
Comparative Transcriptome Analysis Revealed Two Alternative Splicing bHLHs Account for Flower Color Alteration in Chrysanthemum

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
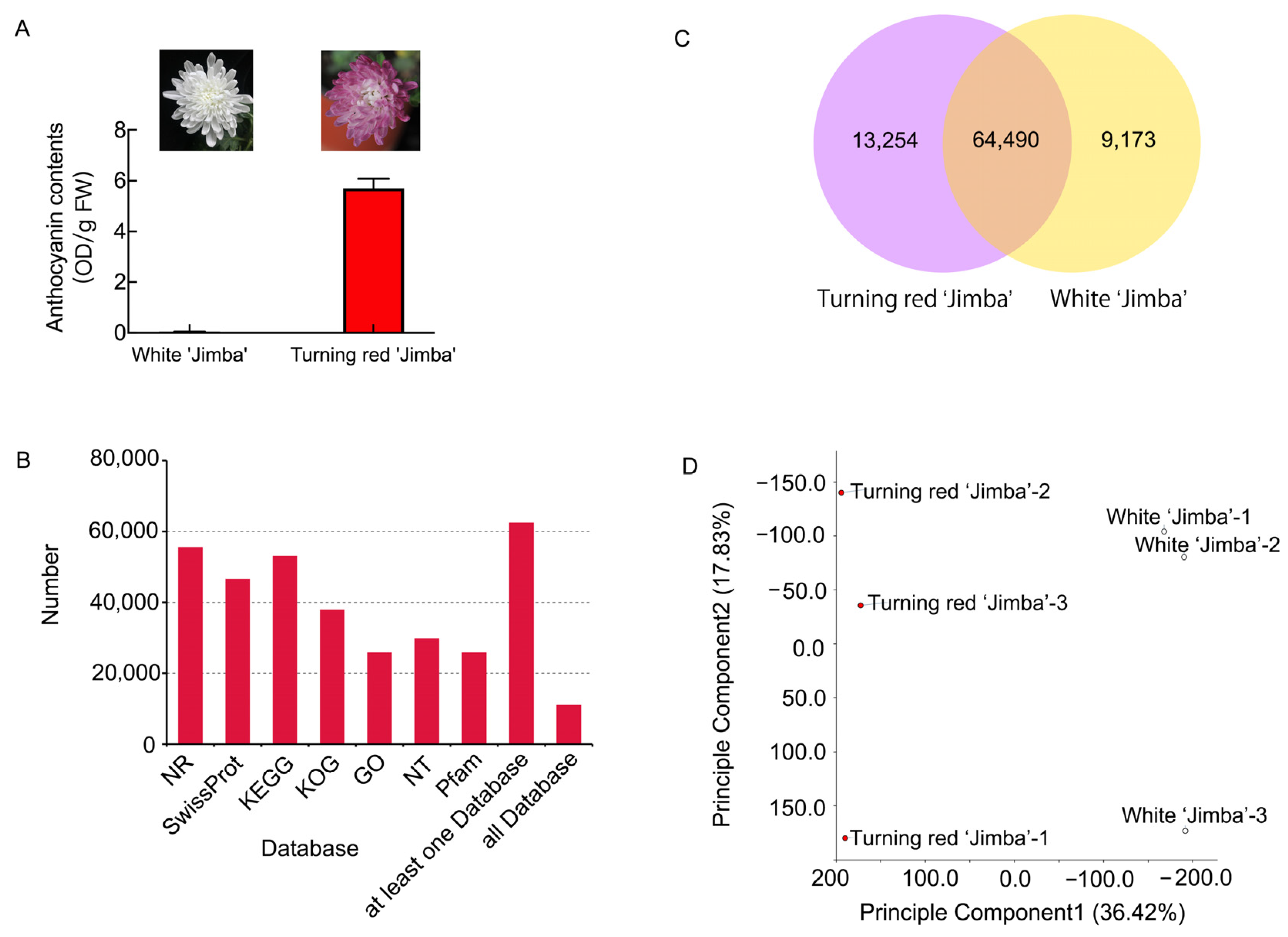
2.1. Anthocyanin Contents in White and Turning Red ‘Jimba’
2.2. Transcriptome Analysis of White and Turning Red ‘Jimba’
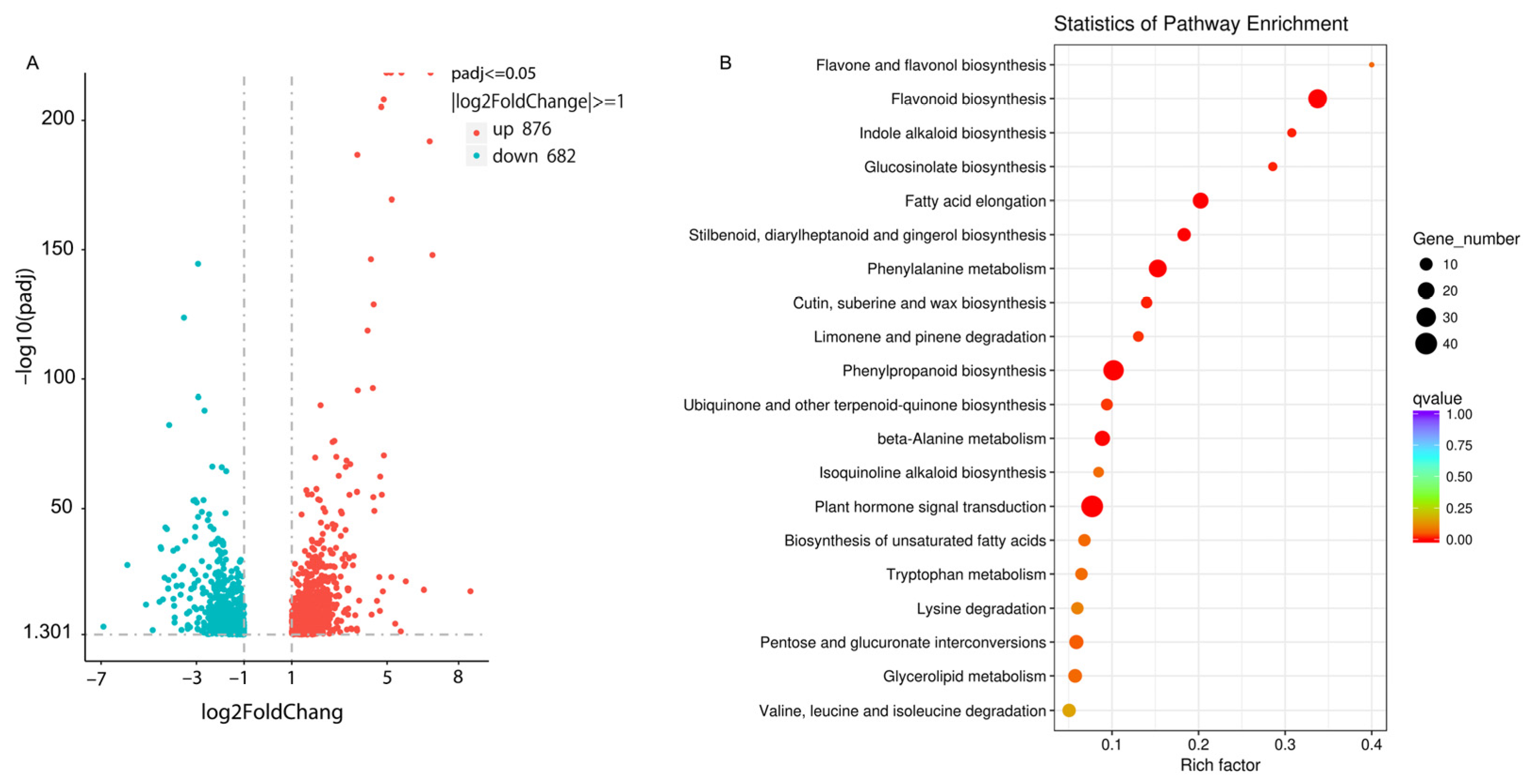
2.3. Differentially Expressed Genes in White ‘Jimba’ and Turning Red ‘Jimba’
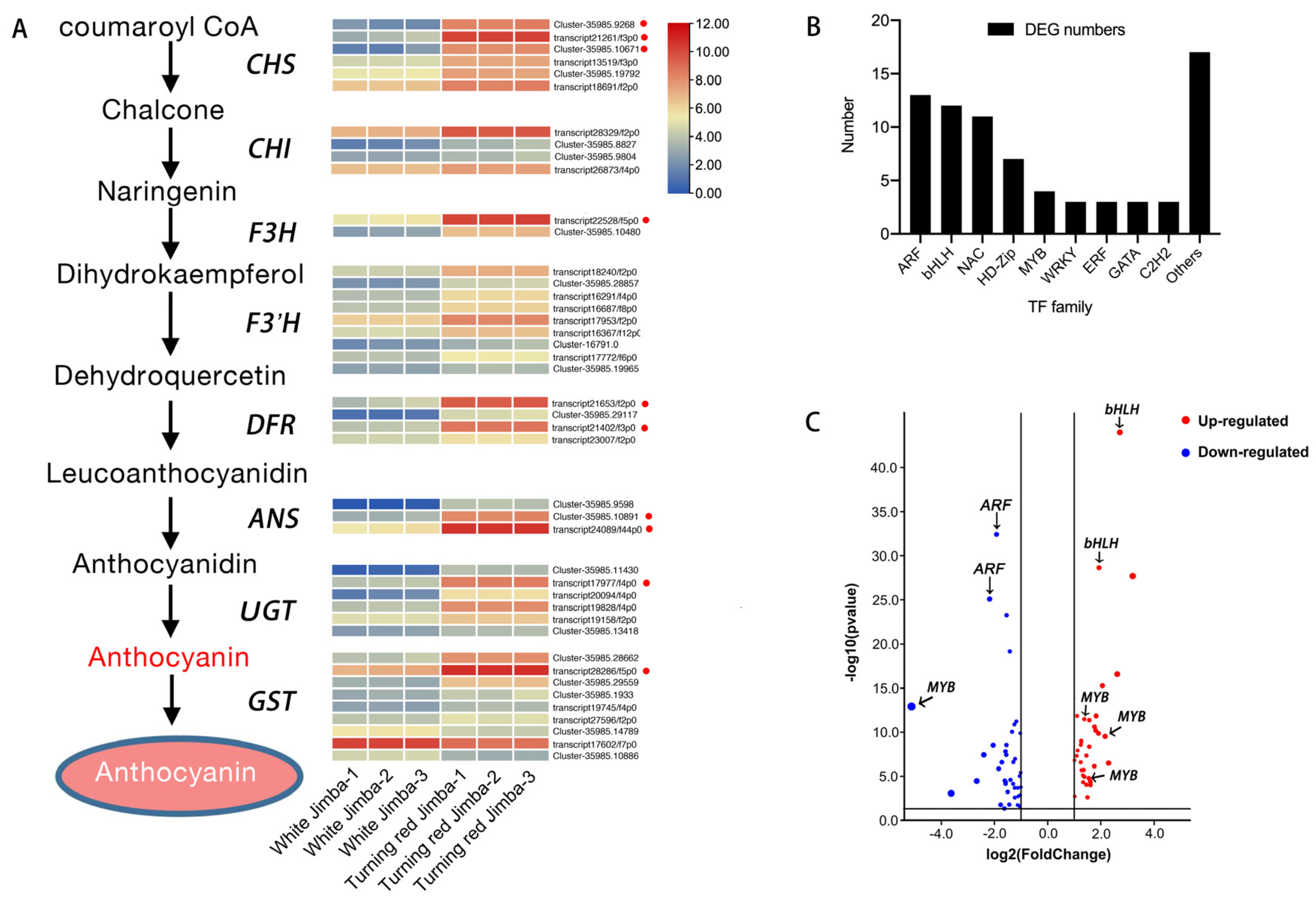
2.4. Differentially Expressed Genes of Anthocyanin Pathway
2.5. Differentially Expressed Transcription Factors
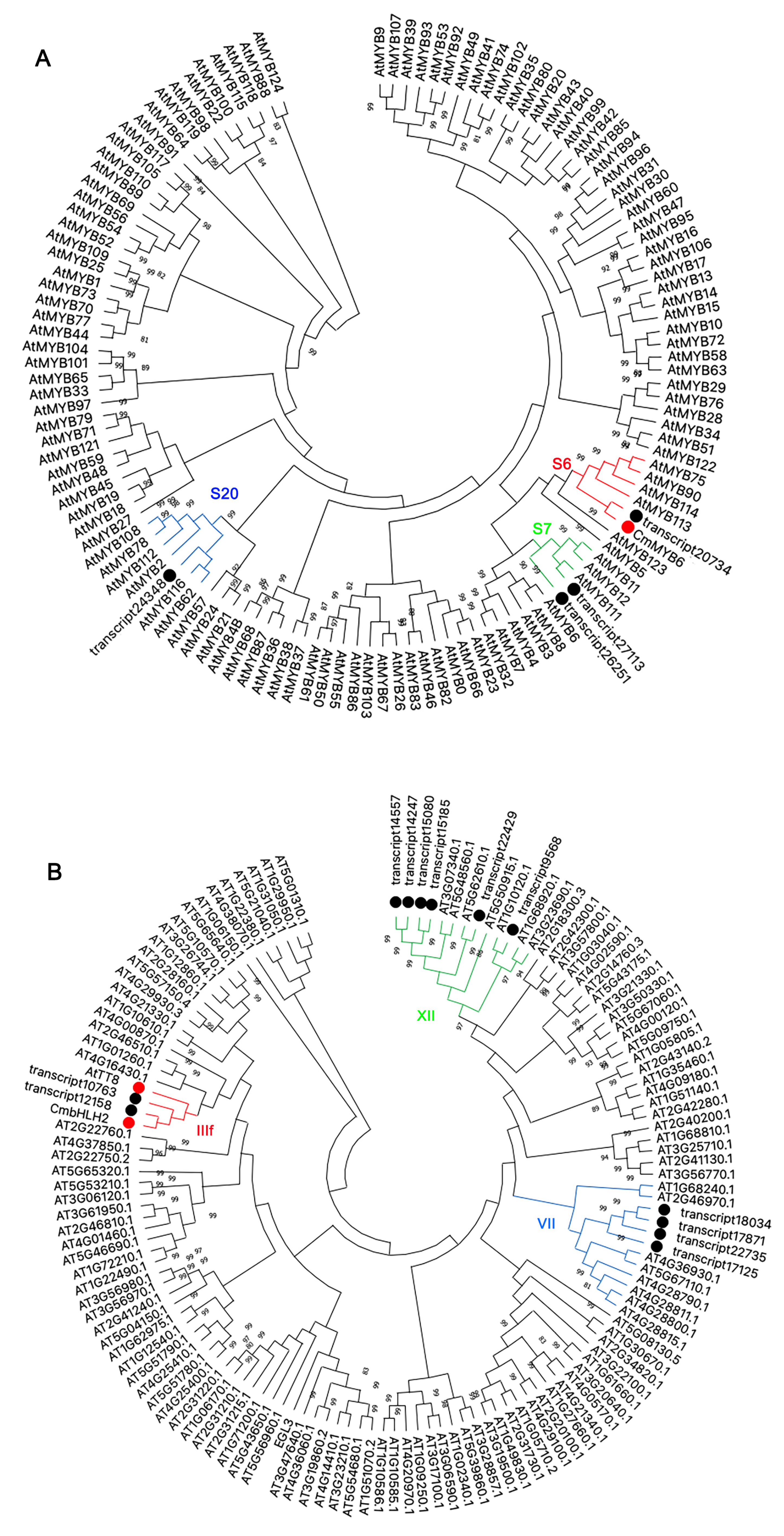
2.6. Phylogenetic Analysis of Differentially Expressed MYBs and bHLHs
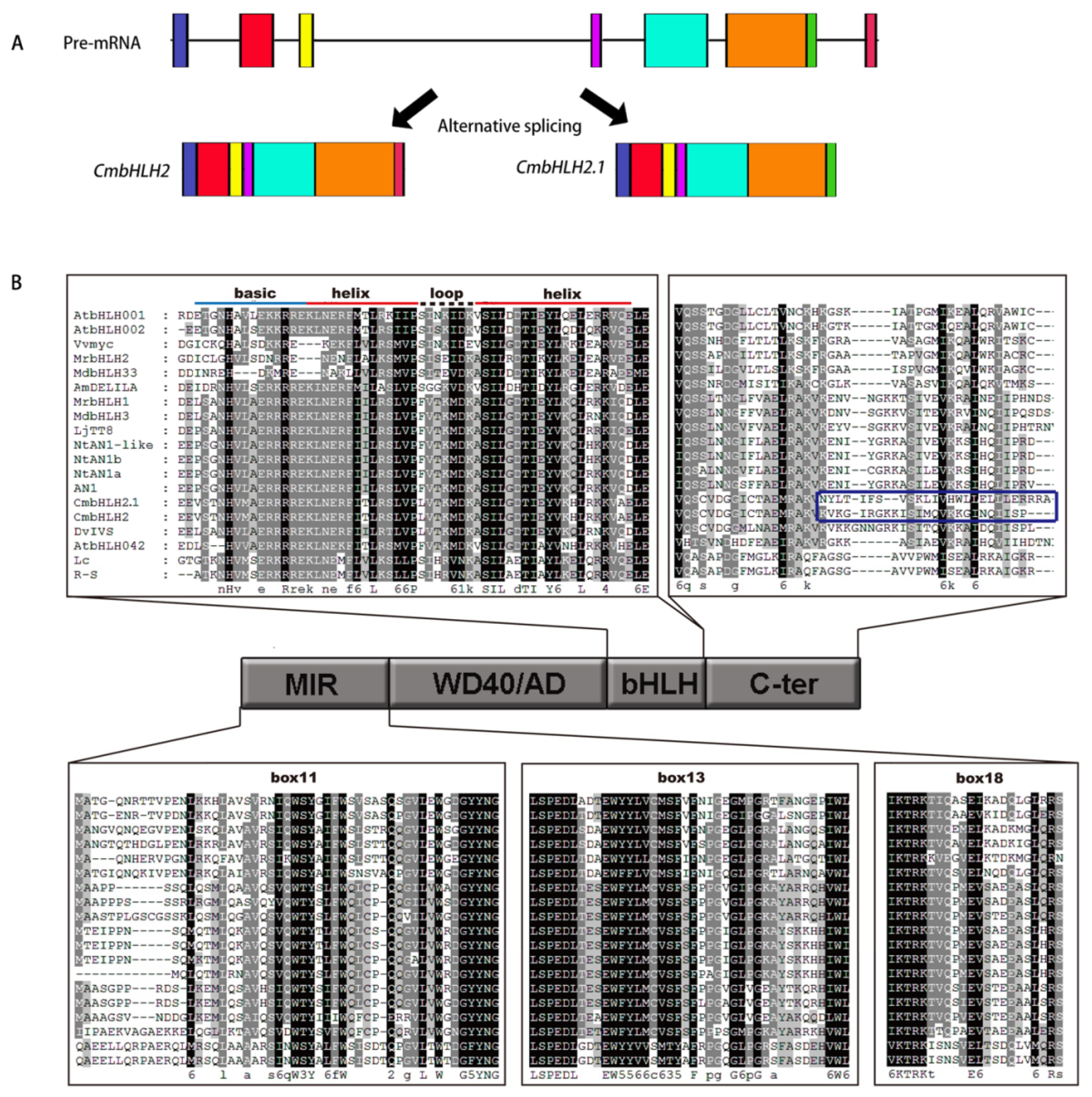
2.7. Full-Length CDS Clone and Sequence Alignment of Anthocyanin-Related MYBs and bHLHs
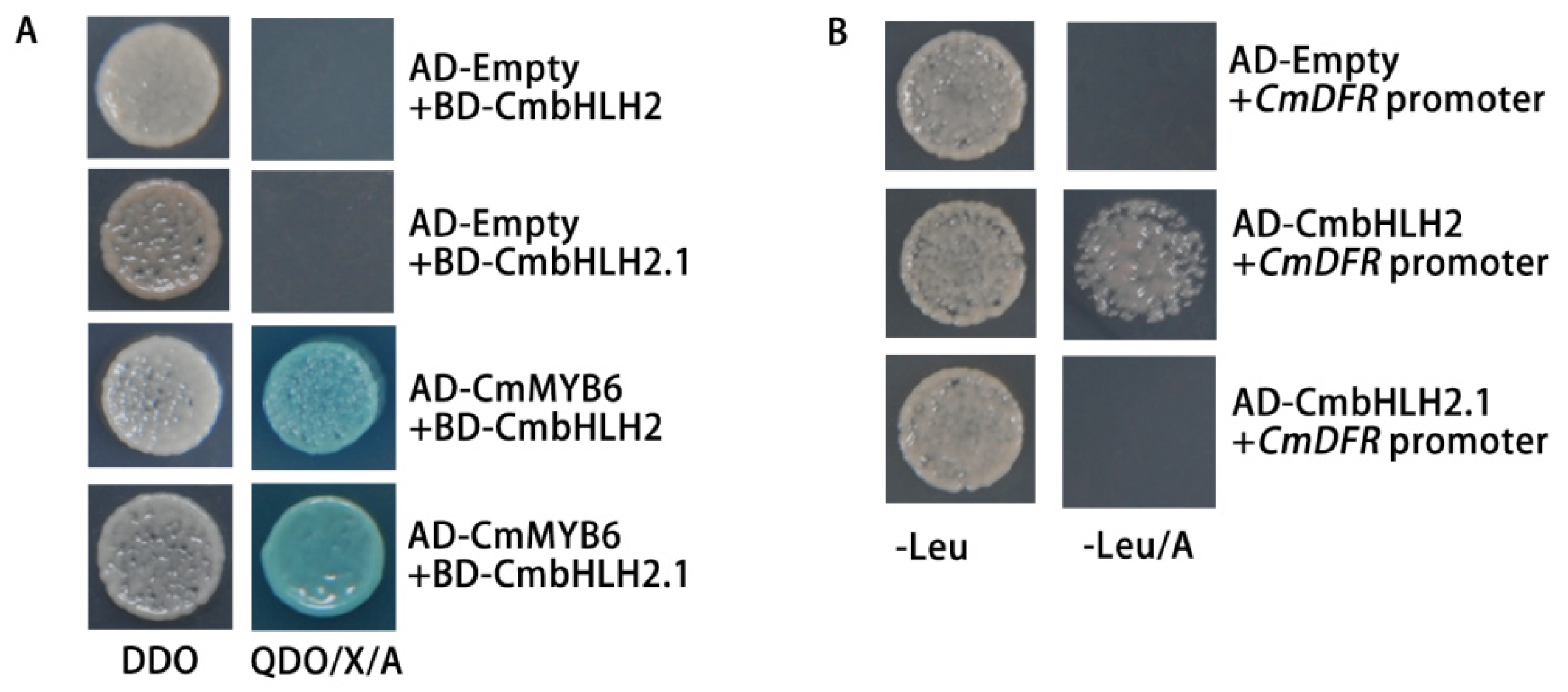
2.8. Analysis of the Protein–Protein Interaction between Regulators and the Protein–DNA Binding Activity of bHLHs on the CmDFR Promoter
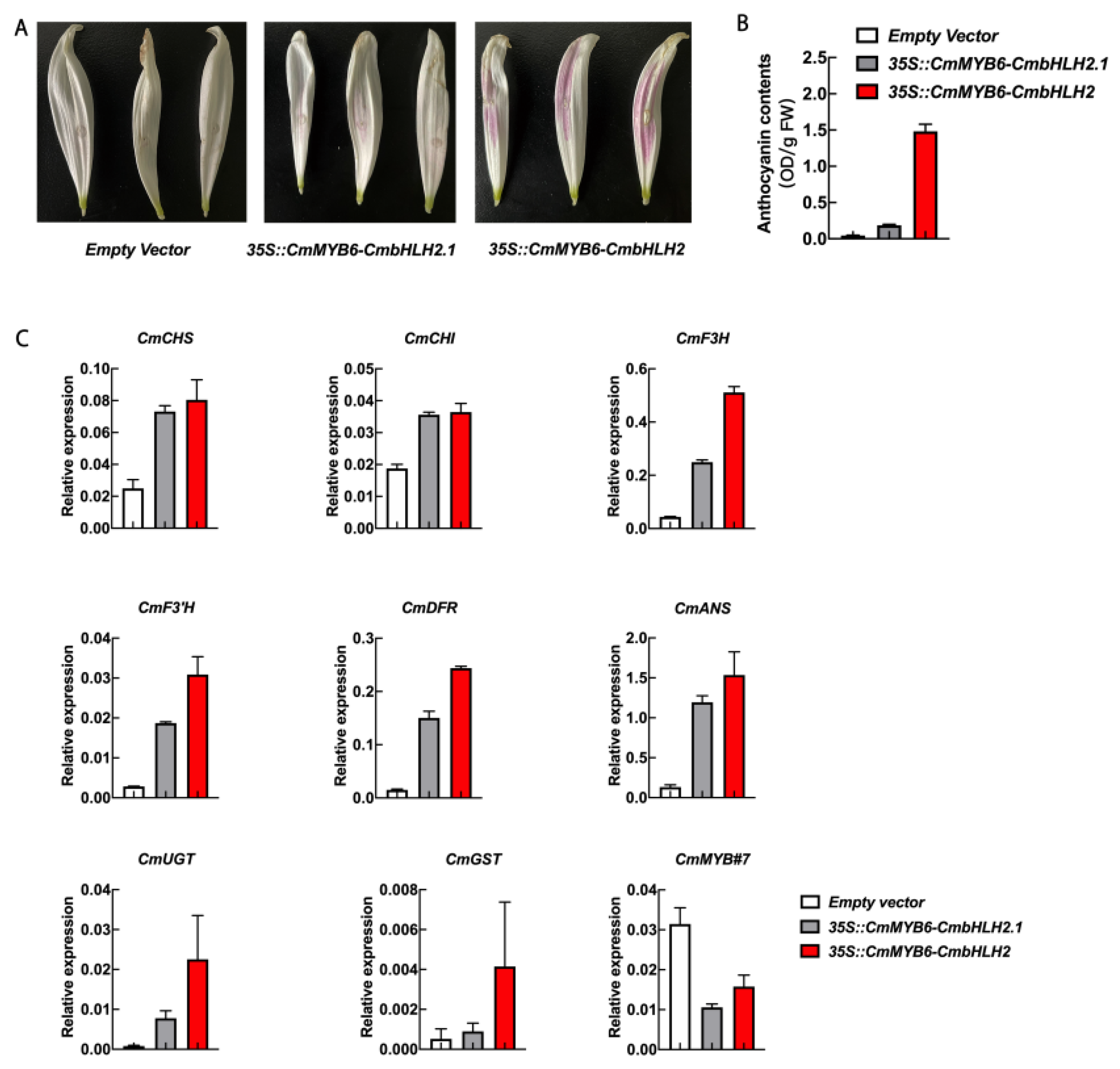
2.9. Transiently Over-Expressed 35S::CmMYB6-CmbHLH2 and 35S::CmMYB6-CmbHLH2.1 in White ‘Jimba’
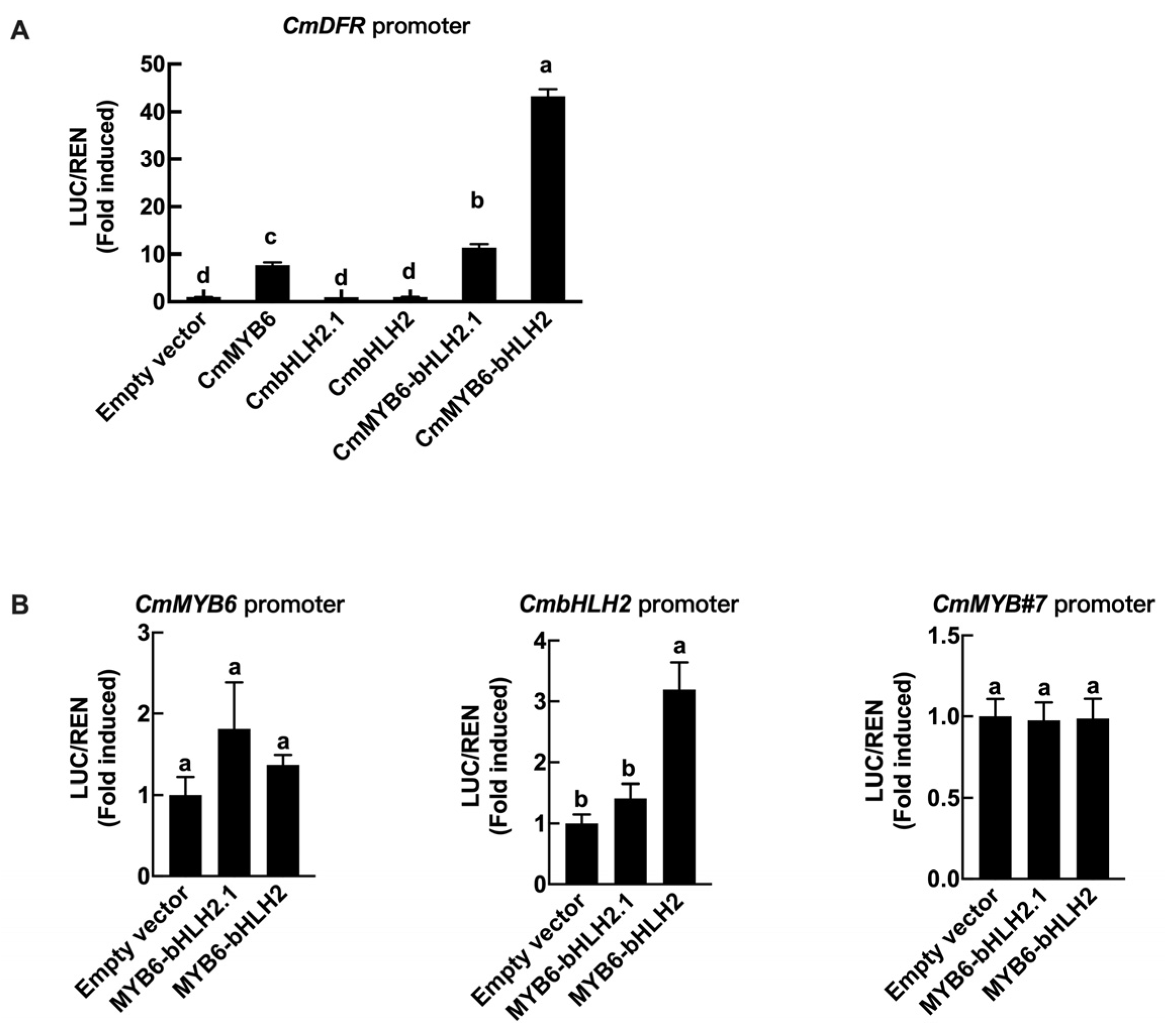
2.10. Regulation Effects of CmMYB6 Together with bHLH2 and bHLH2.1 on the CmDFR Promoter
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. RNA Extraction and cDNA Synthesis
4.3. Anthocyanin Content Analysis
4.4. Transcriptome Library Construction, Sequencing and Data Analysis
4.5. Screening of Differentially Expressed Genes and KEGG Enrichment Analysis
4.6. Phylogenetic Analysis and Protein Sequence Alignment
4.7. Yeast Hybrid Assays
4.8. Transient Over-Expressed Assay
4.9. RT-qPCR
4.10. Dual-Luciferase Assay
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ohmiya, A.; Sumitomo, K.; Aida, R. “Yellow jimba”: Suppression of carotenoid cleavage dioxygenase (CmCCD4a) expression turns white chrysanthemum petals yellow. J. Japan Soc. Hort. Sci. 2009, 78, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.L.; Liu, X.F.; Li, H.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. CmMYB#7, an R3 MYB transcription factor, acts as a negative regulator of anthocyanin biosynthesis in chrysanthemum. J. Exp. Bot. 2019, 70, 3111–3123. [Google Scholar] [PubMed]
- Zhang, Y.Q.H.; Wang, Y.G.; Fang, W.M.; Guan, Z.; Zhang, F.; Chen, F.D. Changes of colors and pigment compositions during the senescence process of chrysanthemum morifolium. Acta Hortic. Sin. 2018, 45, 519–529. [Google Scholar]
- Chen, S.M.; Li, C.H.; Zhu, X.R.; Deng, Y.M.; Sun, W.; Wang, L.S.; Chen, F.D.; Zhang, Z. The identification of flavonoids and the expression of genes of anthocyanin biosynthesis in the chrysanthemum flowers. Biol. Plant. 2012, 56, 458–464. [Google Scholar] [CrossRef]
- Xu, W.J.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.Z.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Liu, X.F.; Li, F.; Xiang, L.L.; Chen, K.S. The isolation and identification of anthocyanin-related GSTs in chrysanthemum. Horticulturae 2021, 7, 231. [Google Scholar] [CrossRef]
- Albert, N.W.; Davies, K.M.; Lewis, D.H.; Zhang, H.B.; Montefiori, M.; Brendolise, C.; Boase, M.; Ngo, H.; Jameson, P.; Schwinn, K. A conserved network of transcriptional activators and repressors regulates anthocyanin pigmentation in eudicots. Plant Cell 2014, 26, 962–980. [Google Scholar] [CrossRef] [Green Version]
- Quattrocchio, F.; Verweij, W.; Kroon, A.; Spelt, C.; Mol, J. PH4 of petunia is an R2R3 MYB protein that activates vacuolar acidification through interactions with basic-helix-loop-helix transcription factors of the anthocyanin pathway. Plant Cell 2006, 18, 1274–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.F.; Yin, X.R.; Allan, A.; Lin-Wang, K.; Shi, Y.N.; Huang, Y.J.; Ferguson, I.; Xu, C.J.; Chen, K.S. The role of MrbHLH1 and MrMYB1 in regulating anthocyanin biosynthetic genes in tobacco and Chinese bayberry (Myrica rubra) during anthocyanin biosynthesis. Plant Cell Tiss. Org. 2013, 115, 285–298. [Google Scholar] [CrossRef]
- Feller, A.; Hernandez, J.M.; Grotewold, E. An ACT-like domain participates in the dimerization of several plant basic-helix-loop-helix transcription factors. J. Biol. Chem. 2006, 281, 28964–28974. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Pattanaik, S.; Feller, A.; Werkman, J.R.; Chai, C.; Wang, Y.Q.; Grotewold, E.; Yuan, L. Regulatory switch enforced by basic helix-loop-helix and ACT-domain mediated dimerizations of the maize transcription factor R. Proc. Natl. Acad. Sci. USA 2012, 109, 11918–11919. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Q.; Shan, X.T.; Gao, R.F.; Yang, S.; Wang, S.C.; Gao, X.; Wang, L. Two IIIf Clade-bHLHs from freesia hybrida play divergent roles in flavonoid biosynthesis and trichome formation when ectopically expressed in arabidopsis. Sci. Rep. 2016, 6, 30514. [Google Scholar] [CrossRef] [Green Version]
- Colanero, S.; Tagliani, A.; Perata, P.; Gonzali, S. Alternative splicing in the anthocyanin fruit gene encoding an R2R3 MYB transcription factor affects anthocyanin biosynthesis in tomato fruits. Plant Commun. 2019, 1, 100006. [Google Scholar] [CrossRef]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 2008, 55, 954–967. [Google Scholar] [CrossRef]
- Zhu, H.F.; Fitzsimmons, K.; Khandelwal, A.; Kranz, R.G. CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Mol. Plant 2009, 2, 790–802. [Google Scholar] [CrossRef]
- Cai, H.Y.; Zhang, M.; Chai, M.N.; He, Q.; Huang, X.Y.; Zhao, L.H.; Qin, Y. Epigenetic regulation of anthocyanin biosynthesis by an antagonistic interaction between H2A.Z and H3K4me3. New Phytol. 2019, 221, 295–308. [Google Scholar] [CrossRef]
- Gates, D.J.; Olson, B.J.S.C.; Clemente, T.E.; Smith, S.D. A novel R3 MYB transcriptional repressor associated with the loss of floral pigmentation in Iochroma. New Phytol. 2017, 217, 1346–1356. [Google Scholar] [CrossRef] [Green Version]
- Morita, Y.; Hoshino, A. Recent advances in flower color variation and patterning of Japanese morning glory and petunia. Breed. Sci. 2018, 68, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.X.; Zhang, Y.C.; Dou, Y.J.; Li, W.J.; Wang, S.M.; Shi, W.J.; Sun, Y.P.; Zhang, Z.H. Single nucleotide mutation in FvMYB10 may lead to the yellow fruit in Fragaria vesca. Mol. Breed. 2017, 37, 35. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Zhang, B.; Allan, A.C.; Lin-Wang, K.; Zhao, Y.; Wang, K.; Chen, K.S.; Xu, C.J. DNA demethylation is involved in the regulation of temperature-dependent anthocyanin accumulation in peach. Plant J. 2020, 102, 965–976. [Google Scholar] [CrossRef]
- Hong, Y.; Li, M.L.; Dai, S.L. Ectopic expression of multiple chrysanthemum (Chrysanthemum × morifolium) R2R3-MYB transcription factor genes regulates anthocyanin accumulation in tobacco. Genes 2019, 10, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Tang, X.J.; Huang, H.; Zhang, Y.; Dai, S.L. Transcriptomic analyses reveal species-specific light-induced anthocyanin biosynthesis in chrysanthemum. BMC Genom. 2015, 16, 202. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Xiang, L.L.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. The identification of a MYB transcription factor controlling anthocyanin biosynthesis regulation in Chrysanthemum flowers. Sci. Hortic. 2015, 194, 278–285. [Google Scholar] [CrossRef]
- Xiang, L.L.; Liu, X.F.; Li, X.; Yin, X.R.; Grierson, D.; Li, F.; Chen, K.S. A novel bHLH transcription factor involved in regulating anthocyanin biosynthesis in Chrysanthemums (Chrysanthemum morifolium ramat.). PLoS ONE 2015, 10, e0143892. [Google Scholar] [CrossRef]
- Liu, H.T.; Wang, Q.; Liu, Y.W.; Zhao, X.Y.; Imaizumi, T.; Somers, D.E.; Tobin, E.M.; Lin, C. Arabidopsis CRY2 and ZTL mediate blue-light regulation of the transcription factor CIB1 by distinct mechanisms. Proc. Natl. Acad. Sci. USA 2013, 110, 17582–17587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depuydt, T.; Vandepoele, K. Multi-omics network-based functional annotation of unknown Arabidopsis genes. Plant J. 2021, 108, 1193–1212. [Google Scholar] [CrossRef] [PubMed]
- Chopra, S.; Hoshino, A.; Boddu, J.; Iida, S. Flavonoid pigments as tools in molecular genetics. In The Science of Flavonoids; Springer: New York, NY, USA, 2006. [Google Scholar] [CrossRef]
- Iida, S.; Morita, Y.; Choi, J.D.; Park, K.I.; Hoshino, A. Genetics and epigenetics in flower pigmentation associated with transposable element in morning glories. Adv. Biophys. 2004, 38, 141–159. [Google Scholar] [CrossRef]
- Nozaki, K.; Takamura, T.; Fukai, S. Effects of high temperature on flower colour and anthocyanin content in pink flower genotypes of greenhouse chrysanthemum (Chrysanthemum morifolium Ramat.). J. Hortic. Sci. Biotech. 2006, 81, 728–734. [Google Scholar] [CrossRef]
- Baudry, A.; Caboche, M.; Lepiniec, L. TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant J. 2006, 46, 768–779. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.H.; Kim, D.H.; Jung, J.A.; Lee, J.Y. Alternative splicing of the basic helix-loop-helix transcription factor gene CmbHLH2 Affects anthocyanin biosynthesis in ray florets of chrysanthemum (Chrysanthemum morifolium). Front. Plant Sci. 2021, 12, 669315. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Neff, M.M.; Chory, J. Genetic interactions between phytochrome A, phytochrome B and cryptochrome 1 during Arabidopsis development. Plant Physiol. 1998, 118, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, K.B.; Nicholas, H.B.J.; Deerfield, D.W.I. GeneDoc: Analysis and visualization of genetic variation. Embnew. News 1997, 4, 14. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, L.; Liu, X.; Shi, Y.; Li, Y.; Li, W.; Li, F.; Chen, K. Comparative Transcriptome Analysis Revealed Two Alternative Splicing bHLHs Account for Flower Color Alteration in Chrysanthemum. Int. J. Mol. Sci. 2021, 22, 12769. https://doi.org/10.3390/ijms222312769
Xiang L, Liu X, Shi Y, Li Y, Li W, Li F, Chen K. Comparative Transcriptome Analysis Revealed Two Alternative Splicing bHLHs Account for Flower Color Alteration in Chrysanthemum. International Journal of Molecular Sciences. 2021; 22(23):12769. https://doi.org/10.3390/ijms222312769
Chicago/Turabian StyleXiang, Lili, Xiaofen Liu, Yanna Shi, Yajing Li, Weidong Li, Fang Li, and Kunsong Chen. 2021. "Comparative Transcriptome Analysis Revealed Two Alternative Splicing bHLHs Account for Flower Color Alteration in Chrysanthemum" International Journal of Molecular Sciences 22, no. 23: 12769. https://doi.org/10.3390/ijms222312769