
Cathepsin B Regulates Mice Granulosa Cells’ Apoptosis and Proliferation In Vitro

 ,
,
Abstract
:1. Introduction
2. Results
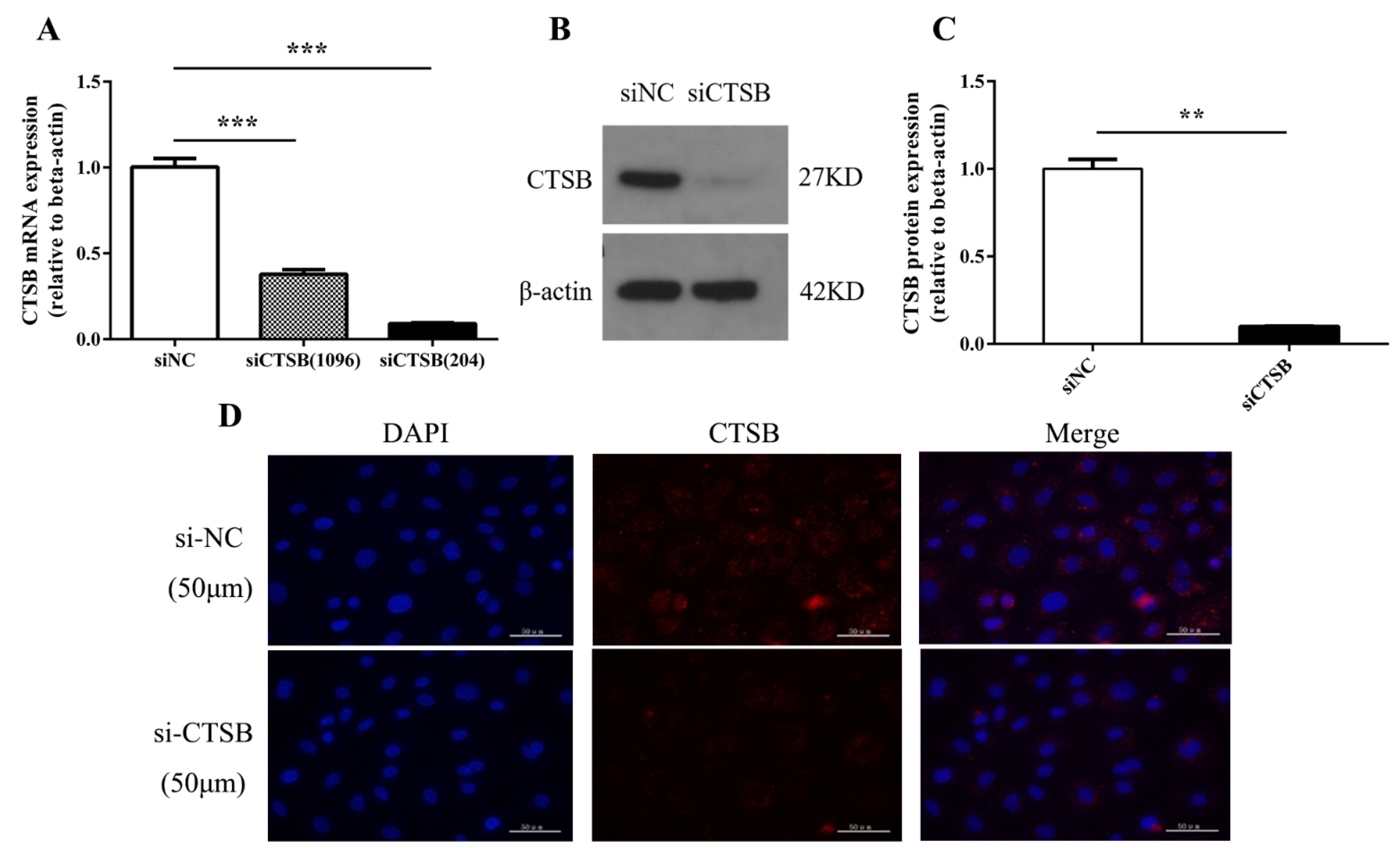
2.1. siRNA Successfully Represses CTSB Expression
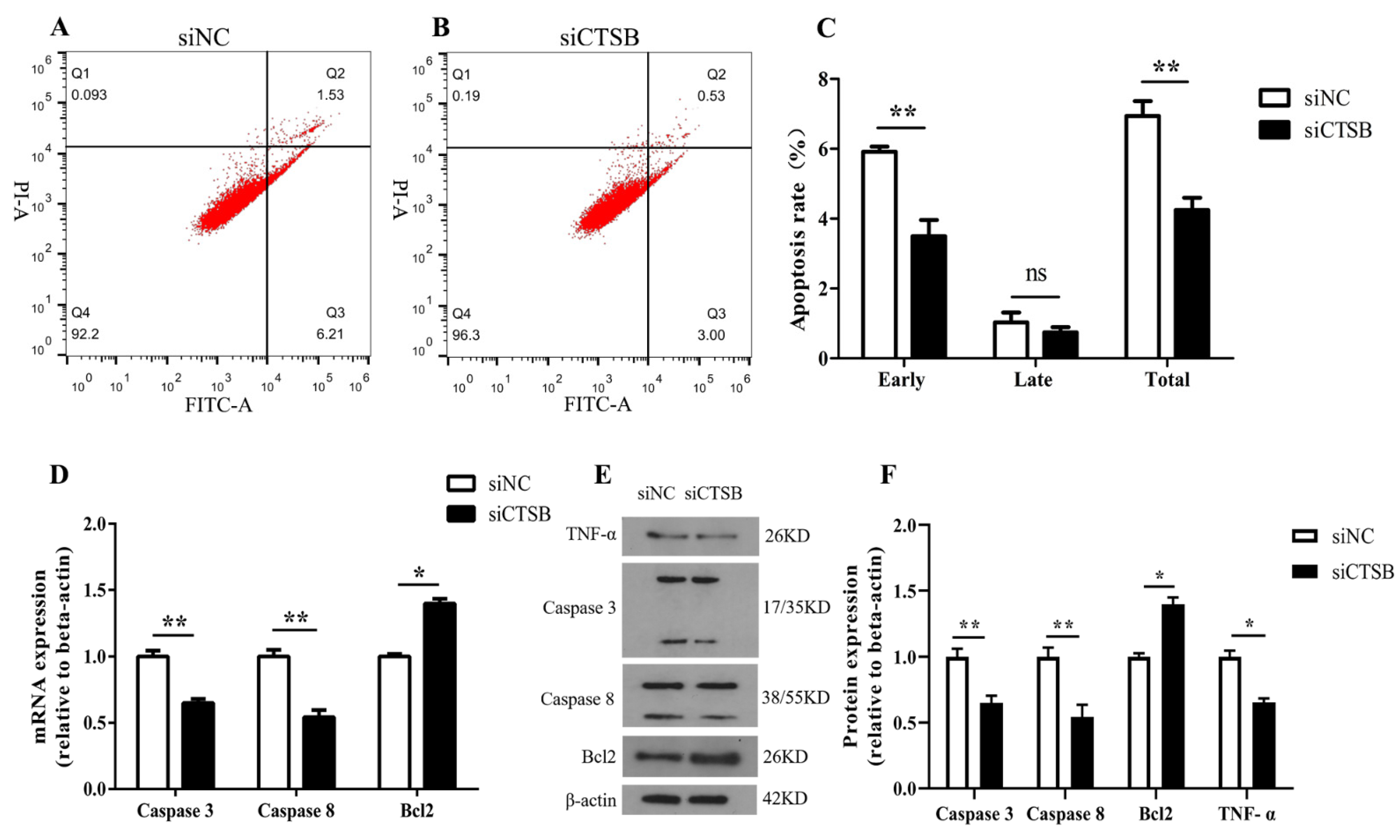
2.2. CTSB Depletion Suppresses Apoptosis in Murine GCs In Vitro
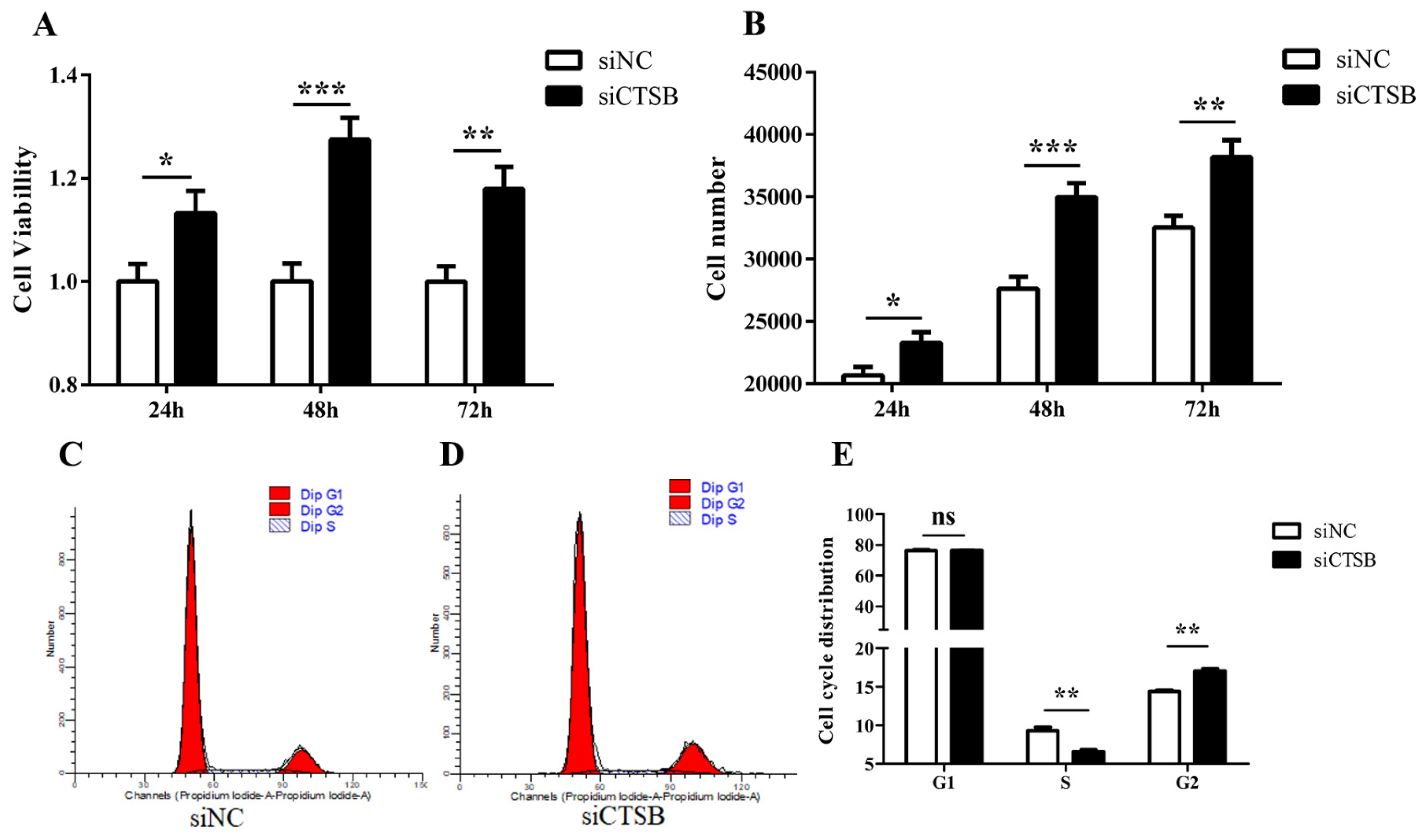
2.3. CTSB Downregulation Promotes Cell Proliferation and Affects Cell Cycle Progression in Mice GCs
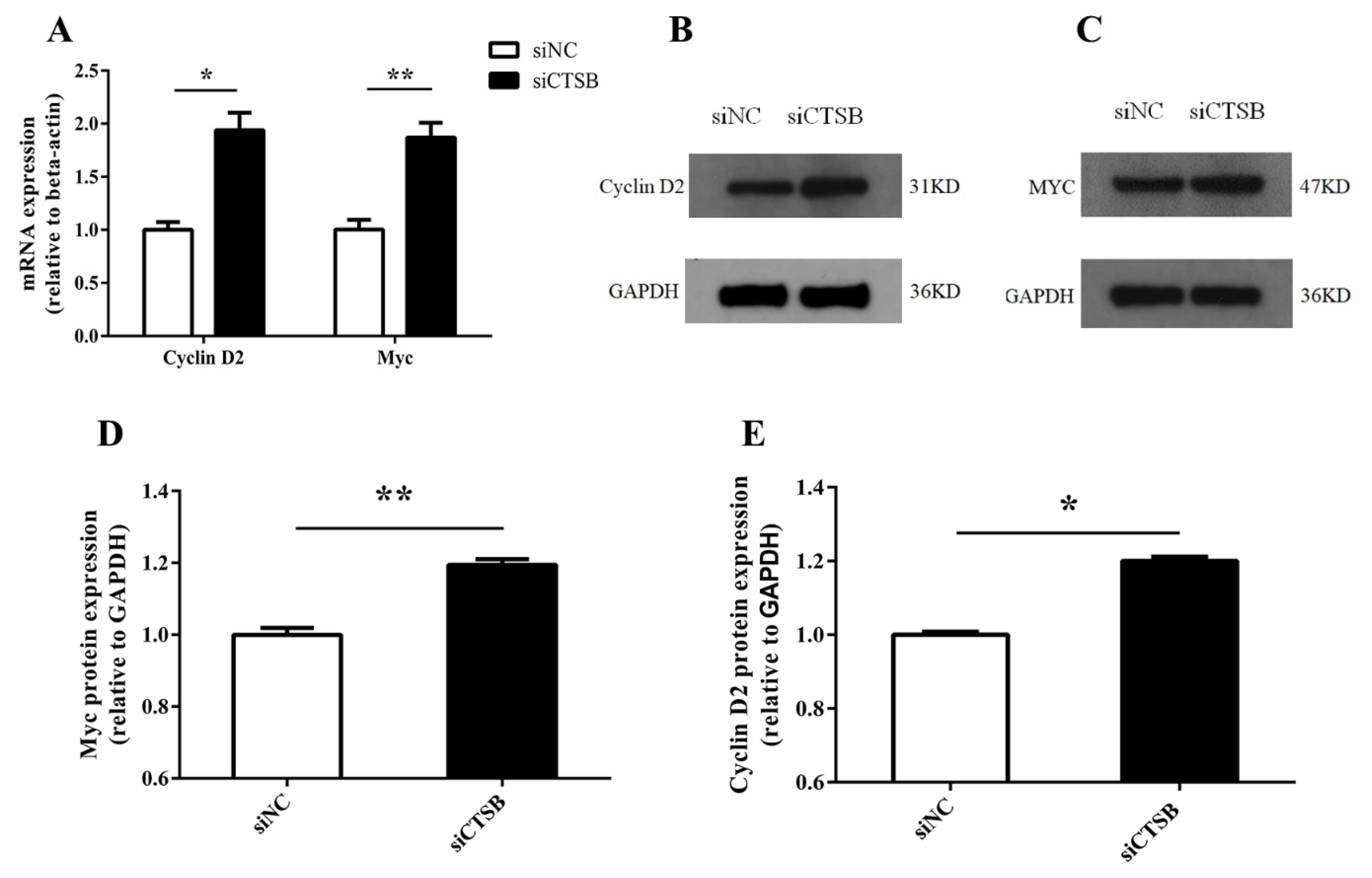
2.4. Downregulation of CTSB Mediates Mouse GC Proliferation by Activation of the ERK and Akt Phosphorylation Pathways
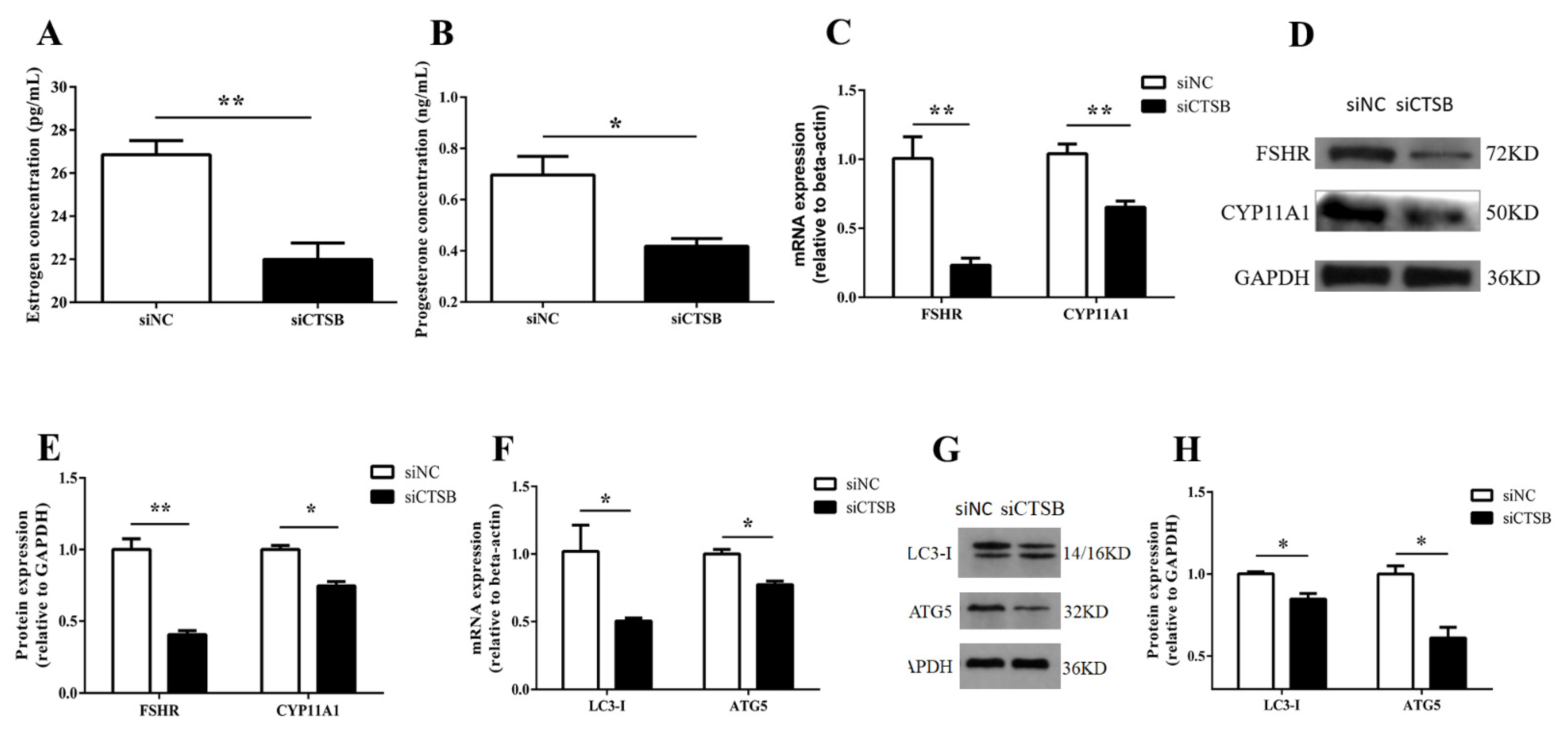
2.5. Steroidogenesis and Autophagy-Related Gene Expression Were Altered by CTSB Depletion in Mouse GCs In Vitro
3. Discussion
4. Materials and Methods
4.1. Management of Experimental Animals
4.2. Mouse GCs Isolation and Culture
4.3. Cathepsin B (CTSB) siRNA Transfection
4.4. RNA Extraction and Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
4.5. Quantitative Real-Time PCR (qRT-PCR)
4.6. Protein Extraction and Western Blotting
4.7. Cell Apoptosis Detection
4.8. Cell Counts and Proliferation Assay
4.9. Cellular Immunofluorescence
4.10. Cell Cycle Assay Middling
4.11. Hormones Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heatwole, H. The Heat Dissipation Limit Theory and Evolution of Life Histories in Endotherms—Time to Dispose of the Disposable Soma Theory?—Supplementary Data. Integr. Comp. Biol. 2012, 50, 793–807. [Google Scholar]
- Worku, T.; Wang, K.; Ayers, D.; Wu, D.; Ur Rehman, Z.; Zhou, H.; Yang, L. Regulatory roles of ephrinA5 and its novel signaling pathway in mouse primary granulosa cell apoptosis and proliferation. Cell Cycle 2018, 17, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Hongshusui, Y.S.; Weili, Z.B.; Jinghetan, M.J. Apoptosis in Granulosa cells during follicular atresia: Relationship with steroids and insulin-like growth factors. Cell Res. 2004, 14, 341–346. [Google Scholar]
- Hughes, F.M.; Gorospe, W.C. Biochemical identification of apoptosis (programmed cell death) in granulosa cells: Evidence for a potential mechanism underlying follicular atresia. Endocrinology 1991, 129, 2415–2422. [Google Scholar] [CrossRef]
- Byskov, A.G. Follicular atresia. In The Vertebrate Ovary: Comparative Biology and Evolution; Jones, R.E., Ed.; Plenum Press: New York, NY, USA, 1978; pp. 533–562. [Google Scholar]
- Choi, J.Y.; Jo, M.W.; Lee, E.Y.; Yoon, B.-K.; Choi, D.S. The role of autophagy in follicular development and atresia in rat granulosa cells. Fertil. Steril. 2010, 93, 2532–2537. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef]
- Levine, B.; Klionsky, D.J. Development by self-digestion: Molecular mechanisms and biological functions of autophagy. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- Hwang, S.O.; Lee, G.M. Nutrient deprivation induces autophagy as well as apoptosis in Chinese hamster ovary cell culture. Biotechnol. Bioeng. 2008, 99, 678–685. [Google Scholar] [CrossRef]
- Kessel, D.; Reiners, J.J., Jr. Initiation of apoptosis and autophagy by the Bcl-2 antagonist HA14-1. Cancer Lett. 2007, 249, 294–299. [Google Scholar] [CrossRef] [Green Version]
- Duerrschmidt, N.; Zabirnyk, O.; Nowicki, M.; Ricken, A.; Hmeidan, F.A.; Blumenauer, V.; Borlak, J.; Spanel-Borowski, K. Lectin-like oxidized low-density lipoprotein receptor-1-mediated autophagy in human granulosa cells as an alternative of programmed cell death. Endocrinology 2006, 147, 3851–3860. [Google Scholar] [CrossRef] [Green Version]
- Billig, H.; Furuta, I.; Hsueh, A. Estrogens inhibit and androgens enhance ovarian granulosa cell apoptosis. Endocrinology 1993, 133, 2204–2212. [Google Scholar] [CrossRef]
- AJ, B. Kirschke H: Cathepsin B, cathepsin H and cathepsin L. Methods Enzym. 1981, 80 Pt C, 535–561. [Google Scholar]
- Mohamed, M.M.; Sloane, B.F. Multifunctional enzymes in cancer. Nat. Rev. Cancer 2006, 6, 764–775. [Google Scholar] [CrossRef]
- Buhling, F.; Fengler, A.; Brandt, W.; Welte, T.; Ansorge, S.; Nagler, D.K. Part II: Cellular Endopeptidases: New Cathepsins; Results from Knock-out-mice; Regulatory Aspects-26. Review: Novel Cysteine Proteases of the Papain Family. Adv. Exp. Med. Biol. 2000, 477, 241–254. [Google Scholar]
- Turk, B.; Turk, D.; Turk, V. Lysosomal cysteine proteases: More than scavengers. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2000, 1477, 98–111. [Google Scholar] [CrossRef]
- Klose, A.; Zigrino, P.; Dennhöfer, R.; Mauch, C.; Hunzelmann, N. Identification and discrimination of extracellularly active cathepsins B and L in high-invasive melanoma cells. Anal. Biochem. 2006, 353, 57–62. [Google Scholar] [CrossRef]
- Vancompernolle, K.; Van Herreweghe, F.; Pynaert, G.; Van De Craen, M.; De Vos, K.; Totty, N.; Sterling, A.; Fiers, W.; Vandenabeele, P.; Grooten, J. Atractyloside-induced release of cathepsin B, a protease with caspase-processing activity. FEBS Lett. 1998, 438, 150–158. [Google Scholar] [CrossRef]
- Bossy-Wetzel, E.; Newmeyer, D.D.; Green, D.R. Mitochondrial cytochrome c release in apoptosis occurs upstream of DEVD-specific caspase activation and independently of mitochondrial transmembrane depolarization. EMBO J. 1998, 17, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Zhao, M.-H.; Liang, S.; Cui, X.-S.; Kim, N.-H. Inhibition of cathepsin B activity reduces apoptosis by preventing cytochrome c release from mitochondria in porcine parthenotes. J. Reprod. Dev. 2015, 61, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Bettegowda, A.; Patel, O.V.; Lee, K.B.; Park, K.E.; Salem, M.; Yao, J.; Ireland, J.J.; Smith, G.W. Identification of novel bovine cumulus cell molecular markers predictive of oocyte competence: Functional and diagnostic implications. Biol. Reprod. 2008, 79, 301–309. [Google Scholar] [CrossRef]
- Balboula, A.; Yamanaka, K.; Sakatani, M.; Hegab, A.; Zaabel, S.; Takahashi, M. Intracellular cathepsin B activity is inversely correlated with the quality and developmental competence of bovine preimplantation embryos. Mol. Reprod. Dev. 2011, 77, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Balboula, A.; Yamanaka, K.; Sakatani, M.; Hegab, A.; Zaabel, S.; Takahashi, M. Cathepsin B Activity is Related to the Quality of Bovine Cumulus Oocyte Complexes and Its Inhibition Can Improve Their Developmental Competence. Mol. Reprod. Dev. 2010, 77, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Balboula, A.Z.; Yamanaka, K.; Sakatani, M.; Kawahara, M.; Hegab, A.O.; Zaabel, S.M.; Takahashi, M. Cathepsin B activity has a crucial role in the developmental competence of bovine cumulus-oocyte complexes exposed to heat shock during in vitro maturation. Reproduction 2013, 146, 407–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.; Zheng, H.; Rong, X.; Rong, X.; Zhang, J.; Fang, W.; Zhao, P.; Luo, R. Over-expression of cathepsin B in hepatocellular carcinomas predicts poor prognosis of HCC patients. Mol. Cancer 2016, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.T.; Baba, Y.; Shima, K.; Nosho, K.; Chung, D.C.; Hung, K.E.; Mahmood, U.; Madden, K.; Poss, K.; Ranieri, A.; et al. Cathepsin B Expression and Survival in Colon Cancer: Implications for Molecular Detection of Neoplasia. Cancer Epidemiol. Biomark. Prev. 2010, 19, 2777–2785. [Google Scholar] [CrossRef] [Green Version]
- Hughes, S.J.; Glover, T.W.; Zhu, X.X.; Kuick, R.; Thoraval, D.; Orringer, M.B.; Beer, D.G.; Hanash, S. A novel amplicon at 8p22-23 results in overexpression of cathepsin B in esophagel adenocarcinoma. Proc. Natl. Acad. Sci. USA 1998, 95, 12410–12415. [Google Scholar] [CrossRef] [Green Version]
- Niedergethmann, M.; Wostbrock, B.; Sturm, J.W.; Willeke, F.; Post, S.; Hildenbrand, R. Prognostic Impact of Cysteine Proteases Cathepsin B and Cathepsin L in Pancreatic Adenocarcinoma. Pancreas 2004, 29, 204. [Google Scholar] [CrossRef]
- Sendler, M.; Maertin, S.; John, D.; Persike, M.; Weiss, F.U.; Krueger, B.; Wartmann, T.; Wagh, P.; Halangk, W.; Schaschke, N.; et al. Cathepsin B Activity Initiates Apoptosis via Digestive Protease Activation in Pancreatic Acinar Cells and Experimental Pancreatitis. J. Biol. Chem. 2016, 291, 14717–14731. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Yang, B.; Yu, W.; Xiao, Y.; Yu, D.; Zhang, Q. Cathepsin B links oxidative stress to the activation of NLRP3 inflammasome. Exp. Cell Res. 2017, 362, 180–187. [Google Scholar] [CrossRef]
- Iwama, H.; Mehanna, S.; Imasaka, M.; Hashidume, S.; Nishiura, H.; Yamamura, K.-I.; Suzuki, C.; Uchiyama, Y.; Hatano, E.; Ohmuraya, M. Cathepsin B and D deficiency in the mouse pancreas induces impaired autophagy and chronic pancreatitis. Sci. Rep. 2021, 11, 6596. [Google Scholar] [CrossRef]
- Tai, W. Chemical modulation of siRNA lipophilicity for efficient delivery. J. Control. Release 2019, 307, 98–107. [Google Scholar] [CrossRef]
- Sargent, K.M.; Lu, N.; Clopton, D.T.; Pohlmeier, W.E.; Brauer, V.M.; Ferrara, N.; Silversides, D.W.; Cupp, A.S. Loss of Vascular Endothelial Growth Factor A (VEGFA) Isoforms in Granulosa Cells Using pDmrt-1-Cre or Amhr2-Cre Reduces Fertility by Arresting Follicular Development and by Reducing Litter Size in Female Mice. PLoS ONE 2015, 10, e0116332. [Google Scholar] [CrossRef] [Green Version]
- Quirk, S.M.; Cowan, R.G.; Harman, R.M.; Hu, C.-L.; Porter, D.A. Ovarian follicular growth and atresia: The relationship between cell proliferation and survival. J. Anim. Sci. 2004, 82 (Suppl. S13), E40. [Google Scholar] [CrossRef]
- Kaipia, A.; Hsueh, A.J. Regulation of ovarian follicle atresia. Annu. Rev. Physiol. 1997, 59, 349–363. [Google Scholar] [CrossRef]
- Chwieralski, C.; Welte, T.; Bühling, F. Cathepsin-regulated apoptosis. Apoptosis 2006, 11, 143–149. [Google Scholar] [CrossRef]
- Boya, P.; Andreau, K.; Poncet, D.; Zamzami, N.; Perfettini, J.-L.; Metivier, D.; Ojcius, D.; Jaattela, M.; Kroemer, G. Lysosomal membrane permeabilization induces cell death in a mitochondrion-dependent fashion. J. Exp. Med. 2003, 197, 1323–1334. [Google Scholar] [CrossRef]
- Johansson, A.-C.; Appelqvist, H.; Nilsson, C.; Kågedal, K.; Roberg, K.; Öllinger, K. Regulation of apoptosis-associated lysosomal membrane permeabilization. Apoptosis 2010, 15, 527–540. [Google Scholar] [CrossRef] [Green Version]
- Droga-Mazovec, G.; Bojic, L.; Petelin, A.; Ivanova, S.; Repnik, U.; Salvesen, G.S.; Stoka, V.; Turk, V.; Turk, B. Cysteine cathepsins trigger caspase-dependent cell death through cleavage of bid and antiapoptotic Bcl-2 homologues. J. Biol. Chem. 2008, 283, 19140–19150. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D. Noncaspase proteases in apoptosis. Leukemia 2000, 14, 1695–1703. [Google Scholar] [CrossRef] [Green Version]
- Leist, M.; Jäättelä, M. Triggering of apoptosis by cathepsins. Cell Death Differ. 2001, 8, 324–326. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Antunes, F.; Eaton, J.W.; Brunk, U.T. Lysosomal enzymes promote mitochondrial oxidant production, cytochrome c release and apoptosis. Eur. J. Biochem. 2003, 270, 3778–3786. [Google Scholar] [CrossRef] [Green Version]
- Guicciardi, M.E.; Deussing, J.; Miyoshi, H.; Bronk, S.F.; Svingen, P.A.; Peters, C.; Kaufmann, S.H.; Gores, G.J. Cathepsin B contributes to TNF-α–mediated hepatocyte apoptosis by promoting mitochondrial release of cytochrome c. J. Clin. Investig. 2000, 106, 1127–1137. [Google Scholar] [CrossRef] [Green Version]
- Shan, X.; Yu, T.; Yan, X.; Wu, J.; Fan, Y.; Guan, X.; Fang, F.; Lin, Y.; Zhang, Y.; Li, Y.; et al. Proteomic analysis of healthy and atretic porcine follicular granulosa cells. J. Proteom. 2020, 232, 104027. [Google Scholar] [CrossRef]
- Talpur, H.S.; Worku, T.; Rehman, Z.U.; Dad, R.; Bhattarai, D.; Bano, I.; Liang, A.; He, C.; Yang, L. Knockdown of melatonin receptor 1 and induction of follicle-stimulating hormone on the regulation of mouse granulosa cell function. Reprod. Biol. 2017, 17, 380–388. [Google Scholar] [CrossRef]
- Li, J.-X.; Ding, X.-M.; Han, S.; Wang, K.; Jiao, C.-Y.; Li, X.-C. mir-637 inhibits the proliferation of cholangiocarcinoma cell QBC939 through interfering CTSB expression. Eur. Rev. Med Pharmacol. Sci. 2018, 22, 1265–1276. [Google Scholar]
- Zhang, L.; Hou, Y.; Ashktorab, H.; Gao, L.; Xu, Y.; Wu, K.; Zhai, J.; Zhang, L. The impact of C-MYC gene expression on gastric cancer cell. Mol. Cell. Biochem. 2010, 344, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, C.; Thieke, K.; Maier, A.; Saffrich, R.; Hanley-Hyde, J.; Ansorge, W.; Reed, S.; Sicinski, P.; Bartek, J.; Eilers, M. Direct induction of cyclin D2 by Myc contributes to cell cycle progression and sequestration of p27. EMBO J. 1999, 18, 5321–5333. [Google Scholar] [CrossRef]
- Bouchard, C.; Dittrich, O.; Kiermaier, A.; Dohmann, K.; Menkel, A.; Eilers, M.; Lüscher, B. Regulation of cyclin D2 gene expression by the Myc/Max/Mad network: Myc-dependent TRRAP recruitment and histone acetylation at the cyclin D2 promoter. Genes Dev. 2001, 15, 2042–2047. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, X.; Jin, H.; Guo, X.; Xia, L.; Chen, Z.; Bai, M.; Liu, J.; Shang, X.; Wu, K.; et al. MiR-206 inhibits gastric cancer proliferation in part by repressing CyclinD2. Cancer Lett. 2013, 332, 94–101. [Google Scholar] [CrossRef]
- Chen, S.; Shimoda, M.; Chen, J.; Matsumoto, S.; Grayburn, P.A. Transient overexpression of cyclin D2/CDK4/GLP1 genes induces proliferation and differentiation of adult pancreatic progenitors and mediates islet regeneration. Cell Cycle 2012, 11, 695–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, E.; Northwood, I.C.; Gonzalez, F.A.; Latour, D.A.; Seth, A.; Abate, C.; Curran, T.; Davis, R.J. Pro-Leu-Ser/Thr-Pro is a consensus primary sequence for substrate protein phosphorylation. Characterization of the phosphorylation of c-myc and c-jun proteins by an epidermal growth factor receptor threonine 669 protein kinase. J. Biol. Chem. 1991, 266, 15277–15285. [Google Scholar] [CrossRef]
- Sears, R.; Nuckolls, F.; Haura, E.; Taya, Y.; Tamai, K.; Nevins, J.R. Multiple Ras-dependent phosphorylation pathways regulate Myc protein stability. Genes Dev. 2000, 14, 2501–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefloch, R.; Pouyssegur, J.; Lenormand, P. Single and combined silencing of ERK1 and ERK2 reveals their positive contribution to growth signaling depending on their expression levels. Mol. Cell. Biol. 2008, 28, 511–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.; Liao, Q.; Su, M.; Huang, K.; Jin, J.; Cao, D. AKT and ERK dual inhibitors: The way forward? Cancer Lett. 2019, 459, 30–40. [Google Scholar] [CrossRef]
- Lu, Z.; Xu, S. ERK1/2 MAP kinases in cell survival and apoptosis. IUBMB Life 2006, 58, 621–631. [Google Scholar] [CrossRef]
- Casarini, L.; Crépieux, P. Molecular mechanisms of action of FSH. Front. Endocrinol. 2019, 10, 305. [Google Scholar] [CrossRef]
- Hu, M.-C.; Hsu, H.-J.; Guo, I.-C.; Chung, B.-C. Function of Cyp11a1 in animal models. Mol. Cell. Endocrinol. 2004, 215, 95–100. [Google Scholar] [CrossRef]
- Chien, Y.; Cheng, W.-C.; Wu, M.-R.; Jiang, S.-T.; Shen, C.-K.J.; Chung, B.-C. Misregulated progesterone secretion and impaired pregnancy in Cyp11a1 transgenic mice. Biol. Reprod. 2013, 89, 91. [Google Scholar] [CrossRef]
- Macklon, N.S.; Fauser, B. Follicle development during the normal menstrual cycle. Maturitas 1998, 30, 181–188. [Google Scholar] [CrossRef]
- Ye, X.; Li, Y.; Wang, Z.; Andersen, C.L. Functions of lysosomes in mammalian female reproductive system. Reprod. Dev. Med. 2020, 4, 109. [Google Scholar] [CrossRef]
- Yoshii, S.R.; Kuma, A.; Akashi, T.; Hara, T.; Yamamoto, A.; Kurikawa, Y.; Itakura, E.; Tsukamoto, S.; Shitara, H.; Eishi, Y.; et al. Systemic analysis of Atg5-null mice rescued from neonatal lethality by transgenic ATG5 expression in neurons. Dev. Cell 2016, 39, 116–130. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | AC/NO | Primer Sequences | Production Length (bp) | Annealing Temp. (°C) |
|---|---|---|---|---|
| β-actin | NM_007393 | F: GTGACGTTGACATCCGTAAAGA | 287 | 60 |
| R: GTAACAGTCCGCCTAGAAGCAC | ||||
| Caspase 3 | NM_009810 | F: GTCTGACTGGAAAGCCGAAAC | 205 | 59.5 |
| R: GACTGGATGAACCACGACCC | ||||
| Caspase 8 | NM_001080126 | F: TCTCGGAATCGGTAGCAAACC | 173 | 60 |
| R: AGAAGAGCTGTAACCTGTGGC | ||||
| Cyclin D2 | NM_009829.3 | F: TACCTCCCGCAGTGTTCCTA | 158 | 60 |
| R: GCCAAGAAACGGTCCAGGTA | ||||
| MYC | NM_001177352.1 | F: GTTGGAAACCCCGCAGACAG | 264 | 60.5 |
| R: GTAGCGACCGCAACATAGGA | ||||
| CYP11A1 | NM_001346787.1 | F: TACTAACCTAGCCCGCCTCG | 163 | 60.5 |
| R: CTCCTGCGCATAGAGAGAGC | ||||
| FSHR | NM_013523.3 | F: AACACTTGCCAGCCTTTCAC | 184 | 60 |
| R: TGGGTTCCGTTGAATGCACA | ||||
| CTSB | NM_007798.3 | F: CAATGGCCGAGTCAACGTG | 176 | 59 |
| R: TGGTGTATGGTAAGCAGCCT | ||||
| BCL2 | NM_009741.5 | F: GAACTGGGGGAGGATTGTGG | 194 | 60 |
| R: GCATGCTGGGGCCATATAGT | ||||
| LC3-I | NM_025735.3 | F:AGGAGAAGGATGAAGACGGA | 160 | 57 |
| R:CCACTGGGGACTGAAATAGC | ||||
| ATG5 | NM_001358596.1 | F:CAGTGTGATCCCGGCAGA | 199 | 58 |
| R:GAGTAAAGCAAGTTGGAATTCG | ||||
| si-CTSB (204) | F: GCUGUCGGAUGACCUGAUUTT | |||
| R: AAUCAGGUCAUCCGACAGCTT | ||||
| si-CTSB (1096) | F: UCAGAAAUUGUGGCUGGAATT | |||
| R: UUCCAGCCACAAUUUCUGATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Ahmad, M.J.; Ye, T.; Du, C.; Zhang, X.; Liang, A.; Yang, L. Cathepsin B Regulates Mice Granulosa Cells’ Apoptosis and Proliferation In Vitro. Int. J. Mol. Sci. 2021, 22, 11827. https://doi.org/10.3390/ijms222111827
Chen C, Ahmad MJ, Ye T, Du C, Zhang X, Liang A, Yang L. Cathepsin B Regulates Mice Granulosa Cells’ Apoptosis and Proliferation In Vitro. International Journal of Molecular Sciences. 2021; 22(21):11827. https://doi.org/10.3390/ijms222111827
Chicago/Turabian StyleChen, Chao, Muhammad Jamil Ahmad, Tingzhu Ye, Chao Du, Xinxin Zhang, Aixin Liang, and Liguo Yang. 2021. "Cathepsin B Regulates Mice Granulosa Cells’ Apoptosis and Proliferation In Vitro" International Journal of Molecular Sciences 22, no. 21: 11827. https://doi.org/10.3390/ijms222111827