H.; Hayashi, K. One-Electron Reduction of Hepatic NADH-Cytochrome B5 Reductase as Studied by Pulse Radiolysis. J. Biol. Chem. 1988, 263, 7493–7499. [Google Scholar] [CrossRef]
- Anderson, R.F. Pulse radiolysis studies on the equilibria between reduced and oxidized free flavin species and the effect of molecular oxygen. In Pulse Radiolysis Studies on the Equilibria between Reduced and Oxidized Free Flavin Species and the Effect of Molecular Oxygen; De Gruyter: Berlin, Germany, 2019; pp. 57–60. ISBN 978-3-11-152135-0. [Google Scholar]
- Deng, B.; Parthasarathy, S.; Wang, W.; Gibney, B.R.; Battaile, K.P.; Lovell, S.; Benson, D.R.; Zhu, H. Study of the Individual Cytochrome B5 and Cytochrome B5 Reductase Domains of Ncb5or Reveals a Unique Heme Pocket and a Possible Role of the CS Domain. J. Biol. Chem. 2010, 285, 30181–30191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siendones, E.; Ballesteros, M.; Navas, P. Cellular and Molecular Mechanisms of Recessive Hereditary Methaemoglobinaemia Type II. J. Clin. Med. 2018, 7, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samhan-Arias, A.K.; López-Sánchez, C.; Marques-da-Silva, D.; Lagoa, R.; Garcia-Lopez, V.; García-Martínez, V.; Gutierrez-Merino, C. Biochemical and Anatomical Basis of Brain Dysfunctions Caused by Cytochrome B5 Reductase Deficiency or Dysregulation. J. Neurol. Neuromed. 2016, 1, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Ozols, J.; Carr, S.A.; Strittmatter, P. Identification of the NH2-Terminal Blocking Group of NADH-Cytochrome B5 Reductase as Myristic Acid and the Complete Amino Acid Sequence of the Membrane-Binding Domain. J. Biol. Chem. 1984, 259, 13349–13354. [Google Scholar] [CrossRef]
- Borgese, N.; D’Arrigo, A.; De Silvestris, M.; Pietrini, G. NADH-Cytochrome b 5 Reductase and Cytochrome b 5 Isoforms as Models for the Study of Post-Translational Targeting to the Endoplasmic Reticulum. FEBS Lett. 1993, 325, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.F.; Vergeres, G.; Waskell, L. Kinetics of the Reduction of Cytochrome B5 with Mutations in Its Membrane-Binding Domain. Arch. Biochem. Biophys. 1994, 308, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Borgese, N.; Longhi, R. Both the Outer Mitochondrial Membrane and the Microsomal Forms of Cytochrome B5 Reductase Contain Covalently Bound Myristic Acid. Quantitative Analysis on the Polyvinylidene Difluoride-Immobilized Proteins. Biochem. J. 1990, 266, 341–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirabe, K.; Landi, M.T.; Takeshita, M.; Uziel, G.; Fedrizzi, E.; Borgese, N. A Novel Point Mutation in a 3’ Splice Site of the NADH-Cytochrome B5 Reductase Gene Results in Immunologically Undetectable Enzyme and Impaired NADH-Dependent Ascorbate Regeneration in Cultured Fibroblasts of a Patient with Type II Hereditary Methemoglobinemia. Am. J. Hum. Genet. 1995, 57, 302–310. [Google Scholar]
- Samhan-Arias, A.K.; Gutierrez-Merino, C. Cytochrome B5 as a pleitropic metabolic modulator in mammalian cells. In Cytochromes b and c: Biochemical Properties, Biological Functions and Electrochemical Analysis; Rurik, T., Ed.; Nova Publishers: New York, NY, USA, 2014; ISBN 978-1-63117-467-4. [Google Scholar]
- Borgese, N.; Aggujaro, D.; Carrera, P.; Pietrini, G.; Bassetti, M. A Role for N-Myristoylation in Protein Targeting: NADH-Cytochrome B5 Reductase Requires Myristic Acid for Association with Outer Mitochondrial but Not ER Membranes. J. Cell Biol. 1996, 135, 1501–1513. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.I.; Duronio, R.J.; Rudnick, D.A.; Adams, S.P.; Gokel, G.W. Protein N-Myristoylation. J. Biol. Chem. 1991, 266, 8647–8650. [Google Scholar] [CrossRef]
- Gómez-Tabales, J.; García-Martín, E.; Agúndez, J.A.G.; Gutierrez-Merino, C. Modulation of CYP2C9 Activity and Hydrogen Peroxide Production by Cytochrome B5. Sci. Rep. 2020, 10, 15571. [Google Scholar] [CrossRef] [PubMed]
- Samhan-Arias, A.K.; Garcia-Bereguiain, M.A.; Martin-Romero, F.J.; Gutierrez-Merino, C. Clustering of Plasma Membrane-Bound Cytochrome B5 Reductase within “lipid Raft” Microdomains of the Neuronal Plasma Membrane. Mol. Cell. Neurosci. 2009, 40, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Samhan-Arias, A.K.; Gutierrez-Merino, C. Purified NADH-Cytochrome B5 Reductase Is a Novel Superoxide Anion Source Inhibited by Apocynin: Sensitivity to Nitric Oxide and Peroxynitrite. Free Radic. Biol. Med. 2014, 73, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Pearl, N.M.; Wilcoxen, J.; Im, S.; Kunz, R.; Darty, J.; Britt, R.D.; Ragsdale, S.W.; Waskell, L. Protonation of the Hydroperoxo Intermediate of Cytochrome P450 2B4 Is Slower in the Presence of Cytochrome P450 Reductase Than in the Presence of Cytochrome B5. Biochemistry 2016, 55, 6558–6567. [Google Scholar] [CrossRef]
- Hahne, K.; Haucke, V.; Ramage, L.; Schatz, G. Incomplete Arrest in the Outer Membrane Sorts NADH-Cytochrome B5 Reductase to Two Different Submitochondrial Compartments. Cell 1994, 79, 829–839. [Google Scholar] [CrossRef]
- The Oxidation of External NADH by an Intermembrane Electron Transfer in Mitochondria from the Ubiquinone-Deficient Mutant E3-24 of Saccharomyces cerevisiae—ScienceDirect. Available online: https://www.sciencedirect.com/science/article/abs/pii/0003986184905514?via%3Dihub (accessed on 9 November 2021).
- Ott, G.; Havemeyer, A.; Clement, B. The Mammalian Molybdenum Enzymes of MARC. J. Biol. Inorg. Chem. 2015, 20, 265–275. [Google Scholar] [CrossRef]
- Yuan, S.; Hahn, S.A.; Miller, M.P.; Sanker, S.; Calderon, M.J.; Sullivan, M.; Dosunmu-Ogunbi, A.M.; Fazzari, M.; Li, Y.; Reynolds, M.; et al. Cooperation between CYB5R3 and NOX4 via Coenzyme Q Mitigates Endothelial Inflammation. Redox Biol. 2021, 47, 102166. [Google Scholar] [CrossRef] [PubMed]
- Takac, I.; Schröder, K.; Zhang, L.; Lardy, B.; Anilkumar, N.; Lambeth, J.D.; Shah, A.M.; Morel, F.; Brandes, R.P. The E-Loop Is Involved in Hydrogen Peroxide Formation by the NADPH Oxidase Nox4. J. Biol. Chem. 2011, 286, 13304–13313. [Google Scholar] [CrossRef] [Green Version]
- Powis, G.; Svingen, B.A.; Appel, P. Factors Affecting the Intracellular Generation of Free Radicals from Quinones. Adv. Exp. Med. Biol. 1981, 136 Pt A, 349–358. [Google Scholar] [CrossRef]
- Valério, G.N.; Gutiérrez-Merino, C.; Nogueira, F.; Moura, I.; Moura, J.J.G.; Samhan-Arias, A.K. Human Erythrocytes Exposure to Juglone Leads to an Increase of Superoxide Anion Production Associated with Cytochrome B5 Reductase Uncoupling. Biochim. Biophys. Acta—Bioenerg. 2020, 1861, 148134. [Google Scholar] [CrossRef] [PubMed]
- Samoilova, R.I.; Crofts, A.R.; Dikanov, S.A. Reaction of Superoxide Radical with Quinone Molecules. J. Phys. Chem. A 2011, 115, 11589–11593. [Google Scholar] [CrossRef] [Green Version]
- Linnane, A.W.; Kios, M.; Vitetta, L. Coenzyme Q(10)—Its Role as a Prooxidant in the Formation of Superoxide Anion/Hydrogen Peroxide and the Regulation of the Metabolome. Mitochondrion 2007, 7, S51–S61. [Google Scholar] [CrossRef]
- Samhan-Arias, A.K.; Marques-da-Silva, D.; Yanamala, N.; Gutierrez-Merino, C. Stimulation and Clustering of Cytochrome B5 Reductase in Caveolin-Rich Lipid Microdomains Is an Early Event in Oxidative Stress-Mediated Apoptosis of Cerebellar Granule Neurons. J. Proteom. 2012, 75, 2934–2949. [Google Scholar] [CrossRef]
- Fortalezas, S.; Poejo, J. Cholesterol-Rich Plasma Membrane Submicrodomains Can Be a Major Extramitochondrial Source of Reactive Oxygen Species in Partially Depolarized Mature Cerebellar Granule Neurons in Culture. J. Neurophysiol. Neurol. Disord. 2019, 1, 1. [Google Scholar]
- Samhan-Arias, A.K.; Fortalezas, S.; Cordas, C.M.; Moura, I.; Moura, J.J.G.; Gutierrez-Merino, C. Cytochrome B5 Reductase Is the Component from Neuronal Synaptic Plasma Membrane Vesicles That Generates Superoxide Anion upon Stimulation by Cytochrome c. Redox Biol. 2018, 15, 109–114. [Google Scholar] [CrossRef]
- Pietrini, G.; Aggujaro, D.; Carrera, P.; Malyszko, J.; Vitale, A.; Borgese, N. A Single MRNA, Transcribed from an Alternative, Erythroid-Specific, Promoter, Codes for Two Non-Myristylated Forms of NADH-Cytochrome B5 Reductase. J. Cell Biol. 1992, 117, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Borgese, N.; Macconi, D.; Parola, L.; Pietrini, G. Rat Erythrocyte NADH-Cytochrome B5 Reductase. Quantitation and Comparison between the Membrane-Bound and Soluble Forms Using an Antibody against the Rat Liver Enzyme. J. Biol. Chem. 1982, 257, 13854–13861. [Google Scholar] [CrossRef]
- Martín-Romero, F.J.; Gutiérrez-Martín, Y.; Henao, F.; Gutiérrez-Merino, C. The NADH Oxidase Activity of the Plasma Membrane of Synaptosomes Is a Major Source of Superoxide Anion and Is Inhibited by Peroxynitrite. J. Neurochem. 2002, 82, 604–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samhan-Arias, A.K.; Almeida, R.M.; Ramos, S.; Cordas, C.M.; Moura, I.; Gutierrez-Merino, C.; Moura, J.J.G. Topography of Human Cytochrome B5/Cytochrome B5 Reductase Interacting Domain and Redox Alterations upon Complex Formation. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 78–87. [Google Scholar] [CrossRef]
- Hampton, M.B.; Zhivotovsky, B.; Slater, A.F.; Burgess, D.H.; Orrenius, S. Importance of the Redox State of Cytochrome c during Caspase Activation in Cytosolic Extracts. Biochem. J. 1998, 329 Pt 1, 95–99. [Google Scholar] [CrossRef]
- Constantinescu, A.; Han, D.; Packer, L. Vitamin E Recycling in Human Erythrocyte Membranes. J. Biol. Chem. 1993, 268, 10906–10913. [Google Scholar] [CrossRef]
- Lagoa, R.; Samhan-Arias, A.K.; Gutierrez-Merino, C. Correlation between the Potency of Flavonoids for Cytochrome c Reduction and Inhibition of Cardiolipin-Induced Peroxidase Activity. Biofactors 2017, 43, 451–468. [Google Scholar] [CrossRef]
- Kobayashi, K.; Harada, Y.; Hayashi, K. Kinetic Behavior of the Monodehydroascorbate Radical Studied by Pulse Radiolysis. Biochemistry 1991, 30, 8310–8315. [Google Scholar] [CrossRef]
- Hara, T.; Minakami, S. On Functional Role of Cytochrome B5. II. NADH-Linked Ascorbate Radical Reductase Activity in Microsomes. J. Biochem. 1971, 69, 325–330. [Google Scholar] [CrossRef]
- Villalba, J.M.; Navarro, F.; Gómez-Díaz, C.; Arroyo, A.; Bello, R.I.; Navas, P. Role of Cytochrome B5 Reductase on the Antioxidant Function of Coenzyme Q in the Plasma Membrane. Mol. Asp. Med. 1997, 18, 7–13. [Google Scholar] [CrossRef]
- Xiaoyuan Wang, P.J.Q. The Location and Function of Vitamin E in Membranes (Review). Mol. Membr. Biol. 2000, 17, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Wefers, H.; Sies, H. The Protection by Ascorbate and Glutathione against Microsomal Lipid Peroxidation Is Dependent on Vitamin E. Eur. J. Biochem. 1988, 174, 353–357. Available online: https://febs.onlinelibrary.wiley.com/doi/full/10.1111/j.1432-1033.1988.tb14105.x?sid=nlm%3Apubmed (accessed on 9 November 2021). [CrossRef] [PubMed]
- Frei, B.; Kim, M.C.; Ames, B.N. Ubiquinol-10 Is an Effective Lipid-Soluble Antioxidant at Physiological Concentrations. Proc. Natl. Acad. Sci. USA 1990, 87, 4879–4883. [Google Scholar] [CrossRef] [Green Version]
- Clemente, S.M.; Martínez-Costa, O.H.; Monsalve, M.; Samhan-Arias, A.K. Targeting Lipid Peroxidation for Cancer Treatment. Molecules 2020, 25, 5144. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.; Siegel, D. Functions of NQO1 in Cellular Protection and CoQ10 Metabolism and Its Potential Role as a Redox Sensitive Molecular Switch. Front. Physiol. 2017, 8, 595. [Google Scholar] [CrossRef]
- Maroz, A.; Anderson, R.F.; Smith, R.A.J.; Murphy, M.P. Reactivity of Ubiquinone and Ubiquinol with Superoxide and the Hydroperoxyl Radical: Implications for in Vivo Antioxidant Activity. Free Radic. Biol. Med. 2009, 46, 105–109. [Google Scholar] [CrossRef]
- Samhan-Arias, A.K.; Duarte, R.O.; Martín-Romero, F.J.; Moura, J.J.G.; Gutiérrez-Merino, C. Reduction of Ascorbate Free Radical by the Plasma Membrane of Synaptic Terminals from Rat Brain. Arch. Biochem. Biophys. 2008, 469, 243–254. [Google Scholar] [CrossRef]
- Cabelli, D.E.; Bielski, B.H.J. Kinetics and Mechanism for the Oxidation of Ascorbic Acid/Ascorbate by HO2/O2- (Hydroperoxyl/Superoxide) Radicals. A Pulse Radiolysis and Stopped-Flow Photolysis Study. J. Phys. Chem. 1983, 87, 1809–1812. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide Dismutases. Annu. Rev. Biochem. 1975, 44, 147–159. [Google Scholar] [CrossRef]
- Dym, O.; Eisenberg, D. Sequence-Structure Analysis of FAD-Containing Proteins. Protein Sci. 2001, 10, 1712–1728. [Google Scholar] [CrossRef]
- Bewley, M.C.; Marohnic, C.C.; Barber, M.J. The Structure and Biochemistry of NADH-Dependent Cytochrome B5 Reductase Are Now Consistent. Biochemistry 2001, 40, 13574–13582. [Google Scholar] [CrossRef] [PubMed]
- Marohnic, C.C.; Crowley, L.J.; Davis, C.A.; Smith, E.T.; Barber, M.J. Cytochrome B5 Reductase: Role of the Si-Face Residues, Proline 92 and Tyrosine 93, in Structure and Catalysis. Biochemistry 2005, 44, 2449–2461. [Google Scholar] [CrossRef]
- Kimura, S.; Emi, Y.; Ikushiro, S.; Iyanagi, T. Systematic Mutations of Highly Conserved His49 and Carboxyl-Terminal of Recombinant Porcine Liver NADH-Cytochrome B5 Reductase Solubilized Domain. Biochim. Biophys. Acta—Protein Struct. Mol. Enzymol. 1999, 1430, 290–301. [Google Scholar] [CrossRef]
- Marohnic, C.C.; Barber, M.J. Arginine 91 Is Not Essential for Flavin Incorporation in Hepatic Cytochrome B5 Reductase. Arch. Biochem. Biophys. 2001, 389, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Kawamura, M.; Iyanagi, T. Role of Thr66 in Porcine NADH-Cytochromeb 5 Reductase in Catalysis and Control of the Rate-Limiting Step in Electron Transfer. J. Biol. Chem. 2003, 278, 3580–3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugler, W.; Pekrun, A.; Laspe, P.; Erdlenbruch, B.; Lakomek, M. Molecular Basis of Recessive Congenital Methemoglobinemia, Types I and II: Exon Skipping and Three Novel Missense Mutations in the NADH-Cytochrome B5 Reductase (Diaphorase 1) Gene. Hum. Mutat. 2001, 17, 348. [Google Scholar] [CrossRef]
- Yamada, M.; Tamada, T.; Takeda, K.; Matsumoto, F.; Ohno, H.; Kosugi, M.; Takaba, K.; Shoyama, Y.; Kimura, S.; Kuroki, R.; et al. Elucidations of the Catalytic Cycle of NADH-Cytochrome B5 Reductase by X-Ray Crystallography: New Insights into Regulation of Efficient Electron Transfer. J. Mol. Biol. 2013, 425, 4295–4306. [Google Scholar] [CrossRef]
- Ainsley Davis, C.; Barber, M.J. Cytochrome B5 Oxidoreductase: Expression and Characterization of the Original Familial Ideopathic Methemoglobinemia Mutations E255- and G291D. Arch. Biochem. Biophys. 2004, 425, 123–132. [Google Scholar] [CrossRef]
- Roma, G.W.; Crowley, L.J.; Davis, C.A.; Barber, M.J. Mutagenesis of Glycine 179 Modulates Both Catalytic Efficiency and Reduced Pyridine Nucleotide Specificity in Cytochrome B5 Reductase. Biochemistry 2005, 44, 13467–13476. [Google Scholar] [CrossRef]
- Iyanagi, T. Molecular Mechanism of Metabolic NAD(P)H-Dependent Electron-Transfer Systems: The Role of Redox Cofactors. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 233–258. [Google Scholar] [CrossRef]
- Baymann, F.; Schoepp-Cothenet, B.; Duval, S.; Guiral, M.; Brugna, M.; Baffert, C.; Russell, M.J.; Nitschke, W. On the Natural History of Flavin-Based Electron Bifurcation. Front. Microbiol. 2018, 9, 1357. [Google Scholar] [CrossRef]
- Christgen, S.L.; Becker, S.M.; Becker, D.F. Chapter one—Methods for determining the reduction potentials of flavin enzymes. In Methods in Enzymology; New Approaches for Flavin Catalysis; Palfey, B.A., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 620, pp. 1–25. [Google Scholar]
- Takaba, K.; Takeda, K.; Kosugi, M.; Tamada, T.; Miki, K. Distribution of Valence Electrons of the Flavin Cofactor in NADH-Cytochrome B5 Reductase. Sci. Rep. 2017, 7, 43162. [Google Scholar] [CrossRef] [PubMed]
- Bonomi, F.; Iametti, S. Redox titration of flavoproteins: An overview. In Flavins and Flavoproteins: Methods and Protocols; Methods in Molecular Biology; Barile, M., Ed.; Springer: New York, NY, USA, 2021; pp. 119–133. ISBN 978-1-07-161286-6. [Google Scholar]
- Ishikita, H.; Saito, K. Proton Transfer Reactions and Hydrogen-Bond Networks in Protein Environments. J. R. Soc. Interface 2014, 11, 20130518. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh-Pasdar, N.; Li-Chan, E.C. Comparison of Protein Surface Hydrophobicity Measured at Various PH Values Using Three Different Fluorescent Probes. J. Agric. Food Chem. 2000, 48, 328–334. Available online: https://pubs.acs.org/doi/10.1021/jf990393p (accessed on 27 June 2021). [CrossRef] [PubMed]
- Li, H.; Chang, Y.-Y.; Lee, J.Y.; Bahar, I.; Yang, L.-W. DynOmics: Dynamics of Structural Proteome and Beyond. Nucleic Acids Res. 2017, 45, W374–W380. [Google Scholar] [CrossRef] [PubMed]
- Samhan-Arias, A.K.; Cordas, C.M.; Carepo, M.S.; Maia, L.B.; Gutierrez-Merino, C.; Moura, I.; Moura, J.J.G. Ligand Accessibility to Heme Cytochrome B5 Coordinating Sphere and Enzymatic Activity Enhancement upon Tyrosine Ionization. J. Biol. Inorg. Chem. 2019, 24, 317–330. [Google Scholar] [CrossRef]
- Samhan-Arias, A.K.; Maia, L.B.; Cordas, C.M.; Moura, I.; Gutierrez-Merino, C.; Moura, J.J.G. Peroxidase-like Activity of Cytochrome B5 Is Triggered upon Hemichrome Formation in Alkaline PH. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 373–378. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Merino, C.; Martínez-Costa, O.H.; Monsalve, M.; Samhan-Arias, A.K. Structural Features of Cytochrome b5–Cytochrome b5 Reductase Complex Formation and Implications for the Intramolecular Dynamics of Cytochrome b5 Reductase. Int. J. Mol. Sci. 2022, 23, 118. https://doi.org/10.3390/ijms23010118
Gutiérrez-Merino C, Martínez-Costa OH, Monsalve M, Samhan-Arias AK. Structural Features of Cytochrome b5–Cytochrome b5 Reductase Complex Formation and Implications for the Intramolecular Dynamics of Cytochrome b5 Reductase. International Journal of Molecular Sciences. 2022; 23(1):118. https://doi.org/10.3390/ijms23010118
Chicago/Turabian StyleGutiérrez-Merino, Carlos, Oscar H. Martínez-Costa, Maria Monsalve, and Alejandro K. Samhan-Arias. 2022. "Structural Features of Cytochrome b5–Cytochrome b5 Reductase Complex Formation and Implications for the Intramolecular Dynamics of Cytochrome b5 Reductase" International Journal of Molecular Sciences 23, no. 1: 118. https://doi.org/10.3390/ijms23010118