
Historical Roles of Selenium and Selenoproteins in Health and Development: The Good, the Bad and the Ugly

 ,
,
Abstract
:1. Introduction
2. Selenocysteine (Sec) tRNA[Ser]Sec
2.1. Transcription of the tRNA[Ser]Sec Gene (Trsp)
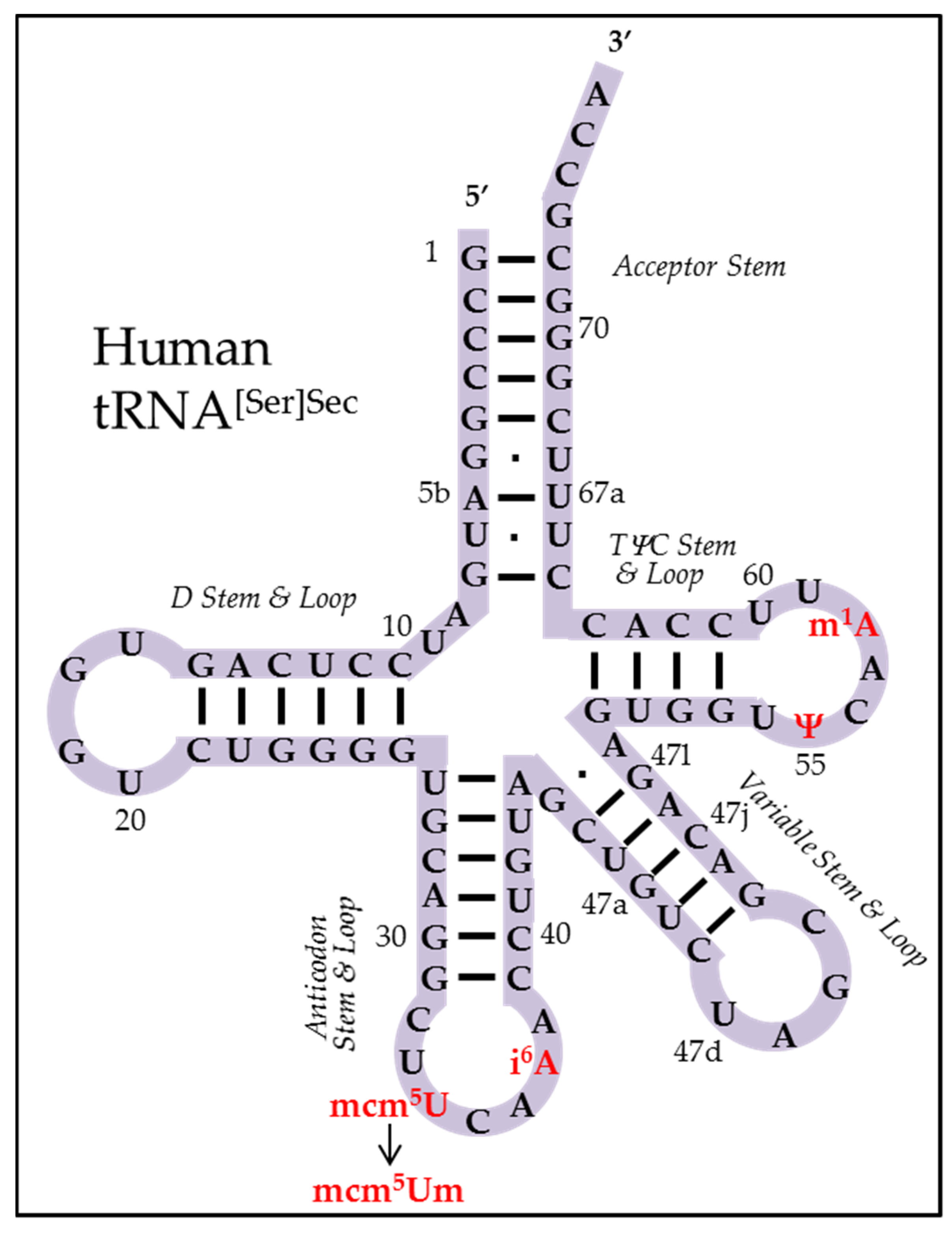
2.2. Primary Sequence of Sec tRNA[Ser]Sec
2.3. The Sec-tRNA[Ser]Sec Population
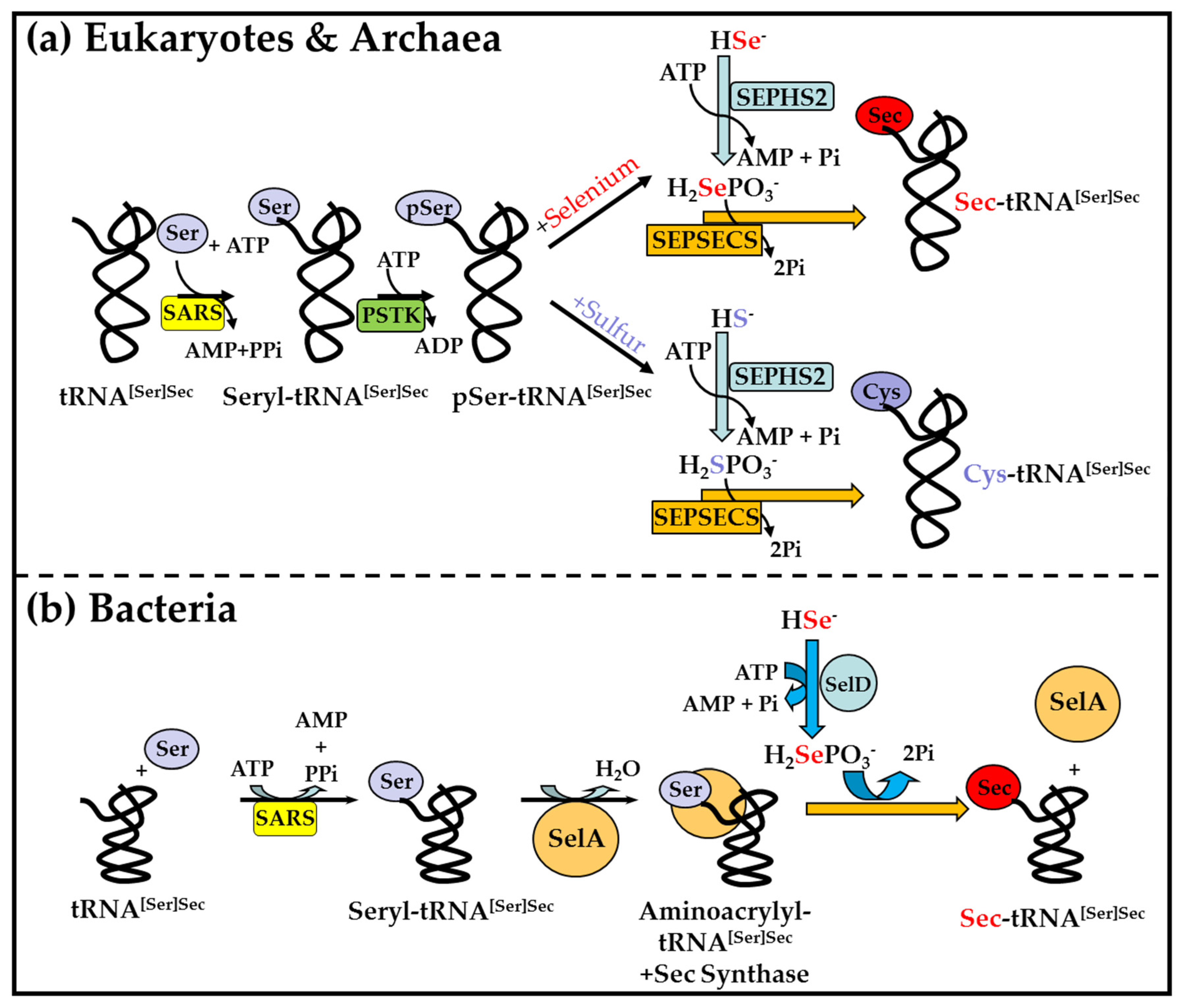
2.4. Biosynthesis of Sec on Sec tRNA[Ser]Sec
2.5. Sec, the 21st Amino Acid in the Genetic Code
3. Selenoproteins
3.1. Mammalian Selenoproteins
3.1.1. Glutathione Peroxidases (GPX)
3.1.2. Thioredoxin Reductases (TXNRD)
3.1.3. Iodothyronine Deiodinases (DIO)
3.1.4. Methionine-R-Sulfoxide Reductase 1 (MSRB1)
3.1.5. Selenophosphate Synthetase 2 (SEPHS2)
3.1.6. Selenoprotein P (SELENOP)
3.1.7. Selenoprotein N (SELENON)
3.1.8. Selenoprotein O (SELENOO)
3.1.9. Selenoprotein I (SELENOI)
3.1.10. Other Selenoproteins
3.2. Phylogenetic Distribution of Selenoproteins
4. Mouse Models
4.1. Trsp Transgenic Mouse Models
4.2. Trsp Conditional Knockout Mouse Models
4.3. Trsp Knockout/Transgenic and Trsp Conditional Knockout/Transgenic Mouse Models
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berzelius, J.J. Undersökning af en ny Mineral-kropp, funnen i de orenare sorterna af det I Falun tillverkade svafl et. Afhandlingar Fysik Kemi Och Mineral. 1818, 6, 42. [Google Scholar]
- Marsden, W. The travels of Marco Polo, the Venetian: The translation of Marsden Revised, with A Selection of His Notes; Wright, T., Ed.; Franklin Classics: London, UK, 1854. [Google Scholar]
- Madison, T.C. Sanitary report—Fort Randall. In Statistical Report on the Sickness and Mortality in the Army of the United States; 36th Congress Senate Executive Document; Coolidge, R.H., Ed.; United States, Surgeon General’s Office: Washington, DC, USA, 1856; pp. 37–41. [Google Scholar]
- Franke, K.W. A new toxicant occurring naturally in certain samples of plant foodstuffs. J. Nutr. 1934, 8, 597. [Google Scholar] [CrossRef]
- Hintz, H.F.; Thompson, L.J. Custer, selenium and swainsonine. Veter Hum. Toxicol. 2000, 42, 242–243. [Google Scholar]
- Schwarz, K.; Foltz, C.M. Factor 3 Activity of Selenium Compounds. J. Biol. Chem. 1958, 233, 245–251. [Google Scholar] [CrossRef]
- Enoch, H.G.; Lester, R.L. Effects of Molybdate, Tungstate, and Selenium Compounds on Formate Dehydrogenase and Other Enzyme Systems in Escherichia coli. J. Bacteriol. 1972, 110, 1032–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatfield, D.L.; Schweizer, U.; Tsuji, P.A.; Gladyshev, V.N. (Eds.) Selenium Its Molecular Biology and Role in Human Health, 4th ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Oldfield, J.E. Selenium: A historical perspective. In Selenium—Its Molecular Biology and Role in Human Health, 2nd ed.; Hatfield, D.L., Berry, M.J., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2006; pp. 1–6. [Google Scholar]
- Combs, G.F.; Yan, L. Status of dietary selenium in cancer prevention. In Selenium—Its Molecular Biology and Role in Human Health, 4th ed.; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; pp. 321–332. [Google Scholar]
- Ge, K.; Xue, A.; Bai, J.; Wang, S. Keshan disease-an endemic cardiomyopathy in China. Virchows Archiv A Pathol. Anat. Histopathol. 1983, 401, 1–15. [Google Scholar] [CrossRef]
- Yu, F.F.; Qi, Z.; Shang, Y.-N.; Ping, Z.-G.; Guo, X. Prevention and control strategies for children Kashin-Beck disease in China: A systematic review and meta-analysis. Medicine 2019, 98, e16823. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, T.; Li, Q.; Li, D. Prevention of Keshan Disease by Selenium Supplementation: A Systematic Review and Meta-analysis. Biol. Trace Element Res. 2018, 186, 98–105. [Google Scholar] [CrossRef]
- Institute of Medicine. Dietary Reference Intakes: Vitamin C, Vitamin E, Selenium, and Carotenoids; Food and Nutrition Board: Washington, DC, USA, 2000. [Google Scholar]
- Peters, K.M.; Galinn, S.E.; Tsuji, P.A. Selenium: Dietary Sources, Human Nutritional Requirements and Intake Across Populations. In Selenium—Its Molecular Biology and Role in Human Health, 4th ed.; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; Volume 1, pp. 295–305. [Google Scholar]
- Diwadkar-Navsariwala, V.; Diamond, A.M. The Link between Selenium and Chemoprevention: A Case for Selenoproteins. J. Nutr. 2004, 134, 2899–2902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in Human Health and Disease. Antioxid. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef]
- Hatfield, D.L.; Tsuji, P.A.; Carlson, B.A.; Gladyshev, V.N. Selenium and selenocysteine: Roles in cancer, health, and development. Trends Biochem. Sci. 2014, 39, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Rayman, M.P. Selenium in cancer prevention: A review of the evidence and mechanism of action. In Proceedings of the Nutrition Society; CABI Publishing: Oxfordshire, UK, 2005; Volume 64, pp. 527–542. [Google Scholar]
- Hoffmann, P.R.; Berry, M.J. The influence of selenium on immune responses. Mol. Nutr. Food Res. 2008, 52, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, Selenoproteins and Viral Infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campa, A.; Shor-Posner, G.; Indachochea, F.; Zhang, G.; Lai, H.; Asthana, D.; Scott, G.B.; Baum, M.K. Mortality risk in selenium-deficient HIV-positive children. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1999, 20, 508–513. [Google Scholar] [CrossRef]
- Xu, X.-M.; Carlson, B.A.; Grimm, T.A.; Kutza, J.; Berry, M.J.; Arreola, R.; Fields, K.H.; Shanmugam, I.; Jeang, K.-T.; Oroszlan, S.; et al. Rhesus Monkey Simian Immunodeficiency Virus Infection as a Model for Assessing the Role of Selenium in AIDS. JAIDS J. Acquir. Immune Defic. Syndr. 2002, 31, 453–463. [Google Scholar] [CrossRef]
- Cai, Z.; Zhang, J.; Li, H. Selenium, aging and aging-related diseases. Aging Clin. Exp. Res. 2018, 31, 1035–1047. [Google Scholar] [CrossRef]
- Bartolini, D.; Sancineto, L.; Fabro de Bem, A.; Tew, K.D.; Santi, C.; Radi, R.; Toquato, P.; Galli, F. Selenocompounds in cancer therapy: An overview. Adv. Cancer Res. 2017, 136, 259–302. [Google Scholar] [PubMed]
- Whanger, P.D. Selenocompounds in Plants and Animals and their Biological Significance. J. Am. Coll. Nutr. 2002, 21, 223–232. [Google Scholar] [CrossRef]
- Ferreira, R.L.U.; Sena-Evangelista, K.C.M.; de Azevedo, E.P.; Pinheiro, F.I.; Cobucci, R.N.; Pedrosa, L.F.C. Selenium in Human Health and Gut Microflora: Bioavailability of Selenocompounds and Relationship with Diseases. Front. Nutr. 2021, 8, 685317. [Google Scholar] [CrossRef] [PubMed]
- Flohe, L.; Günzler, W.; Schock, H. Glutathione peroxidase: A selenoenzyme. FEBS Lett. 1973, 32, 132–134. [Google Scholar] [CrossRef] [Green Version]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical Role as a Component of Glutathione Peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Cone, J.E.; Del Rio, R.M.; Davis, J.N.; Stadtman, T.C. Chemical characterization of the selenoprotein component of clostridial glycine reductase: Identification of selenocysteine as the organoselenium moiety. Proc. Natl. Acad. Sci. USA 1976, 73, 2659–2663. [Google Scholar] [CrossRef] [Green Version]
- Günzler, W.A.; Steffens, G.J.; Grossmann, A.; Kim, S.-M.A.; Ötting, F.; Wendel, A.; Flohé, L. The Amino-Acid Sequence of Bovine Glutathione Peroxidase. Hoppe-Seyler’s Z Physiol. Chem. 1984, 365, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I.; Frampton, J.; Goldfarb, P.; Affara, N.; McBain, W.; Harrison, P. The structure of the mouse glutathione peroxidase gene: The selenocysteine in the active site is encoded by the ‘termination’ codon, TGA. EMBO J. 1986, 5, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Zinoni, F.; Birkmann, A.; Stadtman, T.C.; Bock, A. Nucleotide sequence and expression of the selenocysteine-containing polypeptide of formate dehydrogenase (formate-hydrogen-lyase-linked) from Escherichia coli. Proc. Natl. Acad. Sci. USA 1986, 83, 4650–4654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santesmasses, D.; Mariotti, M.; Guigó, R. Computational identification of the selenocysteine tRNA (tRNASec) in genomes. PLoS Comput. Biol. 2017, 13, e1005383. [Google Scholar] [CrossRef]
- Xu, X.M.; Zhou, X.; Carlson, B.A.; Kim, L.K.; Huh, T.L.; Lee, B.J.; Hatfield, D.L. The zebrafish genome contains two distinct selenocysteine tRNA[Ser]sec genes. FEBS Lett. 1999, 454, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Aeby, E.; Ullu, E.; Yepiskoposyan, H.; Schimanski, B.; Roditi, I.; Mühlemann, O.; Schneider, A. tRNASec is transcribed by RNA polymerase II in Trypanosoma brucei but not in humans. Nucleic Acids Res. 2010, 38, 5833–5843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.M.; Lee, J.Y.; Hatfield, D.L.; Lee, B.J. Differential mode of TBP utilization in transcription of the tRNA[Ser]Sec gene and TATA-less class III genes. Gene 1997, 196, 99–103. [Google Scholar] [CrossRef]
- Park, J.M.; Yang, E.S.; Hatfield, L.L.; Lee, B.J. Analysis of the Selenocysteine tRNA[SER]SEC Gene Transcription in vitro Using Xenopus Oocyte Extracts. Biochem. Biophys. Res. Commun. 1996, 226, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; De-La-Pena-Cortines, P.; Tobian, J.A.; Zasloff, M.; Hatfield, D. Unique pathway of expression of an opal suppressor phosphoserine tRNA. Proc. Natl. Acad. Sci. USA 1987, 84, 6384–6388. [Google Scholar] [CrossRef] [Green Version]
- Carlson, B.A.; Lee, B.C.; Tsuji, P.A.; Tobe, R.; Park, J.M.; Schweizer, U.; Gladyshev, V.N.; Hatfield, D.L. Selenocysteine tRNA[Ser]Sec: From nonsense suppressor tRNA to the quintessential constituent in selenoprotein biosynthesis. In Selenium—Its Molecular Biology and Role in Human Health; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kim, L.K.; Matsufuji, T.; Matsufuji, S.; Carlson, B.A.; Kim, S.S.; Hatfield, D.L.; Lee, B.J. Methylation of the ribosyl moiety at position 34 of selenocysteine tRNA[Ser]Sec is governed by both primary and tertiary structure. RNA 2000, 6, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Chittum, H.S.; Baek, H.J.; Diamond, A.M.; Fernandez-Salguero, P.; Gonzalez, F.; Ohama, T.; Hatfield, D.L.; Kuehn, M.; Lee, B.J. Selenocysteine tRNA[Ser]Sec Levels and Selenium-Dependent Glutathione Peroxidase Activity in Mouse Embryonic Stem Cells Heterozygous for a Targeted Mutation in the tRNA[Ser]Sec Gene. Biochemistry 1997, 36, 8634–8639. [Google Scholar] [CrossRef]
- Carlson, B.A.; Xu, X.-M.; Gladyshev, V.N.; Hatfield, D.L. Selective Rescue of Selenoprotein Expression in Mice Lacking a Highly Specialized Methyl Group in Selenocysteine tRNA. J. Biol. Chem. 2005, 280, 5542–5548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, B.A. Um34 in selenocysteine tRNA is required for the expression of stress-related selenoproteins in mammals. In Fine-tuning of RNA Functions by Modification and Editing; Grosjean, H., Ed.; Topis in Current Genetics; Springer: Berlin/Heidelberg, Germany, 2005; Volume 12, pp. 431–438. [Google Scholar]
- Xu, X.-M.; Carlson, B.A.; Mix, H.; Zhang, Y.; Saira, K.; Glass, R.S.; Berry, M.J.; Gladyshev, V.N.; Hatfield, D.L. Biosynthesis of Selenocysteine on Its tRNA in Eukaryotes. PLoS Biol. 2006, 5, e4. [Google Scholar] [CrossRef]
- Yuan, J.; Palioura, S.; Salazar, J.C.; Su, D.; O’Donoghue, P.; Hohn, M.J.; Cardoso, A.; Whitman, W.; Söll, D. RNA-dependent conversion of phosphoserine forms selenocysteine in eukaryotes and archaea. Proc. Natl. Acad. Sci. USA 2006, 103, 18923–18927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copeland, P.R.; Howard, M.T. Ribosome Fate during Decoding of UGA-Sec Codons. Int. J. Mol. Sci. 2021, 22, 13204. [Google Scholar] [CrossRef]
- Zhang, Y.; Gladyshev, V.N. Comparative Genomics of Trace Element Dependence in Biology. J. Biol. Chem. 2011, 286, 23623–23629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, M.J.; Banu, L.; Chen, Y.; Mandel, S.J.; Kieffer, J.D.; Harney, J.W.; Larsen, P.R. Recognition of UGA as a selenocysteine codon in Type I deiodinase requires sequences in the 3′ untranslated region. Nat. Cell Biol. 1991, 353, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, M.; Ridge, P.G.; Zhang, Y.; Lobanov, A.V.; Pringle, T.H.; Guigo, R.; Hatfield, D.L.; Gladyshev, V.N. Composition and Evolution of the Vertebrate and Mammalian Selenoproteomes. PLoS ONE 2012, 7, e33066. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular Pathways and Physiological Roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toppo, S.; Vanin, S.; Bosello, V.; Tosatto, S.C. Evolutionary and Structural Insights into the Multifaceted Glutathione Peroxidase (Gpx) Superfamily. Antioxid. Redox Signal. 2008, 10, 1501–1514. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Hirao, F.; Sakamoto, T.; Sekine, K.; Mizukura, Y.; Saito, M.; Kitamoto, T.; Hayasaka, M.; Hanaoka, K.; Nakagawa, Y. Early embryonic lethality caused by targeted disruption of the mouse PHGPx gene. Biochem. Biophys. Res. Commun. 2003, 305, 278–286. [Google Scholar] [CrossRef]
- Yant, L.; Ran, Q.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free. Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef]
- Ingold, I.; Berndt, C.; Schmitt, S.; Doll, S.; Poschmann, G.; Buday, K.; Roveri, A.; Peng, X.; Porto Freitas, F.P.; Seibt, T.; et al. Selenium Utilization by GPX4 Is Required to Prevent Hydroperoxide-Induced Ferroptosis. Cell 2017, 172, 409–422.e21. [Google Scholar] [CrossRef] [Green Version]
- Stockwell, B.R.; Angeli, J.P.F.; Bayir, H.; Bush, A.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnér, E.S. Focus on mammalian thioredoxin reductases—Important selenoproteins with versatile functions. Biochim. Biophys. Acta BBA Gen. Subj. 2009, 1790, 495–526. [Google Scholar] [CrossRef]
- Conrad, M.; Jakupoglu, C.; Moreno, S.; Lippl, S.; Banjac, A.; Schneider, M.; Beck, H.; Hatzopoulos, A.K.; Just, U.; Sinowatz, F.; et al. Essential Role for Mitochondrial Thioredoxin Reductase in Hematopoiesis, Heart Development, and Heart Function. Mol. Cell. Biol. 2004, 24, 9414–9423. [Google Scholar] [CrossRef] [Green Version]
- Jakupoglu, C.; Przemeck, G.K.H.; Schneider, M.; Moreno, S.; Mayr, N.; Hatzopoulos, A.K.; de Angelis, M.H.; Wurst, W.; Bornkamm, G.W.; Brielmeier, M.; et al. Cytoplasmic Thioredoxin Reductase Is Essential for Embryogenesis but Dispensable for Cardiac Development. Mol. Cell. Biol. 2005, 25, 1980–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.-A.; Kirnarsky, L.; Sherman, S.; Gladyshev, V.N. Selenoprotein oxidoreductase with specificity for thioredoxin and glutathione systems. Proc. Natl. Acad. Sci. USA 2001, 98, 3673–3678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, A.C.; Salvatore, D.; Gereben, B.; Berry, M.J.; Larsen, P.R. Biochemistry, Cellular and Molecular Biology, and Physiological Roles of the Iodothyronine Selenodeiodinases. Endocr. Rev. 2002, 23, 38–89. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.R.; Dick, T.E.; Markovitz, B.P.; Kaplan, M.M.; Gard, T.G. Inhibition of intrapituitary thyroxine to 3.5.3′-triiodothyronine conversion prevents the acute suppression of thyrotropin release by thyroxine in hypothyroid rats. J. Clin. Investig. 1979, 64, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.E.; Karaczyn, A.; Stohn, J.P.; Donnelly, W.T.; Croteau, W.; Peeters, R.P.; Galton, V.A.; Forrest, D.; St Germain, D.; Hernandez, A. The Type 3 Deiodinase Is a Critical Determinant of Appropriate Thyroid Hormone Action in the Developing Testis. Endocrinology 2016, 157, 1276–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.J.; Fiering, S.N.; Pallud, S.E.; Parlow, A.F.; Germain, D.L.S.; Galton, V.A. Targeted Disruption of the Type 2 Selenodeiodinase Gene (DIO2) Results in a Phenotype of Pituitary Resistance to T4. Mol. Endocrinol. 2001, 15, 2137–2148. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.J.; Fiering, S.N.; Thai, B.; Wu, S.-Y.; Germain, E.S.; Parlow, A.F.; Germain, D.L.S.; Galton, V.A. Targeted Disruption of the Type 1 Selenodeiodinase Gene (Dio1) Results in Marked Changes in Thyroid Hormone Economy in Mice. Endocrinology 2006, 147, 580–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryukov, G.; Kryukov, V.M.; Gladyshev, V.N. New Mammalian Selenocysteine-containing Proteins Identified with an Algorithm That Searches for Selenocysteine Insertion Sequence Elements. J. Biol. Chem. 1999, 274, 33888–33897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescure, A.; Gautheret, D.; Carbon, P.; Krol, A. Novel Selenoproteins Identified in Silico and in Vivo by Using a Conserved RNA Structural Motif. J. Biol. Chem. 1999, 274, 38147–38154. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-Y.; Gladyshev, V.N. Methionine Sulfoxide Reduction in Mammals: Characterization of Methionine-R-Sulfoxide Reductases. Mol. Biol. Cell 2004, 15, 1055–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veres, Z.; Kim, I.; Scholz, T.; Stadtman, T.; Veres, Z.; Kim, I.; Scholz, T.; Stadtman, T. Selenophosphate synthetase. Enzyme properties and catalytic reaction. J. Biol. Chem. 1994, 269, 10597–10603. [Google Scholar] [CrossRef]
- Ching, W.-M.; Wittwer, A.J.; Tsai, L.; Stadtman, T.C. Distribution of two selenonucleosides among the selenium-containing tRNAs from Methanococcus vannielii. Proc. Natl. Acad. Sci. USA 1984, 81, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Payne, N.C.; Geissler, A.; Button, A.; Sasuclark, A.R.; Schroll, A.L.; Ruggles, E.L.; Gladyshev, V.N.; Hondal, R.J. Comparison of the redox chemistry of sulfur- and selenium-containing analogs of uracil. Free. Radic. Biol. Med. 2017, 104, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haft, D.H.; Self, W.T. Orphan SelD proteins and selenium-dependent molybdenum hydroxylases. Biol. Direct 2008, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Turanov, A.A.; Hatfield, D.L.; Gladyshev, V.N. In silico identification of genes involved in selenium metabolism: Evidence for a third selenium utilization trait. BMC Genom. 2008, 9, 251. [Google Scholar] [CrossRef] [Green Version]
- Ma, C. Animal models of disease. Mod. Drug Discov. 2004, 7, 30–36. [Google Scholar]
- Mariotti, M.; Santesmasses, D.; Guigó, R. Evolution of selenophosphate synthetase. In Selenium—Its Molecular Biology and Role in Human Health, 4th ed.; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; pp. 85–99. [Google Scholar]
- Xu, X.-M.; Carlson, B.A.; Irons, R.; Mix, H.; Zhong, N.; Gladyshev, V.N.; Hatfield, D.L. Selenophosphate synthetase 2 is essential for selenoprotein biosynthesis. Biochem. J. 2007, 404, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsina, B.; Corominas, M.; Berry, M.; Baguna, J.; Serras, F. Disruption of selenoprotein biosynthesis affects cell proliferation in the imaginal discs and brain of Drosophila melanogaster. J. Cell Sci. 1999, 112, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Na, J.; Jung, J.; Bang, J.; Lu, Q.; Carlson, B.A.; Guo, X.; Gladyshev, V.N.; Kim, J.-H.; Hatfield, D.L.; Lee, B.J. Selenophosphate synthetase 1 and its role in redox homeostasis, defense and proliferation. Free. Radic. Biol. Med. 2018, 127, 190–197. [Google Scholar] [CrossRef]
- Chapple, C.E.; Guigó, R. Relaxation of Selective Constraints Causes Independent Selenoprotein Extinction in Insect Genomes. PLoS ONE 2008, 3, e2968. [Google Scholar] [CrossRef] [Green Version]
- Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteinless animals: Selenophosphate synthetase SPS1 functions in a pathway unrelated to selenocysteine biosynthesis. Protein Sci. 2007, 17, 176–182. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Santesmasses, D.; Capella-Gutierrez, S.; Mateo, A.; Arnan, C.; Johnson, R.; D’Aniello, S.; Yim, S.H.; Gladyshev, V.N.; Serras, F.; et al. Evolution of selenophosphate synthetases: Emergence and relocation of function through independent duplications and recurrent subfunctionalization. Genome Res. 2015, 25, 1256–1267. [Google Scholar] [CrossRef] [Green Version]
- Carlson, B.A.; Novoselov, S.V.; Kumaraswamy, E.; Lee, B.J.; Anver, M.R.; Gladyshev, V.N.; Hatfield, D.L. Specific excision of the selenocysteine tRNA[Ser]Sec (Trsp) gene in mouse liver demonstrates an essential role of selenoproteins in liver function. J Biol. Chem. 2004, 279, 8011–8017. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, U.; Schomburg, L.; Köhrle, J. Selenoprotein P and selenium distribution in mammals. In Selenium—Its Molecular Biology and Role in Human Health, 4th ed.; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; pp. 261–274. [Google Scholar]
- Hill, K.E.; Zhou, J.; McMahan, W.J.; Motley, A.K.; Atkins, J.; Gesteland, R.F.; Burk, R.F. Deletion of Selenoprotein P Alters Distribution of Selenium in the Mouse. J. Biol. Chem. 2003, 278, 13640–13646. [Google Scholar] [CrossRef] [Green Version]
- Motsenbocker, M.A.; Tappel, A. A selenocysteine-containing selenium-transport protein in rat plasma. Biochim. Biophys. Acta BBA Gen. Subj. 1982, 719, 147–153. [Google Scholar] [CrossRef]
- Baclaocos, J.; Santesmasses, D.; Mariotti, M.; Bierła, K.; Vetick, M.B.; Lynch, S.; McAllen, R.; Mackrill, J.J.; Loughran, G.; Guigó, R.; et al. Processive Recoding and Metazoan Evolution of Selenoprotein P: Up to 132 UGAs in Molluscs. J. Mol. Biol. 2019, 431, 4381–4407. [Google Scholar] [CrossRef]
- Chernorudskiy, A.; Varone, E.; Colombo, S.F.; Fumagalli, S.; Cagnotto, A.; Cattaneo, A.; Briens, M.; Baltzinger, M.; Kuhn, L.; Bachi, A.; et al. Selenoprotein N is an endoplasmic reticulum calcium sensor that links luminal calcium levels to a redox activity. Proc. Natl. Acad. Sci. USA 2020, 117, 21288–21298. [Google Scholar] [CrossRef] [PubMed]
- Moghadaszadeh, B.; Petit, N.; Jaillard, C.; Brockington, M.; Roy, S.Q.; Merlini, L.; Romero, N.; Estournet, B.; Desguerre, I.; Chaigne, D.; et al. Mutations in SEPN1 cause congenital muscular dystrophy with spinal rigidity and restrictive respiratory syndrome. Nat. Genet. 2001, 29, 17–18. [Google Scholar] [CrossRef]
- Villar-Quiles, R.N.; von der Hagen, M.; Métay, C.; Gonzalez, V.; Donkervoort, S.; Bertini, E.; Castiglioni, C.; Chaigne, D.; Colomer, J.; Cuadrado, M.L.; et al. The clinical, histologic, and genotypic spectrum of SEPN1-related myopathy. Neurology 2020, 95, e1512–e1527. [Google Scholar] [CrossRef]
- Allamand, V.; Richard, P.; Lescure, A.; Ledeuil, C.; Desjardin, D.; Petit, N.; Gartioux, C.; Ferreiro, A.; Krol, A.; Pellegrini, N.; et al. A single homozygous point mutation in a 3′untranslated region motif of selenoprotein N mRNA causes SEPN1-related myopathy. EMBO Rep. 2006, 7, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, B.; Arbogast, S.; Moyle, M.W.; Anderson, C.B.; Richard, P.; Guicheney, P.; Ferreiro, A.; Flanigan, K.; Howard, M.T. A mutation in the SEPN1 selenocysteine redefinition element (SRE) reduces selenocysteine incorporation and leads toSEPN1-related myopathy. Hum. Mutat. 2009, 30, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of Mammalian Selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [Green Version]
- Dudkiewicz, M.; Szczepińska, T.; Grynberg, M.; Pawłowski, K. A Novel Protein Kinase-Like Domain in a Selenoprotein, Widespread in the Tree of Life. PLoS ONE 2012, 7, e32138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreelatha, A.; Yee, S.S.; Lopez, V.A.; Park, B.C.; Kinch, L.N.; Pilch, S.; Servage, K.; Zhang, J.; Jiou, J.; Karasiewicz-Urbańska, M.; et al. Protein AMPylation by an Evolutionarily Conserved Pseudokinase. Cell 2018, 175, 809–821.e19. [Google Scholar] [CrossRef] [Green Version]
- Han, S.-J.; Lee, B.C.; Yim, S.H.; Gladyshev, V.N.; Lee, S.-R. Characterization of Mammalian Selenoprotein O: A Redox-Active Mitochondrial Protein. PLoS ONE 2014, 9, e95518. [Google Scholar] [CrossRef] [Green Version]
- Avery, J.C.; Yamazaki, Y.; Hoffmann, F.W.; Folgelgren, B.; Hoffmann, P.R. Selenoprotein I is essential for murine embryogenesis. Arch. Biochem. Biophys. 2020, 689, 108444. [Google Scholar] [CrossRef]
- Horibata, Y.; Hirabayashi, Y. Identification and characterization of human ethanolaminephosphotransferase. J. Lipid Res. 2007, 48, 503–508. [Google Scholar] [CrossRef] [Green Version]
- Horibata, Y.; Ando, H.; Sugimoto, H. Locations and contributions of the phosphotransferases EPT1 and CEPT1 to the biosynthesis of ethanolamine phospholipids. J. Lipid Res. 2020, 61, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Horibata, Y.; Elpeleg, O.; Eran, A.; Hirabayashi, Y.; Savitzki, D.; Tal, G.; Mandel, H.; Sugimoto, H. EPT1 (selenoprotein I) is critical for the neural development and maintenance of plasmalogen in humans. J. Lipid Res. 2018, 59, 1015–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.Y.; Al-Khayat, A.; Al-Murshedi, F.; Al-Futaisi, A.; Chioza, B.A.; Fernandez-Murray, J.P.; Self, J.E.; Salter, C.G.; Harlalka, G.V.; Rawlins, L.E.; et al. A mutation ofEPT1 (SELENOI)underlies a new disorder of Kennedy pathway phospholipid biosynthesis. Brain 2017, 140, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Dikiy, A.; Novoselov, S.V.; Fomenko, D.E.; Sengupta, A.; Carlson, B.A.; Cerny, R.L.; Ginalski, K.; Grishin, N.V.; Hatfield, D.L.; Gladyshev, V.N. SelT, SelW, SelH, and Rdx12: Genomics and Molecular Insights into the Functions of Selenoproteins of a Novel Thioredoxin-like Family. Biochemistry 2007, 46, 6871–6882. [Google Scholar] [CrossRef]
- Kasaikina, M.V.; Fomenko, D.E.; Labunskyy, V.M.; Lachke, S.A.; Qiu, W.; Moncaster, J.A.; Zhang, J.; Wojnarowicz, M.W.; Natarajan, S.K.; Malinouski, M.; et al. Roles of the 15-kDa Selenoprotein (Sep15) in Redox Homeostasis and Cataract Development Revealed by the Analysis of Sep 15 Knockout Mice. J. Biol. Chem. 2011, 286, 33203–33212. [Google Scholar] [CrossRef] [Green Version]
- Canter, J.A.; Ernst, S.E.; Peters, K.M.; Carlson, B.A.; Thielman, N.R.J.; Grysczyk, L.; Udofe, P.; Yu, Y.; Cao, L.; Davis, C.D.; et al. Selenium and the 15kDa Selenoprotein Impact Colorectal Tumorigenesis by Modulating Intestinal Barrier Integrity. Int. J. Mol. Sci. 2021, 22, 10651. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, P.A.; Carlson, B.A.; Naranjo-Suarez, S.; Yoo, M.-H.; Xu, X.-M.; Fomenko, D.E.; Gladyshev, V.N.; Hatfield, D.L.; Davis, C.D. Knockout of the 15 kDa Selenoprotein Protects against Chemically-Induced Aberrant Crypt Formation in Mice. PLoS ONE 2012, 7, e50574. [Google Scholar] [CrossRef]
- Carlson, B.A.; Hartman, J.M.; Tsuji, P.A. The 15 kDa Selenoprotein: Insights into Its Regulation and Function. In Selenium—Its Molecular Biology and Role in Human Health, 4th ed.; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; pp. 235–243. [Google Scholar]
- Gong, T.; Berry, M.J.; Pitts, M.W. Selenoprotein M: Structure, Expression and Functional Relevance. In Selenium—Its Molecular Biology and Role in Human Health, 4th ed.; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016; pp. 253–260. [Google Scholar]
- Reeves, M.A.; Bellinger, F.P.; Berry, M.J. The Neuroprotective Functions of Selenoprotein M and its Role in Cytosolic Calcium Regulation. Antioxid. Redox Signal. 2010, 12, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Pitts, M.W.; Reeves, M.A.; Hashimoto, A.C.; Ogawa, A.; Kremer, P.; Seale, L.A.; Berry, M.J. Deletion of Selenoprotein M Leads to Obesity without Cognitive Deficits. J. Biol. Chem. 2013, 288, 26121–26134. [Google Scholar] [CrossRef] [Green Version]
- Fredericks, G.J.; Hoffmann, F.W.; Hondal, R.J.; Rozovsky, S.; Urschitz, J.; Hoffmann, P.R. Selenoprotein K Increases Efficiency of DHHC6 Catalyzed Protein Palmitoylation by Stabilizing the Acyl-DHHC6 Intermediate. Antioxidants 2017, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, T.; Lin, J.; Xu, Y.-Z.; Zhang, Y. Comparative genomics reveals new evolutionary and ecological patterns of selenium utilization in bacteria. ISME J. 2016, 10, 2048–2059. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Peng, T.; Jiang, L.; Ni, J.-Z.; Liu, Q.; Chen, L.; Zhang, Y. Comparative genomics reveals new candidate genes involved in selenium metabolism in prokaryotes. Genome Biol. Evol. 2015, 7, 664–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Makarova, K.S.; Huang, W.-C.; Wolf, Y.I.; Nikolskaya, A.N.; Zhang, X.; Cai, M.; Zhang, C.-J.; Xu, W.; Luo, Z.; et al. Expanded diversity of Asgard archaea and their relationships with eukaryotes. Nat. Cell Biol. 2021, 593, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, M.; Lobanov, A.V.; Manta, B.; Santesmasses, D.; Bofill, A.; Guigó, R.; Gabaldón, T.; Gladyshev, V.N. Lokiarchaeota Marks the Transition between the Archaeal and Eukaryotic Selenocysteine Encoding Systems. Mol. Biol. Evol. 2016, 33, 2441–2453. [Google Scholar] [CrossRef] [Green Version]
- Lobanov, A.V.; Fomenko, D.E.; Zhang, Y.; Sengupta, A.; Hatfield, D.L.; Gladyshev, V.N. Evolutionary dynamics of eukaryotic selenoproteomes: Large selenoproteomes may associate with aquatic life and small with terrestrial life. Genome Biol. 2007, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Eukaryotic selenoproteins and selenoproteomes. Biochim. Biophys. Acta BBA Gen. Subj. 2009, 1790, 1424–1428. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Lu, Y.; Zheng, L.; Li, G.; Chen, L.; Zhang, M.; Ni, J.; Liu, Q.; Zhang, Y. The algal selenoproteomes. BMC Genom. 2020, 21, 1–16. [Google Scholar] [CrossRef]
- Liang, H.; Wei, T.; Xu, Y.; Li, L.; Sahu, S.K.; Wang, H.; Li, H.; Fu, X.; Zhang, G.; Melkonian, M.; et al. Phylogenomics Provides New Insights into Gains and Losses of Selenoproteins among Archaeplastida. Int. J. Mol. Sci. 2019, 20, 3020. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Salinas, G.; Gabaldón, T.; Gladyshev, V.N. Utilization of selenocysteine in early-branching fungal phyla. Nat. Microbiol. 2019, 4, 759–765. [Google Scholar] [CrossRef]
- Mariotti, M. Selenocysteine extinction in insect. In Short Views on Insect Genomics and Proteomics; Springer: Cham, Switzerland, 2016; Volume 4, pp. 113–140. [Google Scholar]
- Rispe, C.; Legeai, F.; Nabity, P.D.; Fernández, R.; Arora, A.K.; Baa-Puyoulet, P.; Banfill, C.R.; Bao, L.; Barberà, M.; Bouallègue, M.; et al. The genome sequence of the grape phylloxera provides insights into the evolution, adaptation, and invasion routes of an iconic pest. BMC Biol. 2020, 18, 90. [Google Scholar] [CrossRef]
- Otero, L.; Romanelli-Cedrez, L.; Turanov, A.A.; Gladyshev, V.N.; Miranda-Vizuete, A.; Salinas, G. Adjustments, extinction, and remains of selenocysteine incorporation machinery in the nematode lineage. RNA 2014, 20, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Bösl, M.R.; Takaku, K.; Oshima, M.; Nishimura, S.; Taketo, M.M. Early embryonic lethality caused by targeted disruption of the mouse selenocysteine tRNA gene (Trsp). Proc. Natl. Acad. Sci. USA 1997, 94, 5531–5534. [Google Scholar] [CrossRef] [Green Version]
- Kumaraswamy, E.; Carlson, B.A.; Morgan, F.; Miyoshi, K.; Robinson, G.W.; Su, D.; Wang, S.; Southon, E.; Tessarollo, L.; Lee, B.J.; et al. Selective removal of the selenocysteine tRNA [Ser]Sec gene (Trsp) in mouse mammary epithelium. Mol. Cell Biol. 2003, 23, 1477–1488. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, M.E.; Carlson, B.A.; El-Saadani, M.A.; Kryukov, G.V.; Sun, Q.-A.; Harney, J.W.; Hill, K.E.; Combs, G.F.; Feigenbaum, L.; Mansur, D.B.; et al. Selective Inhibition of Selenocysteine tRNA Maturation and Selenoprotein Synthesis in Transgenic Mice Expressing Isopentenyladenosine-Deficient Selenocysteine tRNA. Mol. Cell. Biol. 2001, 21, 3840–3852. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, M.E.; Kumaraswamy, E.; Zhong, N.; Rao, M.; Carlson, B.A.; Hatfield, D.L. Models for assessing the role of selenoproteins in health. J. Nutr. 2003, 133, 2494S–2496S. [Google Scholar] [CrossRef] [Green Version]
- Hatfield, D.L.; Gladyshev, V.; Park, J.; Park, S.; Chittum, H.; Huh, J.; Carlson, B.; Kim, M.; Moustafa, M.; Lee, B.J. Biosynthesis of selenocysteine and its incorporation into protein as the 21st amino acid. In Comprehensive Natural Products Chemistry; Barton, D., Nakanishi, K., Meth-Cohn, O., Kelly, J.W., Eds.; Amino Acids, Peptides, Porphyrins, and Alkaloids; Pergamon; Elsevier Science: Oxford, UK, 1999; Volume 4, pp. 353–380. [Google Scholar]
- Carlson, B.A.; Moustafa, M.; Sengupta, A.; Schweizer, U.; Shrimali, R.; Rao, M.; Zhong, N.; Wang, S.; Feigenbaum, L.; Lee, B.J.; et al. Selective Restoration of the Selenoprotein Population in a Mouse Hepatocyte Selenoproteinless Background with Different Mutant Selenocysteine tRNAs Lacking Um. J. Biol. Chem. 2007, 282, 32591–32602. [Google Scholar] [CrossRef] [Green Version]
- Irons, R.; Carlson, B.A.; Hatfield, D.L.; Davis, C.D. Both selenoproteins and low molecular weight selenocompounds reduce colon cancer risk in mice with genetically impaired selenoprotein expression. J. Nutr. 2006, 135, 1311–1317. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, P.A.; Zhong, N.; Carlson, B.A.; Perella, C.M.; Hatfield, D.L.; Beck, M.A. Decreased selenoprotein expression alters the immune response during influenzavirus infection in mice. J. Nutr. 2007, 137, 1466–1471. [Google Scholar] [CrossRef]
- Baliga, M.S.; Diwadkar-Navsariwala, V.; Koh, T.; Fayad, R.; Fantuzzi, G.; Diamond, A.M. Selenoprotein deficiency enhances radiation-induced micronucleiformation. Mol. Nutr. Food Res. 2008, 52, 1300–1304. [Google Scholar] [CrossRef] [PubMed]
- Diwadkar-Navsariwala, V.; Prins, G.S.; Swanson, S.M.; Birch, L.A.; Ray, V.H.; Hedayat, S.; Lantvit, D.L.; Diamond, A.M. Selenoprotein deficiency accelerates pros-tate carcinogenesis in a transgenic model. Proc. Natl. Acad. Sci. USA 2006, 103, 8179–8184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labunskyy, V.; Lee, B.C.; Handy, D.; Loscalzo, J.; Hatfield, D.L.; Gladyshev, V.N. Both Maximal Expression of Selenoproteins and Selenoprotein Deficiency Can Promote Development of Type 2 Diabetes-Like Phenotype in Mice. Antioxid. Redox Signal 2011, 14, 2327–2336. [Google Scholar] [CrossRef]
- Shrimali, R.K.; Weaver, J.A.; Miller, G.F.; Starost, M.F.; Carlson, B.A.; Novoselov, S.V.; Kumaraswamy, E.; Gladyshev, V.N.; Hatfield, D.L. Selenoprotein expression is essential in endothelial cell development and cardiac muscle function. Neuromuscul. Disord. 2007, 17, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Downey, C.M.; Horton, C.R.; Carlson, B.A.; Parsons, T.E.; Hatfield, D.L.; Hallgrímsson, B.; Jirik, F.R. Osteo-Chondroprogenitor–Specific Deletion of the Selenocysteine tRNA Gene, Trsp, Leads to Chondronecrosis and Abnormal Skeletal Development: A Putative Model for Kashin-Beck Disease. PLoS Genet. 2009, 5, e1000616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, A.; Lichti, U.F.; Carlson, B.A.; Ryscavage, A.O.; Gladyshev, V.N.; Yuspa, S.H.; Hatfield, D.L. Selenoproteins are essential for proper keratinocyte function andskin development. PLoS ONE 2010, 5, e12249. [Google Scholar] [CrossRef] [Green Version]
- Kawatani, Y.; Suzuki, T.; Shimizu, R.; Kelly, T.K.; Yamamoto, M. Nrf2 and selenoproteins are essential for maintaining oxidative homeostasis in erythrocytes andprotecting against hemolytic anemia. Blood 2011, 117, 986–996. [Google Scholar] [CrossRef] [Green Version]
- Shrimali, R.K.; Irons, R.D.; Carlson, B.A.; Sano, Y.; Gladyshev, V.N.; Park, J.M.; Hatfield, D.L. Selenoproteins Mediate T Cell Immunity through an Antioxidant Mechanism. J. Biol. Chem. 2008, 283, 20181–20185. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Kelly, V.P.; Motohashi, H.; Nakajima, O.; Takahashi, S.; Nishimura, S.; Yamamoto, M. Deletion of the selenocysteine tRNA gene in macrophages and liver results in compensatory gene induction of cytoprotective enzymes by Nrf2. J. Biol. Chem. 2008, 283, 2021–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, E.K.; Conrad, M.; Winterer, J.; Wozny, C.; Carlson, B.A.; Roth, S.; Schmitz, D.; Bornkamm, G.W.; Coppola, V.; Tessarollo, L.; et al. Neuronal selenoprotein expression is required for interneuron development and prevents seizures and neurodegeneration. FASEB J. 2010, 24, 844–852. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, U.; Streckfuß, F.; Pelt, P.; Carlson, B.A.; Hatfield, D.L.; Köhrle, J.; Schomburg, L. Hepatically derived selenoprotein P is a key factor for kidney but not for brain selenium supply. Biochem. J. 2005, 386, 221–226. [Google Scholar] [CrossRef]
- Blauwkamp, M.N.; Yu, J.; Schin, M.A.; Burke, K.A.; Berry, M.J.; Carlson, B.A.; Brosius III, F.C.; Koenig, R.J. Podocyte specific knock out of selenoproteins does not enhance nephropathy in streptozotocin diabetic C57BL/6 mice. BMC Nephrol. 2008, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Chiu-Ugalde, J.; Wirth, E.K.; Klein, M.O.; Sapin, R.; Fradejas-Villar, N.; Renko, K.; Schomburg, L.; Köhrle, J.; Schweizer, U. Thyroid Function Is Maintained Despite Increased Oxidative Stress in Mice Lacking Selenoprotein Biosynthesis in Thyroid Epithelial Cells. Antioxid. Redox Signal. 2012, 17, 902–913. [Google Scholar] [CrossRef] [PubMed]
- Luchman, H.A.; Villemaire, M.L.; Bismar, T.A.; Carlson, B.A.; Jirik, F.R. Prostate epithelium-specific deletion of the selenocysteine tRNA gene Trsp leads to early onset intraepithelial neoplasia. Am. J. Pathol. 2014, 184, 871–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Kim, Y.; Na, J.; Qiao, L.; Bang, J.; Kwon, D.; Yoo, T.-J.; Kang, D.; Kim, L.K.; Carlson, B.A.; et al. Constitutive Oxidative Stress by SEPHS1 Deficiency Induces Endothelial Cell Dysfunction. Int. J. Mol. Sci. 2021, 22, 11646. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Carlson, B.A.; Weaver, J.A.; Novoselov, S.V.; Fomenko, D.E.; Gladyshev, V.N.; Hatfield, D.L. A functional link between housekeeping selenoproteins and phase II enzymes. Biochem. J. 2008, 413, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Malinouski, M.; Kehr, S.; Finney, L.; Vogt, S.; Carlson, B.A.; Seravalli, J.; Jin, R.; Handy, D.E.; Park, T.J.; Loscalzo, J.; et al. High-Resolution Imaging of Selenium in Kidneys: A Localized Selenium Pool Associated with Glutathione Peroxidase Antioxid. Redox Signal. 2012, 16, 185–192. [Google Scholar] [CrossRef]
- Seeher, S.; Atassi, T.; Mahdi, Y.; Carlson, B.A.; Braun, D.; Wirth, E.K.; Klein, M.O.; Reix, N.; Miniard, A.C.; Schomburg, L.; et al. Secisbp2 Is Essential for Embryonic Development and Enhances Selenoprotein Expression. Antioxid. Redox Signal. 2014, 21, 835–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, B.A.; Yoo, M.-H.; Sano, Y.; Sengupta, A.; Kim, J.Y.; Irons, R.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Selenoproteins regulate macrophage invasiveness and extracellular matrix-related gene expression. BMC Immunol. 2009, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Mattmiller, S.A.; Carlson, B.A.; Gandy, J.C.; Sordillo, L.M. Reduced macrophage selenoprotein expression alters oxidized lipid metabolite biosynthesis from arachidonic and linoleic acid. J. Nutr. Biochem. 2014, 25, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, N.; Kudva, A.K.; Patterson, A.D.; Chiaro, C.; Kennett, M.J.; Desai, D.; Amin, S.; Carlson, B.A.; Cantorna, M.T.; Prabhu, K.S. Crucial Role of Macrophage Selenoproteins in Experimental Colitis. J. Immunol. 2014, 193, 3683–3692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayan, V.; Ravindra, K.C.; Liao, C.; Kaushal, N.; Carlson, B.A.; Prabhu, K.S. Epigenetic regulation of inflammatory gene expression in macrophages by selenium. J. Nutr. Biochem. 2015, 26, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Nelson, S.M.; Shay, A.E.; James, J.L.; Carlson, B.A.; Urban, J.; Prabhu, K.S. Selenoprotein Expression in Macrophages Is Critical for Optimal Clearance of Parasitic Helminth Nippostrongylus brasiliensis. J. Biol. Chem. 2016, 291, 2787–2798. [Google Scholar] [CrossRef] [Green Version]
- Hudson, T.S.; Carlson, B.A.; Hoeneroff, M.J.; Young, H.A.; Sordillo, L.; Muller, W.J.; Hatfield, D.L.; Green, J.E. Selenoproteins reduce susceptibility to DMBA-induced mammary carcinogenesis. Carcinogenesis 2012, 33, 1225–1230. [Google Scholar] [CrossRef] [Green Version]
- Wirth, E.K.; Bharathi, B.S.; Hatfield, D.; Conrad, M.; Brielmeier, M.; Schweizer, U. Cerebellar Hypoplasia in Mice Lacking Selenoprotein Biosynthesis in Neurons. Biol. Trace Element Res. 2014, 158, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Carlson, B.A. Selenocysteine tRNA[Ser]Sec mouse models for elucidating roles of selenoproteins in health and development. In Selenium—Its Molecular Biology and Role in Human Health; Hatfield, D.L., Schweizer, U., Tsuji, P.A., Gladyshev, V.N., Eds.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Park, J.M.; Choi, I.S.; Kang, S.G.; Lee, J.Y.; Hatfield, D.L.; Lee, B.J. Upstream promoter elements are sufficient for selenocysteine tRNA[Ser]Sec gene transcription and to determine the transcription start point. Gene 1995, 162, 13–19. [Google Scholar] [CrossRef]
- Carlson, B.A.; Schweizer, U.; Perella, C.; Shrimali, R.K.; Feigenbaum, L.; Shen, L.; Speransky, S.; Floss, T.; Jeong, S.-J.; Watts, J.; et al. The selenocysteine tRNA STAF-binding region is essential for adequate selenocysteine tRNA status, selenoprotein expression and early age survival of mice. Biochem. J. 2009, 418, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Sec tRNA[Ser]Sec | |||||||
|---|---|---|---|---|---|---|---|
| mcm5U | mcm5Um | ||||||
| Cell Line | Selenium Supplementation a | % of Total b | % | % of Total c | % | % of Total d | mcm5Um/ mcm5U e |
| HL-60 | +(chem. defined media) | 9.6 | 38.5 | 3.70 | 61.5 | 5.90 | 1.60 |
| −(chem. defined media) | 7.5 | 61.3 | 4.60 | 38.7 | 2.90 | 0.63 | |
| HL-60 | +(FBS) | 9.4 | 55.3 | 5.20 | 44.7 | 4.20 | 0.81 |
| −(FBS) | 7.4 | 77.0 | 5.70 | 23.0 | 1.70 | 0.30 | |
| CHO | +(FBS) | 1.01 | 45.1 | 0.46 | 54.9 | 0.55 | 1.22 |
| −(FBS) | 0.86 | 56.2 | 0.48 | 43.8 | 0.38 | 0.78 | |
| RMT | +(chem. defined media) | 1.7 | 11.8 | 0.20 | 88.2 | 1.50 | 7.47 |
| −(chem. defined media) | 1.4 | 35.7 | 0.50 | 64.3 | 0.90 | 1.80 | |
| Sec tRNA[Ser]Sec | |||||||
|---|---|---|---|---|---|---|---|
| mcm5U | mcm5Um | ||||||
| Organ | Dietary Selenium Supplementation | % of Total a | % | % of Total b | % | % of Total c | mcm5Um/ mcm5U d |
| Heart | + | 4.3 | 38.1 | 1.64 | 61.9 | 2.66 | 1.62 |
| − | 3.2 | 66.4 | 2.12 | 33.6 | 1.08 | 0.51 | |
| Kidney | + | 7.5 | 33.7 | 2.52 | 66.3 | 4.97 | 1.97 |
| − | 3.7 | 59.2 | 2.19 | 40.8 | 1.51 | 0.69 | |
| Liver | + | 4.5 | 33.3 | 1.50 | 66.7 | 3.00 | 2.00 |
| − | 2.8 | 57.7 | 1.62 | 42.3 | 1.18 | 0.73 | |
| Muscle | + | 1.9 | 38.6 | 0.73 | 61.4 | 1.17 | 1.59 |
| − | 1.5 | 73.3 | 1.10 | 26.7 | 0.40 | 0.35 | |
| Targeted Tissue or Organ 1 | Main Findings Regarding Role of Selenoproteins in Genetically-Altered Mice, Relative to Control Mice in the Study | Cre Promoter |
|---|---|---|
| Endothelial cells | Endothelial cell development/function: embryonic lethal. 14.5 d.p.c. embryos were smaller, more fragile, had poorly or under-developed vascular systems, limbs, head, and tail [133]. | TieTek2-Cre |
| Heart & Skeletal Muscle | Heart disease prevention: mice died from acute myocardial failure 12 days after birth. | MCK-Cre |
| Kidney | No increase in oxidative stress or nephropathy found in podocytes of selenoprotein-deficient mice [141]. | NPHS2-Cre |
| Liver | Liver function: severe hepatocellular degeneration—mice died between 1 and 3 months of age [82]. SELENOP and GPX3 were reduced in serum and kidney, supporting a selenium-transport role for liver-derived SELENOP [140]. Enhanced expression of phase II response genes compensated for loss of hepatic Trsp [145]. Mice used as controls to monitor selenium pools in kidney due to reduction of GPX3 imported from liver [146]. Secisbp2 gene inactivation was less detrimental than Trsp inactivation [147]. | Alb-Cre |
| Macrophages | Immune function: increased oxidative stress and expression of cytoprotective antioxidant and detoxification genes, accumulation of ROS levels, and impaired invasiveness. Altered expression of ECM and fibrosis-associated genes [148]. Balance of pro- and anti-inflammatory oxylipids during inflammation [149]. Selenoproteins protect mice from chemically-induced colitis by alleviating inflammation [150]. Role in epigenetic modulation of pro-inflammatory genes [151]. When infected with N. brasiliensis, selenium-supplemented KO mice showed a complete abrogation in M2-marker expression with a significant increase in intestinal worms and fecal eggs [152]. | LysM-Cre |
| Mammary glands | First Trsp conditional KO mouse, providing an important tool for elucidating the role of selenoproteins in health and development [123]. MMTV-Cre mice treated with DMBA had significantly more tumors, suggesting that selenoproteins protect against carcinogen-induced mammary cancer [153]. | MMTV-Cre; Wap-Cre |
| Neurons | Neuronal function: enhanced neuronal excitation followed by neurodegeneration of hippocampus. Cerebellar hypoplasia associated with degeneration of Purkinje and granule cells. Cerebellar interneurons essentially absent [139]. Selenoproteins required in post-mitotic neurons of the developing cerebellum [154]. | Tal-Cre; CamK-Cre |
| Osteo-chondroprogenitor | Kashin–Beck disease model: mice had post-natal growth retardation, chondrodysplasia, chondronecrosis, and delayed skeletal ossification characteristic of Kashin–Beck disease [134]. | Col2a1-Cre |
| Prostate | Mice developed PIN-like lesions and microinvasive carcinoma by 24 weeks, which were associated with loss of basement membrane, increased cell cycle, and apoptotic activity [143]. | PB-Cre4 |
| Skin | Role in skin and hair follicle development: runt phenotype, premature death, alopecia with flaky and fragile skin, epidermal hyperplasia with disturbed hair cycle, and an early regression of hair follicles [135]. | K14-Cre |
| T-cells | Immune function: reduction of mature T cells and a defect in T-cell-dependent antibody response. Antioxidant hyperproduction and suppression of T cell proliferation in response to T cell receptor stimulation [137]. | LCK-Cre |
| Thyroid | Mice lacking selenoproteins in thyrocytes showed increased oxidative stress in thyroid. Gross morphology remained intact for at least 6 months. Thyroid hormone levels remained normal in knockout mice; thyrotropin levels moderately elevated [142]. | Pax8-Cre; Tg-CreER |
| Target Site | Model Description | Major Findings Observed in Genetically Altered Mice in Comparison to Control Mice |
|---|---|---|
| Whole Mouse | Trsp KO rescued with WT Trsp transgene | Selenoprotein synthesis was completely recovered [125]. |
| Trsp KO rescued with G37 Trsp transgene | Proper base modification in the anticodon is essential, as mutant mice synthesize stress-related selenoproteins very poorly. Male mutant mice show abnormal sperm and reduced fertility; females produced reduced litter size [43]. Trsp KO could not be rescued with A34 mutant transgene most likely due to misreading (see Text). | |
| Whole Mouse | Trsp KO rescued with promoter mutant Trsp transgene | Mice expressed tissue- and organ-specific amounts of tRNA[Ser]Sec. Lower levels of the mcm5Um isoform were observed in promoter mutant Trsp mice. Mice developed a similar neurological phenotype as SELENOP-KO mice and a reduced life span [157]. |
| Liver Alb-Cre | Trsp liver KO rescued with Trsp WT transgene | Selenoprotein synthesis was completely recovered [82]. |
| Trsp liver KO rescued with G37 mutant Trsp transgene | Housekeeping selenoprotein synthesis was recovered while stress-related selenoprotein synthesis was poorly recovered [82]. | |
| Trsp liver KO rescued with A34 mutant Trsp transgene | Housekeeping selenoprotein synthesis was recovered while stress-related selenoprotein synthesis was poorly recovered. Replacement of selenoprotein synthesis in conditional Trsp mutants resulted in normal gene expression of Phase II response enzymes [127,145]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuji, P.A.; Santesmasses, D.; Lee, B.J.; Gladyshev, V.N.; Hatfield, D.L. Historical Roles of Selenium and Selenoproteins in Health and Development: The Good, the Bad and the Ugly. Int. J. Mol. Sci. 2022, 23, 5. https://doi.org/10.3390/ijms23010005
Tsuji PA, Santesmasses D, Lee BJ, Gladyshev VN, Hatfield DL. Historical Roles of Selenium and Selenoproteins in Health and Development: The Good, the Bad and the Ugly. International Journal of Molecular Sciences. 2022; 23(1):5. https://doi.org/10.3390/ijms23010005
Chicago/Turabian StyleTsuji, Petra A., Didac Santesmasses, Byeong J. Lee, Vadim N. Gladyshev, and Dolph L. Hatfield. 2022. "Historical Roles of Selenium and Selenoproteins in Health and Development: The Good, the Bad and the Ugly" International Journal of Molecular Sciences 23, no. 1: 5. https://doi.org/10.3390/ijms23010005