
Effects of Cynaroside on Cell Proliferation, Apoptosis, Migration and Invasion though the MET/AKT/mTOR Axis in Gastric Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Drug Treatment
2.3. Cell Viability Assay
2.4. BrdU Staining
2.5. Plate Clone Formation Experiment
2.6. Flow Cytometry
2.7. Western Blot Analysis
2.8. Transfection and Infection
2.9. The qRT-PCR Assay
2.10. Ubiquitination Assay
2.11. Soft Agar Assay
2.12. Tumor Xenografts
2.13. Statistical Analysis
3. Results and Discussion
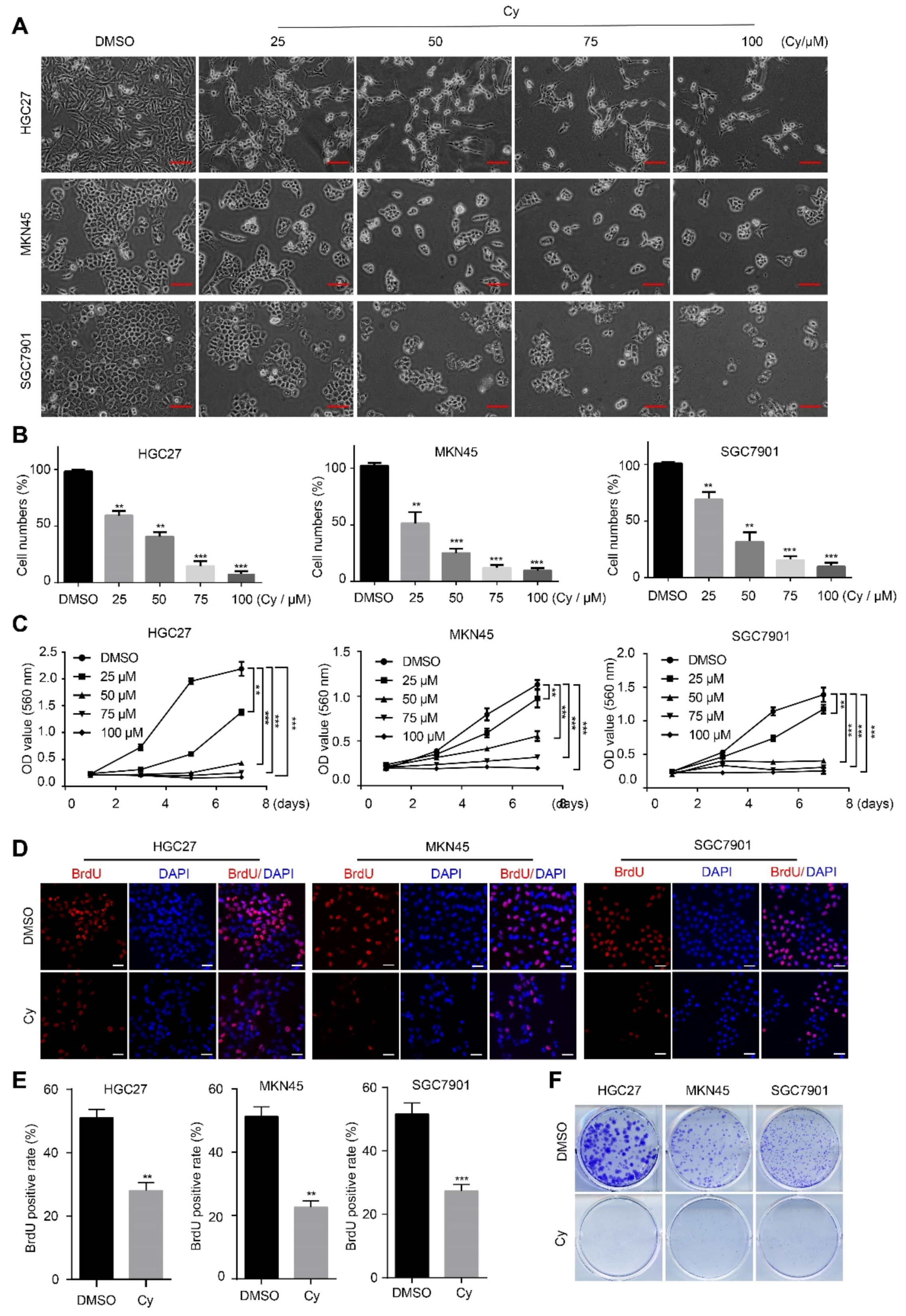
3.1. Cynaroside Inhibits the Proliferation of Gastric Cancer Cells
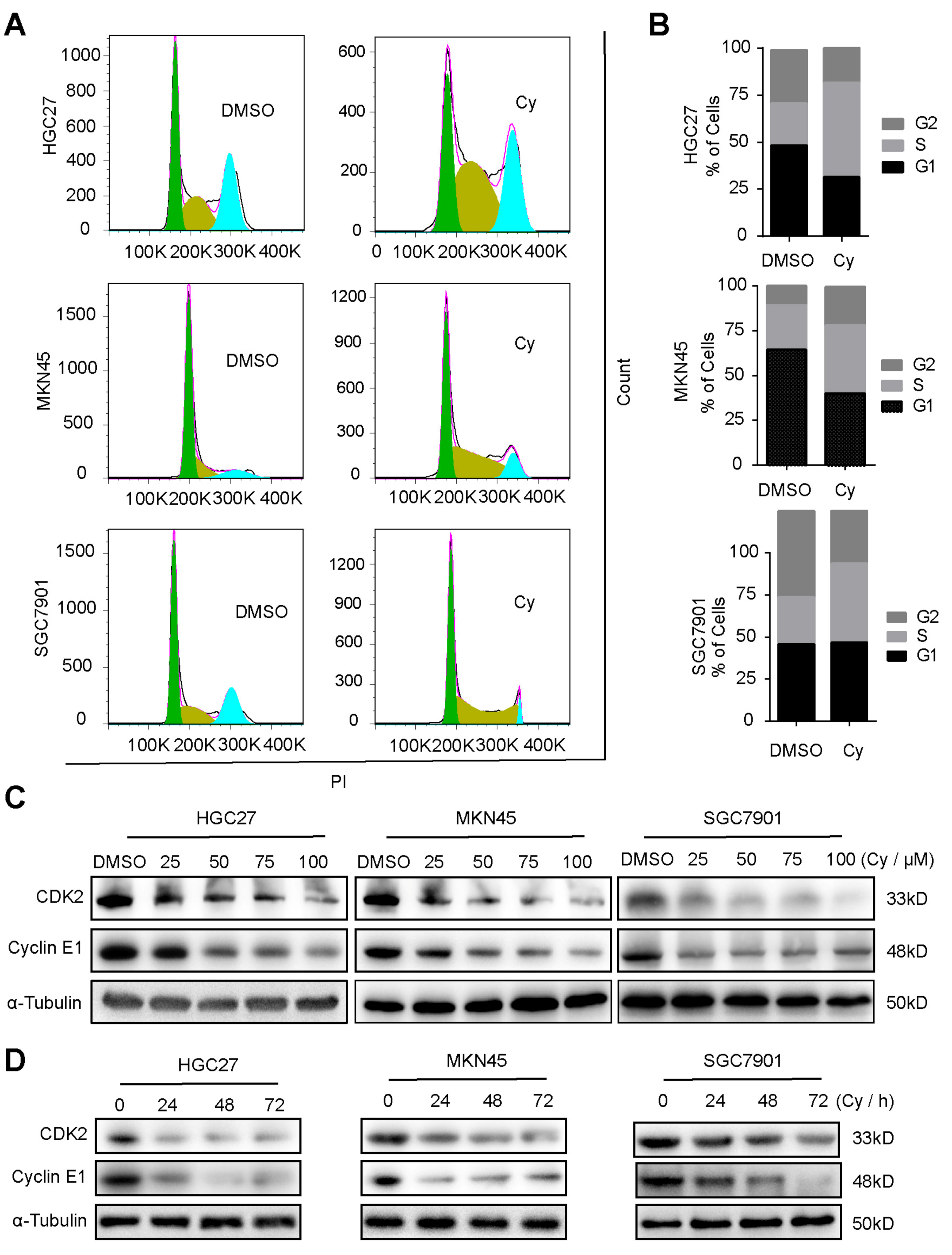
3.2. Cynaroside Causes Cell Cycle Arrest at S Phase
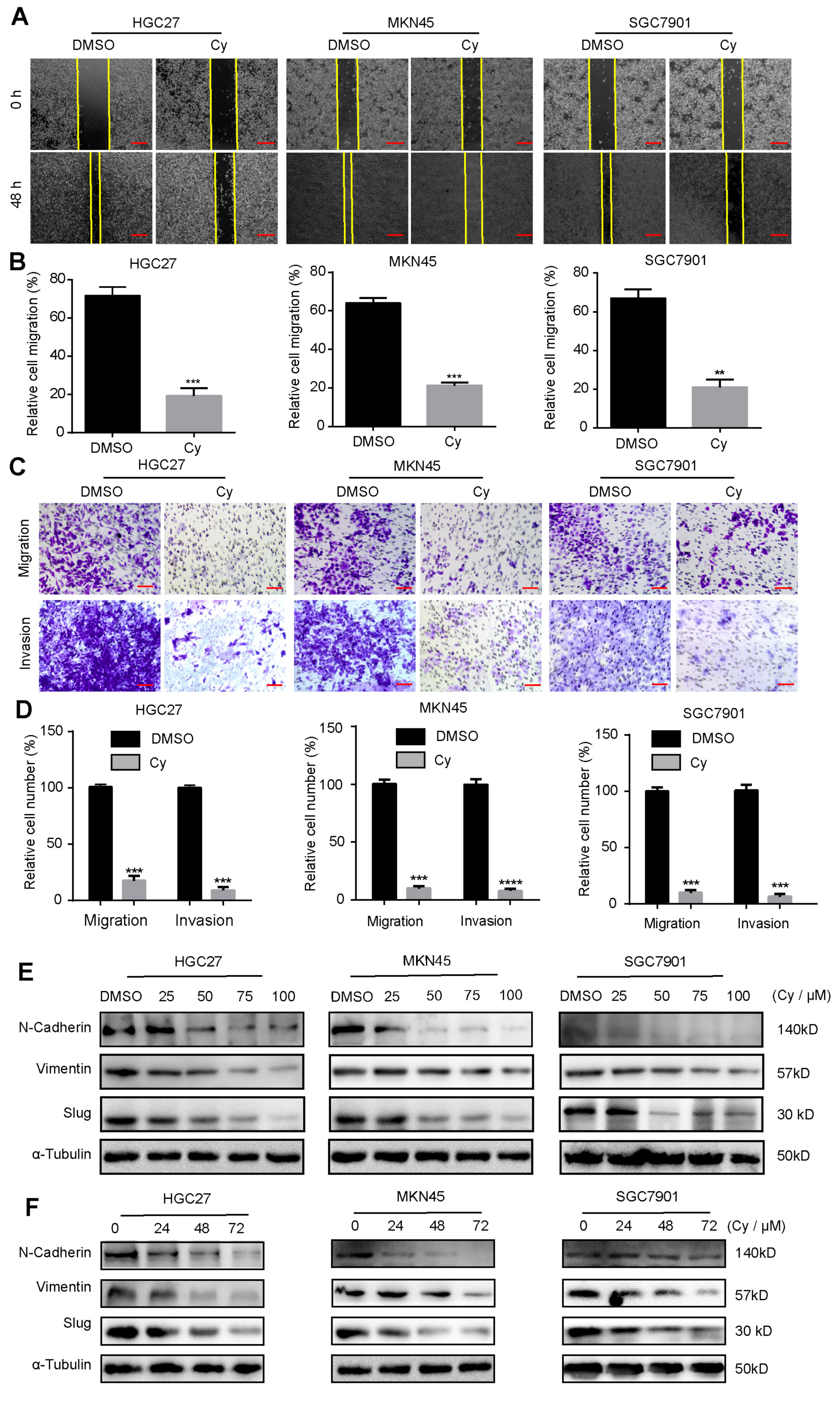
3.3. Cynaroside Inhibits the Migration and Invasion of Gastric Cancer Cells
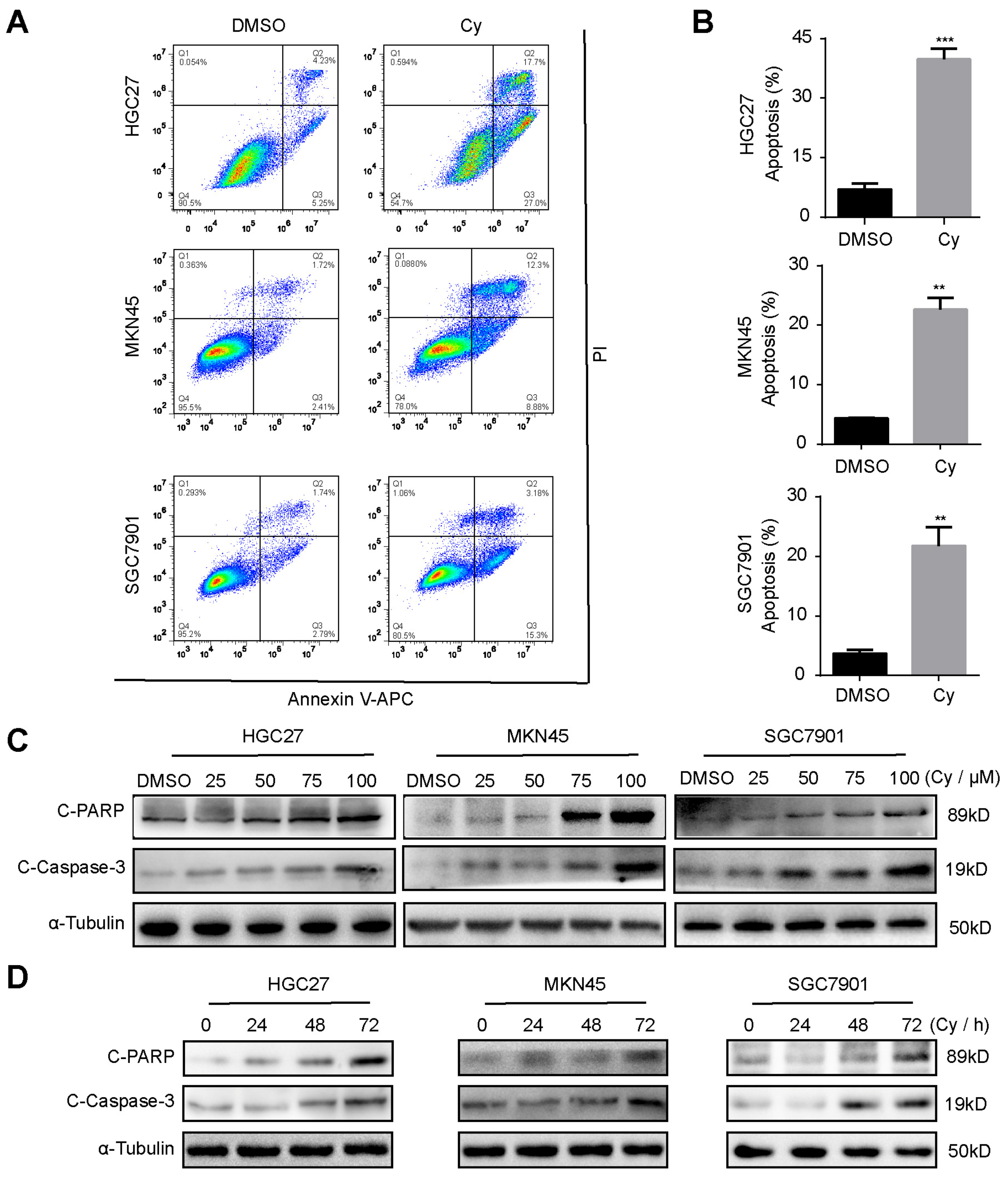
3.4. Cynaroside Induces Apoptosis of Gastric Cancer Cells
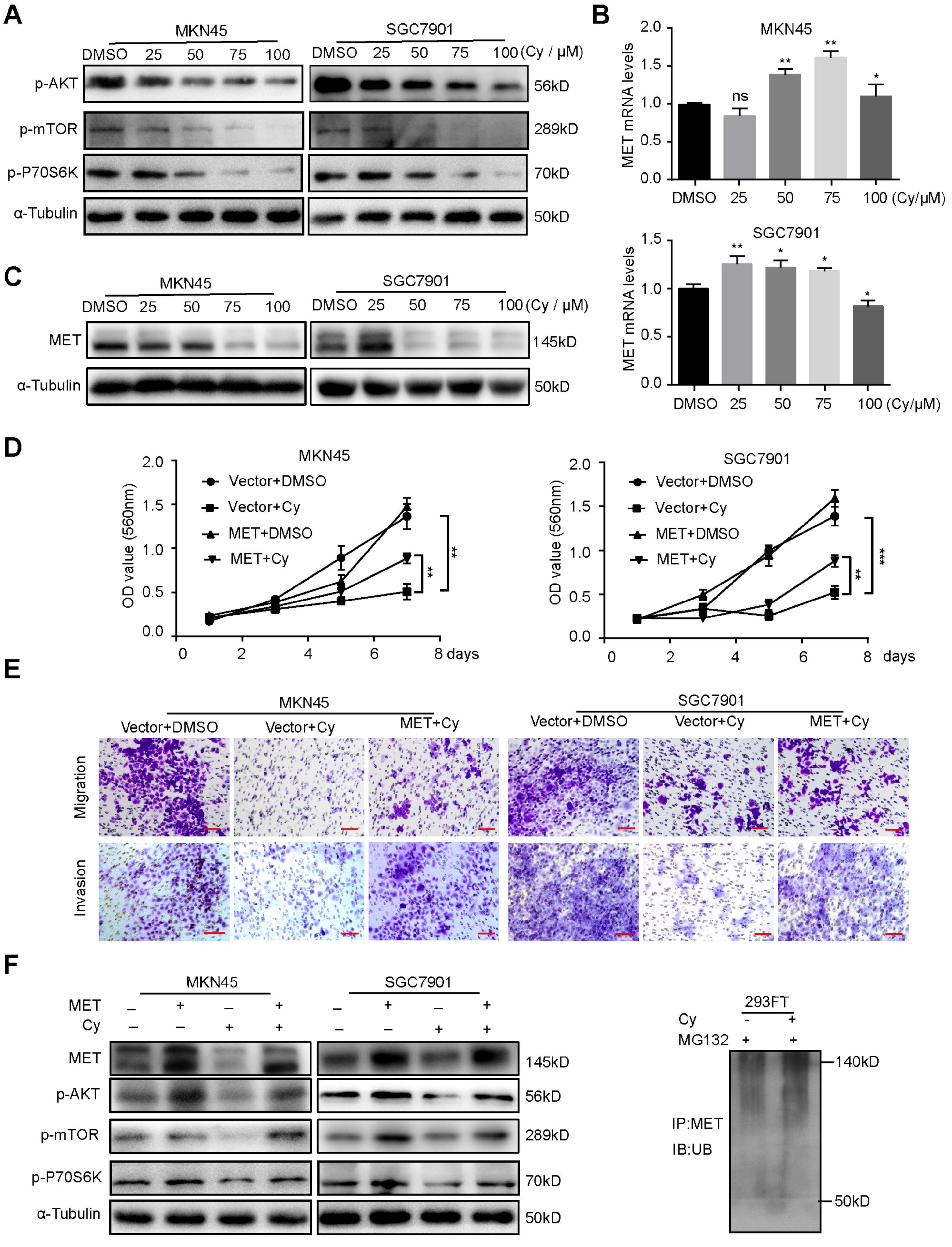
3.5. Cynaroside Inhibits the Activation of the AKT/mTOR Pathway by Enhancing MET Ubiquitination Degradation
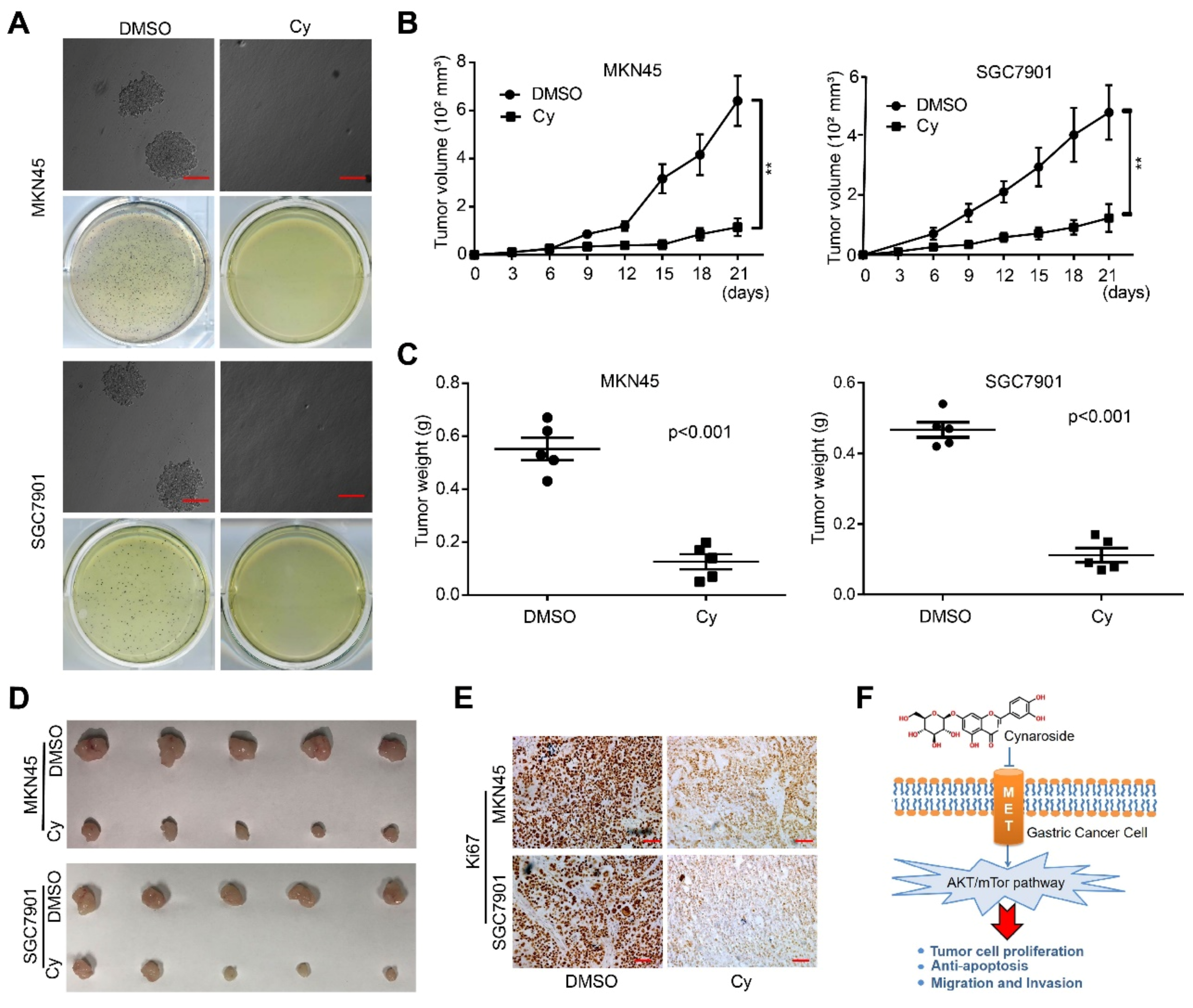
3.6. Cynaroside Inhibits the Tumorigenesis Ability of Gastric Cancer Cells In Vivo and In Vitro
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | serine/threonine kinase 1 |
| C-PARP | cleaved poly (ADP-ribose) polymerase 1 |
| CCNE | cyclin E |
| CDKs | cyclin-dependent kinases |
| Cy | cynaroside |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | dimethyl sulfoxide |
| FBS | fetal bovine serum |
| IgG | immunoglobulin G |
| IHC | immunohistochemistry |
| IUPAC | International Union of Pure and Applied Chemistry |
| MET | mesenchymal–epithelial transition protein |
| mTOR | mammalian target of rapamycin |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| NOD/SCID | nonobese diabetic/severe combined immunodeficiency |
| PI | propidium iodide |
| PVDF | polyvinylidene difluoride |
| RTK | receptor tyrosine kinase |
| SPF | specific pathogen-free |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Eccles, S.A.; Box, C.; Court, W. Cell migration/invasion assays and their application in cancer drug discovery. Biotechnol. Annu. Rev. 2005, 11, 391–421. [Google Scholar]
- Maconi, G.; Manes, G.; Porro, G.B. Role of symptoms in diagnosis and outcome of gastric cancer. World J. Gastroenterol. 2008, 14, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.Y.; Wu, Y.Y.; Yang, J.B.; Yang, D.Q.; Fang, X.D. Progress in the treatment of advanced gastric cancer. Tumor Biol. 2017, 39, 1010428317714626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Yuan, T.; Yin, N.; Ma, X.; Zhang, Z.; Zhu, Z.; Shaukat, A.; Deng, G. Luteoloside Protects the Uterus from Staphylococcus aureus-Induced Inflammation, Apoptosis, and Injury. Inflammation 2018, 41, 1702–1716. [Google Scholar] [CrossRef]
- Stefano, A.D.; Caporali, S.; Daniele, N.D.; Rovella, V.; Cardillo, C.; Schinzari, F.; Minieri, M.; Pieri, M.; Candi, E.; Bernardini, S.; et al. Anti-Inflammatory and Proliferative Properties of Luteolin-7-O-Glucoside. Int. J. Mol. Sci. 2021, 22, 1321. [Google Scholar] [CrossRef]
- Palombo, R.; Caporali, S.; Falconi, M.; Iacovelli, F.; Rocca, B.M.D.; Surdo, A.L.; Campione, E.; Candi, E.; Melino, G.; Bernardini, S.; et al. Luteolin-7-O-β-d-Glucoside Inhibits Cellular Energy Production Interacting with HEK2 in Keratinocytes. Int. J. Mol. Sci. 2019, 20, 2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witkowska-Banaszczak, E.; Krajka-Kuźniak, V.; Papierska, K. The effect of luteolin 7-glucoside, apigenin 7-glucoside and Succisa pratensis extracts on NF-κB activation and α-amylase activity in HepG2 cells. Acta Biochim. Pol. 2020, 67, 41–47. [Google Scholar] [CrossRef]
- Nho, J.H.; Jung, H.K.; Lee, M.J.; Jang, J.H.; Sim, M.O.; Jeong, D.E.; Cho, H.W.; Kim, J.C. Beneficial Effects of Cynaroside on Cisplatin-Induced Kidney Injury In Vitro and In Vivo. Toxicol. Res. 2018, 34, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Shen, S.; Zhao, X.; Gong, X. Luteoloside induces G0/G1 arrest and pro-death autophagy through the ROS-mediated AKT/mTOR/p70S6K signalling pathway in human non-small cell lung cancer cell lines. Biochem. Biophys. Res. Commun. 2017, 494, 263–269. [Google Scholar] [CrossRef]
- Fan, S.H.; Wang, Y.Y.; Lu, J.; Zheng, Y.L.; Wu, D.M.; Li, M.Q.; Hu, B.; Zhang, Z.F.; Cheng, W.; Shan, Q. Luteoloside suppresses proliferation and metastasis of hepatocellular carcinoma cells by inhibition of NLRP3 inflammasome. PLoS ONE 2014, 9, e89961. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Wang, C.; Li, L.; Liang, H.; Dai, J.; Ling, X.; Tang, H. Luteoloside Inhibits Proliferation and Promotes Intrinsic and Extrinsic Pathway-Mediated Apoptosis Involving MAPK and mTOR Signaling Pathways in Human Cervical Cancer Cells. Int. J. Mol. Sci. 2018, 19, 1664. [Google Scholar] [CrossRef] [Green Version]
- Chabot, T.; Cheraud, Y.; Fleury, F. Relationships between DNA repair and RTK-mediated signaling pathways. Biochim. Biophys. Acta. Rev. Cancer 2021, 1875, 188495. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Lovly, C.M. Mechanisms of receptor tyrosine kinase activation in cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef]
- Yamaoka, T.; Kusumoto, S.; Ando, K.; Ohba, M.; Ohmori, T. Receptor Tyrosine Kinase-Targeted Cancer Therapy. Int. J. Mol. Sci. 2018, 19, 3491. [Google Scholar] [CrossRef] [Green Version]
- Tang, R.Q.; Ma, F.F.; Li, W.; Ouyang, S.R.; Liu, Z.; Wu, J.X. miR-206-3p Inhibits 3T3-L1 Cell Adipogenesis via the c-Met/PI3K/Akt Pathway. Int. J. Mol. Sci. 2017, 18, 1510. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.K.; Zhang, L.X.; Jia, C.Y.; Sun, H.M.; Zou, Q.G.; Wang, Z.; Wang, L.; Qu, M.J.; Zhao, X.; Zhao, H.; et al. MACC1 overexpression induces cisplatin resistance in lung adenocarcinoma A549 cells by activating c-Met/Akt pathway. Int. J. Clin. Exp. Med. 2017, 10, 11778–11786. [Google Scholar]
- Zhao, E.; Hou, J.; Ke, X.; Abbas, M.N.; Kausar, S.; Zhang, L.; Cui, H. The Roles of Sirtuin Family Proteins in Cancer Progression. Cancers 2019, 11, 1949. [Google Scholar] [CrossRef] [Green Version]
- Tong, G.L.; Cheng, B.R.; Li, J.Z.; Wu, X.; Nong, Q.H.; He, L.R.; Li, X.; Li, L.Q.; Wang, S.B. MACC1 regulates PDL1 expression and tumor immunity through the c-Met/AKT/mTOR pathway in gastric cancer cells. Cancer Med. USA 2019, 8, 7044–7054. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Deng, H.Y.; Zhao, Y.J.; Li, C.; Liang, Y. LncRNA XIST/miR-34a axis modulates the cell proliferation and tumor growth of thyroid cancer through MET-PI3K-AKT signaling. J. Exp. Clin. Cancer Res. 2018, 37, 1–12. [Google Scholar] [CrossRef]
- Jia, Y.F.; Feng, Q.; Tang, B.; Luo, X.D.; Yang, Q.; Yang, H.; Li, Q. Decorin Suppresses Invasion and EMT Phenotype of Glioma by Inducing Autophagy via c-Met/Akt/mTOR Axis. Front. Oncol. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Cai, H.P.; Yu, Y.J.; Ni, X.R.; Li, C.; Hu, Y.J.; Wang, J.; Chen, F.R.; Xi, S.Y.; Chen, Z.P. LncRNA LINC00998 inhibits the malignant glioma phenotype via the CBX3-mediated c-Met/Akt/mTOR axis. Cell Death Dis. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Zhao, E.; Feng, L.; Bai, L.; Cui, H. NUCKS promotes cell proliferation and suppresses autophagy through the mTOR-Beclin1 pathway in gastric cancer. J. Exp. Clin. Cancer Res. 2020, 39, 194. [Google Scholar] [CrossRef]
- Bai, L.; Ma, Y.; Wang, X.; Feng, Q.; Zhang, Z.; Wang, S.; Zhang, H.; Lu, X.; Xu, Y.; Zhao, E.; et al. Polydatin Inhibits Cell Viability, Migration, and Invasion Through Suppressing the c-Myc Expression in Human Cervical Cancer. Front. Cell Dev. Biol. 2021, 9, 587218. [Google Scholar] [CrossRef]
- Zhao, E.; Ding, J.; Xia, Y.; Liu, M.; Ye, B.; Choi, J.H.; Yan, C.; Dong, Z.; Huang, S.; Zha, Y.; et al. KDM4C and ATF4 Cooperate in Transcriptional Control of Amino Acid Metabolism. Cell Rep. 2016, 14, 506–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Li, J.; Hu, X.; Feng, J.; Wang, H.; Xiong, F. Protective effects of cynaroside on oxidative stress in retinal pigment epithelial cells. J. Biochem. Mol. Toxicol. 2019, 33, e22352. [Google Scholar] [CrossRef]
- Lee, S.A.; Park, B.R.; Moon, S.M.; Shin, S.H.; Kim, J.S.; Kim, D.K.; Kim, C.S. Cynaroside protects human periodontal ligament cells from lipopolysaccharide-induced damage and inflammation through suppression of NF-kappaB activation. Arch. Oral Biol. 2020, 120, 104944. [Google Scholar] [CrossRef]
- Feng, J.H.; Dong, X.W.; Yu, H.L.; Shen, W.; Lv, X.Y.; Wang, R.; Cheng, X.X.; Xiong, F.; Hu, X.L.; Wang, H. Cynaroside protects the blue light-induced retinal degeneration through alleviating apoptosis and inducing autophagy in vitro and in vivo. Phytomedicine 2021, 88, 153604. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sun, G.B.; Wang, M.; Xiao, J.; Sun, X.B. Protective effects of cynaroside against H2O2-induced apoptosis in H9c2 cardiomyoblasts. J. Cell. Biochem. 2011, 112, 2019–2029. [Google Scholar] [CrossRef]
- Levinson, A.D.; Oppermann, H.; Varmus, H.E.; Bishop, J.M. The purified product of the transforming gene of avian sarcoma virus phosphorylates tyrosine. J. Biol. Chem. 1980, 255, 11973–11980. [Google Scholar] [CrossRef]
- Yao, H.P.; Hudson, R.; Wang, M.H. Progress and challenge in development of biotherapeutics targeting MET receptor for treatment of advanced cancer. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188425. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wroblewski, L.E.; Peek, R.M., Jr. Helicobacter pylori, Cancer, and the Gastric Microbiota. Adv. Exp. Med. Biol. 2016, 908, 393–408. [Google Scholar] [PubMed]
- Lyons, K.; Le, L.C.; Pham, Y.T.; Borron, C.; Park, J.Y.; Tran, C.T.D.; Tran, T.V.; Tran, H.T.; Vu, K.T.; Do, C.D.; et al. Gastric cancer: Epidemiology, biology, and prevention: A mini review. Eur. J. Cancer Prev. 2019, 28, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Gantuya, B.; Tuan, V.P.; Yamaoka, Y. Diffuse Gastric Cancer: A Summary of Analogous Contributing Factors for Its Molecular Pathogenicity. Int. J. Mol. Sci. 2018, 19, 2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, J.; Wang, Z.; Sun, W.; Li, Z.; Cai, H.; Zhao, E.; Cui, H. Effects of Cynaroside on Cell Proliferation, Apoptosis, Migration and Invasion though the MET/AKT/mTOR Axis in Gastric Cancer. Int. J. Mol. Sci. 2021, 22, 12125. https://doi.org/10.3390/ijms222212125
Ji J, Wang Z, Sun W, Li Z, Cai H, Zhao E, Cui H. Effects of Cynaroside on Cell Proliferation, Apoptosis, Migration and Invasion though the MET/AKT/mTOR Axis in Gastric Cancer. International Journal of Molecular Sciences. 2021; 22(22):12125. https://doi.org/10.3390/ijms222212125
Chicago/Turabian StyleJi, Juanli, Zhongze Wang, Wei Sun, Zekun Li, Huarui Cai, Erhu Zhao, and Hongjuan Cui. 2021. "Effects of Cynaroside on Cell Proliferation, Apoptosis, Migration and Invasion though the MET/AKT/mTOR Axis in Gastric Cancer" International Journal of Molecular Sciences 22, no. 22: 12125. https://doi.org/10.3390/ijms222212125