
Wound Healing Effect of Gintonin Involves Lysophosphatidic Acid Receptor/Vascular Endothelial Growth Factor Signaling Pathway in Keratinocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
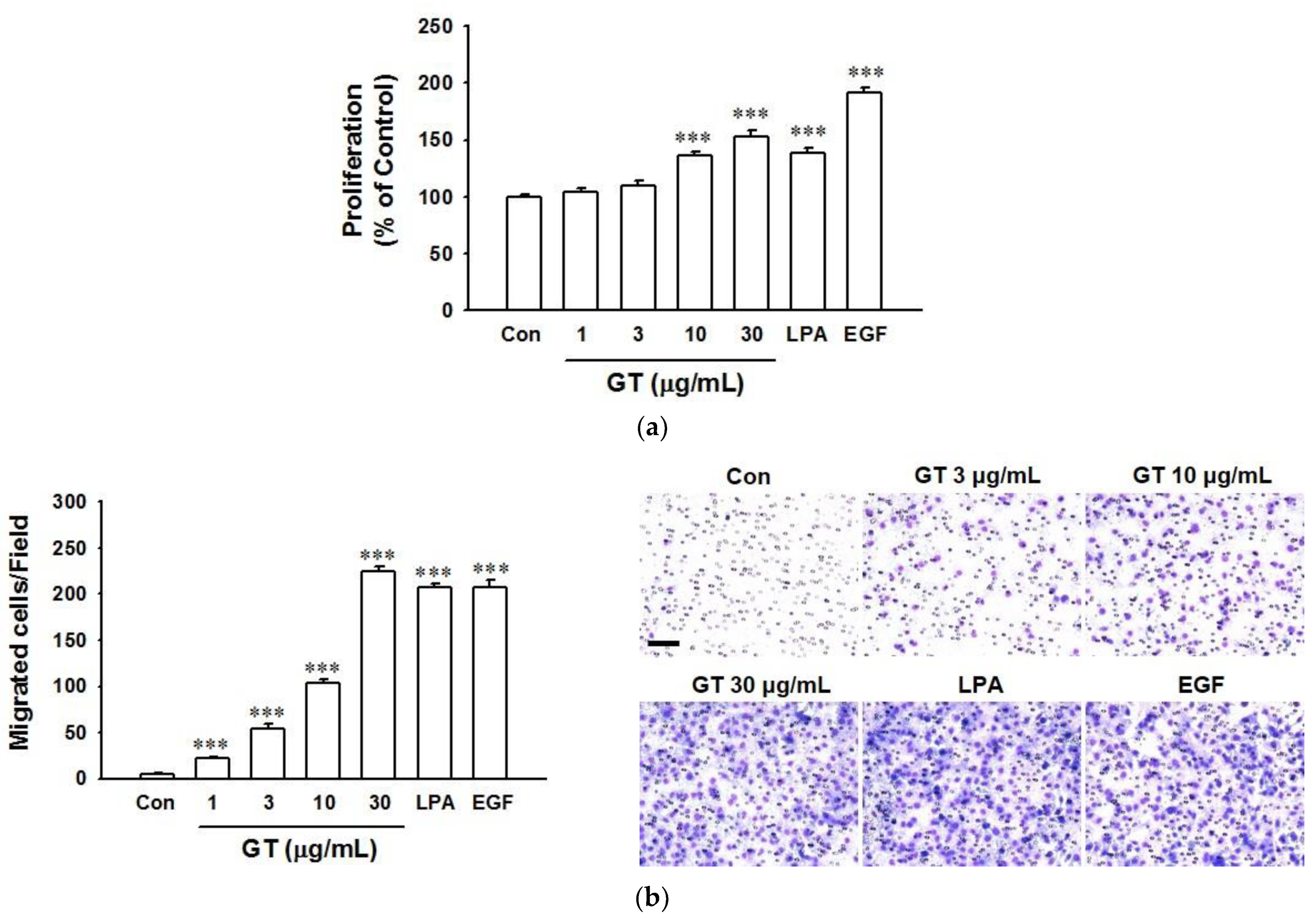
2.1. Gintonin-Mediated Enhancement of HaCaT Cell Proliferation and Migration
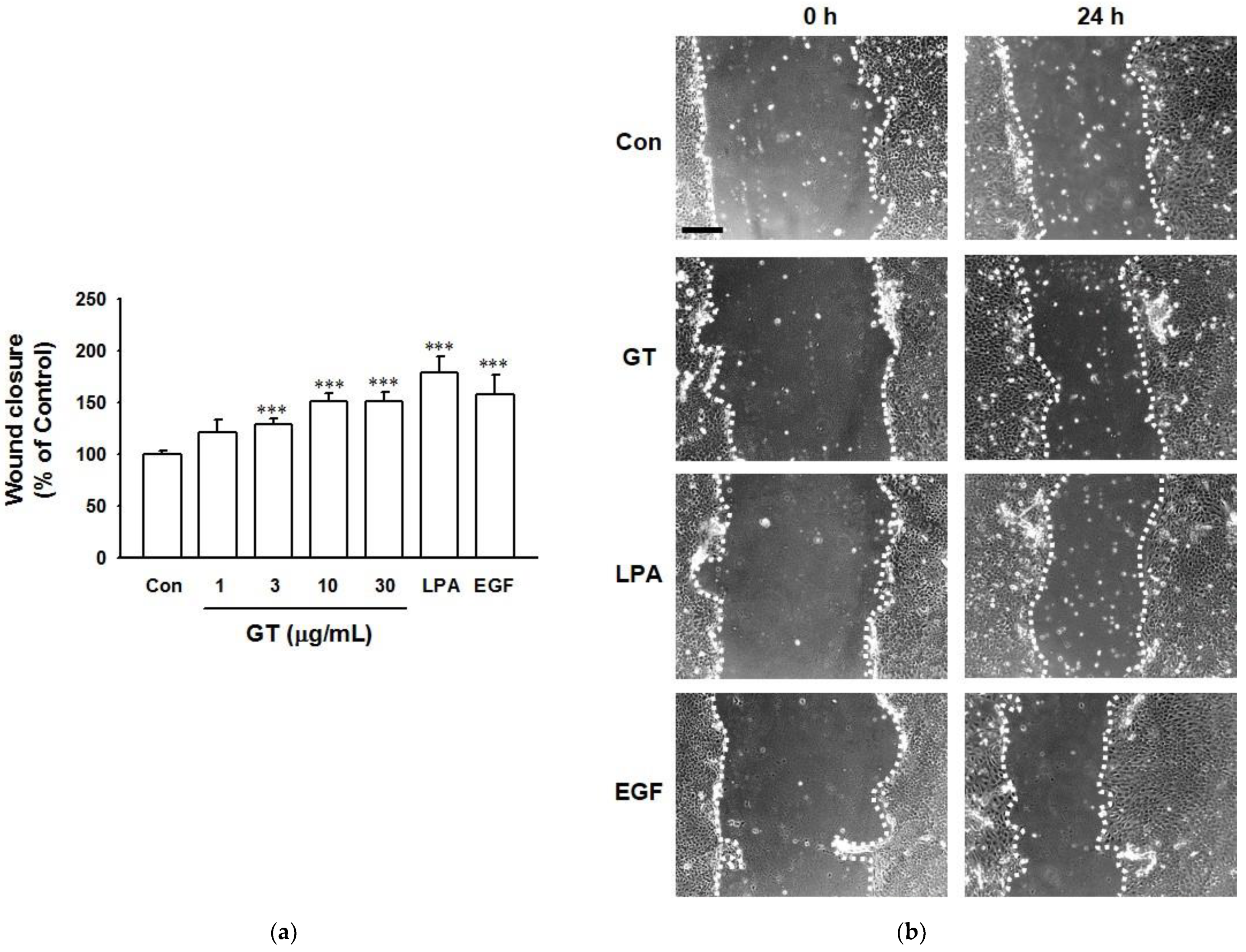
2.2. In Vitro Wound Healing Effect of Gintonin
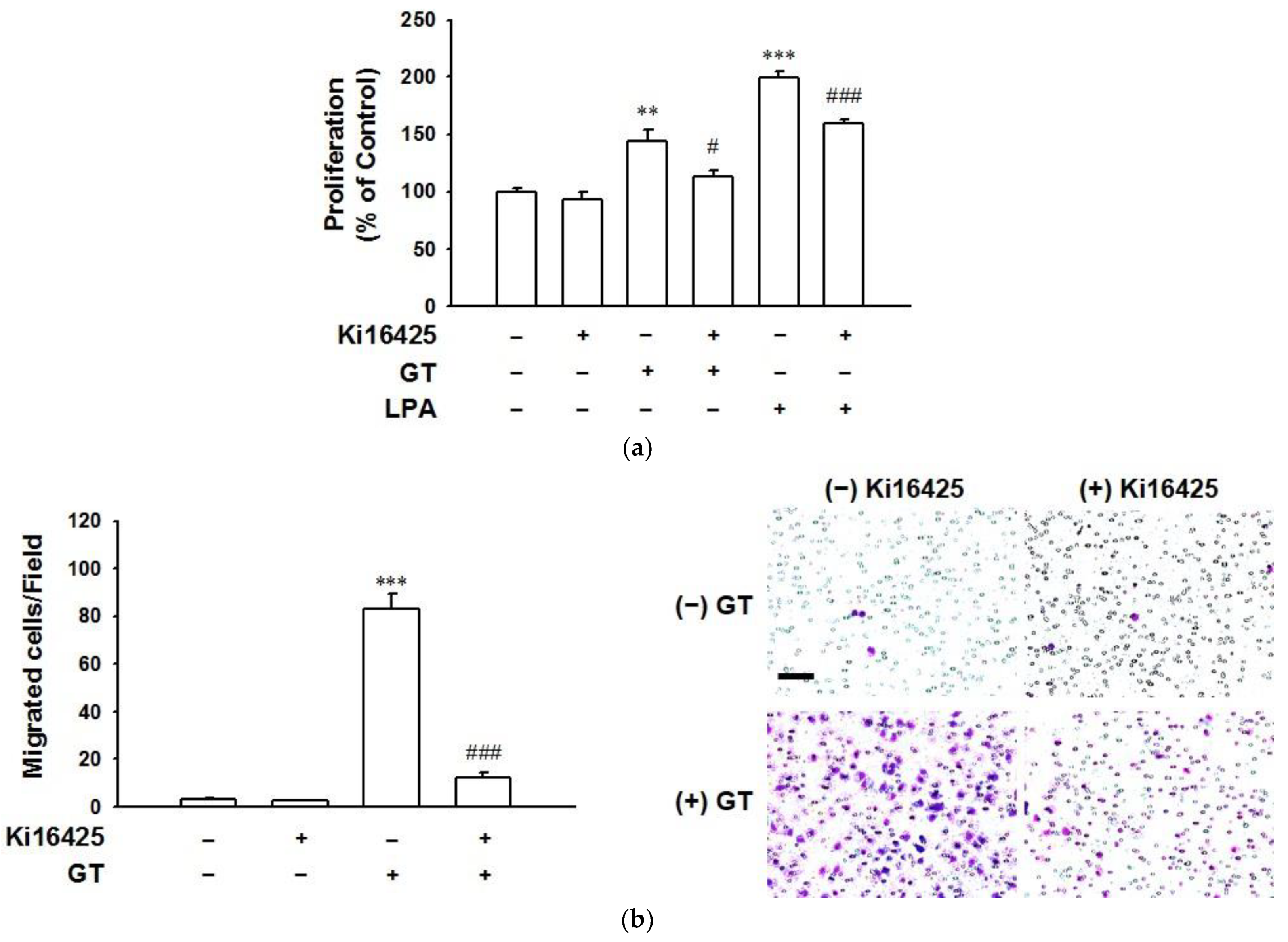
2.3. Involvement of LPA1 Receptor Subtypes in Gintonin-Mediated HaCaT Cell Proliferation
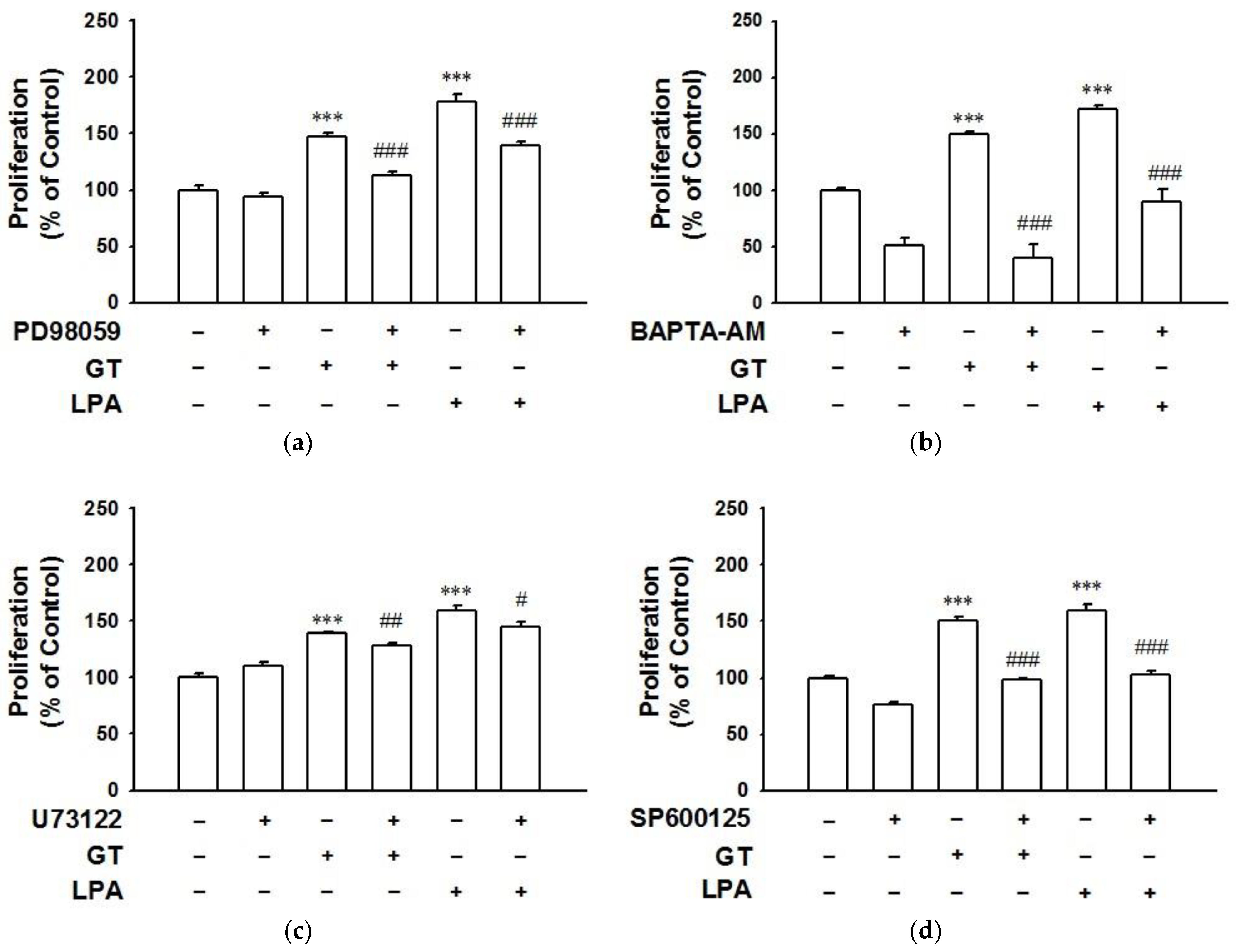
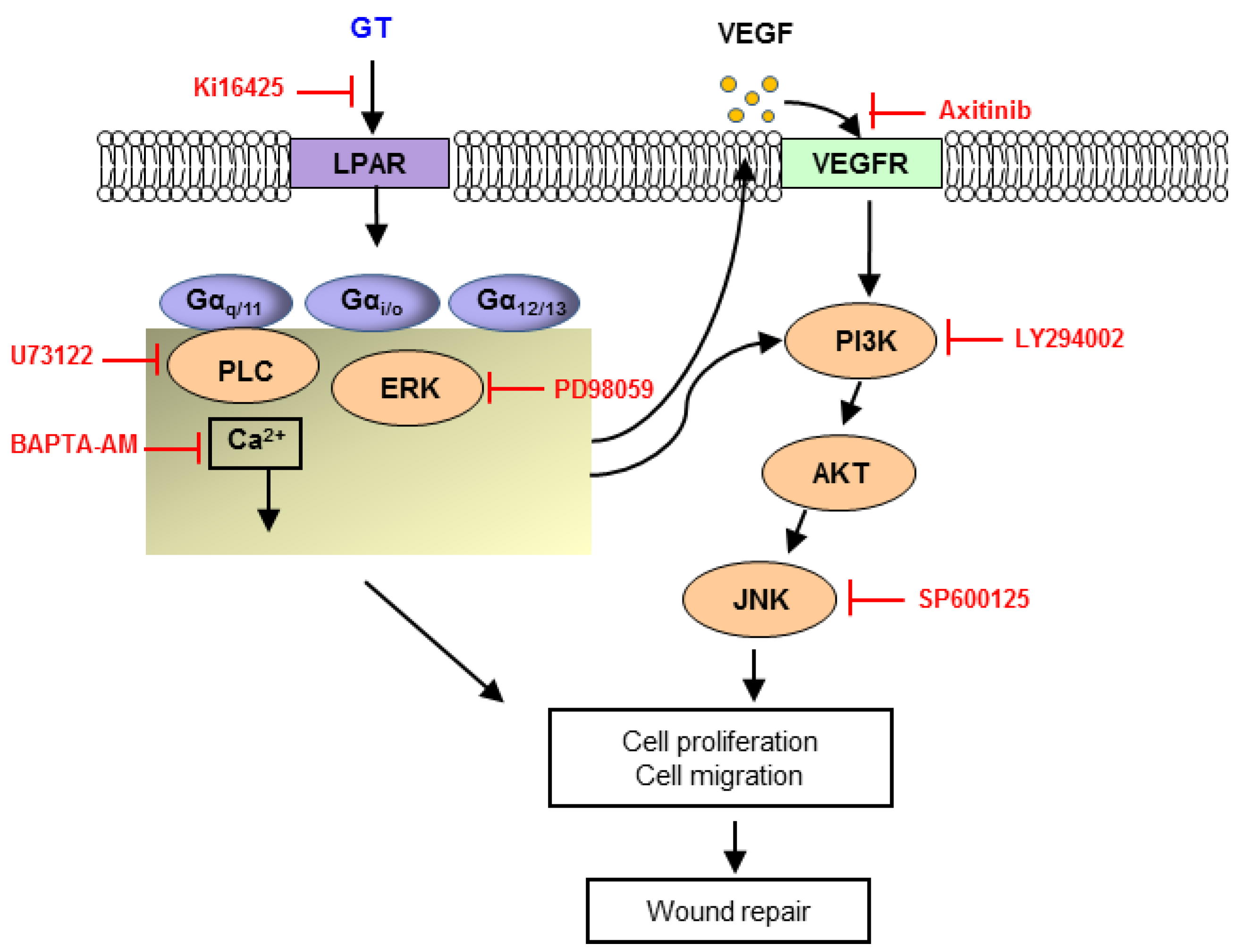
2.4. Signaling Pathways of Gintonin Effect
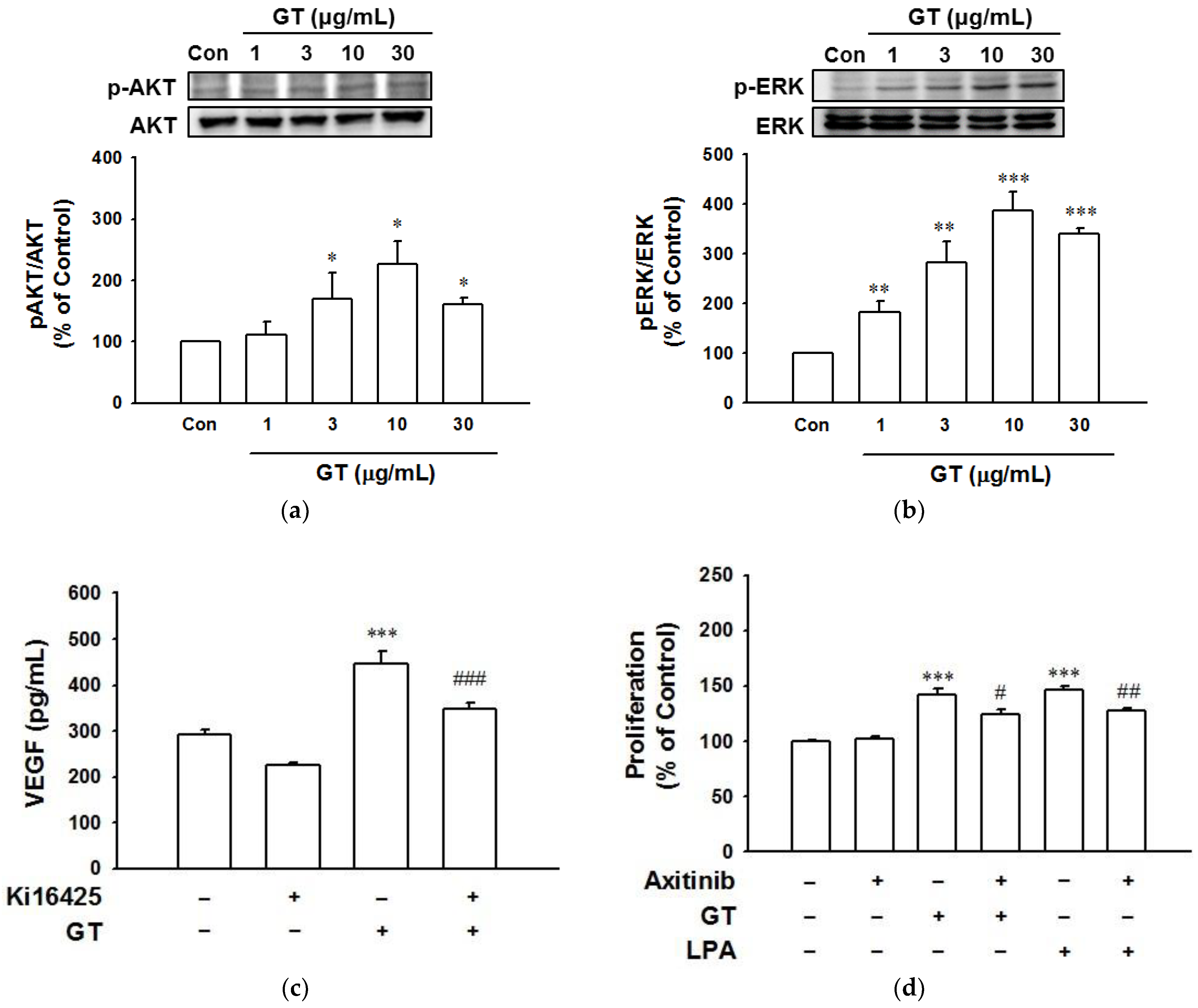
2.5. Phosphorylation of Akt and ERK in HaCaT Cells Exposed to Gintonin
2.6. Increased VEGF Release from HaCaT Cells Exposed to Gintonin
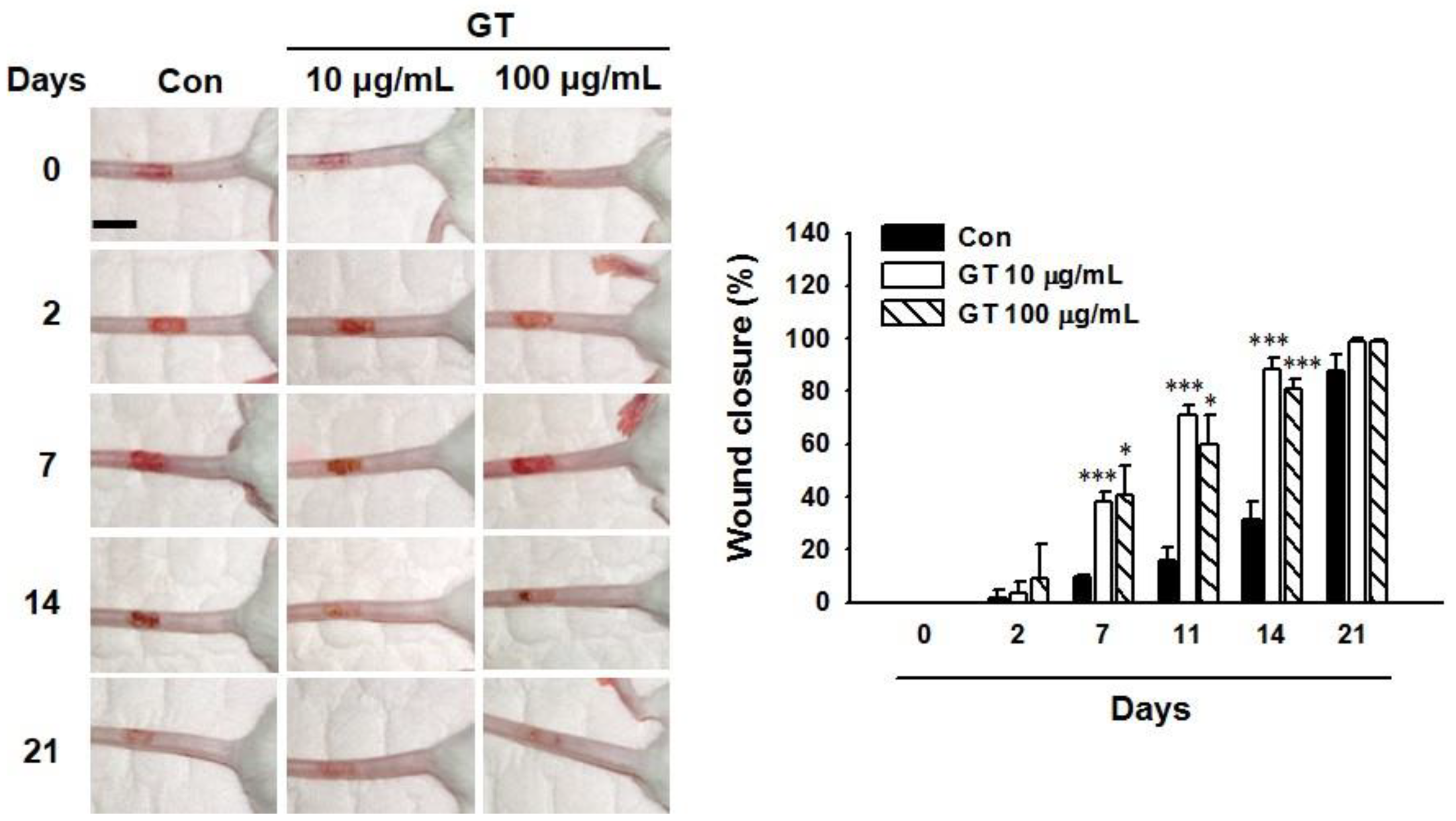
2.7. Gintonin Facilitates In Vivo Wound Repair of Mouse Tail
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Cell Proliferation Assay
4.4. Migration Assay
4.5. Scratch Wound Healing Assay
4.6. VEGF ELISA
4.7. Immunoblotting
4.8. In Vivo Wound Healing
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ratan, Z.A.; Haidere, M.F.; Hong, Y.H.; Park, S.H.; Lee, J.-O.; Lee, J.; Cho, J.Y. Pharmacological potential of ginseng and its major component ginsenosides. J. Ginseng Res. 2020, 45, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Jang, D.; Park, S.K.; Lee, M.-G.; Cha, B.; Park, C.; Shin, Y.S.; Park, H.; Baek, J.-M.; Heo, H.; et al. Ginsenoside 20(S)-protopanaxadiol induces cell death in human endometrial cancer cells via apoptosis. J. Ginseng Res. 2020, 45, 126–133. [Google Scholar] [CrossRef]
- Yoon, S.J.; Kim, S.K.; Lee, N.Y.; Choi, Y.R.; Kim, H.S.; Gupta, H.; Youn, G.S.; Sung, H.; Shin, M.J.; Suk, K.T. Effect of Korean Red Ginseng on metabolic syndrome. J. Ginseng Res. 2020, 45, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Mo, E.J.; Choi, J.E.; Jo, Y.H.; Jang, H.; Jeong, J.Y.; Jin, Q.; Chung, H.N.; Hwang, B.Y.; Lee, M.K. Effect of Korean Red Ginseng extraction conditions on antioxidant activity, extraction yield, and ginsenoside Rg1 and phenolic content: Optimization using response surface methodology. J. Ginseng Res. 2015, 40, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Yang, Z.; Gao, L.; Liu, W.; Liu, R.; Zhao, J.; You, J. Changes in element accumulation, phenolic metabolism, and antioxidative enzyme activities in the red-skin roots of Panax ginseng. J. Ginseng Res. 2016, 41, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Han, S.Y.; Kim, J.; Kim, E.; Kim, S.H.; Seo, D.B.; Kim, J.-H.; Shin, S.S.; Cho, J.Y. AKT-targeted anti-inflammatory activity of Panax ginseng calyx ethanolic extract. J. Ginseng Res. 2017, 42, 496–503. [Google Scholar] [CrossRef]
- Kim, J.H.; Yi, Y.-S.; Kim, M.-Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2016, 41, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Lee, D.; Lee, H.L.; Kim, C.-E.; Jung, K.; Kang, K.S. Beneficial effects of Panax ginseng for the treatment and prevention of neurodegenerative diseases: Past findings and future directions. J. Ginseng Res. 2017, 42, 239–247. [Google Scholar] [CrossRef]
- Huang, J.; Liu, D.; Wang, Y.; Liu, L.; Li, J.; Yuan, J.; Jiang, Z.; Jiang, Z.; Hsiao, W.W.; Liu, H.; et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut 2021, gutjnl-2020-321031. [Google Scholar] [CrossRef]
- Pyo, M.-K.; Choi, S.-H.; Shin, T.-J.; Hwang, S.-H.; Lee, B.-H.; Kang, J.-Y.; Kim, H.-J.; Lee, S.-H.; Nah, S.-Y. A simple method for the preparation of Crude Gintonin from Ginseng root, stem, and leaf. J. Ginseng Res. 2011, 35, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.H.; Shin, T.-J.; Choi, S.-H.; Cho, H.-J.; Lee, B.-H.; Pyo, M.K.; Lee, J.-H.; Kang, J.; Kim, H.-J.; Park, C.-W.; et al. Gintonin, newly identified compounds from ginseng, is novel lysophosphatidic acids-protein complexes and activates G protein-coupled lysophosphatidic acid receptors with high affinity. Mol. Cells 2012, 33, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aikawa, S.; Hashimoto, T.; Kano, K.; Aoki, J. Lysophosphatidic acid as a lipid mediator with multiple biological actions. J. Biochem. 2014, 157, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraldo, L.H.M.; de Sampaio Spohr, T.C.L.; Amaral, R.F.D.; da Fonseca, A.C.C.; Garcia, C.; Mendes, F.D.A.; Freitas, C.; Dossantos, M.F.; Lima, F.R.S. Role of lysophosphatidic acid and its receptors in health and disease: Novel therapeutic strategies. Signal Transduct. Target. Ther. 2021, 6, 1–18. [Google Scholar] [CrossRef]
- Hwang, S.-H.; Lee, B.-H.; Choi, S.-H.; Kim, H.-J.; Won, K.J.; Lee, H.M.; Rhim, H.; Kim, H.-C.; Nah, S.-Y. Effects of gintonin on the proliferation, migration, and tube formation of human umbilical-vein endothelial cells: Involvement of lysophosphatidic-acid receptors and vascular-endothelial-growth-factor signaling. J. Ginseng Res. 2015, 40, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.H.; Shin, E.-J.; Shin, T.-J.; Lee, B.-H.; Choi, S.-H.; Kang, J.; Kim, H.-J.; Kwon, S.-H.; Jang, C.-G.; Lee, J.-H.; et al. Gintonin, a Ginseng-Derived Lysophosphatidic Acid Receptor Ligand, Attenuates Alzheimer‘s Disease-Related Neuropathies: Involvement of Non-Amyloidogenic Processing. J. Alzheimer’s Dis. 2012, 31, 207–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.-E.; Park, S.-D.; Hwang, H.; Choi, S.-H.; Lee, R.M.; Nam, S.M.; Choi, J.H.; Rhim, H.; Cho, I.-H.; Kim, H.-C.; et al. Effects of a gintonin-enriched fraction on hair growth: An in vitro and in vivo study. J. Ginseng Res. 2019, 44, 168–177. [Google Scholar] [CrossRef]
- You, L.; Cho, J.Y. The regulatory role of Korean ginseng in skin cells. J. Ginseng Res. 2020, 45, 363–370. [Google Scholar] [CrossRef]
- Park, M.Y.; Han, S.J.; Moon, D.; Kwon, S.; Lee, J.-W.; Kim, K.S. Effects of red ginseng on the elastic properties of human skin. J. Ginseng Res. 2019, 44, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-O.; Hwang, S.-H.; Shen, T.; Kim, J.H.; You, L.; Hu, W.; Cho, J.Y. Enhancement of skin barrier and hydration-related molecules by protopanaxatriol in human keratinocytes. J. Ginseng Res. 2020, 45, 354–360. [Google Scholar] [CrossRef]
- Lee, R.; Lee, N.-E.; Hwang, H.; Rhim, H.; Cho, I.-H.; Nah, S.-Y. Ginseng gintonin enhances hyaluronic acid and collagen release from human dermal fibroblasts through lysophosphatidic acid receptor Interaction. Molecules 2019, 24, 4438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; DiPietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef]
- Broughton, G.; Janis, J.; Attinger, C.E. The basic science of wound healing. Plast. Reconstr. Surg. 2006, 117, 12S–34S. [Google Scholar] [CrossRef]
- Kirfel, G.; Herzog, V. Migration of epidermal keratinocytes: Mechanisms, regulation, and biological significance. Protoplasma 2004, 223, 67–78. [Google Scholar] [CrossRef]
- McKay, I.; Leigh, I. Epidermal cytokines and their roles in cutaneous wound healing. Br. J. Dermatol. 1991, 124, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Tsoi, L.C.; Billi, A.C.; Ward, N.L.; Harms, P.W.; Zeng, C.; Maverakis, E.; Kahlenberg, J.M.; Gudjonsson, J.E. Cytokinocytes: The diverse contribution of keratinocytes to immune responses in skin. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Squarize, C.; Castilho, R.M.; Bugge, T.H.; Gutkind, J.S. Accelerated Wound Healing by mTOR Activation in Genetically Defined Mouse Models. PLoS ONE 2010, 5, e10643. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Martinotti, S.; Burlando, B. Wound healing properties of jojoba liquid wax: An in vitro study. J. Ethnopharmacol. 2011, 134, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.K.; Choi, S.W.; Lee, H.-R.; Lee, E.-J.; Lee, K.-H.; Kim, H.O. Purinoceptor-mediated calcium mobilization and proliferation in HaCaT keratinocytes. J. Dermatol. Sci. 2001, 25, 97–105. [Google Scholar] [CrossRef]
- Sharma, G.-D.; He, J.; Bazan, H.E. p38 and ERK1/2 Coordinate cellular migration and proliferation in epithelial wound healing: Evidence of cross-talk activation between MAP kinase cascades. J. Biol. Chem. 2003, 278, 21989–21997. [Google Scholar] [CrossRef] [Green Version]
- Haase, I.; Evans, R.; Pofahl, R.; Watt, F. Regulation of keratinocyte shape, migration and wound epithelialization by IGF-1- and EGF-dependent signalling pathways. J. Cell Sci. 2003, 116, 3227–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Graves, D.T. Keratinocyte function in normal and diabetic wounds and modulation by FOXO1. J. Diabetes Res. 2020, 2020, 3714704. [Google Scholar] [CrossRef]
- El Ashram, S.; El-Samad, L.M.; Basha, A.A.; El Wakil, A. Naturally-derived targeted therapy for wound healing: Beyond classical strategies. Pharmacol. Res. 2021, 170, 105749. [Google Scholar] [CrossRef]
- Sivamani, R.K.; Ma, B.R.; Wehrli, L.N.; Maverakis, E. Phytochemicals and naturally derived substances for wound healing. Adv. Wound Care 2012, 1, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, H.-N.; Yang, M.-J.; Chang, C.-F.; Chau, Y.-P.; Lu, K.-S. In vitro and in vivo wound healing-promoting activities of β-lapachone. Am. J. Physiol. Physiol. 2008, 295, C931–C943. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.; Martínez, N.; Rodríguez-García, I.; Alonso, A. EGFR Family-mediated signal transduction in the human keratinocyte cell line HaCaT. Exp. Cell Res. 1999, 252, 432–438. [Google Scholar] [CrossRef]
- Burgess, A.W. EGFR family: Structure physiology signalling and therapeutic targets. Growth Factors 2008, 26, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, F.; Wang, X.; Yang, J.; Li, Z.; Cong, X.; Chen, X. Lysophosphatidic acid promotes secretion of VEGF by increasing expression of 150-kD Oxygen-regulated protein (ORP150) in mesenchymal stem cells. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2013, 1831, 1426–1434. [Google Scholar] [CrossRef]
- Mazereeuw-Hautier, J.; Gres, S.; Fanguin, M.; Cariven, C.; Fauvel, J.; Perret, B.; Chap, H.; Salles, J.-P.; Saulnier-Blache, J.-S. Production of lysophosphatidic acid in blister fluid: Involvement of a Lysophospholipase D Activity. J. Investig. Dermatol. 2005, 125, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumitomo, A.; Siriwach, R.; Thumkeo, D.; Ito, K.; Nakagawa, R.; Tanaka, N.; Tanabe, K.; Watanabe, A.; Kishibe, M.; Ishida-Yamamoto, A.; et al. LPA Induces keratinocyte differentiation and promotes skin barrier function through the LPAR1/LPAR5-RHO-ROCK-SRF axis. J. Investig. Dermatol. 2019, 139, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- Eichholtz, T.; Jalink, K.; Fahrenfort, I.; Moolenaar, W.H. The bioactive phospholipid lysophosphatidic acid is released from activated platelets. Biochem. J. 1993, 291, 677–680. [Google Scholar] [CrossRef] [Green Version]
- Piazza, G.A.; Ritter, J.L.; Baracka, C.A. Lysophosphatidic Acid induction of transforming growth factors α and β: Modulation of proliferation and differentiation in cultured human keratinocytes and mouse skin. Exp. Cell Res. 1995, 216, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Rhim, J.H.; Jang, I.-S.; Kwon, S.T.; Song, K.Y.; Yeo, E.-J.; Park, S.C. Activation of Wound Healing in Aged Rats by Altering the Cellular Mitogenic Potential. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2010, 65, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Saba, E.; Jeon, B.R.; Jeong, D.-H.; Lee, K.; Goo, Y.-K.; Kwak, D.; Kim, S.; Roh, S.-S.; Kim, S.D.; Nah, S.-Y.; et al. A Novel Korean Red Ginseng Compound Gintonin Inhibited Inflammation by MAPK and NF-κB Pathways and recovered the levels of mir-34a and mir-93 in RAW 264.7 cells. Evid. Based Complementary Altern. Med. 2015, 2015, 624132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-H.; Shin, T.-J.; Lee, B.-H.; Hwang, S.-H.; Kang, J.-Y.; Kim, H.-J.; Park, C.-W.; Nah, S.-Y. An Edible Gintonin preparation from Ginseng. J. Ginseng Res. 2011, 35, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, C.-H.; Sung, H.-K.; Kim, K.-T.; Cheon, H.G.; Oh, G.T.; Hong, H.J.; Yoo, O.-J.; Koh, G.Y. COMP-angiopoietin-1 promotes wound healing through enhanced angiogenesis, lymphangiogenesis, and blood flow in a diabetic mouse model. Proc. Natl. Acad. Sci. USA 2006, 103, 4946–4951. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.-H.; Won, K.-J.; Lee, R.; Cho, H.-S.; Hwang, S.-H.; Nah, S.-Y. Wound Healing Effect of Gintonin Involves Lysophosphatidic Acid Receptor/Vascular Endothelial Growth Factor Signaling Pathway in Keratinocytes. Int. J. Mol. Sci. 2021, 22, 10155. https://doi.org/10.3390/ijms221810155
Choi S-H, Won K-J, Lee R, Cho H-S, Hwang S-H, Nah S-Y. Wound Healing Effect of Gintonin Involves Lysophosphatidic Acid Receptor/Vascular Endothelial Growth Factor Signaling Pathway in Keratinocytes. International Journal of Molecular Sciences. 2021; 22(18):10155. https://doi.org/10.3390/ijms221810155
Chicago/Turabian StyleChoi, Sun-Hye, Kyung-Jong Won, Rami Lee, Han-Sung Cho, Sung-Hee Hwang, and Seung-Yeol Nah. 2021. "Wound Healing Effect of Gintonin Involves Lysophosphatidic Acid Receptor/Vascular Endothelial Growth Factor Signaling Pathway in Keratinocytes" International Journal of Molecular Sciences 22, no. 18: 10155. https://doi.org/10.3390/ijms221810155