
Melatonin Modulates Plant Tolerance to Heavy Metal Stress: Morphological Responses to Molecular Mechanisms


 ,
,  ,
,  , , ,
, , ,  ,
,  , and
, and
Abstract
:1. Introduction
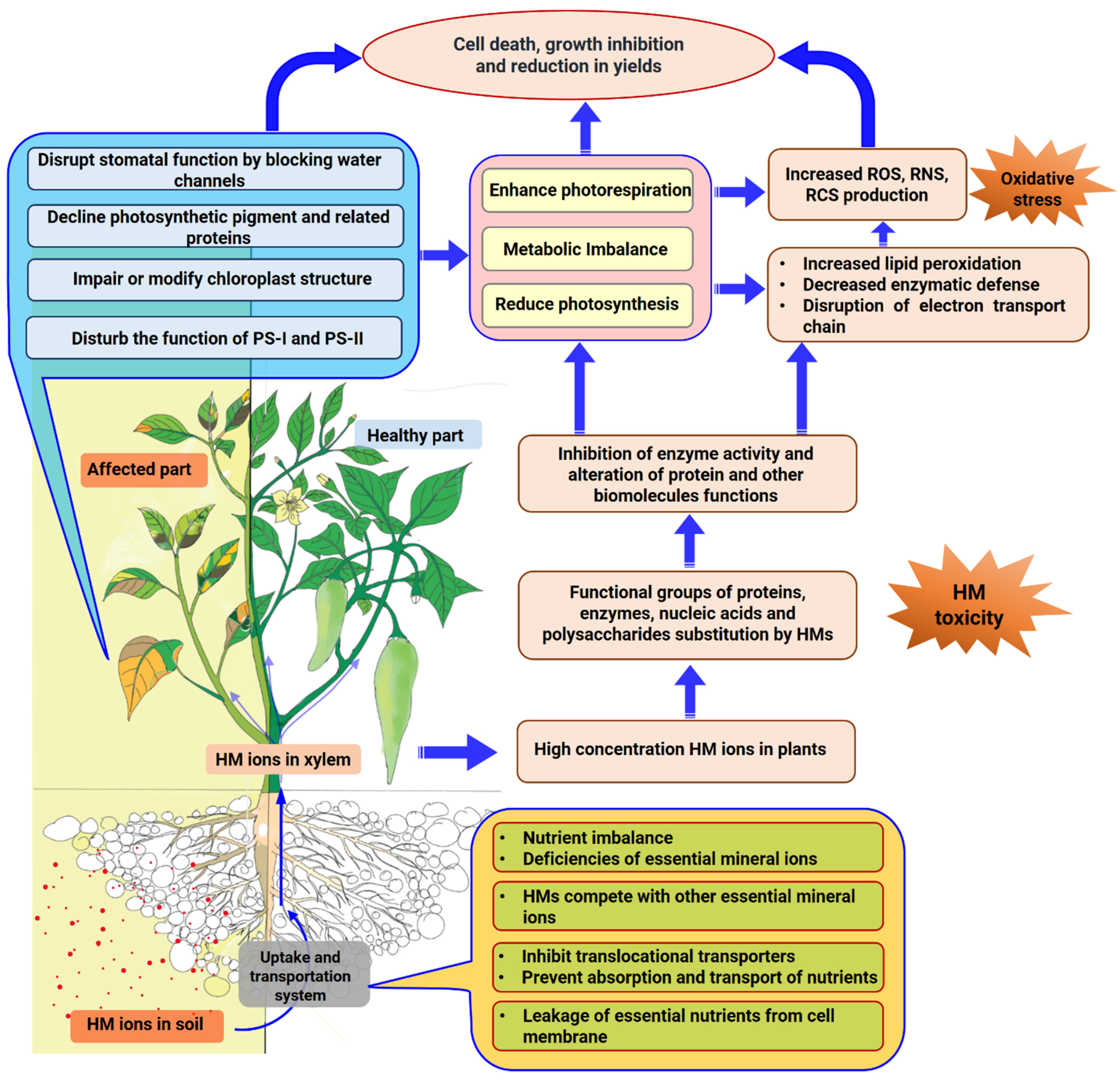
2. Mechanism of Heavy-Metal-Induced Growth Inhibition
3. Heavy Metals Induce Endogenous Melatonin Accumulation in Plants
4. Role of Exogenous Melatonin on Heavy Metal Stress Tolerance
5. Mechanisms of Melatonin-Mediated Heavy Metal Stress Tolerance
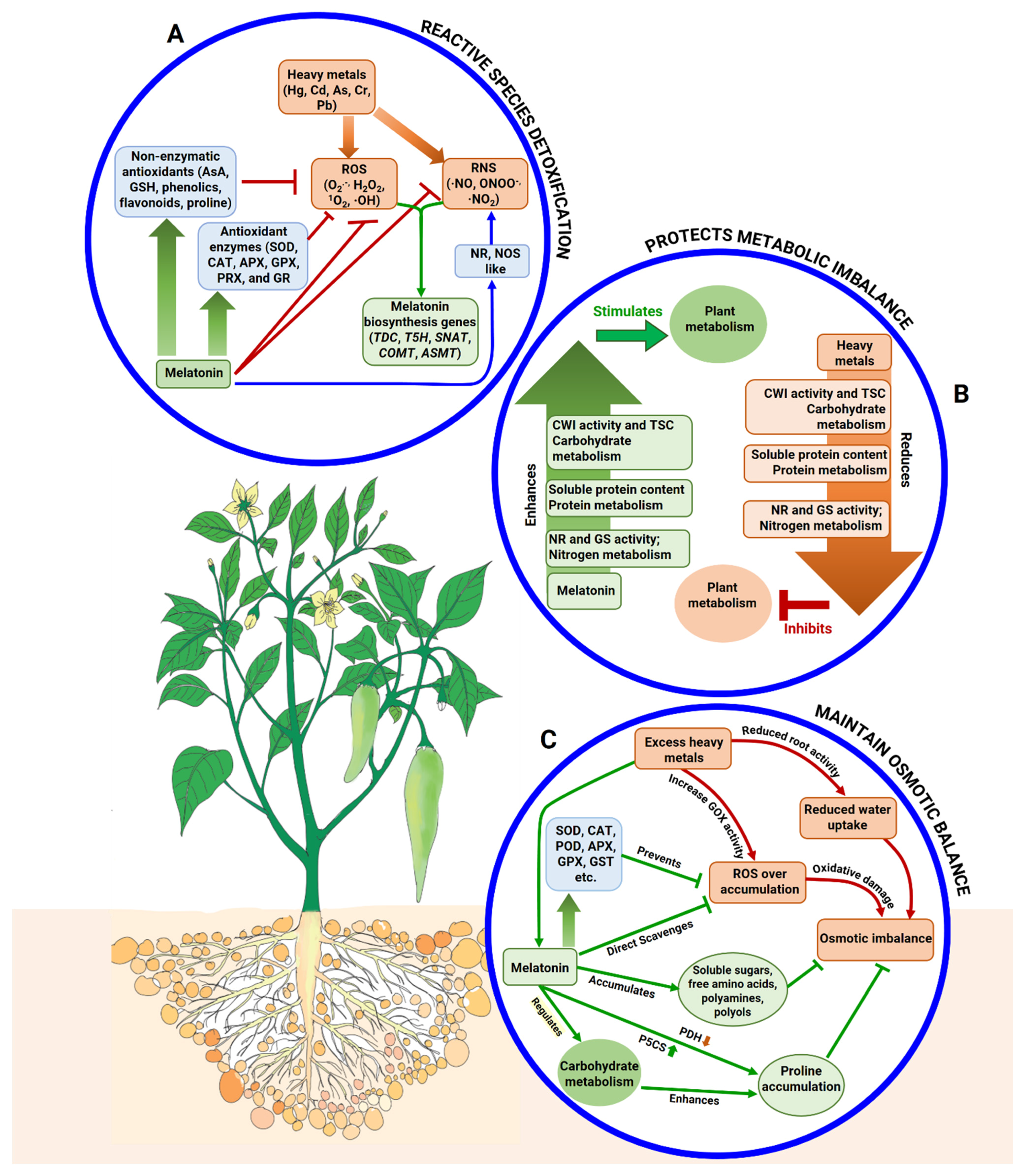
5.1. Melatonin Modulates Reactive Species Detoxification and Antioxidant Upregulation
5.2. Melatonin Protects against Different Metabolic Imbalances
5.3. Melatonin Adjusts Osmotic Imbalance in Plants
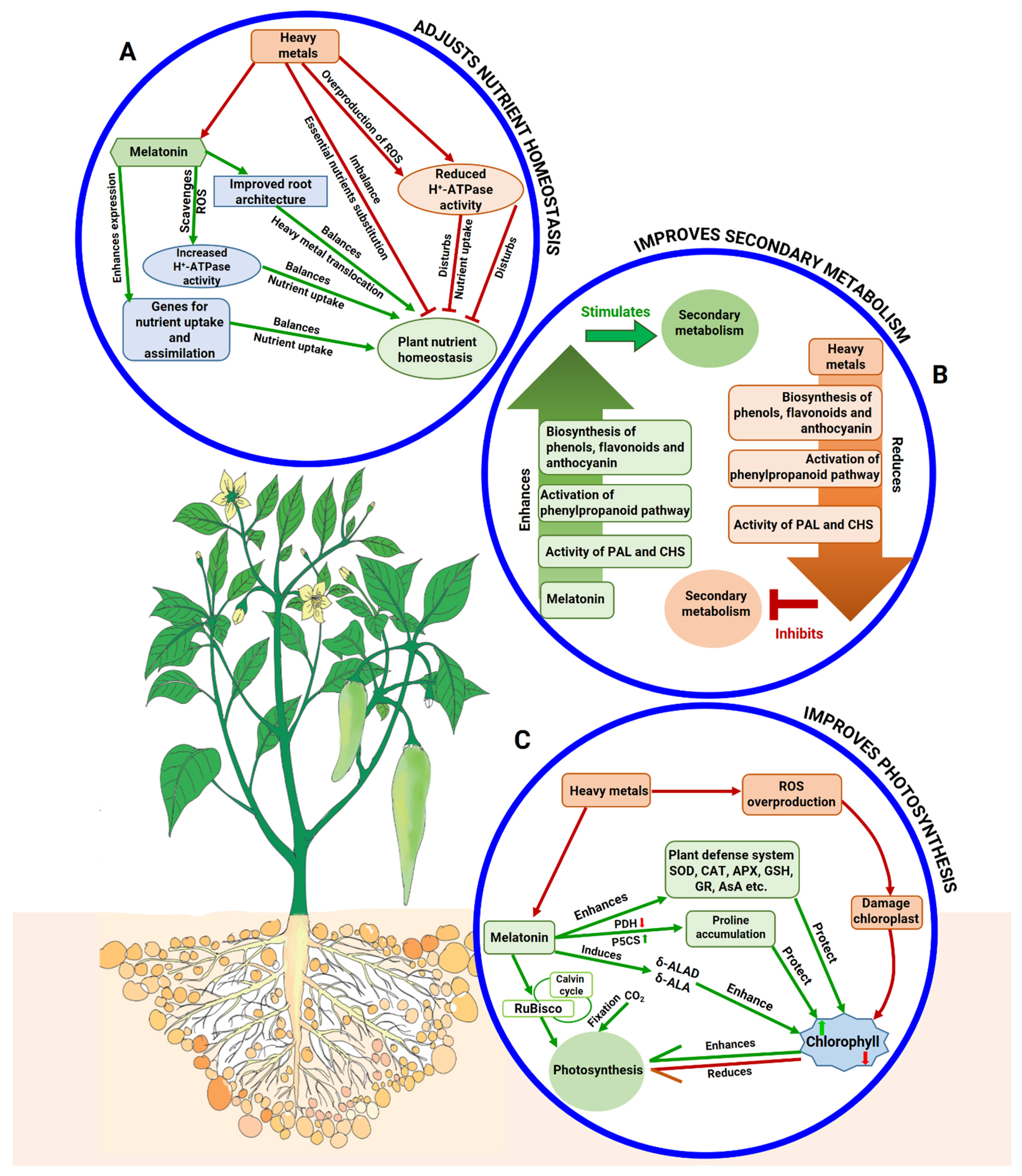
5.4. Melatonin Maintains Homeostasis of Essential Nutrients
5.5. Melatonin Regulates Secondary Metabolites
5.6. Melatonin Protects Photosynthetic Attributes
5.7. Melatonin Upregulates Defensive Genes
6. Endogenous MT in HM Stress Tolerance
7. Conclusions and Future Prospective
- Exogenous MT significantly improves the concentration of photosynthetic pigments by upregulating Chl synthesizing enzymes and downregulating genes responsible for Chl degradation.
- Exogenous MT alleviates the harsh effects of HM stress on plant growth, photosynthesis, and development.
- Exogenous MT mitigates HMs toxicity through upregulating a wide range of defensive genes that are responsible for higher antioxidant activities and metal chelating properties.
- The application of MT enhances HM tolerance in plants by the accumulation of osmolytes, increased antioxidant enzyme activity, and osmotic adjustment, maintaining membrane integrity and limiting the lipid peroxidation and ROS generation, improving the activity of antioxidant enzymes and non-enzymatic antioxidants.
- MT also improves different nutrient homeostasis.
- The exogenous application of MT induces plant secondary metabolites biosynthesis.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gull, A.; Lone, A.A.; Wani, N.U.I. Biotic and Abiotic Stresses in Plants; IntechOpen: London, UK, 2019; pp. 1–6. [Google Scholar]
- Mertens, D.; Boege, K.; Kessler, A.; Koricheva, J.; Thaler, J.S.; Whiteman, N.K.; Poelman, E.H. Predictability of Biotic Stress Structures Plant Defence Evolution. Trends Ecol. Evol. 2021, 36, 444–456. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS Regulation during Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Piyatida, P.; da Silva, J.A.T.; Fujita, M. Molecular Mechanism of Heavy Metal Toxicity and Tolerance in Plants: Central Role of Glutathione in Detoxification of Reactive Oxygen Species and Methylglyoxal and in Heavy Metal Chelation. J. Bot. 2012, 2012, 1–37. [Google Scholar] [CrossRef]
- Chibuike, G.U.; Obiora, S.C. Heavy Metal Polluted Soils: Effect on Plants and Bioremediation Methods. Appl. Environ. Soil Sci. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chandrakar, V.; Naithani, S.C.; Keshavkant, S. Arsenic-Induced Metabolic Disturbances and Their Mitigation Mechanisms in Crop Plants: A Review. Biologia Bratisl. 2016, 71, 367–377. [Google Scholar] [CrossRef]
- Chen, T.; Su, Y. Uptake by Rice Seedlings and In-Plant Degradation of Atrazine as Influenced by the Oxidative Stress Induced by Added Arsenic or Phosphate Deficiency. Hum. Ecol. Risk Assess. Int. J. 2018, 24, 1550–1564. [Google Scholar] [CrossRef]
- Farnese, F.S.; Oliveira, J.; Farnese, M.S.; Gusman, G.S.; Silveira, N.M.; Siman, L.I. Uptake Arsenic by Plants: Effects on Mineral Nutrition, Growth and Antioxidant Capacity. Idesia 2014, 32, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Edelstein, M.; Ben-Hur, M. Heavy Metals and Metalloids: Sources, Risks and Strategies to Reduce Their Accumulation in Horticultural Crops. Sci. Hortic. 2018, 234, 431–444. [Google Scholar] [CrossRef]
- Dong, J.; Wu, F.; Zhang, G. Influence of Cadmium on Antioxidant Capacity and Four Microelement Concentrations in Tomato Seedlings (Lycopersicon esculentum). Chemosphere 2006, 64, 1659–1666. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demiral, T.; Türkan, I. Comparative Lipid Peroxidation, Antioxidant Defense Systems and Proline Content in Roots of Two Rice Cultivars Differing in Salt Tolerance. Environ. Exp. Bot. 2005, 53, 247–257. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Regulation in Photosynthetic Organisms: Signaling, Acclimation, and Practical Implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and Its Relationship to Plant Hormones. Ann. Bot. 2018, 121, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Alamri, S.; Alsubaie, Q.D.; Ali, H.M.; Ibrahim, A.A.; Alsadon, A. Potential Roles of Melatonin and Sulfur in Alleviation of Lanthanum Toxicity in Tomato Seedlings. Ecotoxicol. Environ. Saf. 2019, 180, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Javeed, H.M.R.; Ali, M.; Skalicky, M.; Nawaz, F.; Qamar, R.; Faheem, M.; Mubeen, M.; Iqbal, M.M.; Vachova, P.; Brestic, M. Lipoic Acid Combined with Melatonin Mitigates Oxidative Stress and Promotes Root Formation and Growth in Salt-Stressed Canola Seedlings (Brassica napus L.). Molecules 2021, 26, 3147. [Google Scholar] [CrossRef]
- Li, X.; Ahammed, G.J.; Zhang, X.-N.; Zhang, L.; Yan, P.; Zhang, L.-P.; Fu, J.-Y.; Han, W.-Y. Melatonin-Mediated Regulation of Anthocyanin Biosynthesis and Antioxidant Defense Confer Tolerance to Arsenic Stress in Camellia sinensis L. J. Hazard. Mater. 2021, 403, 123922. [Google Scholar] [CrossRef]
- Alam, M.N.; Zhang, L.; Yang, L.; Islam, M.R.; Liu, Y.; Luo, H.; Yang, P.; Wang, Q.; Chan, Z. Transcriptomic Profiling of Tall Fescue in Response to Heat Stress and Improved Thermotolerance by Melatonin and 24-Epibrassinolide. BMC Genom. 2018, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Y.; Roohi, M.; Iqbal, Z.; Ashraf, M.; Öztürk, M.; Gücel, S. Cadmium (Cd) and Lead (Pb) Induced Changes in Growth, Some Biochemical Attributes, and Mineral Accumulation in Two Cultivars of Mung Bean [Vigna radiata (L.) Wilczek]. Commun. Soil Sci. Plant Anal. 2016, 47, 405–413. [Google Scholar]
- Alaraidh, I.; Alsahli, A.; Razik, E.A. Alteration of Antioxidant Gene Expression in Response to Heavy Metal Stress in Trigonella Foenum-Graecum L. South Afr. J. Bot. 2018, 115, 90–93. [Google Scholar] [CrossRef]
- Abbas, M.; Akmal, M.; Ullah, S.; Hassan, M.; Farooq, S. Effectiveness of Zinc and Gypsum Application against Cadmium Toxicity and Accumulation in Wheat (Triticum aestivum L.). Commun. Soil Sci. Plant Anal. 2017, 48, 1659–1668. [Google Scholar] [CrossRef]
- Hasan, M.; Cheng, Y.; Kanwar, M.K.; Chu, X.-Y.; Ahammed, G.J.; Qi, Z.-Y. Responses of Plant Proteins to Heavy Metal Stress—a Review. Front. Plant Sci. 2017, 8, 1492. [Google Scholar] [CrossRef] [Green Version]
- Majeed, A.; Muhammad, Z.; Siyar, S. Assessment of Heavy Metal Induced Stress Responses in Pea (Pisum sativum L.). Acta Ecol. Sin. 2019, 39, 284–288. [Google Scholar] [CrossRef]
- Asati, A.; Pichhode, M.; Nikhil, K. Effect of Heavy Metals on Plants: An Overview. Int. J. Appl. Innov. Eng. Manag. 2016, 5, 56–66. [Google Scholar]
- Shekari, L.; Aroiee, H.; Mirshekari, A.; Nemati, H. Protective Role of Selenium on Cucumber (Cucumis sativus L.) Exposed to Cadmium and Lead Stress during Reproductive Stage Role of Selenium on Heavy Metals Stress. J. Plant Nutr. 2019, 42, 529–542. [Google Scholar] [CrossRef]
- Farid, M.; Ali, S.; Rizwan, M.; Ali, Q.; Abbas, F.; Bukhari, S.A.H.; Saeed, R.; Wu, L. Citric Acid Assisted Phytoextraction of Chromium by Sunflower; Morpho-Physiological and Biochemical Alterations in Plants. Ecotoxicol. Environ. Saf. 2017, 145, 90–102. [Google Scholar] [CrossRef]
- Song, B.; Hao, X.; Wang, X.; Yang, S.; Dong, Y.; Ding, Y.; Wang, Q.; Wang, X.; Zhou, J. Boron Stress Inhibits Beet (Beta vulgaris L.) Growth through Influencing Endogenous Hormones and Oxidative Stress Response. Soil Sci. Plant Nutr. 2019, 65, 346–352. [Google Scholar] [CrossRef]
- Huihui, Z.; Xin, L.; Zisong, X.; Yue, W.; Zhiyuan, T.; Meijun, A.; Yuehui, Z.; Wenxu, Z.; Nan, X.; Guangyu, S. Toxic Effects of Heavy Metals Pb and Cd on Mulberry (Morus alba L.) Seedling Leaves: Photosynthetic Function and Reactive Oxygen Species (ROS) Metabolism Responses. Ecotoxicol. Environ. Saf. 2020, 195, 110469. [Google Scholar] [CrossRef]
- Wang, S.; Chen, F.; Mu, S.; Zhang, D.; Pan, X.; Lee, D.-J. Simultaneous Analysis of Photosystem Responses of Microcystis Aeruginoga under Chromium Stress. Ecotoxicol. Environ. Saf. 2013, 88, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Strand, D.D.; Kramer, D.M.; Cousins, A.B. The Response of Cyclic Electron Flow around Photosystem I to Changes in Photorespiration and Nitrate Assimilation. Plant Physiol. 2014, 165, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xu, Z.; Guo, K.; Huo, Y.; He, G.; Sun, H.; Guan, Y.; Xu, N.; Yang, W.; Sun, G. Toxic Effects of Heavy Metal Cd and Zn on Chlorophyll, Carotenoid Metabolism and Photosynthetic Function in Tobacco Leaves Revealed by Physiological and Proteomics Analysis. Ecotoxicol. Environ. Saf. 2020, 202, 110856. [Google Scholar] [CrossRef] [PubMed]
- Gálusová, T.; Piršelová, B.; Rybanský, Ľ.; Krasylenko, Y.; Mészáros, P.; Blehová, A.; Bardáčová, M.; Moravčíková, J.; Matušíková, I. Plasticity of Soybean Stomatal Responses to Arsenic and Cadmium at the Whole Plant Level. Pol. J. Environ. Stud. 2020, 29, 3569–3580. [Google Scholar] [CrossRef]
- Rucińska-Sobkowiak, R. Water Relations in Plants Subjected to Heavy Metal Stresses. Acta Physiol. Plant. 2016, 38, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Zhang, X.; Wang, G. Effects of Heavy Metals on Stomatal Movements in Broad Bean Leaves. Russ. J. Plant Physiol. 2004, 51, 464–468. [Google Scholar] [CrossRef]
- Perfus-Barbeoch, L.; Leonhardt, N.; Vavasseur, A.; Forestier, C. Heavy Metal Toxicity: Cadmium Permeates through Calcium Channels and Disturbs the Plant Water Status. Plant J. 2002, 32, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Sarwat, M.; Bhat, N.A.; Wani, M.R.; Kazi, A.G.; Tran, L.-S.P. Alleviation of Cadmium Toxicity in Brassica juncea L. (Czern. & Coss.) by Calcium Application Involves Various Physiological and Biochemical Strategies. PLoS ONE 2015, 10, e0114571. [Google Scholar]
- Sunil, B.; Saini, D.; Bapatla, R.B.; Aswani, V.; Raghavendra, A.S. Photorespiration Is Complemented by Cyclic Electron Flow and the Alternative Oxidase Pathway to Optimize Photosynthesis and Protect against Abiotic Stress. Photosynth. Res. 2019, 139, 67–79. [Google Scholar] [CrossRef]
- Reiter, R.J.; Paredes, S.D.; Manchester, L.C.; Tan, D.-X. Reducing Oxidative/Nitrosative Stress: A Newly-Discovered Genre for Melatonin. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 175–200. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Malvankar, M.R.; Karle, S.B.; Kumar, K. Reactive Nitrogen Species: Paradigms of Cellular Signaling and Regulation of Salt Stress in Plants. Environ. Exp. Bot. 2019, 161, 86–97. [Google Scholar] [CrossRef]
- Mano, J. Reactive Carbonyl Species: Their Production from Lipid Peroxides, Action in Environmental Stress, and the Detoxification Mechanism. Plant Physiol. Biochem. 2012, 59, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.S.; Mano, J. Lipid Peroxide-Derived Short-Chain Carbonyls Mediate Hydrogen Peroxide-Induced and Salt-Induced Programmed Cell Death in Plants. Plant Physiol. 2015, 168, 885–898. [Google Scholar] [CrossRef] [Green Version]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [Green Version]
- Colangelo, E.P.; Guerinot, M.L. Put the Metal to the Petal: Metal Uptake and Transport throughout Plants. Curr. Opin. Plant Biol. 2006, 9, 322–330. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of Enzymatic and Nonenzymatic Antioxidants in Plants during Abiotic Stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Bharwana, S.; Ali, S.; Farooq, M.; Iqbal, N.; Abbas, F.; Ahmad, M. Alleviation of Lead Toxicity by Silicon Is Related to Elevated Photosynthesis, Antioxidant Enzymes Suppressed Lead Uptake and Oxidative Stress in Cotton. J. Bioremed. Biodeg 2013, 4, 187. [Google Scholar]
- Bashri, G.; Prasad, S.M. Indole Acetic Acid Modulates Changes in Growth, Chlorophyll a Fluorescence and Antioxidant Potential of Trigonella Foenum-Graecum L. Grown under Cadmium Stress. Acta Physiol. Plant. 2015, 37, 49. [Google Scholar] [CrossRef]
- Nadgórska-Socha, A.; Ptasiński, B.; Kita, A. Heavy Metal Bioaccumulation and Antioxidative Responses in Cardaminopsis Arenosa and Plantago Lanceolata Leaves from Metalliferous and Non-Metalliferous Sites: A Field Study. Ecotoxicology 2013, 22, 1422–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flora, S.; Mittal, M.; Mehta, A. Heavy Metal Induced Oxidative Stress & Its Possible Reversal by Chelation Therapy. Indian J. Med. Res. 2008, 128, 501. [Google Scholar]
- Guo, H.; Hong, C.; Xiao, M.; Chen, X.; Chen, H.; Zheng, B.; Jiang, D. Real-Time Kinetics of Cadmium Transport and Transcriptomic Analysis in Low Cadmium Accumulator Miscanthus Sacchariflorus. Planta 2016, 244, 1289–1302. [Google Scholar] [CrossRef]
- Farooq, M.; Ali, S.; Hameed, A.; Bharwana, S.; Rizwan, M.; Ishaque, W.; Farid, M.; Mahmood, K.; Iqbal, Z. Cadmium Stress in Cotton Seedlings: Physiological, Photosynthesis and Oxidative Damages Alleviated by Glycinebetaine. South Afr. J. Bot. 2016, 104, 61–68. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R. Cadmium Uptake and Its Toxicity in Higher Plants. In Cadmium Toxicity and Tolerance in Plants; Narosa Publishing House: New Delhi, India, 2006; pp. 63–86. [Google Scholar]
- Asgher, M.; Khan, N.A.; Khan, M.I.R.; Fatma, M.; Masood, A. Ethylene Production Is Associated with Alleviation of Cadmium-Induced Oxidative Stress by Sulfur in Mustard Types Differing in Ethylene Sensitivity. Ecotoxicol. Environ. Saf. 2014, 106, 54–61. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Mobin, M.; Anjum, N.A.; Khan, N.A. Modulation and Significance of Nitrogen and Sulfur Metabolism in Cadmium Challenged Plants. Plant Growth Regul. 2016, 78, 1–11. [Google Scholar] [CrossRef]
- Gjorgieva, D.; Panovska, T.K.; Ruskovska, T.; Bačeva, K.; Stafilov, T. Mineral Nutrient Imbalance, Total Antioxidants Level and DNA Damage in Common Bean (Phaseolus vulgaris L.) Exposed to Heavy Metals. Physiol. Mol. Biol. Plants 2013, 19, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, M.; Dumat, C.; Khalid, S.; Schreck, E.; Xiong, T.; Niazi, N.K. Foliar Heavy Metal Uptake, Toxicity and Detoxification in Plants: A Comparison of Foliar and Root Metal Uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demecsová, L.; Tamás, L. Reactive Oxygen Species, Auxin and Nitric Oxide in Metal-Stressed Roots: Toxicity or Defence. BioMetals 2019, 32, 717–744. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Abbas, T.; Zia-ur-Rehman, M.; Hannan, F.; Keller, C.; Al-Wabel, M.I.; Ok, Y.S. Cadmium Minimization in Wheat: A Critical Review. Ecotoxicol. Environ. Saf. 2016, 130, 43–53. [Google Scholar] [CrossRef]
- Naeem, A.; Zafar, M.; Khalid, H.; Zia-ur-Rehman, M.; Ahmad, Z.; Ayub, M.A.; Qayyum, M.F. Cadmium-Induced Imbalance in Nutrient and Water Uptake by Plants. In Cadmium Toxicity and Tolerance in Plants; Elsevier: London, UK, 2019; pp. 299–326. [Google Scholar]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Farid, M.; Ali, S.; Rizwan, M.; Saeed, R.; Tauqeer, H.M.; Sallah-Ud-Din, R.; Azam, A.; Raza, N. Microwave Irradiation and Citric Acid Assisted Seed Germination and Phytoextraction of Nickel (Ni) by Brassica napus L.: Morpho-Physiological and Biochemical Alterations under Ni Stress. Environ. Sci. Pollut. Res. 2017, 24, 21050–21064. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Dubey, G.; Prasad, S.M. Exogenous Proline Application Ameliorates Toxic Effects of Arsenate in Solanum melongena L. Seedlings. Ecotoxicol. Environ. Saf. 2015, 117, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Chemical Stress by Different Agents Affects the Melatonin Content of Barley Roots. J. Pineal Res. 2009, 46, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Growth Conditions Determine Different Melatonin Levels in L Upinus albus L. J. Pineal Res. 2013, 55, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Chen, Z.; Yu, X.; Cui, W.; Pan, J.; Zhao, G.; Xu, S.; Wang, R.; Shen, W. Melatonin Confers Plant Tolerance against Cadmium Stress via the Decrease of Cadmium Accumulation and Reestablishment of MicroRNA-Mediated Redox Homeostasis. Plant Sci. 2017, 261, 28–37. [Google Scholar] [CrossRef]
- Li, M.; Hasan, M.K.; Li, C.; Ahammed, G.J.; Xia, X.; Shi, K.; Zhou, Y.; Reiter, R.J.; Yu, J.; Xu, M. Melatonin Mediates Selenium-induced Tolerance to Cadmium Stress in Tomato Plants. J. Pineal Res. 2016, 61, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Lee, H.; Lee, H.Y.; Back, K. Cloning and Functional Characterization of the Arabidopsis N-acetylserotonin O-methyltransferase Responsible for Melatonin Synthesis. J. Pineal Res. 2016, 60, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Zhang, Y.; Xu, Y.; Qi, Z.; Li, M.; Ahammed, G.J.; Xia, X.; Shi, K.; Zhou, Y.; Reiter, R.J. HsfA1a Upregulates Melatonin Biosynthesis to Confer Cadmium Tolerance in Tomato Plants. J. Pineal Res. 2017, 62, e12387. [Google Scholar] [CrossRef]
- Lee, H.-Y.; Back, K. Cadmium Disrupts Subcellular Organelles, Including Chloroplasts, Resulting in Melatonin Induction in Plants. Molecules 2017, 22, 1791. [Google Scholar] [CrossRef] [Green Version]
- Back, K.; Tan, D.; Reiter, R.J. Melatonin Biosynthesis in Plants: Multiple Pathways Catalyze Tryptophan to Melatonin in the Cytoplasm or Chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Wang, L.; Feng, C.; Zheng, X.; Guo, Y.; Zhou, F.; Shan, D.; Liu, X.; Kong, J. Plant Mitochondria Synthesize Melatonin and Enhance the Tolerance of Plants to Drought Stress. J. Pineal Res. 2017, 63, e12429. [Google Scholar] [CrossRef]
- Zhang, J.; Zeng, B.; Mao, Y.; Kong, X.; Wang, X.; Yang, Y.; Zhang, J.; Xu, J.; Rengel, Z.; Chen, Q. Melatonin Alleviates Aluminium Toxicity through Modulating Antioxidative Enzymes and Enhancing Organic Acid Anion Exudation in Soybean. Funct. Plant Biol. 2017, 44, 961–968. [Google Scholar] [CrossRef]
- Tang, Y.; Sun, G.; Miao, M.; Li, Y.; Li, H. Effects of Exogenous Melatonin on Growth and Physiological Characteristics of Chinese Cabbage Seedlings under Aluminum Stress. Adv. Eng. Res. 2017, 143, 1119–1123. [Google Scholar]
- Nazarian, M.; Ghanati, F. The Role of Melatonin in Reinforcement of Antioxidant System of Rice Plant (Oryza sativa L.) under Arsenite Toxicity? Plant Physiol. Rep. 2020, 25, 395–404. [Google Scholar] [CrossRef]
- Farouk, S.; Al-Amri, S.M. Exogenous Melatonin-Mediated Modulation of Arsenic Tolerance with Improved Accretion of Secondary Metabolite Production, Activating Antioxidant Capacity and Improved Chloroplast Ultrastructure in Rosemary Herb. Ecotoxicol. Environ. Saf. 2019, 180, 333–347. [Google Scholar] [CrossRef]
- Lv, X.; Fang, Y.; Zhang, L.; Zhang, W.; Xu, L.; Han, J.; Jin, B.; Zhang, X.; Zhang, X.; Xue, D. Effects of Melatonin on Growth, Physiology and Gene Expression in Rice Seedlings under Cadmium Stress. Phyton 2019, 88, 91. [Google Scholar] [CrossRef]
- Wang, M.; Duan, S.; Zhou, Z.; Chen, S.; Wang, D. Foliar spraying of melatonin confers cadmium tolerance in Nicotiana tabacum L. Ecotoxicol. Environ. Saf. 2019, 170, 68–76. [Google Scholar] [CrossRef]
- He, J.; Zhuang, X.; Zhou, J.; Sun, L.; Wan, H.; Li, H.; Lyu, D. Exogenous Melatonin Alleviates Cadmium Uptake and Toxicity in Apple Rootstocks. Tree Physiol. 2020, 40, 746–761. [Google Scholar] [CrossRef]
- Wu, X.; Li, H.; Luo, H.; Lin, D.; Zhou, X. Effect of Exogenous Melatonin on Photosynthetic Characteristics in Cucumber Seedlings under Cadmium Stress: A Rapid Detection Method for the Cadmium Resistance. E3S Web Conf. 2019, 136, 07023. [Google Scholar] [CrossRef]
- Lin, L.; Li, J.; Chen, F.; Liao, M.A.; Tang, Y.; Liang, D.; Xia, H.; Lai, Y.; Wang, X.; Chen, C.; et al. Effects of melatonin on the growth and cadmium characteristics of Cyphomandra betacea seedlings. Environ. Monit. Assess. 2018, 190, 1–8. [Google Scholar] [CrossRef]
- Asif, M.; Pervez, A.; Irshad, U.; Mehmood, Q.; Ahmad, R. Melatonin and Plant Growth-Promoting Rhizobacteria Alleviate the Cadmium and Arsenic Stresses and Increase the Growth of Spinacia oleracea L. Plant Soil Environ. 2020, 66, 234–241. [Google Scholar] [CrossRef]
- Hodžić, E.; Balaban, M.; Šuškalo, N.; Galijašević, S.; Hasanagić, D.; Kukavica, B. Antioxidative Response of Melissa officinalis L. and Valeriana officinalis L. Leaves Exposed to Exogenous Melatonin and Excessive Zinc and Cadmium Levels. J. Serbian Chem. Soc. 2019, 84, 11–25. [Google Scholar] [CrossRef] [Green Version]
- Ayyaz, A.; Farooq, M.A.; Kanwal, A.; Aslam, M.; Iqbal, M.; Manzoor, A.; Khalid, A.; Umer, S.; Bano, H.; Rasool, B. Differential Responses of Exogenous Melatonin on Growth, Photosynthesis and Antioxidant Defence System in Two Brassica napus L. Cultivars under Chromium Stress. Int. J. Environ. Agric. Biotechnol. 2020, 5, 397–411. [Google Scholar]
- Hu, Z.; Fu, Q.; Zheng, J.; Zhang, A.; Wang, H. Transcriptomic and Metabolomic Analyses Reveal That Melatonin Promotes Melon Root Development under Copper Stress by Inhibiting Jasmonic Acid Biosynthesis. Hortic. Res. 2020, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-Y.; Qi, C.-D.; Li, S.; Wang, Z.; Wang, X.; Wang, J.; Ren, S.; Li, X.; Zhang, N.; Guo, Y.-D. Melatonin Alleviates Copper Toxicity via Improving Copper Sequestration and ROS Scavenging in Cucumber. Plant Cell Physiol. 2019, 60, 562–574. [Google Scholar] [CrossRef]
- Kholodova, V.; Vasil’ev, S.; Efimova, M.; Voronin, P.Y.; Rakhmankulova, Z.; Danilova, E.Y.; Kuznetsov, V.V. Exogenous Melatonin Protects Canola Plants from Toxicity of Excessive Copper. Russ. J. Plant Physiol. 2018, 65, 882–889. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Integrative Roles of Nitric Oxide and Hydrogen Sulfide in Melatonin-induced Tolerance of Pepper (Capsicum annuum L.) Plants to Iron Deficiency and Salt Stress Alone or in Combination. Physiol. Plant. 2020, 168, 256–277. [Google Scholar] [CrossRef] [Green Version]
- Ahammed, G.J.; Wu, M.; Wang, Y.; Yan, Y.; Mao, Q.; Ren, J.; Ma, R.; Liu, A.; Chen, S. Melatonin Alleviates Iron Stress by Improving Iron Homeostasis, Antioxidant Defense and Secondary Metabolism in Cucumber. Sci. Hortic. 2020, 265, 109205. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Baloch, A.R.; Sun, J.; Shu, S.; Wang, Y.; Ahammed, G.J.; Kabir, K.; Roy, R. Melatonin Alleviates Nickel Phytotoxicity by Improving Photosynthesis, Secondary Metabolism and Oxidative Stress Tolerance in Tomato Seedlings. Ecotoxicol. Environ. Saf. 2020, 197, 110593. [Google Scholar] [CrossRef] [PubMed]
- Okant, M.; Kaya, C. The Role of Endogenous Nitric Oxide in Melatonin-Improved Tolerance to Lead Toxicity in Maize Plants. Environ. Sci. Pollut. Res. 2019, 26, 11864–11874. [Google Scholar] [CrossRef]
- Namdjoyan, S.; Soorki, A.A.; Elyasi, N.; Kazemi, N.; Simaei, M. Melatonin Alleviates Lead-Induced Oxidative Damage in Safflower (Carthamus tinctorius L.) Seedlings. Ecotoxicology 2020, 29, 108–118. [Google Scholar] [CrossRef]
- Kobylińska, A.; Posmyk, M.M. Melatonin Restricts Pb-Induced PCD by Enhancing BI-1 Expression in Tobacco Suspension Cells. Biometals 2016, 29, 1059–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Xiong, X.; Huang, Z.; Sun, L.; Ma, J.; Cai, S.; Yu, F.; Zhong, W.; Chen, S.; Li, X. Exogenous Melatonin Improves Lead Tolerance of Bermudagrass through Modulation of the Antioxidant Defense System. Int. J. Phytoremediat. 2018, 20, 1408–1417. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Li, H.; Li, X.; Cao, Y.; Zhang, H.; Li, S.; Zhang, L.; Qi, Y.; Ren, S. Melatonin Improved Anthocyanin Accumulation by Regulating Gene Expressions and Resulted in High Reactive Oxygen Species Scavenging Capacity in Cabbage. Front. Plant Sci. 2016, 7, 197. [Google Scholar] [CrossRef]
- Al-Huqail, A.A.; Khan, M.N.; Ali, H.M.; Siddiqui, M.H.; Al-Huqail, A.A.; AlZuaibr, F.M.; Al-Muwayhi, M.A.; Marraiki, N.; Al-Humaid, L. Exogenous Melatonin Mitigates Boron Toxicity in Wheat. Ecotoxicol. Environ. Saf. 2020, 201, 110822. [Google Scholar] [CrossRef]
- Chan, Z.; Shi, H. Improved Abiotic Stress Tolerance of Bermudagrass by Exogenous Small Molecules. Plant Signal. Behav. 2015, 10, e991577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical Priming of Plants against Multiple Abiotic Stresses: Mission Possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-Z.; Zhang, L.-H.; Jun, M.; Li, X.-Y.; Yan, L.; Zhang, R.-P.; Wang, R.-Q. Effects of Water Stress on Reactive Oxygen Species Generation and Protection System in Rice during Grain-Filling Stage. Agric. Sci. China 2010, 9, 633–641. [Google Scholar] [CrossRef]
- Kacienė, G.; Juknys, R.; Januškaitienė, I. The Role of Oxidative Stress in Spring Barley Cross-Adaptation to Different Heavy Metals. Arch. Agron. Soil Sci. 2017, 63, 1037–1048. [Google Scholar] [CrossRef]
- Zhang, M.; Fang, Y.; Ji, Y.; Jiang, Z.; Wang, L. Effects of Salt Stress on Ion Content, Antioxidant Enzymes and Protein Profile in Different Tissues of Broussonetia Papyrifera. South Afr. J. Bot. 2013, 85, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of Melatonin in Alleviating Cold Stress in A Rabidopsis Thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.-X.; Reiter, R.J.; Zhang, H.; Liu, R.; Chan, Z. Comparative Physiological, Metabolomic, and Transcriptomic Analyses Reveal Mechanisms of Improved Abiotic Stress Resistance in Bermudagrass [Cynodon dactylon (L). Pers.] by Exogenous Melatonin. J. Exp. Bot. 2015, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.A.; Jiao, Y.; Chen, C.; Shireen, F.; Zheng, Z.; Imtiaz, M.; Bie, Z.; Huang, Y. Melatonin Pretreatment Improves Vanadium Stress Tolerance of Watermelon Seedlings by Reducing Vanadium Concentration in the Leaves and Regulating Melatonin Biosynthesis and Antioxidant-Related Gene Expression. J. Plant Physiol. 2018, 220, 115–127. [Google Scholar] [CrossRef]
- Nabaei, M.; Amooaghaie, R. Nitric Oxide Is Involved in the Regulation of Melatonin-Induced Antioxidant Responses in Catharanthus Roseus Roots under Cadmium Stress. Botany 2019, 97, 681–690. [Google Scholar] [CrossRef]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.; Reiter, R.J. Melatonin: An Ancient Molecule That Makes Oxygen Metabolically Tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef]
- Gong, B.; Yan, Y.; Wen, D.; Shi, Q. Hydrogen Peroxide Produced by NADPH Oxidase: A Novel Downstream Signaling Pathway in Melatonin-induced Stress Tolerance in Solanum Lycopersicum. Physiol. Plant. 2017, 160, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-Mediated Nitric Oxide Improves Tolerance to Cadmium Toxicity by Reducing Oxidative Stress in Wheat Plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Xalxo, R.; Keshavkant, S. Melatonin, Glutathione and Thiourea Attenuates Lead and Acid Rain-Induced Deleterious Responses by Regulating Gene Expression of Antioxidants in Trigonella Foenum Graecum L. Chemosphere 2019, 221, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sami, A.; Shah, F.A.; Abdullah, M.; Zhou, X.; Yan, Y.; Zhu, Z.; Zhou, K. Melatonin Mitigates Cadmium and Aluminium Toxicity through Modulation of Antioxidant Potential in Brassica napus L. Plant Biol. 2020, 22, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Ke, M.; Zhang, M.; Peijnenburg, W.; Fan, X.; Xu, J.; Zhang, Z.; Lu, T.; Fu, Z.; Qian, H. The Interactive Effects of Diclofop-Methyl and Silver Nanoparticles on Arabidopsis Thaliana: Growth, Photosynthesis and Antioxidant System. Environ. Pollut. 2018, 232, 212–219. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of Melatonin in Plants: A Review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Mayo, J.C.; Tan, D.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wang, Q.; Shah, F.A.; Liu, W.; Wang, D.; Huang, S.; Fu, S.; Wu, L. Exogenous Melatonin Confers Cadmium Tolerance by Counterbalancing the Hydrogen Peroxide Homeostasis in Wheat Seedlings. Molecules 2018, 23, 799. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Wang, Y.; Zhang, J.; Gong, X.; Zhang, Z.; Sun, J.; Chen, X.; Wang, Y. Exogenous Melatonin Improves Physiological Characteristics and Promotes Growth of Strawberry Seedlings under Cadmium Stress. Hortic. Plant J. 2021, 7, 13–22. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, R.; Meng, J.; Li, Z.; Wu, Y.; Tao, J. Ameliorative Effects of Melatonin on Dark-Induced Leaf Senescence in Gardenia (Gardenia Jasminoides Ellis): Leaf Morphology, Anatomy, Physiology and Transcriptome. Sci. Rep. 2017, 7, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.-G.; Liu, W.; Yu, W.; Zhang, Y.; Ding, S.; Li, H.; Mrak, T.; Kraigher, H.; Luo, Z.-B. Abscisic Acid Enhances Lead Translocation from the Roots to the Leaves and Alleviates Its Toxicity in Populus× Canescens. J. Hazard. Mater. 2019, 362, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Banu Doğanlar, Z. Metal Accumulation and Physiological Responses Induced by Copper and Cadmium in Lemna gibba L. Minor and Spirodela Polyrhiza. Chem. Speciat. Bioavailab. 2013, 25, 79–88. [Google Scholar] [CrossRef]
- Zhao, H.; Su, T.; Huo, L.; Wei, H.; Jiang, Y.; Xu, L.; Ma, F. Unveiling the Mechanism of Melatonin Impacts on Maize Seedling Growth: Sugar Metabolism as a Case. J. Pineal Res. 2015, 59, 255–266. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, C.; Wang, Z.; Sun, S.; Zhan, R.; Zhao, Y.; Ma, B.; Ma, F.; Li, M. Melatonin-Mediated Sugar Accumulation and Growth Inhibition in Apple Plants Involves down-Regulation of Fructokinase 2 Expression and Activity. Front. Plant Sci. 2019, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S. Melatonin Promotes Ripening and Improves Quality of Tomato Fruit during Postharvest Life. J. Exp. Bot. 2015, 66, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Wolf, S.; Han, M.; Zhao, H.; Wei, H.; Greiner, S.; Rausch, T. Reassessment of an Arabidopsis Cell Wall Invertase Inhibitor AtCIF1 Reveals Its Role in Seed Germination and Early Seedling Growth. Plant Mol. Biol. 2016, 90, 137–155. [Google Scholar] [CrossRef]
- Gouia, H.; Ghorbal, M.H.; Meyer, C. Effects of Cadmium on Activity of Nitrate Reductase and on Other Enzymes of the Nitrate Assimilation Pathway in Bean. Plant Physiol. Biochem. 2000, 38, 629–638. [Google Scholar] [CrossRef]
- Balestrasse, K.B.; Benavides, M.P.; Gallego, S.M.; Tomaro, M.L. Effect of Cadmium Stress on Nitrogen Metabolism in Nodules and Roots of Soybean Plants. Funct. Plant Biol. 2003, 30, 57–64. [Google Scholar] [CrossRef]
- Lea, P.J.; Miflin, B.J. Glutamate Synthase and the Synthesis of Glutamate in Plants. Plant Physiol. Biochem. 2003, 41, 555–564. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant Salt-Tolerance Mechanism: A Review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Qiao, Y.; Yin, L.; Wang, B.; Ke, Q.; Deng, X.; Wang, S. Melatonin Promotes Plant Growth by Increasing Nitrogen Uptake and Assimilation under Nitrogen Deficient Condition in Winter Wheat. Plant Physiol. Biochem. 2019, 139, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Malar, S.; Manikandan, R.; Favas, P.J.; Sahi, S.V.; Venkatachalam, P. Effect of Lead on Phytotoxicity, Growth, Biochemical Alterations and Its Role on Genomic Template Stability in Sesbania Grandiflora: A Potential Plant for Phytoremediation. Ecotoxicol. Environ. Saf. 2014, 108, 249–257. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.-J. The Significance of Amino Acids and Amino Acid-Derived Molecules in Plant Responses and Adaptation to Heavy Metal Stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [Green Version]
- Seregin, I.; Ivanov, V. Physiological Aspects of Cadmium and Lead Toxic Effects on Higher Plants. Russ. J. Plant Physiol. 2001, 48, 523–544. [Google Scholar] [CrossRef]
- Małecka, A.; Piechalak, A.; Morkunas, I.; Tomaszewska, B. Accumulation of Lead in Root Cells of Pisum Sativum. Acta Physiol. Plant. 2008, 30, 629–637. [Google Scholar] [CrossRef]
- Siripornadulsil, S.; Traina, S.; Verma, D.P.S.; Sayre, R.T. Molecular mechanisms of proline-mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell 2002, 14, 2837–2847. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A.; Fotopoulos, V. Melatonin Systemically Ameliorates Drought Stress-induced Damage in M Edicago Sativa Plants by Modulating Nitro-oxidative Homeostasis and Proline Metabolism. J. Pineal Res. 2017, 62, e12401. [Google Scholar] [CrossRef]
- Jahan, M.S.; Shu, S.; Wang, Y.; Chen, Z.; He, M.; Tao, M.; Sun, J.; Guo, S. Melatonin Alleviates Heat-Induced Damage of Tomato Seedlings by Balancing Redox Homeostasis and Modulating Polyamine and Nitric Oxide Biosynthesis. BMC Plant Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef]
- Fukutoku, Y.; Yamada, Y. Accumulation of Carbohydrates and Proline in Water-Stressed Soybean (Glycine max L.). Soil Sci. Plant Nutr. 1982, 28, 147–151. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, Z.; Zhu, L.; Ma, Z.; Wang, J.; Zhu, J. Exogenous Melatonin Improves Plant Iron Deficiency Tolerance via Increased Accumulation of Polyamine-Mediated Nitric Oxide. Int. J. Mol. Sci. 2016, 17, 1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulhassan, Z.; Huang, Q.; Gill, R.A.; Ali, S.; Mwamba, T.M.; Ali, B.; Hina, F.; Zhou, W. Protective Mechanisms of Melatonin against Selenium Toxicity in Brassica Napus: Insights into Physiological Traits, Thiol Biosynthesis and Antioxidant Machinery. BMC Plant Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Van Hoewyk, D. A Tale of Two Toxicities: Malformed Selenoproteins and Oxidative Stress Both Contribute to Selenium Stress in Plants. Ann. Bot. 2013, 112, 965–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fashola, M.O.; Ngole-Jeme, V.M.; Babalola, O.O. Heavy Metal Pollution from Gold Mines: Environmental Effects and Bacterial Strategies for Resistance. Int. J. Environ. Res. Public. Health 2016, 13, 1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llamas, A.; Ullrich, C.I.; Sanz, A. Cd2+ Effects on Transmembrane Electrical Potential Difference, Respiration and Membrane Permeability of Rice (Oryza Sativa L.) Roots. Plant Soil 2000, 219, 21–28. [Google Scholar] [CrossRef]
- Gévaudant, F.; Duby, G.; von Stedingk, E.; Zhao, R.; Morsomme, P.; Boutry, M. Expression of a Constitutively Activated Plasma Membrane H+−ATPase Alters Plant Development and Increases Salt Tolerance. Plant Physiol. 2007, 144, 1763–1776. [Google Scholar] [CrossRef] [Green Version]
- Hardeland, R.; Balzer, I.; Poeggeler, B.; Fuhrberg, B.; Una, H.; Behrmann, G.; Wolf, R.; Meyer, T.J.; Reiter, R.J. On the Primary Functions of Melatonin in Evolution: Mediation of Photoperiodic Signals in a Unicell, Photooxidation, and Scavenging of Free Radicals. J. Pineal Res. 1995, 18, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.K.; Liu, C.-X.; Pan, Y.-T.; Ahammed, G.J.; Qi, Z.-Y.; Zhou, J. Melatonin Alleviates Low-Sulfur Stress by Promoting Sulfur Homeostasis in Tomato Plants. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Li, X.; Zhang, L.; Ahammed, G.J.; Li, Z.-X.; Wei, J.-P.; Shen, C.; Yan, P.; Zhang, L.-P.; Han, W.-Y. Stimulation in Primary and Secondary Metabolism by Elevated Carbon Dioxide Alters Green Tea Quality in Camellia sinensis L. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Michalak, A. Phenolic Compounds and Their Antioxidant Activity in Plants Growing under Heavy Metal Stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Verdan, A.M.; Wang, H.C.; García, C.R.; Henry, W.P.; Brumaghim, J.L. Iron Binding of 3-Hydroxychromone, 5-Hydroxychromone, and Sulfonated Morin: Implications for the Antioxidant Activity of Flavonols with Competing Metal Binding Sites. J. Inorg. Biochem. 2011, 105, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Gould, K.S. Nature’s Swiss Army Knife: The Diverse Protective Roles of Anthocyanins in Leaves. J. Biomed. Biotechnol. 2004, 2004, 314. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.H.; Chee Kong, Y.; Mohd Zain, N.A. Effect of Cadmium and Copper Exposure on Growth, Secondary Metabolites and Antioxidant Activity in the Medicinal Plant Sambung Nyawa (Gynura Procumbens (Lour.) Merr). Molecules 2017, 22, 1623. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Ueda, Y.; Lai, S.; Frei, M. Shoot Tolerance Mechanisms to Iron Toxicity in Rice (Oryza sativa L.). Plant Cell Environ. 2017, 40, 570–584. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.-Q. Phenylalanine Ammonia-Lyase, a Key Component Used for Phenylpropanoids Production by Metabolic Engineering. RSC Adv. 2015, 5, 62587–62603. [Google Scholar] [CrossRef]
- Sarafi, E.; Tsouvaltzis, P.; Chatzissavvidis, C.; Siomos, A.; Therios, I. Melatonin and Resveratrol Reverse the Toxic Effect of High Boron (B) and Modulate Biochemical Parameters in Pepper Plants (Capsicum annuum L.). Plant Physiol. Biochem. 2017, 112, 173–182. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Cao, Y.; Li, X.; Zhang, H.; Zhang, L.; Tan, D.X.; Guo, Y.D. A label-free differential proteomics analysis reveals the effect of melatonin on promoting fruit ripening and anthocyanin accumulation upon postharvest in tomato. J. Pineal Res. 2016, 61, 138–153. [Google Scholar] [CrossRef]
- Masood, A.; Iqbal, N.; Khan, N.A. Role of Ethylene in Alleviation of Cadmium-induced Photosynthetic Capacity Inhibition by Sulphur in Mustard. Plant Cell Environ. 2012, 35, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Larsson, E.H.; Bornman, J.F.; Asp, H. Influence of UV-B Radiation and Cd2+ on Chlorophyll Fluorescence, Growth and Nutrient Content in Brassica Napus. J. Exp. Bot. 1998, 49, 1031–1039. [Google Scholar] [CrossRef]
- Sarropoulou, V.; Dimassi-Theriou, K.; Therios, I.; Koukourikou-Petridou, M. Melatonin Enhances Root Regeneration, Photosynthetic Pigments, Biomass, Total Carbohydrates and Proline Content in the Cherry Rootstock PHL-C (Prunus Avium× Prunus Cerasus). Plant Physiol. Biochem. 2012, 61, 162–168. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Pan, K.-Y.; Hung, C.-H.; Huang, H.-E.; Chen, C.-L.; Feng, T.-Y.; Huang, L.-F. Overexpression of Ferredoxin, PETF, Enhances Tolerance to Heat Stress in Chlamydomonas Reinhardtii. Int. J. Mol. Sci. 2013, 14, 20913–20929. [Google Scholar] [CrossRef] [Green Version]
- Badger, M.R.; Price, G.D. The Role of Carbonic Anhydrase in Photosynthesis. Annu. Rev. Plant Biol. 1994, 45, 369–392. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.; Shri, M.; Misra, P.; Lakhwani, D.; Bag, S.K.; Asif, M.H.; Trivedi, P.K.; Tripathi, R.D.; Chakrabarty, D. Heavy Metals Induce Oxidative Stress and Genome-Wide Modulation in Transcriptome of Rice Root. Funct. Integr. Genomics 2014, 14, 401–417. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, P.K. Heavy metal stress signaling in plants. In Plant Metal Interaction; Elsevier: London, UK, 2016; pp. 585–603. [Google Scholar]
- Pál, M.; Janda, T.; Szalai, G. Interactions between Plant Hormones and Thiol-Related Heavy Metal Chelators. Plant Growth Regul. 2018, 85, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, A.; Namdjoyan, S.; Soorki, A.A. Effects of Exogenous Melatonin and Glutathione on Zinc Toxicity in Safflower (Carthamus tinctorius L.) Seedlings. Ecotoxicol. Environ. Saf. 2020, 201, 110853. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, F.; Tang, M.; Wang, Y.; Dong, J.; Ying, J.; Chen, Y.; Hu, B.; Li, C.; Liu, L. Melatonin Confers Cadmium Tolerance by Modulating Critical Heavy Metal Chelators and Transporters in Radish Plants. J. Pineal Res. 2020, 69, e12659. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| HMs | Plant Species | Exposed Organs | Concentration (µM) | Exposure Duration | MT Level | Reference |
|---|---|---|---|---|---|---|
| Al | Glycine max | Root | 50 | 24 h | ↑ | [74] |
| Al | Nicotiana tabacum | Leaves | 100 | 15 days | ↑ | [71] |
| Al | Oryza sativa | Seedlings | 500 | 72 h | ↓ | [71] |
| Cd | Medicago sativa | Root | 50, 100, and 200 | 15 h | ↑ | [67] |
| Cd | Solanum lycopersicum | Root | 100 | 15 days | ↑ | [68] |
| Cd | Solanum lycopersicum | Leaves | 100 | 15 days | ↑ | [70] |
| Cd | Nicotiana tabacum | Leaves | 500 | 15 days | ↑ | [71] |
| Cd | Arabidopsis thaliana | Leaves | 300 | 24 h | ↑ | [69] |
| Cd | Oryza sativa | Seedlings | 500 | 72 h | ↑ | [71] |
| Zn | Hordeum vulgare | Root | 100 | 15 h | ↑ | [65] |
| Zn | Hordeum vulgare | Root | 100 | 72 h | ↑ | [65] |
| Zn | Lupinus albus | Root | 100 | 24 h | ↑ | [66] |
| Plant Species | HM (conc.) | MT Doses | Observed Effects of MT on Plant Systems | Reference | |
|---|---|---|---|---|---|
| Increase | Decrease | ||||
| Brassica rapa | Al (50 µM) | 50, 100, 200, and 400 µmol L−1 | SOD, POD, and CAT activity; SP and Chl content; RWC; plant growth and biomass. | MDA content. | [75] |
| Glycine max | 0.1, 1, 10, 100, and 200 mM | CAT, SOD, and POD activity; exudation of malate and citrate; gene encoding acetyltransferase NSI-like (nuclear shuttle protein-interacting); root growth. | H2O2 content. | [74] | |
| Oryza sativa | As (25 µM) | 250 mM | GSH, PCs content; dry matter production. | Oxidative stress; H2O2 and MDA content. | [76] |
| Rosmarinus officinalis | As (75 mg kg−1 soil) | 25, 50, 100, and 200 µM | SOD, POD, and CAT secondary metabolites (AsA, phenols, flavanoids); chloroplast ultrastructure; Chl pigment, essential ions, essential oil, stability, the cell membrane integrity; growth. | H2O2 and MDA content. | [77] |
| Oryza sativa | Cd (10 and 50 µM) | 10 and 50 µM | SOD and POD activity; plant biomass including both underground and above-ground areas. | Accumulation of Cd; transcription of Cd uptake and transport-related genes. | [78] |
| Medicago sativa | Cd (50, 100, and 200 µM) | 10, 50, and 200 µM | Cd tolerance; microRNA-mediated redox homeostasis. | Accumulation of Cd and ROS; oxidative damage. | [67] |
| Nicotiana tabacum | Cd (10, 50, 100, and 200 µM) | 25, 50, 100, and 250 µM | APX, CAT, and POD content; promotion of cell wall or vacuolar sequestration of Cd; plant growth. | Expression of Cd uptake-related genes (IRT1, Nramp1, HMA2, HMA4, and HMA3); photoinhibition. | [79] |
| Malus micromalus | Cd (30 µM) | 0 and 100 µM | Photosynthesis, photosynthetic pigments; transcriptionally regulated key genes involved in detoxification; plant biomass. | Cd-induced reductions in growth; ROS and MDA. | [80] |
| Cucumis sativus | Cd (100 µM) | 150 µmol L−1 | LA; photosynthetic rate; Chl content; stomatal conductance; transpiration rate. | Growth inhibition; excess Cd poisoning. | [81] |
| Solanum lycopersicum | 100 µM | APX, POD, and CAT activity; redox homeostasis; S metabolism, and biosynthesis of downstream S metabolites; H+-ATPase activity; GSH and PCs; plant growth. | Oxidative stress. | [68] | |
| Cyphomandra betacea | Cd (10 mg L−1) | 50 µM | SOD, POD, and CAT activity; biomass of C. betacea seedlings; Cd contents in the stems, leaves, and shoots of C. betacea seedlings. | Seedlings growth inhibition | [82] |
| Spinacia oleracea | Cd-As (25, 75, and 125 ppm) | 100 µM | SOD, POD, and CAT activity; fresh and dry weight. | Chl damage; lipid peroxidation. | [83] |
| Melissa officinalis and Valeriana officinalis | Zn-Cd (3 g L−1 and 15 mg L−1) | 1 µM | POD and CAT activity; SP content. | MDA content; oxidative stress. | [84] |
| Brassica napus | Cr (50 and 100 µM) | 0, 1, 5, and 10 µM | SOD, POD, APX, and CAT activity; photosynthesis rate; photosystem II efficiency; cellular redox potential; plant growth and development. | Cr accumulation; ROS accumulation. | [85] |
| Cucumis melo | Cu (300 µM) | 10, 50, 100, 300, 500, and 800 µmol L−1 | SOD, POD, and CAT activity; GSH which chelates excess Cu2+; redox-related gene expression; cell-wall-related gene expression. | ROS production. | [86] |
| Cucumis sativus | Cu (80 µM) | 10 nmol L−1 | SOD, APX, POD, and GR activity; GSH and PC content; Cu2+ sequestration; carbon metabolism (glycolysis and the pentose phosphate pathway); cell wall trapping; plant fresh weight. | Cu2+ toxicity and ROS production. | [87] |
| Brassica napus | Cu (10–100 µM) | 0.1–100 µM | Plant biomass; photosynthetic pigments; efficiency of photosynthetic apparatus; proline content. | Oxidative stress. | [88] |
| Capsicum annum | Fe (0.1 mM) | 100 µM | POD, SOD, and CAT activity; Chl content; active Fe2+ and K+ content; endogenous NO and H2S; total biomass; fruit yield of plants. | H2O2 and MDA content. | [89] |
| Cucumis sativus | Fe (3 and 90 mg L−1) | 100 µM | Endogenous MT content; SOD, POD, and CAT activity; phenols and flavonoids contents; phenylalanine ammonia lyase, polyphenol oxidase activity; photosynthetic pigment and rate; plant growth and biomass. | ROS production and Fe acquisition. | [90] |
| Solanum lycopersicum | Ni (50 µM) | 100 µM | APX, CAT, SOD, POD and GR activity; redox balance; Chl-synthesis-related genes; photosynthesis rate; phenols, flavonoids, and anthocyanin content; nutrient homeostasis; biomass production. | ROS accumulation. | [91] |
| Zea mays | Pb (0.1 mM) | 0.05 and 0.10 mM | SOD, POD, and CAT activity; nutrient element content; plant growth. | Oxidative stress; H2O2 and MDA content; electrolyte leakage. | [92] |
| Carthamus tinctorius | Pb (50 µM) | 0–300 µM | APX, CAT, SOD, and POD activity; glyoxalase (Gly I and Gly II); Chl and PC content; biomass production of roots, stems and leaves. | Pb stress. | [93] |
| Nicotiana tabacum | Pb (15 µM) | 200 nM | Pb stress tolerance. | Programmed cell death; ROS content; DNA fragmentation. | [94] |
| Cynodon dactylon | Pb (1000 and 2000 mg kg−1 soil) | 0, 5, 20, and 100 µM | SOD, CAT, POD, APX, and GR activity; non-enzymatic antioxidant (AsA and GSH) content; water status; photosynthetic pigments; biomass production. | ROS content; membrane lipid peroxidation and permeability. | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoque, M.N.; Tahjib-Ul-Arif, M.; Hannan, A.; Sultana, N.; Akhter, S.; Hasanuzzaman, M.; Akter, F.; Hossain, M.S.; Sayed, M.A.; Hasan, M.T.; et al. Melatonin Modulates Plant Tolerance to Heavy Metal Stress: Morphological Responses to Molecular Mechanisms. Int. J. Mol. Sci. 2021, 22, 11445. https://doi.org/10.3390/ijms222111445
Hoque MN, Tahjib-Ul-Arif M, Hannan A, Sultana N, Akhter S, Hasanuzzaman M, Akter F, Hossain MS, Sayed MA, Hasan MT, et al. Melatonin Modulates Plant Tolerance to Heavy Metal Stress: Morphological Responses to Molecular Mechanisms. International Journal of Molecular Sciences. 2021; 22(21):11445. https://doi.org/10.3390/ijms222111445
Chicago/Turabian StyleHoque, Md. Najmol, Md. Tahjib-Ul-Arif, Afsana Hannan, Naima Sultana, Shirin Akhter, Md. Hasanuzzaman, Fahmida Akter, Md. Sazzad Hossain, Md. Abu Sayed, Md. Toufiq Hasan, and et al. 2021. "Melatonin Modulates Plant Tolerance to Heavy Metal Stress: Morphological Responses to Molecular Mechanisms" International Journal of Molecular Sciences 22, no. 21: 11445. https://doi.org/10.3390/ijms222111445