
Atrial Fibrillation in Heart Failure Is Associated with High Levels of Circulating microRNA-199a-5p and 22–5p and a Defective Regulation of Intracellular Calcium and Cell-to-Cell Communication

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. HFrEF Patients with AF Have Higher Plasmatic Levels of miRNA-199a-5p and miRNA-22-5p
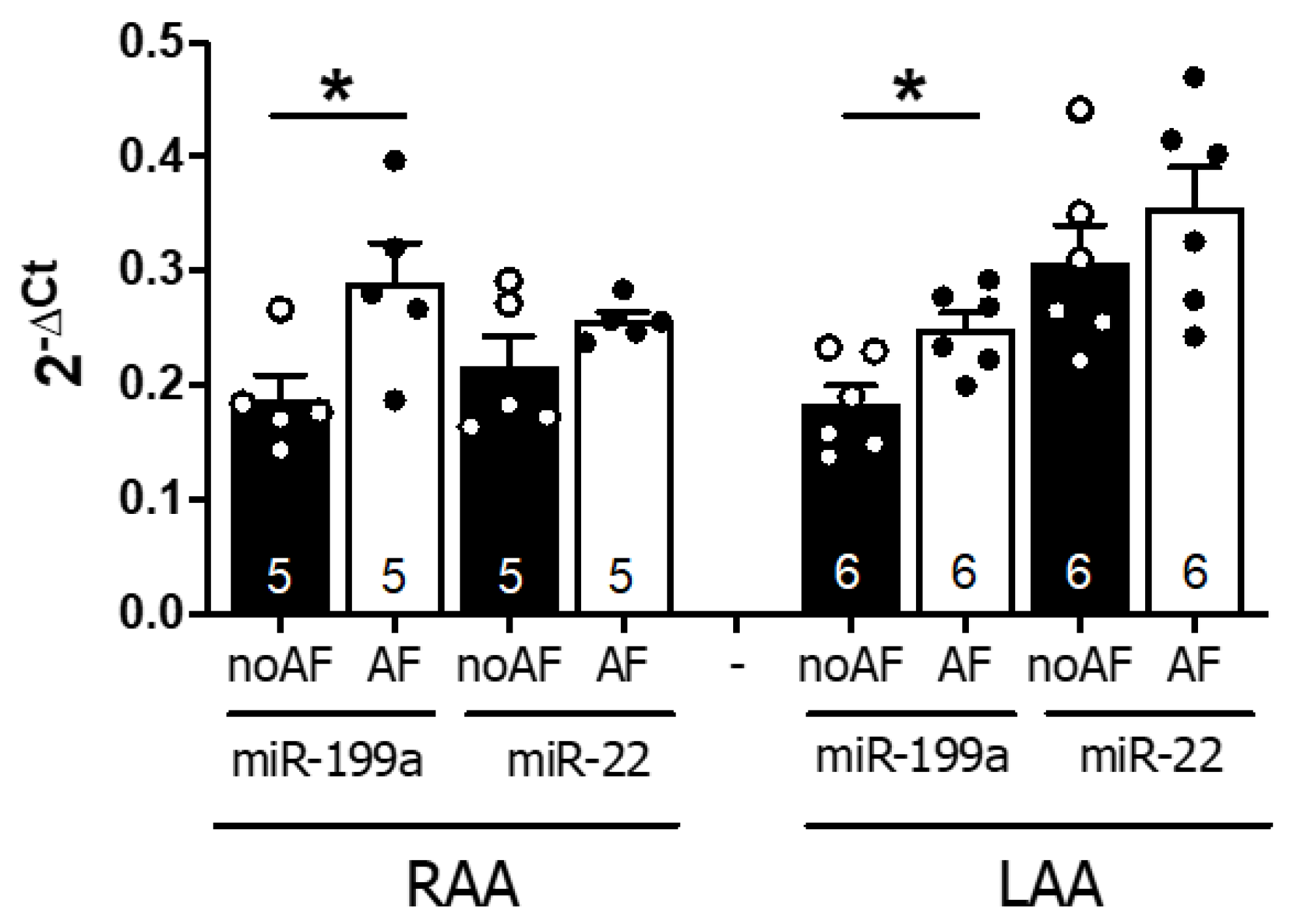
2.2. MiR-199a-5p and 22–5p Expression Levels Are Higher in Atrial Samples of AF Patients
2.3. Overexpression of miR-199a-5p and miR-22-5p Is Associated with Reduced Expression of Proteins Involved in Ca2+ Homeostasis and Cell-to-Cell Communication
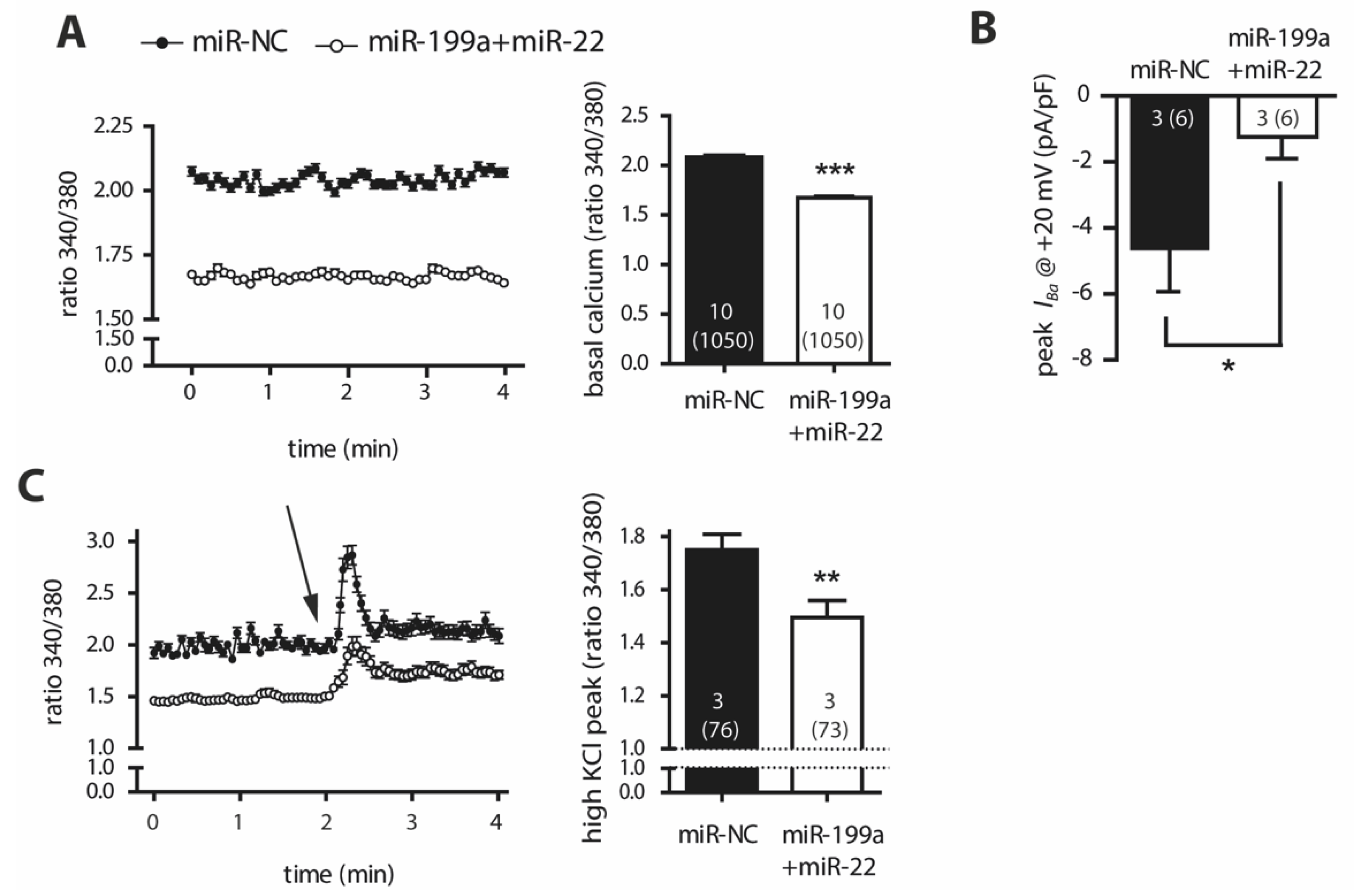
2.4. Overexpression of miR-199a-5p and miR-22-5p Is Associated with Reduced Intracellular Ca2+ Levels and a Decrease in Barium Currents (IBa)
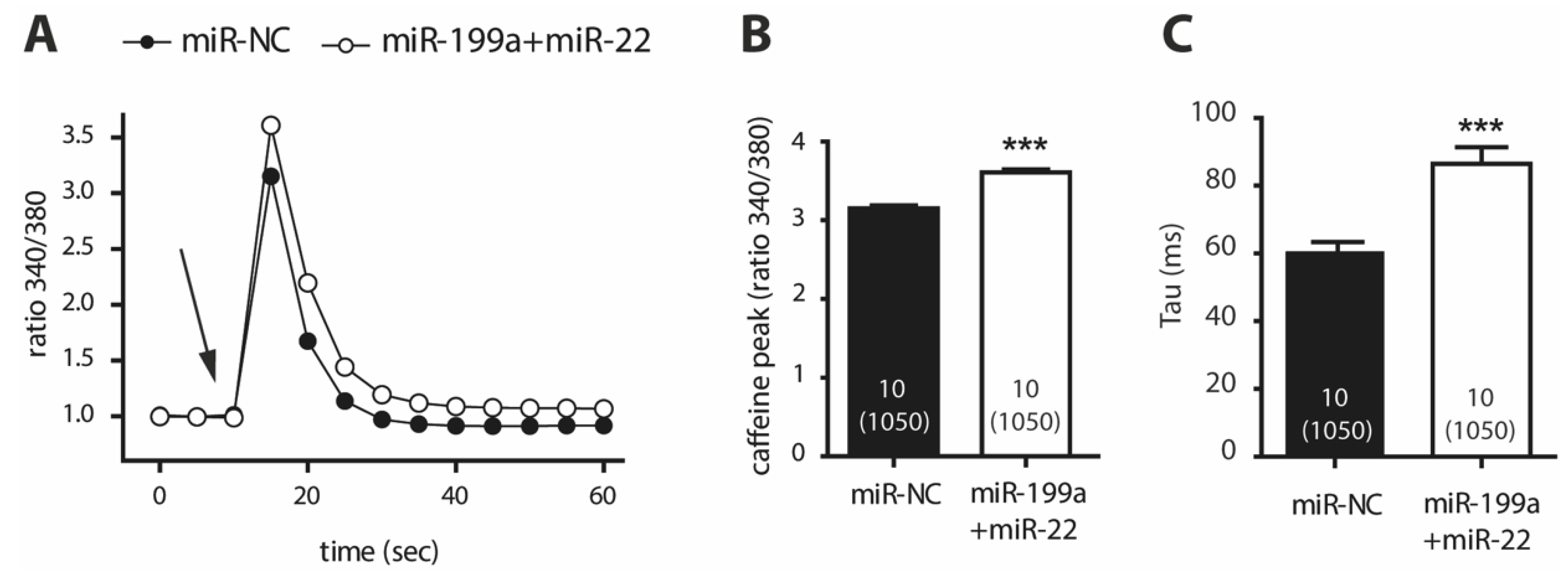
2.5. Cells Transfected with miR-199a-5p and miR-22-5p Have Higher Ca2+ Levels in the Sarcoplasmic Reticulum and a Slower Caffeine-Induced Ca2+ Decay
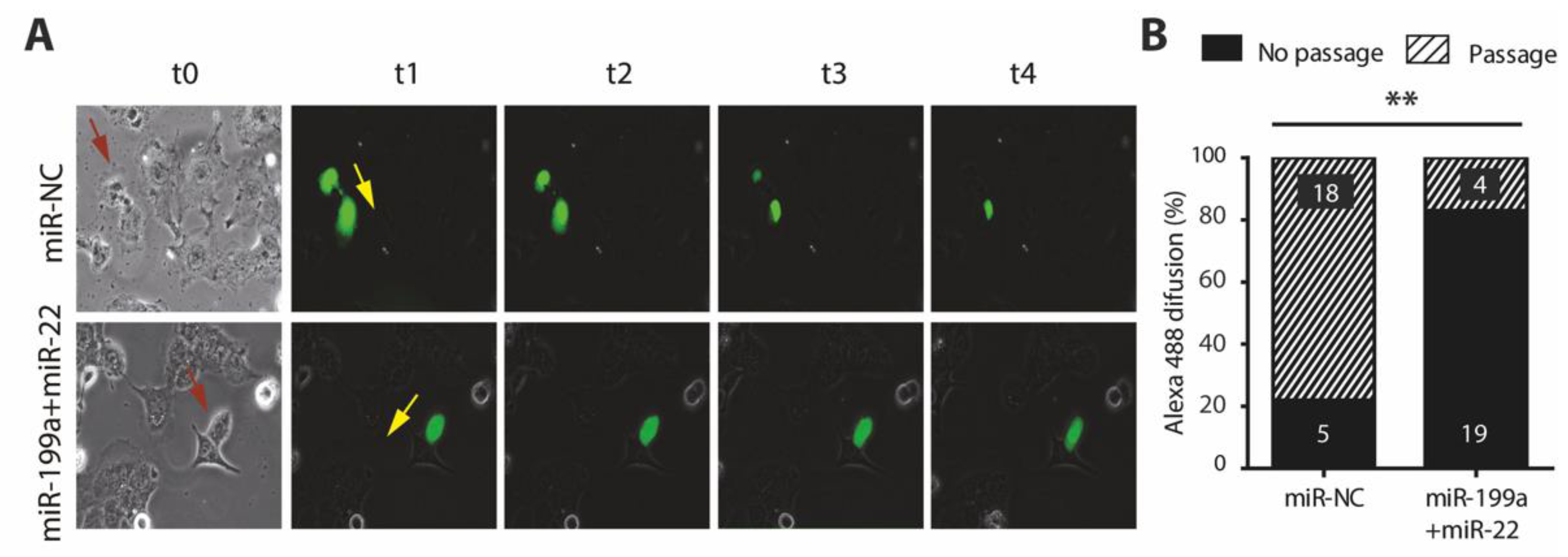
2.6. Overexpression of miR-199a-5p and miR-22-5p Is Associated with a Reduction in Cx40 and Impaired Cell-to-Cell Communication
3. Discussion
4. Materials and Methods
4.1. Human Plasma Study
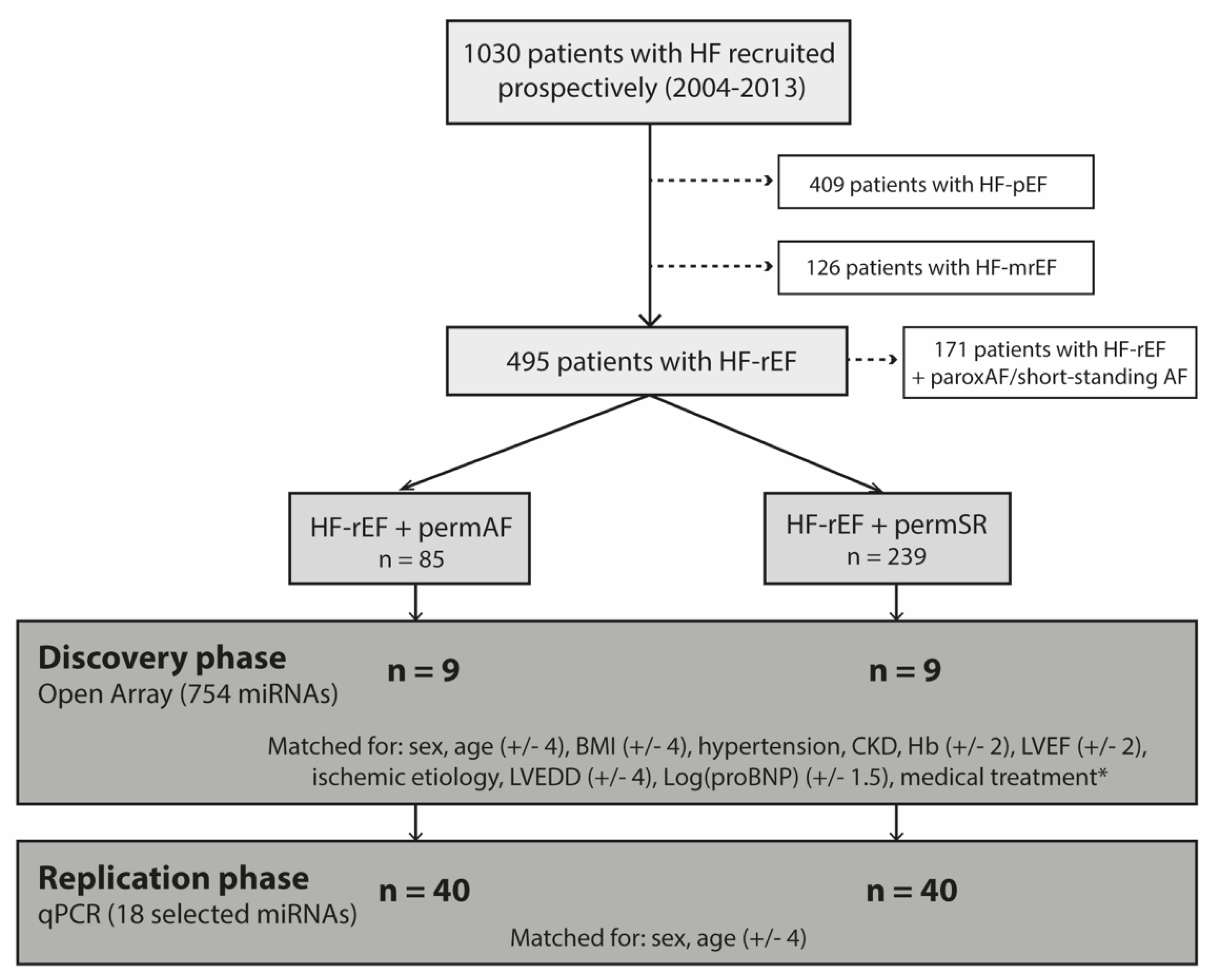
4.1.1. Study Population
4.1.2. Experimental Design
4.1.3. MicroRNA Extraction and Profiling
4.2. Human Atrial Samples
4.2.1. Cardiac Tissue Collection
4.2.2. MicroRNA Profiling
4.3. Molecular Studies
4.3.1. Cell Cultures and Transfection
4.3.2. qPCR and Western Blot Analyses
4.3.3. Recordings of Intracellular Ca2+ and Patch Clamp Studies
4.3.4. Gap-Junction Monitoring
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mosterd, A.; Hoes, A.W. Clinical epidemiology of heart failure. Heart 2007, 93, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linssen, G.C.; Rienstra, M.; Jaarsma, T.; Voors, A.A.; Van Gelder, I.C.; Hillege, H.L.; Van Veldhuisen, D.J. Clinical and prognostic effects of atrial fibrillation in heart failure patients with reduced and preserved left ventricular ejection fraction. Eur. J. Hear. Fail. 2011, 13, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.M.; Redfield, M.M.; Shen, W.K.; Gersh, B.J. Atrial fibrillation and ventricular dysfunction: A vicious electromechanical cycle. Circulation 2004, 109, 2839–2843. [Google Scholar] [CrossRef] [Green Version]
- Trulock, K.M.; Narayan, S.M.; Piccini, J.P. Rhythm control in heart failure patients with atrial fibrillation: Contemporary challenges including the role of ablation. J. Am. Coll. Cardiol. 2014, 64, 710–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, L.G.; Swedberg, K.; Ducharme, A.; Granger, C.B.; Michelson, E.L.; Mcmurray, J.; Puu, M.; Yusuf, S.; Pfeffer, M.A. Atrial Fibrillation and Risk of Clinical Events in Chronic Heart Failure With and Without Left Ventricular Systolic Dysfunction: Results From the Candesartan in Heart failure-Assessment of Reduction in Mortality and morbidity (CHARM) Program. J. Am. Coll. Cardiol. 2006, 47, 1997–2004. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.J.; Larson, M.G.; Levy, D.; Vasan, R.S.; Leip, E.P.; Wolf, P.A.; D’Agostino, R.B.; Murabito, J.M.; Kannel, W.B.; Benjamin, E.J. Temporal relations of atrial fibrillation and congestive heart failure and their joint influence on mortality: The Framingham Heart Study. Circulation 2003, 107, 2920–2925. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, U.M.; Jhund, P.; Abraham, W.T.; Desai, A.S.; Dickstein, K.; Packer, M.; Rouleau, J.L.; Solomon, S.D.; Swedberg, K.; Zile, M.; et al. Type of Atrial Fibrillation and Outcomes in Patients With Heart Failure and Reduced Ejection Fraction. J. Am. Coll. Cardiol. 2017, 70, 2490–2500. [Google Scholar] [CrossRef]
- Heijman, J.; Voigt, N.; Nattel, S.; Dobrev, D. Cellular and Molecular Electrophysiology of Atrial Fibrillation Initiation, Maintenance, and Progression. Circ. Res. 2014, 114, 1483–1499. [Google Scholar] [CrossRef] [Green Version]
- Sutanto, H.; Lyon, A.; Lumens, J.; Schotten, U.; Dobrev, D.; Heijman, J. Cardiomyocyte calcium handling in health and disease: Insights from in vitro and in silico studies. Prog. Biophys. Mol. Biol. 2020, 157, 54–75. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Duisters, R.F.; Tijsen, A.J.; Schroen, B.; Leenders, J.J.; Lentink, V.; van der Made, I.; Herias, V.; van Leeuwen, R.E.; Schellings, M.W.; Barenbrug, P.; et al. miR-133 and miR-30 regulate connective tissue growth factor: Implications for a role of microRNAs in myocardial matrix remodeling. Circ. Res. 2009, 104, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Dawson, K.; Wakili, R.; Ordög, B.; Clauss, S.; Chen, Y.; Iwasaki, Y.; Voigt, N.; Qi, X.Y.; Sinner, M.F.; Dobrev, D.; et al. MicroRNA29: A mechanistic contributor and potential biomarker in atrial fibrillation. Circulation 2013, 127, 1466–1475. [Google Scholar] [CrossRef] [Green Version]
- Reilly, S.N.; Liu, X.; Carnicer, R.; Recalde, A.; Muszkiewicz, A.; Jayaram, R.; Carena, M.C.; Wijesurendra, R.; Stefanini, M.; Surdo, N.C.; et al. Up-regulation of miR-31 in human atrial fibrillation begets the arrhythmia by depleting dystrophin and neuronal nitric oxide synthase. Sci. Transl. Med. 2016, 8, 340ra74. [Google Scholar] [CrossRef] [Green Version]
- Creemers, E.E.; Tijsen, A.J.; Pinto, Y.M. Circulating microRNAs: Novel biomarkers and extracellular communicators in cardiovascular disease? Circ. Res. 2012, 11, 483–495. [Google Scholar] [CrossRef]
- Wong, L.L.; Zou, R.; Zhou, L.; Lim, J.Y.; Phua, D.; Liu, C.; Chong, J.; Ng, J.; Liew, O.W.; Chan, S.P.; et al. Combining Circulating MicroRNA and NT-proBNP to Detect and Categorize Heart Failure Subtypes. J. Am. Coll. Cardiol. 2019, 73, 1300–1313. [Google Scholar] [CrossRef] [PubMed]
- Mun, D.; Kim, H.; Kang, J.; Park, H.; Park, H.; Lee, S.; Yun, N.; Joung, B. Expression of miRNAs in circulating exosomes derived from patients with persistent atrial fibrillation. FASEB J. 2019, 33, 5979–5989. [Google Scholar] [CrossRef] [Green Version]
- Goren, Y.; Meiri, E.; Hogan, C.; Mitchell, H.; Lebanony, D.; Salman, N.; Schliamser, J.E.; Amir, O. Relation of Reduced Expression of MiR-150 in Platelets to Atrial Fibrillation in Patients With Chronic Systolic Heart Failure. Am. J. Cardiol. 2014, 113, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.J.; Han, M.; Yang, F.Y.; Wei, G.C.; Liang, Z.G.; Yao, H.; Ji, C.W.; Xie, R.S.; Gong, C.L.; Tian, Y. Biological significance of miR-126 expression in atrial fibrillation and heart failure. Braz. J. Med. Biol. Res. 2015, 48, 983–989. [Google Scholar] [CrossRef]
- Gurha, P.; Wang, T.; Larimore, A.H.; Sassi, Y.; Abreu-Goodger, C.; Ramirez, M.O.; Reddy, A.K.; Engelhardt, S.; Taffet, G.E.; Wehrens, X.; et al. microRNA-22 Promotes Heart Failure through Coordinate Suppression of PPAR/ERR-Nuclear Hormone Receptor Transcription. PLoS ONE 2013, 8, e75882. [Google Scholar] [CrossRef] [PubMed]
- Boštjančič, E.; Zidar, N.; Glavač, D. MicroRNAs and cardiac sarcoplasmic reticulum calcium ATPase-2 in human myocardial infarction: Expression and bioinformatic analysis. BMC Genom. 2012, 13, 552. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Elias, A.; Alloza, L.; Puigdecanet, E.; Nonell, L.; Tajes, M.; Curado, J.; Enjuanes, C.; Diaz, O.; Bruguera, J.; Marti-Almor, J.; et al. Defining quantification methods and optimizing protocols for microarray hybridization of circulating microRNAs. Sci. Rep. 2017, 7, 7725. [Google Scholar] [CrossRef]
- Luo, X.; Yang, B.; Nattel, S. MicroRNAs and atrial fibrillation: Mechanisms and translational potential. Nat. Rev. Cardiol. 2014, 12, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Gurha, P.; Abreu-Goodger, C.; Wang, T.; Ramirez, M.O.; Drumond-Bock, A.L.; Van Dongen, S.; Chen, Y.; Bartonicek§, N.; Enright, A.; Lee, B.; et al. Targeted Deletion of MicroRNA-22 Promotes Stress-Induced Cardiac Dilation and Contractile Dysfunction. Circulation 2012, 125, 2751–2761. [Google Scholar] [CrossRef] [PubMed]
- Nishi, H.; Sakaguchi, T.; Miyagawa, S.; Yoshikawa, Y.; Fukushima, S.; Saito, S.; Ueno, T.; Kuratani, T.; Sawa, Y. Data from: Impact of microRNA expression in human atrial tissue in patients with atrial fibrillation undergoing cardiac surgery. PLoS One 2013, 8, e73397. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, J.; Seok, H.Y.; Zhang, Z.; Kataoka, M.; Hu, X.; Wang, D.-Z. MicroRNA-22 Regulates Cardiac Hypertrophy and Remodeling in Response to Stress. Circ. Res. 2013, 112, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Chiang, D.Y.; Zhang, M.; Voigt, N.; Alsina, K.M.; Jakob, H.; Martin, J.F.; Dobrev, D.; Wehrens, X.; Li, N. Identification of microRNA-mRNA dysregulations in paroxysmal atrial fibrillation. Int. J. Cardiol. 2015, 184, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Yamac, A.H.; Kucukbuzcu, S.; Ozansoy, M.; Gok, O.; Oz, K.; Erturk, M.; Yilmaz, E.; Ersoy, B.; Zeybek, R.; Goktekin, O.; et al. Altered expression of micro-RNA 199a and increased levels of cardiac SIRT1 protein are associated with the occurrence of atrial fibrillation after coronary artery bypass graft surgery. Cardiovasc. Pathol. 2016, 25, 232–236. [Google Scholar] [CrossRef]
- Aranda, J.F.; Rathjen, S.; Johannes, L.; Fernández-Hernando, C. MicroRNA 199a-5p Attenuates Retrograde Transport and Protects against Toxin-Induced Inhibition of Protein Biosynthesis. Mol. Cell. Biol. 2018, 38, e00548-17. [Google Scholar] [CrossRef] [Green Version]
- Aranda, J.F.; Canfrán-Duque, A.; Goedeke, L.; Suárez, Y.; Fernández-Hernando, C. The miR-199-dynamin regulatory axis controls receptor-mediated endocytosis. J Cell Sci. 2015, 128, 3197–3209. [Google Scholar] [CrossRef] [Green Version]
- Wakili, R.; Voigt, N.; Kääb, S.; Dobrev, D.; Nattel, S. Recent advances in the molecular pathophysiology of atrial fibrillation. J. Clin. Investig. 2011, 121, 2955–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voigt, N.; Li, N.; Wang, Q.; Wang, W.; Trafford, A.; Abu-Taha, I.; Sun, Q.; Wieland, T.; Ravens, U.; Nattel, S.; et al. Enhanced Sarcoplasmic Reticulum Ca 2+ Leak and Increased Na + -Ca2+ Exchanger Function Underlie Delayed Afterdepolarizations in Patients With Chronic Atrial Fibrillation. Circulation 2012, 125, 2059–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiser, M.; Kerfant, B.-G.; Williams, G.S.; Voigt, N.; Harks, E.; Dibb, K.; Giese, A.; Meszaros, J.; Verheule, S.; Ravens, U.; et al. Tachycardia-induced silencing of subcellular Ca2+ signaling in atrial myocytes. J. Clin. Investig. 2014, 124, 4759–4772. [Google Scholar] [CrossRef] [Green Version]
- Lugenbiel, P.; Wenz, F.; Govorov, K.; Schweizer, P.A.; Katus, H.A.; Thomas, D. Atrial fibrillation complicated by heart failure induces distinct remodeling of calcium cycling proteins. PLoS ONE 2015, 10, e0116395. [Google Scholar] [CrossRef]
- Ujihara, Y.; Iwasaki, K.; Takatsu, S.; Hashimoto, K.; Naruse, K.; Mohri, S.; Katanosaka, Y. Induced NCX1 overexpression attenuates pressure overload-induced pathological cardiac remodelling. Cardiovasc. Res. 2016, 111, 348–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firouzi, M.; Ramanna, H.; Kok, B.; Jongsma, H.J.; Koeleman, B.P.; Doevendans, P.A.; Groenewegen, W.A.; Hauer, R.N. Association of Human Connexin40 Gene Polymorphisms With Atrial Vulnerability as a Risk Factor for Idiopathic Atrial Fibrillation. Circ. Res. 2004, 95, e29–e33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gollob, M.H.; Jones, D.L.; Krahn, A.D.; Danis, L.; Gong, X.-Q.; Shao, Q.; Liu, X.; Veinot, J.P.; Tang, A.S.L.; Stewart, A.; et al. Somatic Mutations in the Connexin 40 Gene (GJA5) in Atrial Fibrillation. N. Engl. J. Med. 2006, 354, 2677–2688. [Google Scholar] [CrossRef]
- Chaldoupi, S.-M.; Loh, P.; Hauer, R.N.W.; De Bakker, J.M.T.; van Rijen, H. The role of connexin40 in atrial fibrillation. Cardiovasc. Res. 2009, 84, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Comin-Colet, J.; Enjuanes, C.; González, G.; Torrens, A.; Cladellas, M.; Meroño, O.; Ribas, N.; Ruiz, S.; Gomez, M.; Verdú, J.M.; et al. Iron deficiency is a key determinant of health-related quality of life in patients with chronic heart failure regardless of anaemia status. Eur. J. Hear. Fail. 2013, 15, 1164–1172. [Google Scholar] [CrossRef]
- Moliner, P.; Enjuanes, C.; Tajes, M.; Cainzos-Achirica, M.; Lupón, J.; Garay, A.; Jimenez-Marrero, S.; Yun, S.; Farré, N.; Cladellas, M.; et al. Association Between Norepinephrine Levels and Abnormal Iron Status in Patients With Chronic Heart Failure: Is Iron Deficiency More Than a Comorbidity? J. Am. Heart Assoc. 2019, 8, e010887. [Google Scholar] [CrossRef] [Green Version]
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.; Coats, A.; Falk, V.; González-Juanatey, J.R.; Harjola, V.P.; Jankowska, E.A.; et al. 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: The Task Force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC)Developed with the special contribution of the Heart Failure Association (HFA) of the ESC. Eur. Heart J. 2016, 37, 2129–2200. [Google Scholar] [CrossRef] [PubMed]
- Weber, P.A.; Chang, H.-C.; Spaeth, K.E.; Nitsche, J.M.; Nicholson, B. The Permeability of Gap Junction Channels to Probes of Different Size Is Dependent on Connexin Composition and Permeant-Pore Affinities. Biophys. J. 2004, 87, 958–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| permAF (n = 40) | SR (n = 40) | p-Value | |
|---|---|---|---|
| Patient Characteristics | |||
| Sex, men (%) | 31 (77.5) | 31 (77.5) | NS |
| Age, y (SD) | 73.2 (10.1) | 73.5 (10.3) | NS |
| BMI, kg/m2 (SD) | 30.5 (6.8) | 27.5 (3.7) | 0.02 |
| Medical history | |||
| Tobacco history (%) | 22 (55) | 22 (55) | NS |
| Hypertension (%) | 30 (75) | 31 (77.5) | NS |
| Diabetes (%) | 14 (35) | 18 (45) | NS |
| Hypercholesterolemia (%) | 20 (50) | 21 (52.5) | NS |
| COPD (%) | 6 (15) | 11 (27.5) | NS |
| CKD (%) | 11 (27.5) | 8 (20.0) | NS |
| Previous stroke (%) | 8 (20.0) | 4 (10.0) | NS |
| HF parameters | |||
| Ischemic etiology (%) | 16 (40) | 24 (60) | NS |
| Heart rate, bpm (SD) | 76.5 (16.6) | 70.0 (14.6) | NS |
| NYHA class (%): | NS | ||
| I-II | 16 (51.6) | 24 (75) | |
| III-IV | 15 (48.4) | 8 (25) | |
| LVEF, % (SD) | 32.5 (6.7) | 32.8 (6.5) | NS |
| Log Pro-BNP, mean (SD) | 3.4 (0.4) | 3.3 (0.6) | NS |
| HF treatment | |||
| ACEI/ARB (%) | 34 (85.0) | 33 (82.5) | NS |
| Betablockers (%) | 37 (92.5) | 37 (92.5) | NS |
| Diuretics (%) | 40 (100) | 37 (92.5) | NS |
| Heart rhythm | <0.001 | ||
| Permanent SR | 0 | 40 | |
| History of parox/pers AF * | 0 | 0 | |
| Permanent AF # | 40 | 0 |
| 2−△△Ct (permAF vs. SR) | p-Value | |
|---|---|---|
| miR-106a-5p | 1.292 | 0.378 |
| miR-106b-5p | 1.311 | 0.275 |
| miR-125a-5p | 1.395 | 0.110 |
| miR-126-5p | 1.244 | 0.258 |
| miR-133a-3p | 1.554 | 0.073 |
| miR-16-5p | 0.955 | 0.783 |
| miR-17-5p | 1.230 | 0.283 |
| miR-199a-5p | 1.921 | 0.028 * |
| miR-19a-3p | 1.526 | 0.117 |
| miR-20a-5p | 1.158 | 0.452 |
| miR-22-5p | 1.549 | 0.033 * |
| miR-23a-3p | 1.278 | 0.219 |
| miR-26a-5p | 1.322 | 0.187 |
| miR-27b-3p | 1.402 | 0.153 |
| miR-301a-3p | 1.115 | 0.765 |
| miR-324-5p | 1.531 | 0.088 |
| miR-374a-5p | 1.239 | 0.433 |
| miR-425-3p | 1.450 | 0.119 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Elias, A.; Tajes, M.; Yañez-Bisbe, L.; Enjuanes, C.; Comín-Colet, J.; Serra, S.A.; Fernández-Fernández, J.M.; Aguilar-Agon, K.W.; Reilly, S.; Martí-Almor, J.; et al. Atrial Fibrillation in Heart Failure Is Associated with High Levels of Circulating microRNA-199a-5p and 22–5p and a Defective Regulation of Intracellular Calcium and Cell-to-Cell Communication. Int. J. Mol. Sci. 2021, 22, 10377. https://doi.org/10.3390/ijms221910377
Garcia-Elias A, Tajes M, Yañez-Bisbe L, Enjuanes C, Comín-Colet J, Serra SA, Fernández-Fernández JM, Aguilar-Agon KW, Reilly S, Martí-Almor J, et al. Atrial Fibrillation in Heart Failure Is Associated with High Levels of Circulating microRNA-199a-5p and 22–5p and a Defective Regulation of Intracellular Calcium and Cell-to-Cell Communication. International Journal of Molecular Sciences. 2021; 22(19):10377. https://doi.org/10.3390/ijms221910377
Chicago/Turabian StyleGarcia-Elias, Anna, Marta Tajes, Laia Yañez-Bisbe, Cristina Enjuanes, Josep Comín-Colet, Selma A. Serra, José M. Fernández-Fernández, Kathryn W. Aguilar-Agon, Svetlana Reilly, Julio Martí-Almor, and et al. 2021. "Atrial Fibrillation in Heart Failure Is Associated with High Levels of Circulating microRNA-199a-5p and 22–5p and a Defective Regulation of Intracellular Calcium and Cell-to-Cell Communication" International Journal of Molecular Sciences 22, no. 19: 10377. https://doi.org/10.3390/ijms221910377