
Functional Changes of T-Cell Subsets with Age and CMV Infection

 ,
,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Results
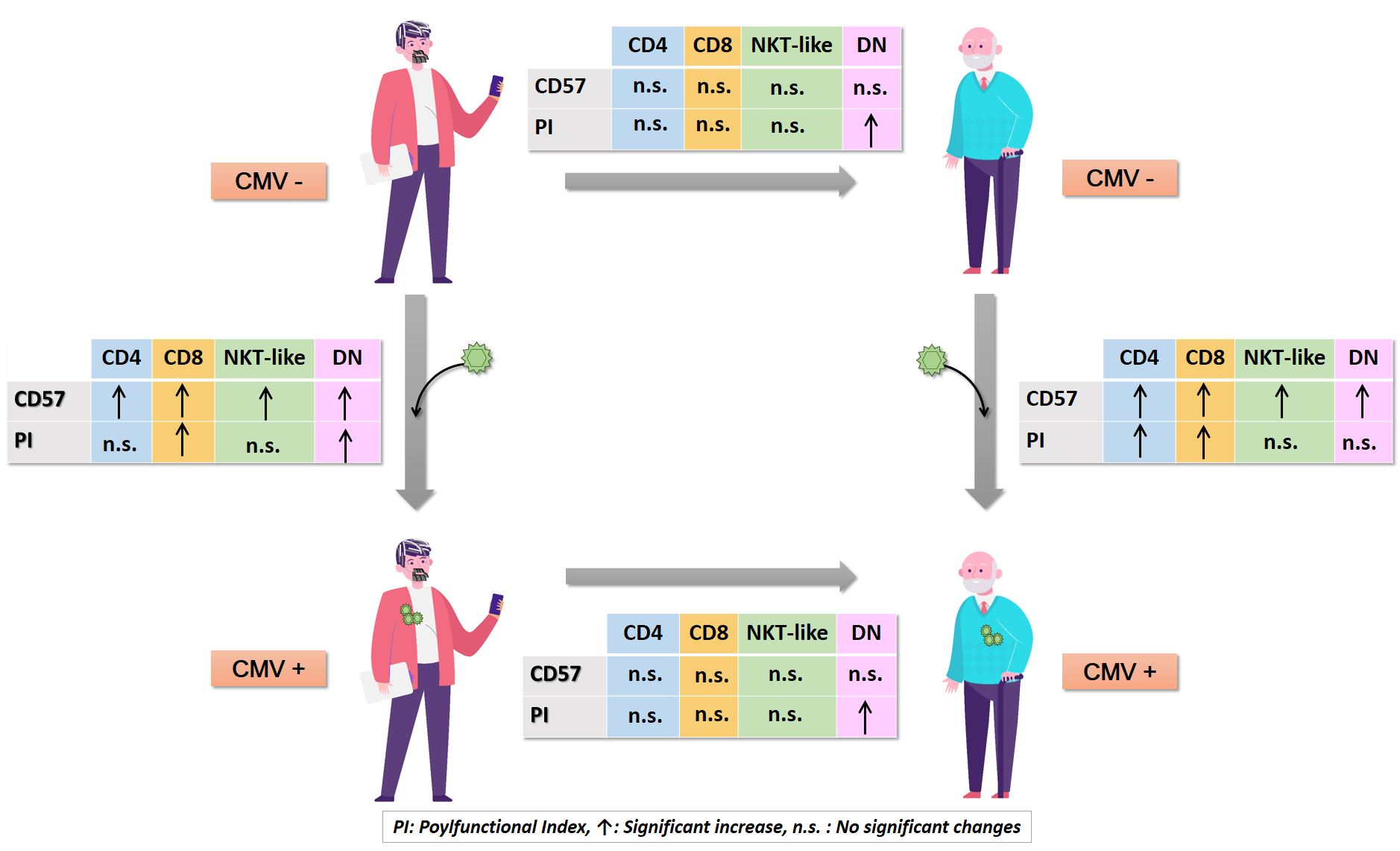
2.1. Effect of Age and CMV Infection on T-Cell Responses to SEB Stimulation
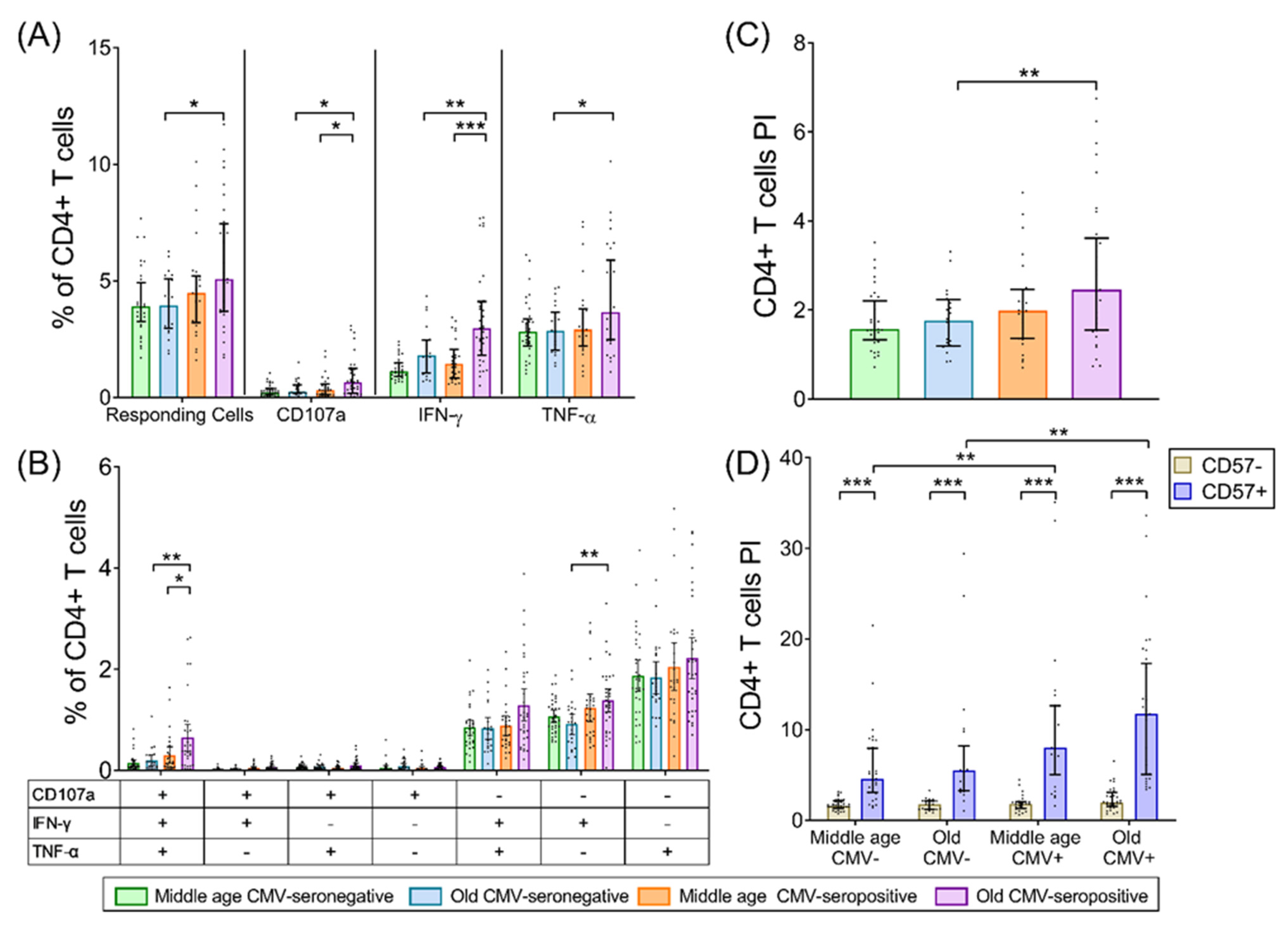
2.2. CD4+ T-Cells
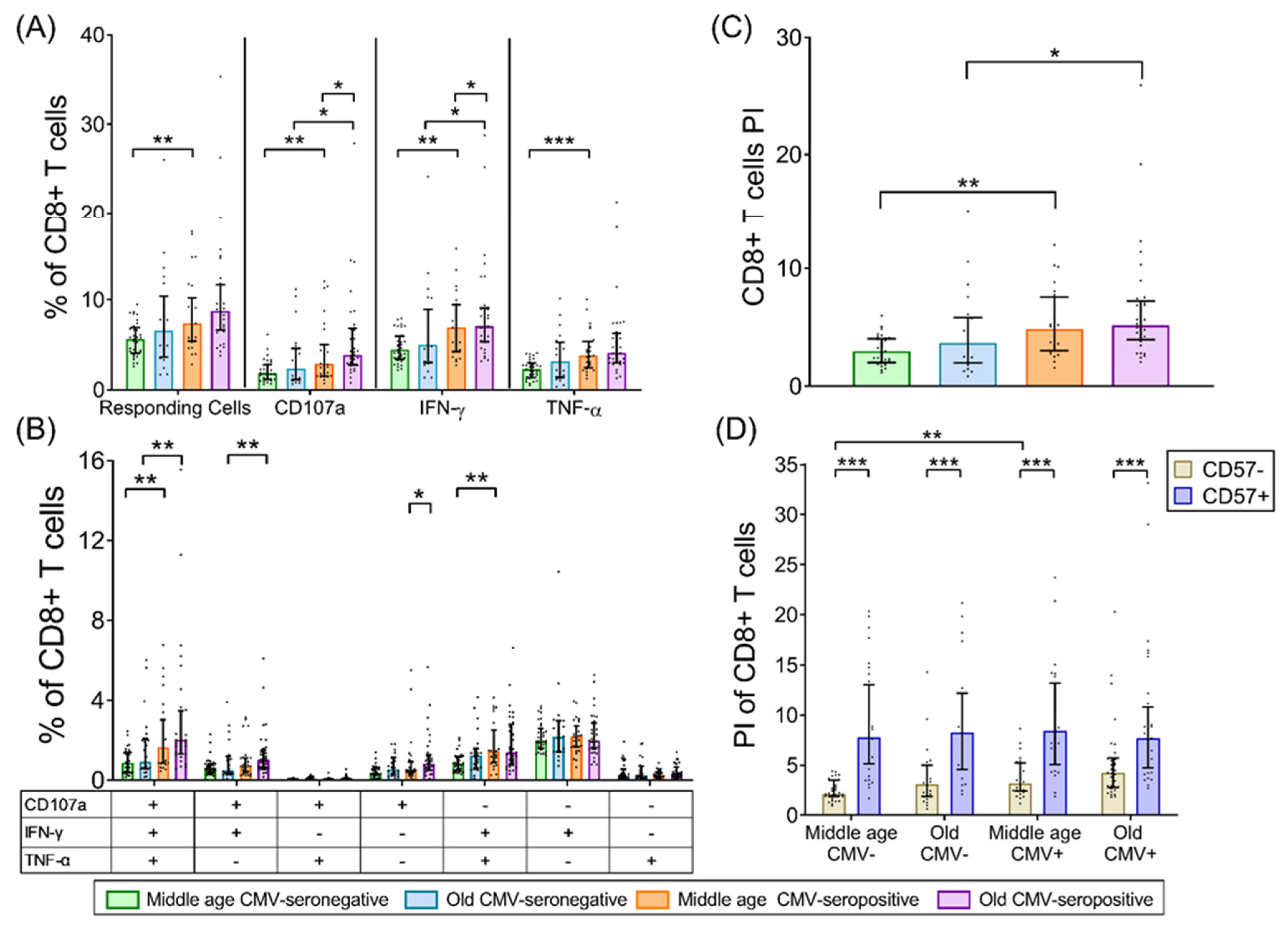
2.3. CD8+ T-Cells
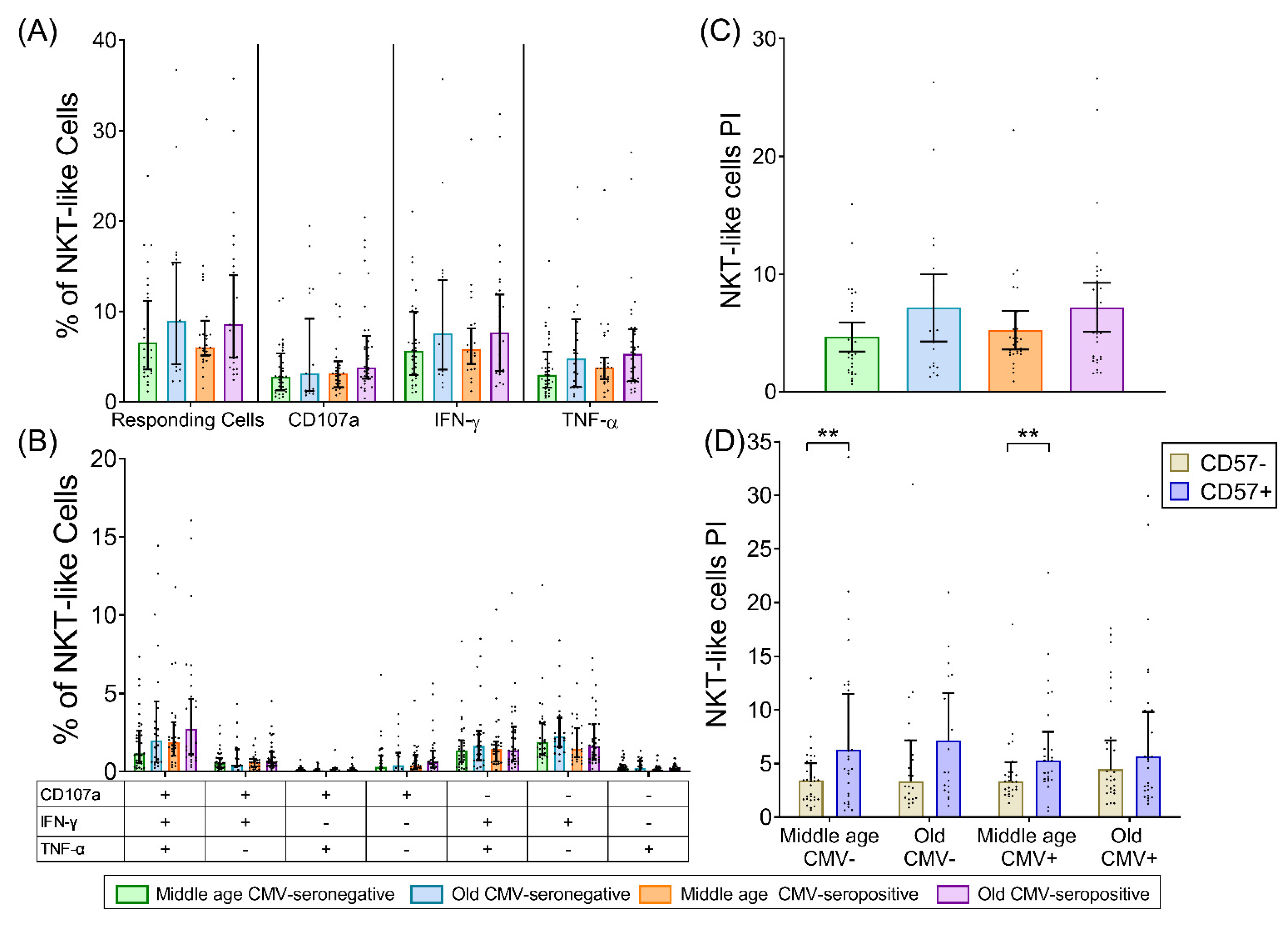
2.4. NKT-Like (CD8+CD56+) T-Cells
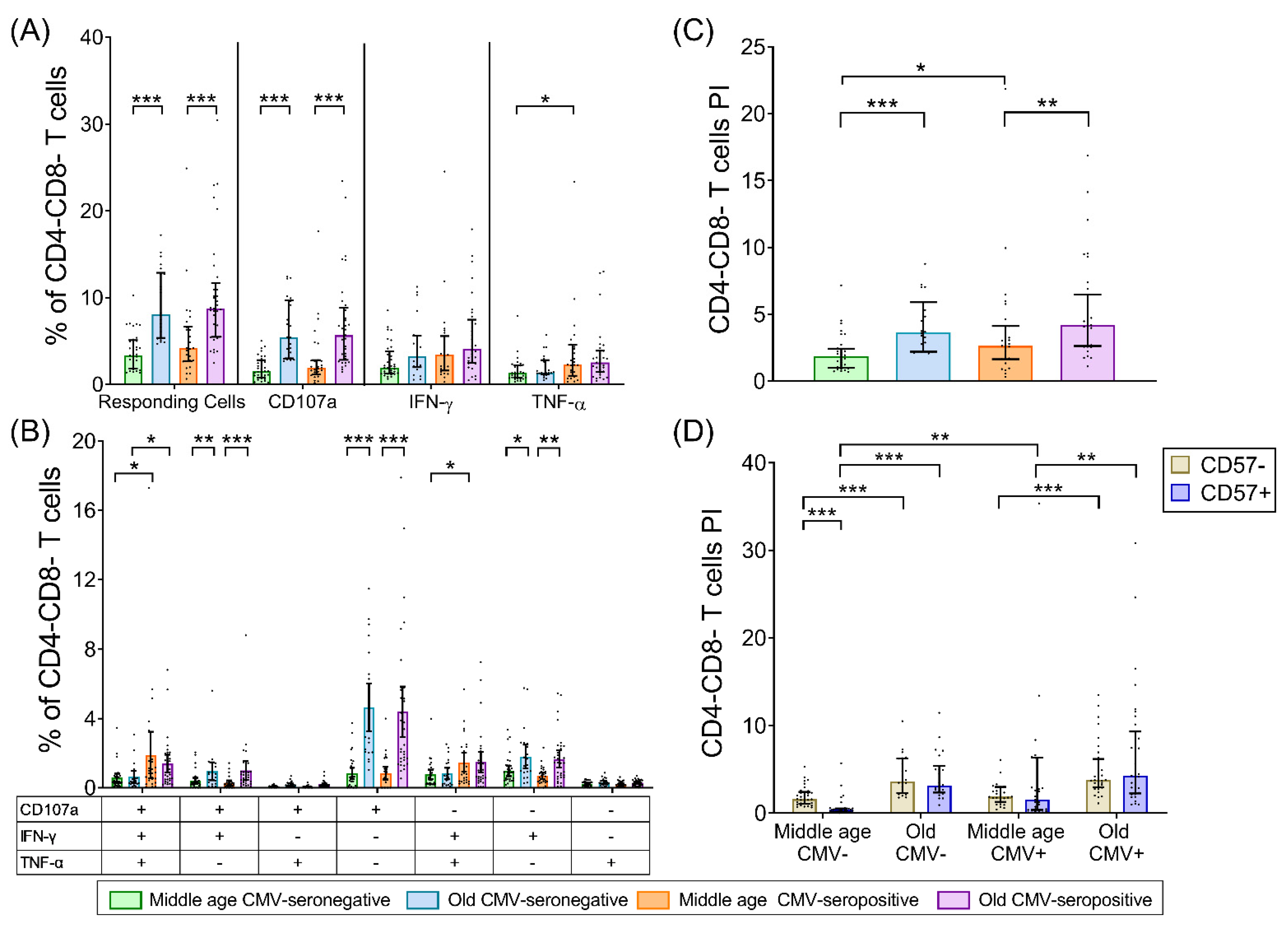
2.5. CD4-CD8- T-Cells (DN T-Cells)
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. CMV Serology
4.3. Stimulation, Intracellular Staining, and Detection of CD107a Expression
4.4. Flow Cytometry and Data Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonilla, F.A.; Oettgen, H.C. Adaptive immunity. J. Allergy Clin. Immunol. 2010, 125, S33–S40. [Google Scholar] [CrossRef]
- Farber, D.L.; Yudanin, N.A.; Restifo, N.P. Human memory T cells: Generation, compartmentalization and homeostasis. Nat. Rev. Immunol. 2014, 14, 24–35. [Google Scholar] [CrossRef]
- Pawelec, G.; Derhovanessian, E. Role of CMV in immune senescence. Virus Res. 2011, 157, 175–179. [Google Scholar] [CrossRef]
- Pawelec, G.; Derhovanessian, E.; Larbi, A.; Strindhall, J.; Wikby, A. Cytomegalovirus and human immunosenescence. Rev. Med. Virol. 2009, 19, 47–56. [Google Scholar] [CrossRef]
- Miller, R.A. The aging immune system: Primer and prospectus. Science 1996, 273, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Cambier, J. Immunosenescence: A problem of lymphopoiesis, homeostasis, microenvironment, and signaling. Immunol. Rev. 2005, 205, 5–6. [Google Scholar] [CrossRef]
- Wertheimer, A.M.; Bennett, M.S.; Park, B.; Uhrlaub, J.L.; Martinez, C.; Pulko, V.; Currier, N.L.; Nikolich-Žugich, D.; Kaye, J.; Nikolich-Žugich, J. Aging and cytomegalovirus infection differentially and jointly affect distinct circulating T cell subsets in humans. J. Immunol. 2014, 192, 2143–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, S.; Solana, R.; Dela Rosa, O.; Pawelec, G. Human cytomegalovirus infection and T cell immunosenescence: A mini review. Mech. Ageing Dev. 2006, 127, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Larbi, A.; Ozcelik, D.; Solana, R.; Gouttefangeas, C.; Attig, S.; Wikby, A.; Strindhall, J.; Franceschi, C.; Pawelec, G. Cytomegalovirus infection: A driving force in human T cell immunosenescence. Ann. N. Y. Acad. Sci. 2007, 1114, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Aiello, A.; Accardi, G.; Candore, G.; Caruso, C.; Colomba, C.; Di Bona, D.; Duro, G.; Gambino, C.M.; Ligotti, M.E.; Pandey, J.P. Role of Immunogenetics in the Outcome of HCMV Infection: Implications for Ageing. Int. J. Mol. Sci. 2019, 20, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Shariff, N.; Cobbold, M.; Bruton, R.; Ainsworth, J.A.; Sinclair, A.J.; Nayak, L.; Moss, P.A. Cytomegalovirus seropositivity drives the CD8 T cell repertoire toward greater clonality in healthy elderly individuals. J. Immunol. 2002, 169, 1984–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Q.; Wagner, W.M.; Zheng, W.; Wikby, A.; Remarque, E.J.; Pawelec, G. Dysfunctional CMV-specific CD8(+) T cells accumulate in the elderly. Exp. Gerontol. 2004, 39, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; McElhaney, J.E.; Aiello, A.E.; Derhovanessian, E. The impact of CMV infection on survival in older humans. Curr. Opin. Immunol. 2012, 24, 507–511. [Google Scholar] [CrossRef]
- Simanek, A.M.; Dowd, J.B.; Pawelec, G.; Melzer, D.; Dutta, A.; Aiello, A.E. Seropositivity to cytomegalovirus, inflammation, all-cause and cardiovascular disease-related mortality in the United States. PLoS ONE 2011, 6, e16103. [Google Scholar] [CrossRef]
- Olsson, J.; Wikby, A.; Johansson, B.; Lofgren, S.; Nilsson, B.O.; Ferguson, F.G. Age-related change in peripheral blood T-lymphocyte subpopulations and cytomegalovirus infection in the very old: The Swedish longitudinal OCTO immune study. Mech. Ageing Dev. 2000, 121, 187–201. [Google Scholar] [CrossRef]
- Ferrando-Martinez, S.; Romero-Sanchez, M.C.; Solana, R.; Delgado, J.; de la Rosa, R.; Munoz-Fernandez, M.A.; Ruiz-Mateos, E.; Leal, M. Thymic function failure and C-reactive protein levels are independent predictors of all-cause mortality in healthy elderly humans. Age 2013, 35, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Weng, N.P.; Akbar, A.N.; Goronzy, J. CD28(-) T cells: Their role in the age-associated decline of immune function. Trends Immunol. 2009, 30, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.P.; Effros, R.B. T cell replicative senescence in human aging. Curr. Pharm. Des. 2013, 19, 1680–1698. [Google Scholar] [PubMed]
- Strioga, M.; Pasukoniene, V.; Characiejus, D. CD8+ CD28- and CD8+ CD57+ T cells and their role in health and disease. Immunology 2011, 134, 17–32. [Google Scholar] [CrossRef]
- Furman, D.; Jojic, V.; Sharma, S.; Shen-Orr, S.S.; Angel, C.J.; Onengut-Gumuscu, S.; Kidd, B.A.; Maecker, H.T.; Concannon, P.; Dekker, C.L.; et al. Cytomegalovirus infection enhances the immune response to influenza. Sci. Transl. Med. 2015, 7, 281. [Google Scholar] [CrossRef] [Green Version]
- Barton, E.S.; White, D.W.; Cathelyn, J.S.; Brett-McClellan, K.A.; Engle, M.; Diamond, M.S.; Miller, V.L.; Virgin, H.W.t. Herpesvirus latency confers symbiotic protection from bacterial infection. Nature 2007, 447, 326–329. [Google Scholar] [CrossRef]
- Miles, D.J.; Sanneh, M.; Holder, B.; Crozier, S.; Nyamweya, S.; Touray, E.S.; Palmero, M.S.; Zaman, S.M.; Rowland-Jones, S.; van der Sande, M.; et al. Cytomegalovirus infection induces T-cell differentiation without impairing antigen-specific responses in Gambian infants. Immunology 2008, 124, 388–400. [Google Scholar] [CrossRef]
- Pera, A.; Vasudev, A.; Tan, C.; Kared, H.; Solana, R.; Larbi, A. CMV induces expansion of highly polyfunctional CD4+ T cell subset coexpressing CD57 and CD154. J. Leukoc. Biol. 2017, 101, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Pera, A.; Campos, C.; Corona, A.; Sanchez-Correa, B.; Tarazona, R.; Larbi, A.; Solana, R. CMV latent infection improves CD8+ T response to SEB due to expansion of polyfunctional CD57+ cells in young individuals. PLoS ONE 2014, 9, e88538. [Google Scholar] [CrossRef] [Green Version]
- Hassouneh, F.; Campos, C.; López-Sejas, N.; Alonso, C.; Tarazona, R.; Solana, R.; Pera, A. Effect of age and latent CMV infection on CD8+ CD56+ T cells (NKT-like) frequency and functionality. Mech. Ageing Dev. 2016, 158, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.; Almeida, J.R.; Darrah, P.A.; Sauce, D.; Seder, R.A.; Appay, V.; Gorochov, G.; Larsen, M. Pathogen-Specific T Cell Polyfunctionality Is a Correlate of T Cell Efficacy and Immune Protection. PLoS ONE 2015, 10, e0128714. [Google Scholar] [CrossRef]
- White, D.W.; Suzanne Beard, R.; Barton, E.S. Immune modulation during latent herpesvirus infection. Immunol. Rev. 2012, 245, 189–208. [Google Scholar] [CrossRef] [Green Version]
- Fleischer, B. Superantigens. APMIS 1994, 102, 3–12. [Google Scholar] [CrossRef]
- Fleischer, B.; Schrezenmeier, H. T cell stimulation by staphylococcal enterotoxins. Clonally variable response and requirement for major histocompatibility complex class II molecules on accessory or target cells. J. Exp. Med. 1988, 167, 1697–1707. [Google Scholar] [CrossRef]
- Whitfield, S.J.C.; Taylor, C.; Risdall, J.E.; Griffiths, G.D.; Jones, J.T.A.; Williamson, E.D.; Rijpkema, S.; Saraiva, L.; Vessillier, S.; Green, A.C.; et al. Interference of the T Cell and Antigen-Presenting Cell Costimulatory Pathway Using CTLA4-Ig (Abatacept) Prevents Staphylococcal Enterotoxin B Pathology. J. Immunol. 2017, 198, 3989–3998. [Google Scholar] [CrossRef]
- Appay, V.; van Lier, R.A.; Sallusto, F.; Roederer, M. Phenotype and function of human T lymphocyte subsets: Consensus and issues. Cytom. A 2008, 73, 975–983. [Google Scholar] [CrossRef]
- Koch, S.; Larbi, A.; Derhovanessian, E.; Ozcelik, D.; Naumova, E.; Pawelec, G. Multiparameter flow cytometric analysis of CD4 and CD8 T cell subsets in young and old people. Immun. Ageing 2008, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Elwenspoek, M.M.C.; Sias, K.; Hengesch, X.; Schaan, V.K.; Leenen, F.A.D.; Adams, P.; Meriaux, S.B.; Schmitz, S.; Bonnemberger, F.; Ewen, A.; et al. T Cell Immunosenescence after Early Life Adversity: Association with Cytomegalovirus Infection. Front. Immunol. 2017, 8, 1263. [Google Scholar] [CrossRef] [Green Version]
- Focosi, D.; Bestagno, M.; Burrone, O.; Petrini, M. CD57+ T lymphocytes and functional immune deficiency. J. Leukoc. Biol. 2010, 87, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Broadley, I.; Pera, A.; Morrow, G.; Davies, K.A.; Kern, F. Expansions of Cytotoxic CD4. Front. Immunol. 2017, 8, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pera, A.; Broadley, I.; Davies, K.A.; Kern, F. Cytomegalovirus as a Driver of Excess Cardiovascular Mortality in Rheumatoid Arthritis: A Red Herring or a Smoking Gun? Circ. Res. 2017, 120, 274–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassouneh, F.; Lopez-Sejas, N.; Campos, C.; Sanchez-Correa, B.; Tarazona, R.; Solana, R.; Pera, A. Differential Effect of Cytomegalovirus Infection with Age on the Expression of CD57, CD300a, and CD161 on T-Cell Subpopulations. Front. Immunol. 2017, 8, 649. [Google Scholar] [CrossRef] [Green Version]
- Pera, A.; Caserta, S.; Albanese, F.; Blowers, P.; Morrow, G.; Terrazzini, N.; Smith, H.E.; Rajkumar, C.; Reus, B.; Msonda, J.R.; et al. CD28. Theranostics 2018, 8, 4509–4519. [Google Scholar] [CrossRef]
- Youn, J.C.; Jung, M.K.; Yu, H.T.; Kwon, J.S.; Kwak, J.E.; Park, S.H.; Kim, I.C.; Park, M.S.; Lee, S.K.; Choi, S.W.; et al. Increased frequency of CD4. Sci. Rep. 2019, 9, 12887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okba, A.M.; Abd El Raouf Raafat, M.; Nazmy Farres, M.; Abd El Nour Melek, N.; Amin, M.M.; Gendy, N.N. Expanded peripheral CD4. Hum. Immunol. 2019, 80, 748–754. [Google Scholar] [CrossRef]
- Peeters, L.M.; Vanheusden, M.; Somers, V.; Van Wijmeersch, B.; Stinissen, P.; Broux, B.; Hellings, N. Cytotoxic CD4+ T Cells Drive Multiple Sclerosis Progression. Front. Immunol. 2017, 8, 1160. [Google Scholar] [CrossRef] [Green Version]
- Bano, A.; Pera, A.; Almoukayed, A.; Clarke, T.H.S.; Kirmani, S.; Davies, K.A.; Kern, F. CD28. F1000Research 2019, 8, 179. [Google Scholar] [CrossRef]
- Chanouzas, D.; Sagmeister, M.; Faustini, S.; Nightingale, P.; Richter, A.; Ferro, C.J.; Morgan, M.D.; Moss, P.; Harper, L. Subclinical Reactivation of Cytomegalovirus Drives CD4+CD28null T-Cell Expansion and Impaired Immune Response to Pneumococcal Vaccination in Antineutrophil Cytoplasmic Antibody-Associated Vasculitis. J. Infect. Dis. 2019, 219, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Cobo, B.; Solano, C.; Benet, I.; Costa, E.; Remigia, M.J.; de la Cámara, R.; Nieto, J.; López, J.; Amat, P.; Garcia-Noblejas, A.; et al. Functional profile of cytomegalovirus (CMV)-specific CD8+ T cells and kinetics of NKG2C+ NK cells associated with the resolution of CMV DNAemia in allogeneic stem cell transplant recipients. J. Med. Virol. 2012, 84, 259–267. [Google Scholar] [CrossRef]
- Chan, K.S.; Kaur, A. Flow cytometric detection of degranulation reveals phenotypic heterogeneity of degranulating CMV-specific CD8+ T lymphocytes in rhesus macaques. J. Immunol. Methods 2007, 325, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, M.C.; Manunta, M.D.I.; Pincott, E.S.; Fenton, M.; Simpson, G.L.; Klein, N.J.; Burch, M. Specific Immunity to Cytomegalovirus in Pediatric Cardiac Transplantation. Transplantation 2018, 102, 1569–1575. [Google Scholar] [CrossRef]
- Makwana, N.; Foley, B.; Fernandez, S.; Lee, S.; Irish, A.; Pircher, H.; Price, P. CMV drives the expansion of highly functional memory T cells expressing NK-cell receptors in renal transplant recipients. Eur. J. Immunol. 2017, 47, 1324–1334. [Google Scholar] [CrossRef]
- Cantisán, S.; Páez-Vega, A.; Santos, F.; Rodríguez-Benot, A.; Aguado, R.; Rivero, A.; Montejo, M.; Torre-Cisneros, J.; Solana, R.; (REIPI), S.N.f.R.i.I.D. Impact of age and cytomegalovirus on CD8. Exp. Gerontol. 2017, 95, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.; Mourin, G.; Larsen, M.; Fastenackels, S.; Urrutia, A.; Gorochov, G.; Autran, B.; Donner, C.; Sidi, D.; Sibony-Prat, J.; et al. Differential impact of age and cytomegalovirus infection on the γδ T cell compartment. J. Immunol. 2013, 191, 1300–1306. [Google Scholar] [CrossRef]
- Vasudev, A.; Ying, C.T.; Ayyadhury, S.; Puan, K.J.; Andiappan, A.K.; Nyunt, M.S.; Shadan, N.B.; Mustafa, S.; Low, I.; Rotzschke, O.; et al. γ/δ T cell subsets in human aging using the classical α/β T cell model. J. Leukoc. Biol. 2014, 96, 647–655. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.T.; Wistuba-Hamprecht, K.; Xu, W.; Nyunt, M.S.; Vasudev, A.; Lee, B.T.; Pawelec, G.; Puan, K.J.; Rotzschke, O.; Ng, T.P.; et al. Vδ2+ and α/ß T cells show divergent trajectories during human aging. Oncotarget 2016, 7, 44906–44918. [Google Scholar] [CrossRef] [PubMed]
- van der Geest, K.S.M.; Kroesen, B.J.; Horst, G.; Abdulahad, W.H.; Brouwer, E.; Boots, A.M.H. Impact of Aging on the Frequency, Phenotype, and Function of CD161-Expressing T Cells. Front. Immunol. 2018, 9, 752. [Google Scholar] [CrossRef] [Green Version]
- Mocchegiani, E.; Giacconi, R.; Cipriano, C.; Malavolta, M. NK and NKT cells in aging and longevity: Role of zinc and metallothioneins. J. Clin. Immunol. 2009, 29, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Almehmadi, M.; Flanagan, B.F.; Khan, N.; Alomar, S.; Christmas, S.E. Increased numbers and functional activity of CD56⁺ T cells in healthy cytomegalovirus positive subjects. Immunology 2014, 142, 258–268. [Google Scholar] [CrossRef]
- Sadat-Sowti, B.; Debre, P.; Mollet, L.; Quint, L.; Hadida, F.; Leblond, V.; Bismuth, G.; Autran, B. An inhibitor of cytotoxic functions produced by CD8+CD57+ T lymphocytes from patients suffering from AIDS and immunosuppressed bone marrow recipients. Eur. J. Immunol. 1994, 24, 2882–2888. [Google Scholar] [CrossRef]
- Frassanito, M.A.; Silvestris, F.; Cafforio, P.; Dammacco, F. CD8+/CD57 cells and apoptosis suppress T-cell functions in multiple myeloma. Br. J. Haematol. 1998, 100, 469–477. [Google Scholar] [CrossRef]
- Derhovanessian, E.; Theeten, H.; Hähnel, K.; Van Damme, P.; Cools, N.; Pawelec, G. Cytomegalovirus-associated accumulation of late-differentiated CD4 T-cells correlates with poor humoral response to influenza vaccination. Vaccine 2013, 31, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Goldeck, D.; Theeten, H.; Hassouneh, F.; Oettinger, L.; Wistuba-Hamprecht, K.; Cools, N.; Tsitsilonis, O.E.; Pawelec, G. Frequencies of peripheral immune cells in older adults following seasonal influenza vaccination with an adjuvanted vaccine. Vaccine 2017, 35, 4330–4338. [Google Scholar] [CrossRef]
- Derhovanessian, E.; Maier, A.B.; Hähnel, K.; McElhaney, J.E.; Slagboom, E.P.; Pawelec, G. Latent infection with cytomegalovirus is associated with poor memory CD4 responses to influenza A core proteins in the elderly. J. Immunol. 2014, 193, 3624–3631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Berg, S.P.H.; Lanfermeijer, J.; Jacobi, R.H.J.; Hendriks, M.; Vos, M.; van Schuijlenburg, R.; Nanlohy, N.M.; Borghans, J.A.M.; van Beek, J.; van Baarle, D.; et al. Latent CMV Infection Is Associated with Lower Influenza Virus-Specific Memory T-Cell Frequencies, but Not With an Impaired T-Cell Response to Acute Influenza Virus Infection. Front. Immunol. 2021, 12, 663664. [Google Scholar] [CrossRef]
- Higdon, L.E.; Gustafson, C.E.; Ji, X.; Sahoo, M.K.; Pinsky, B.A.; Margulies, K.B.; Maecker, H.T.; Goronzy, J.; Maltzman, J.S. Association of Premature Immune Aging and Cytomegalovirus After Solid Organ Transplant. Front. Immunol. 2021, 12, 661551. [Google Scholar] [CrossRef] [PubMed]
- Royston, L.; Isnard, S.; Lin, J.; Routy, J.P. Cytomegalovirus as an Uninvited Guest in the Response to Vaccines in People Living with HIV. Viruses 2021, 13, 1266. [Google Scholar] [CrossRef] [PubMed]
- Westendorp, R.G.; van Heemst, D.; Rozing, M.P.; Frolich, M.; Mooijaart, S.P.; Blauw, G.J.; Beekman, M.; Heijmans, B.T.; de Craen, A.J.; Slagboom, P.E.; et al. Nonagenarian siblings and their offspring display lower risk of mortality and morbidity than sporadic nonagenarians: The Leiden Longevity Study. J. Am. Geriatr. Soc. 2009, 57, 1634–1637. [Google Scholar] [CrossRef] [PubMed]
- Schoenmaker, M.; de Craen, A.J.; de Meijer, P.H.; Beekman, M.; Blauw, G.J.; Slagboom, P.E.; Westendorp, R.G. Evidence of genetic enrichment for exceptional survival using a family approach: The Leiden Longevity Study. Eur. J. Hum. Genet. 2006, 14, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, L.H.; Maier, A.B.; Slagbom, P.E.; Pawelec, G.; Derhovanessian, E.; Petersen, I.; Jahn, G.; Westendorp, R.G.; Christensen, K. Early-life environment influencing susceptibility to cytomegalovirus infection: Evidence from the Leiden Longevity Study and the Longitudinal Study of Aging Danish Twins. Epidemiol. Infect. 2012, 140, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Herrero, S.; Sordo-Bahamonde, C.; Gonzalez, S.; López-Soto, A. CD107a Degranulation Assay to Evaluate Immune Cell Antitumor Activity. Methods Mol. Biol. 2019, 1884, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.; Sauce, D.; Arnaud, L.; Fastenackels, S.; Appay, V.; Gorochov, G. Evaluating cellular polyfunctionality with a novel polyfunctionality index. PLoS ONE 2012, 7, e42403. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CMV | Age (Mean ± SD) | Sex (Male/Female) | No. | Group Name |
|---|---|---|---|---|
| Negative | 45–64 (60 ± 3) | 13/21 | 34 | Middle-aged CMV-seronegative |
| Positive | 46–63 (60 ± 3) | 10/18 | 28 | Middle-aged CMV-seropositive |
| Negative | 65–78 (69 ± 3) | 16/8 | 24 | Older CMV-seronegative |
| Positive | 65–73 (68 ± 2) | 18/15 | 33 | Older CMV-seropositive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassouneh, F.; Goldeck, D.; Pera, A.; van Heemst, D.; Slagboom, P.E.; Pawelec, G.; Solana, R. Functional Changes of T-Cell Subsets with Age and CMV Infection. Int. J. Mol. Sci. 2021, 22, 9973. https://doi.org/10.3390/ijms22189973
Hassouneh F, Goldeck D, Pera A, van Heemst D, Slagboom PE, Pawelec G, Solana R. Functional Changes of T-Cell Subsets with Age and CMV Infection. International Journal of Molecular Sciences. 2021; 22(18):9973. https://doi.org/10.3390/ijms22189973
Chicago/Turabian StyleHassouneh, Fakhri, David Goldeck, Alejandra Pera, Diana van Heemst, P. Eline Slagboom, Graham Pawelec, and Rafael Solana. 2021. "Functional Changes of T-Cell Subsets with Age and CMV Infection" International Journal of Molecular Sciences 22, no. 18: 9973. https://doi.org/10.3390/ijms22189973