
An O-GlcNAcylomic Approach Reveals ACLY as a Potential Target in Sepsis in the Young Rat

 , , , , , , , , add
Show full author list
, , , , , , , , add
Show full author list
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
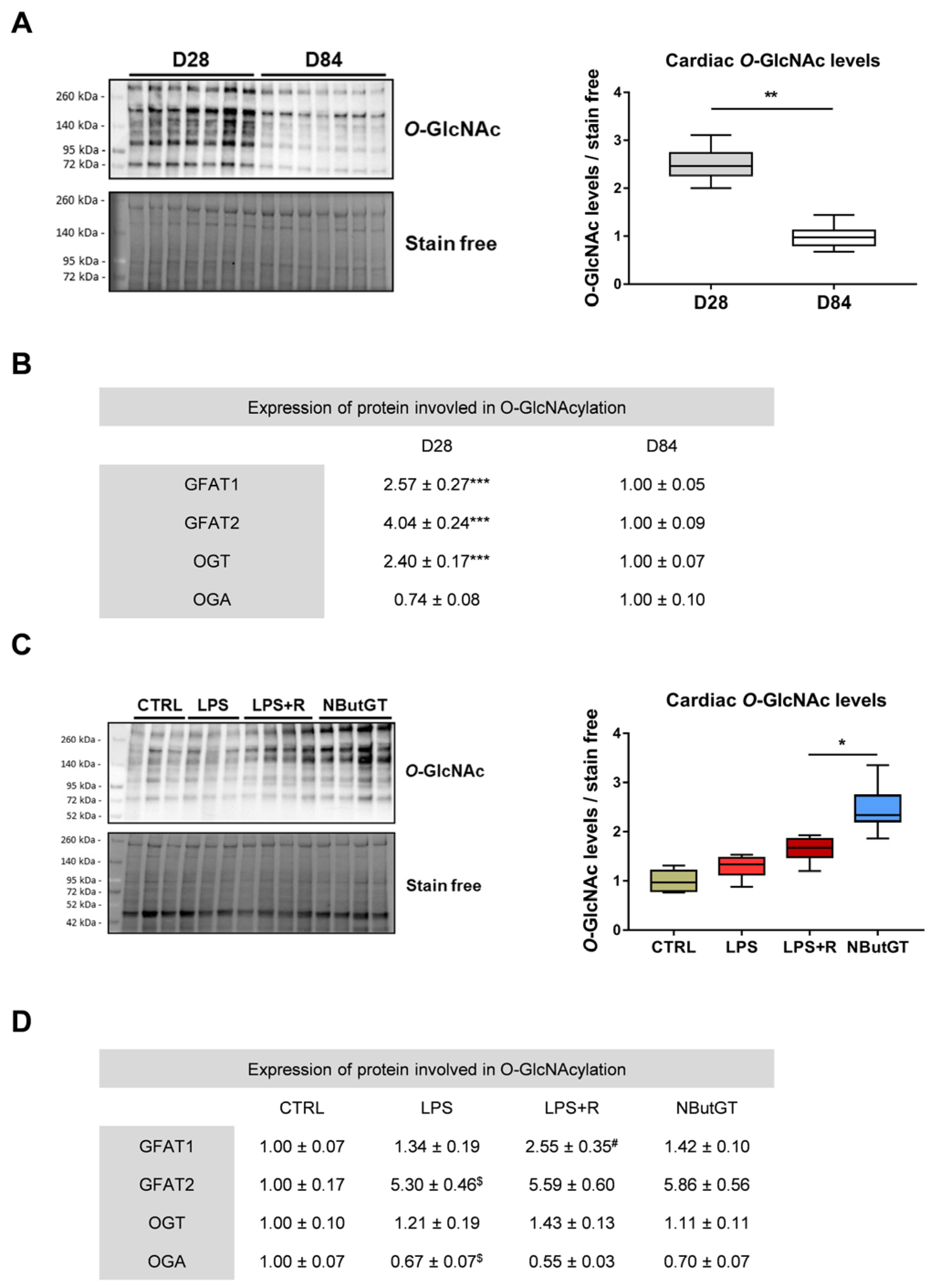
2.1. O-GlcNAcylation Levels Decrease during the Early Stage of Life
2.2. O-GlcNAcylation Levels Increase in Response to Sepsis in Young Rats
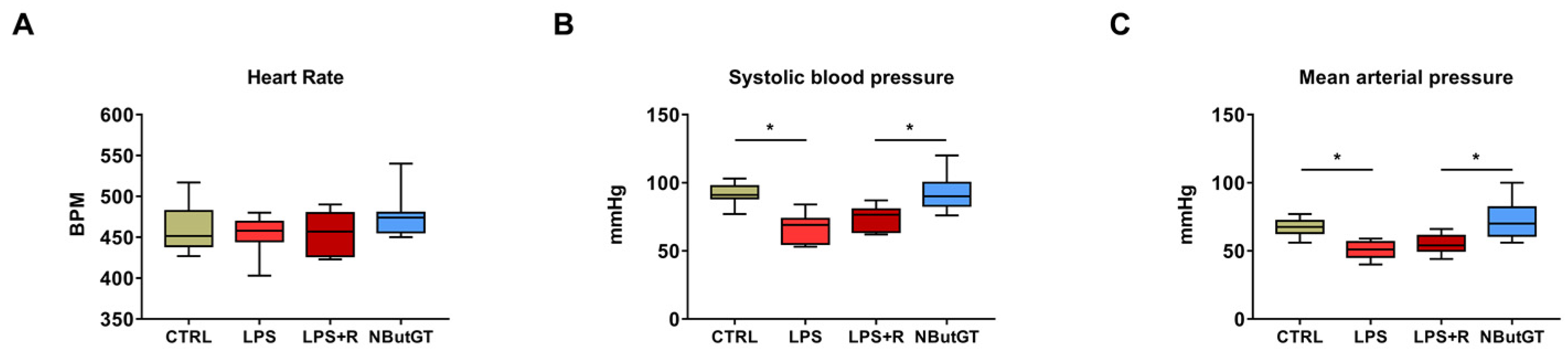
2.3. O-GlcNAc Stimulation Significantly Improves Blood Pressure
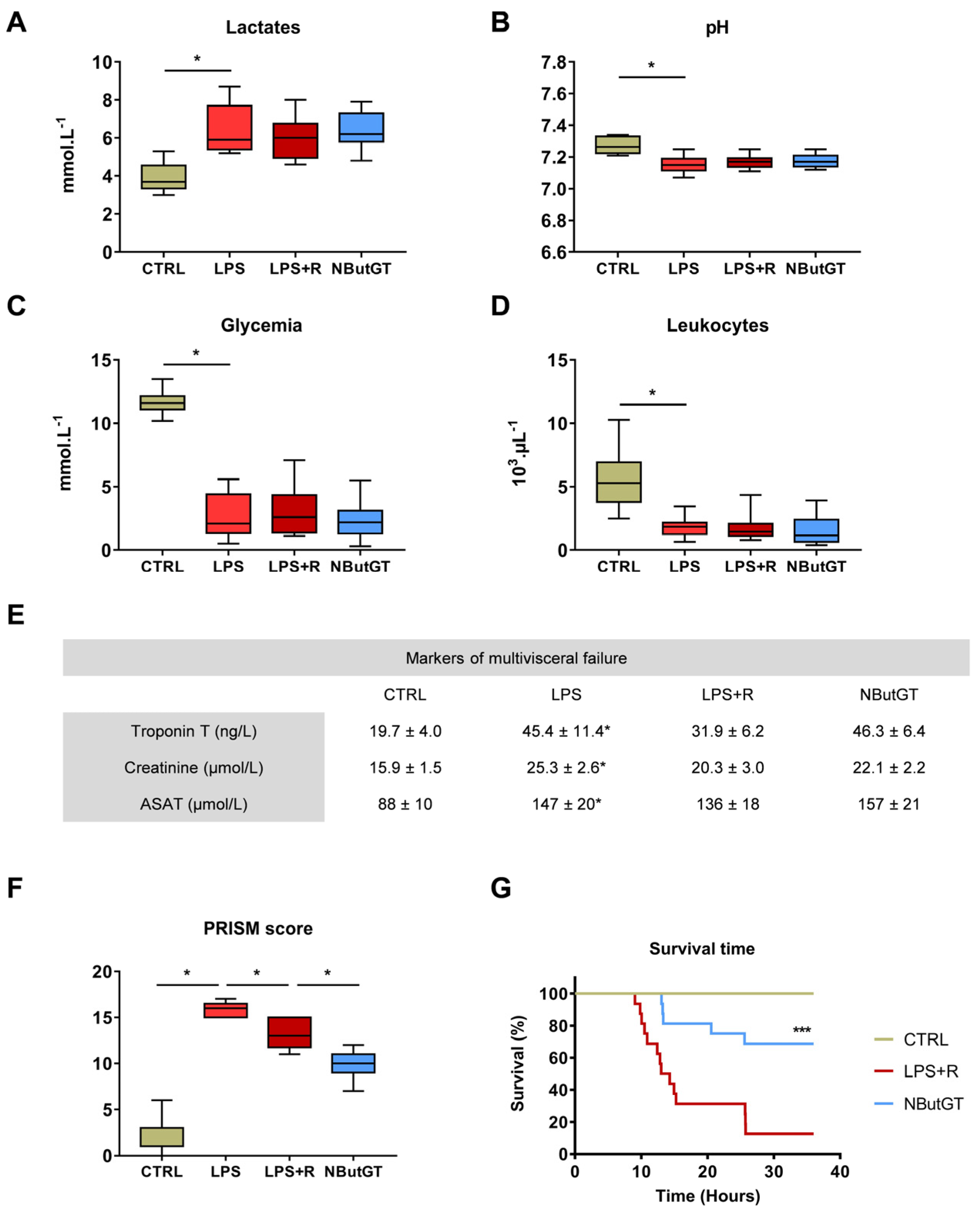
2.4. O-GlcNAc Stimulation in the Young Population Does Not Correct Circulating Parameters
2.5. The General Condition and Survival of Young Animals Is Improved by Nbutgt Treatment
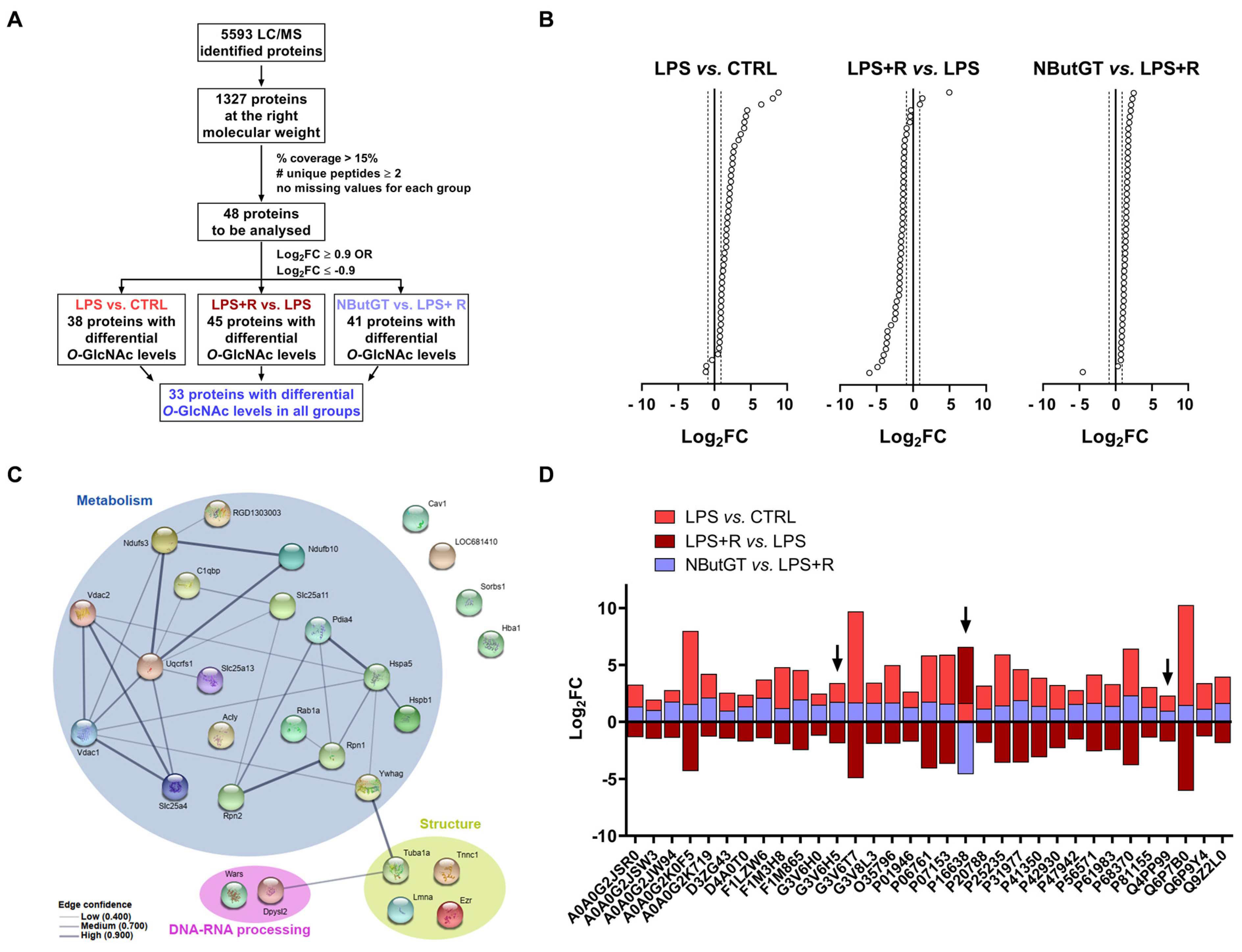
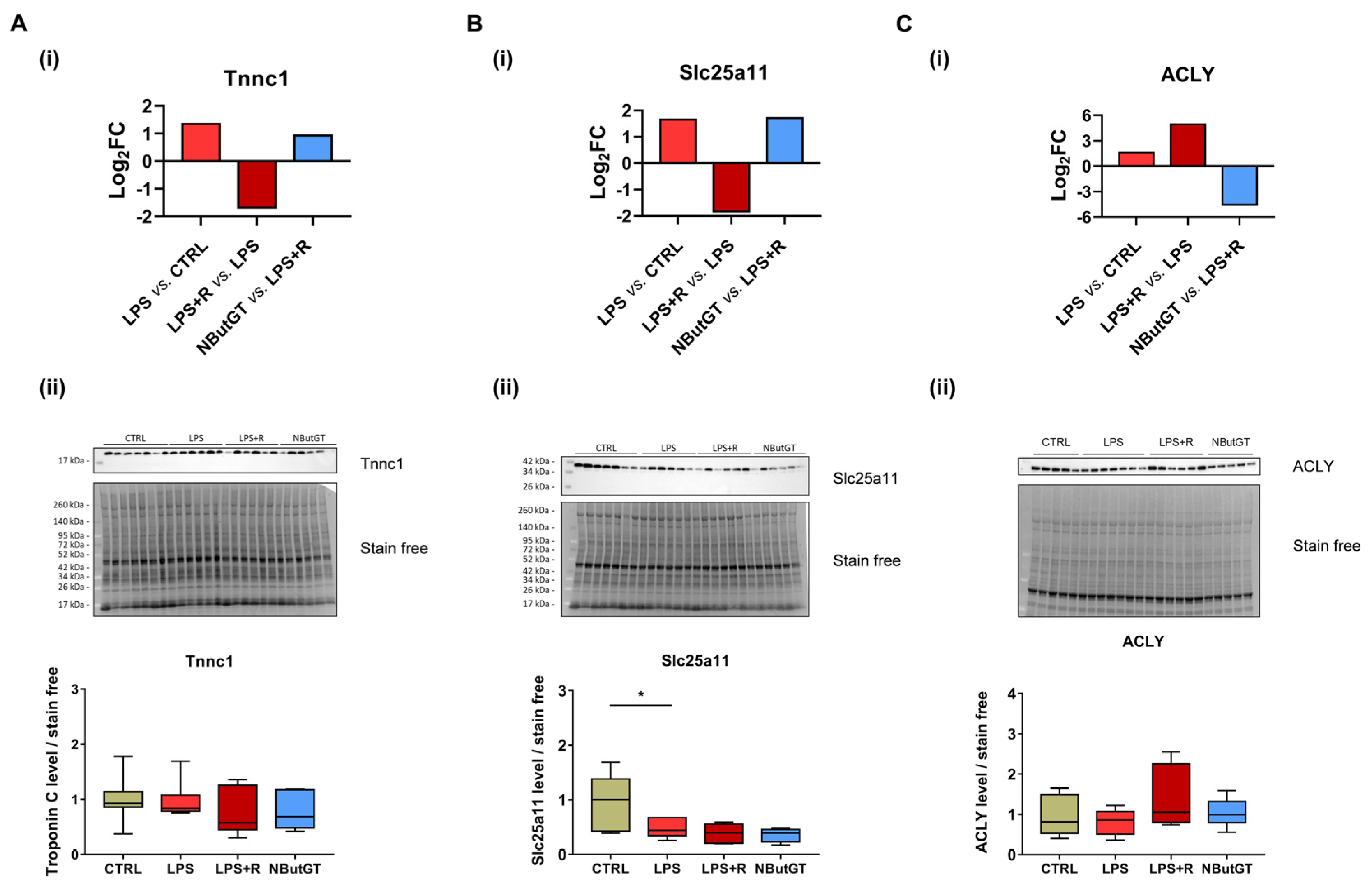
2.6. Identification of Putative Cardiac O-GlcNAcylated Proteins by O-GlcNAcylomic
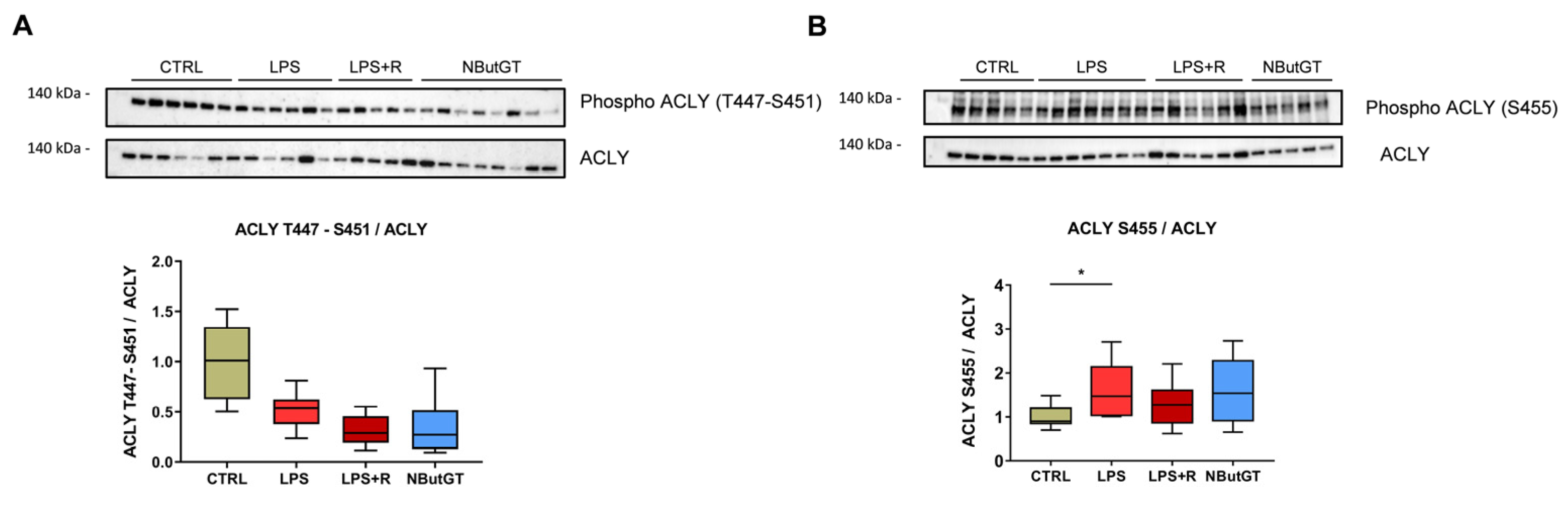
2.7. ATP-Citrate Lyase Is Less O-GlcNAcylated in the NButGT Group
3. Discussion
3.1. Cardiac O-GlcNAc throughout Aging
3.2. Validation of the Septic Shock Model in Young Rats
3.3. Impact of O-GlcNAcylation Stimulation in Septic Shock in Young Rats
3.4. O-GlcNAcylomic Analysis
4. Limits
5. Conclusions
6. Methods
6.1. Reagents
6.2. Animal Model and Measures
6.3. Tissue Preparation and Protein Extraction
6.4. Western Blot
6.5. Proteomic Study
6.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; De Santis, V.; Vitale, D.; Jeffcoate, W. Multiorgan Failure Is an Adaptive, Endocrine-Mediated, Metabolic Response to Overwhelming Systemic Inflammation. Lancet 2004, 364, 545–548. [Google Scholar] [CrossRef]
- Angus, D.C.; van der Poll, T. Severe Sepsis and Septic Shock. N. Engl. J. Med. 2013, 369, 2063. [Google Scholar] [CrossRef]
- Deutschman, C.S.; Tracey, K.J. Sepsis: Current Dogma and New Perspectives. Immunity 2014, 40, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiersinga, W.J.; Leopold, S.J.; Cranendonk, D.R.; van der Poll, T. Host Innate Immune Responses to Sepsis. Virulence 2014, 5, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Girard, T.D.; Opal, S.M.; Ely, E.W. Insights into Severe Sepsis in Older Patients: From Epidemiology to Evidence-Based Management. Clin. Infect. Dis. 2005, 40, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Hartman, M.E.; Linde-Zwirble, W.T.; Angus, D.C.; Watson, R.S. Trends in the Epidemiology of Pediatric Severe Sepsis. Pediatr. Crit. Care Med. 2013, 14, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Randolph, A.G.; McCulloh, R.J. Pediatric Sepsis: Important Considerations for Diagnosing and Managing Severe Infections in Infants, Children, and Adolescents. Virulence 2014, 5, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.L.; Fitzgerald, J.C.; Pappachan, J.; Wheeler, D.; Jaramillo-Bustamante, J.C.; Salloo, A.; Singhi, S.C.; Erickson, S.; Roy, J.A.; Bush, J.L.; et al. Global Epidemiology of Pediatric Severe Sepsis: The Sepsis Prevalence, Outcomes, and Therapies Study. Am. J. Respir. Crit. Care Med. 2015, 191, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.L.; Peters, M.J.; Alhazzani, W.; Agus, M.S.D.; Flori, H.R.; Inwald, D.P.; Nadel, S.; Schlapbach, L.J.; Tasker, R.C.; Argent, A.C.; et al. Surviving Sepsis Campaign International Guidelines for the Management of Septic Shock and Sepsis-Associated Organ Dysfunction in Children. Pediatr. Crit. Care Med. 2020, 21, e52. [Google Scholar] [CrossRef]
- Deep, A.; Goonasekera, C.D.A.; Wang, Y.; Brierley, J. Evolution of Haemodynamics and Outcome of Fluid-Refractory Septic Shock in Children. Intensive Care Med 2013, 39, 1602–1609. [Google Scholar] [CrossRef]
- Emr, B.M.; Alcamo, A.M.; Carcillo, J.A.; Aneja, R.K.; Mollen, K.P. Pediatric Sepsis Update: How Are Children Different? Surg. Infect. 2018, 19, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Bond, M.R.; Hanover, J.A. A Little Sugar Goes a Long Way: The Cell Biology of O-GlcNAc. J. Cell Biol. 2015, 208, 869–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Qian, K. Protein O-GlcNAcylation: Emerging Mechanisms and Functions. Nat. Rev. Mol. Cell Biol. 2017, 18, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.R.; Dias, T.B.; Natov, P.S.; Zachara, N.E. Stress-Induced O-GlcNAcylation: An Adaptive Process of Injured Cells. Biochem. Soc. Trans. 2017, 45, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Ferron, M.; Denis, M.; Persello, A.; Rathagirishnan, R.; Lauzier, B. Protein O-GlcNAcylation in Cardiac Pathologies: Past, Present, Future. Front. Endocrinol. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Yi, W. O-GlcNAcylation, a Sweet Link to the Pathology of Diseases. J. Zhejiang Univ. Sci. B 2019, 20, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Ferron, M.; Cadiet, J.; Persello, A.; Prat, V.; Denis, M.; Erraud, A.; Aillerie, V.; Mevel, M.; Bigot, E.; Chatham, J.C.; et al. O-GlcNAc Stimulation: A New Metabolic Approach to Treat Septic Shock. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Silva, J.F.; Olivon, V.C.; Mestriner, F.L.A.C.; Zanotto, C.Z.; Ferreira, R.G.; Ferreira, N.S.; Silva, C.A.A.; Luiz, J.P.M.; Alves, J.V.; Fazan, R.; et al. Acute Increase in O-GlcNAc Improves Survival in Mice With LPS-Induced Systemic Inflammatory Response Syndrome. Front. Physiol. 2020, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- Dupas, T.; Denis, M.; Dontaine, J.; Persello, A.; Bultot, L.; Erraud, A.; Vertommen, D.; Bouchard, B.; Tessier, A.; Rivière, M.; et al. Protein O-GlcNAcylation Levels Are Regulated Independently of Dietary Intake in a Tissue and Time-Specific Manner during Rat Postnatal Development. Acta Physiol. 2020, 231, e13566. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.-X.; Dai, C.; Liu, F.; Ge, J.; Shi, J.; Gu, J.; Iqbal, K.; Yu, Q.; Zhao, Y.; Chen, Y.; et al. O-GlcNAcylation Reduces Ischemia-Reperfusion–Induced Brain Injury. Sci. Rep. 2017, 7, 10686. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, P.S.; Lagerlöf, O.; Hart, G.W. Roles of O-GlcNAc in Chronic Diseases of Aging. Mol. Aspects Med. 2016, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.L.; Carcillo, J.A.; Aneja, R.K.; Deymann, A.J.; Lin, J.C.; Nguyen, T.C.; Okhuysen-Cawley, R.S.; Relvas, M.S.; Rozenfeld, R.A.; Skippen, P.W.; et al. American College of Critical Care Medicine Clinical Practice Parameters for Hemodynamic Support of Pediatric and Neonatal Septic Shock. Crit. Care Med. 2017, 45, 1061–1093. [Google Scholar] [CrossRef] [PubMed]
- Dobkin, E.D.; Lobe, T.E.; Bhatia, J.; Oldham, K.T.; Traber, D.L. The Study of Fecal-Escherichia Coli Peritonitis-Induced Septic Shock in a Neonatal Pig Model. Circ. Shock. 1985, 16, 325–336. [Google Scholar] [PubMed]
- Al-Mukh, H.; Baudoin, L.; Bouaboud, A.; Sanchez-Salgado, J.-L.; Maraqa, N.; Khair, M.; Pagesy, P.; Bismuth, G.; Niedergang, F.; Issad, T. Lipopolysaccharide Induces GFAT2 Expression to Promote O-Linked β-N-Acetylglucosaminylation and Attenuate Inflammation in Macrophages. J. Immunol. 2020, 205, 2499–2510. [Google Scholar] [CrossRef]
- Inata, Y.; Piraino, G.; Hake, P.W.; O’Connor, M.; Lahni, P.; Wolfe, V.; Schulte, C.; Moore, V.; James, J.M.; Zingarelli, B. Age-Dependent Cardiac Function during Experimental Sepsis: Effect of Pharmacological Activation of AMP-Activated Protein Kinase by AICAR. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H826–H837. [Google Scholar] [CrossRef]
- Eisenberg, M.A.; Green-Hopkins, I.; Alexander, M.E.; Chiang, V.W. Cardiac Troponin T as a Screening Test for Myocarditis in Children. Pediatr. Emerg. Care 2012, 28, 1173–1178. [Google Scholar] [CrossRef]
- Sankar, J.; Das, R.R.; Jain, A.; Dewangan, S.; Khilnani, P.; Yadav, D.; Dubey, N. Prevalence and Outcome of Diastolic Dysfunction in Children with Fluid Refractory Septic Shock—a Prospective Observational Study. Pediatr. Crit. Care Med. 2014, 15, e370–e378. [Google Scholar] [CrossRef] [Green Version]
- Ekmark, L.; Rooyackers, O.; Wernerman, J.; Fläring, U. Plasma Glutamine Deficiency Is Associated with Multiple Organ Failure in Critically Ill Children. Amino Acids 2015, 47, 535–542. [Google Scholar] [CrossRef]
- Barbosa, E.; Moreira, E.A.; Goes, J.E.; Faintuch, J. Pilot Study with a Glutamine-Supplemented Enteral Formula in Critically Ill Infants. Rev. Hosp. Clin. Fac. Med. Sao Paulo 1999, 54, 21–24. [Google Scholar] [CrossRef]
- Katrukha, I.A. Human Cardiac Troponin Complex. Structure and Functions. Biochemistry 2013, 78, 1447–1465. [Google Scholar] [CrossRef]
- Zanotti-Cavazzoni, S.L.; Hollenberg, S.M. Cardiac Dysfunction in Severe Sepsis and Septic Shock. Curr. Opin. Crit. Care 2009, 15, 392–397. [Google Scholar] [CrossRef]
- Mailleux, F.; Gélinas, R.; Beauloye, C.; Horman, S.; Bertrand, L. O-GlcNAcylation, Enemy or Ally during Cardiac Hypertrophy Development? Biochim. Biophys. Acta 2016, 1862, 2232–2243. [Google Scholar] [CrossRef]
- Brealey, D.; Singer, M. Mitochondrial Dysfunction in Sepsis. Curr. Infect. Dis. Rep. 2003, 5, 365–371. [Google Scholar] [CrossRef]
- Zara, V.; De Benedittis, R.; Ragan, C.I.; Palmieri, F. Immunological Characterization of the Mitochondrial 2-Oxoglutarate Carrier from Liver and Heart. Organ Specificity. FEBS Lett. 1990, 263, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Cappello, A.R.; Miniero, D.V.; Curcio, R.; Ludovico, A.; Daddabbo, L.; Stipani, I.; Robinson, A.J.; Kunji, E.R.S.; Palmieri, F. Functional and Structural Role of Amino Acid Residues in the Odd-Numbered Transmembrane Alpha-Helices of the Bovine Mitochondrial Oxoglutarate Carrier. J. Mol. Biol. 2007, 369, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, M.L.; Joseph, J.W.; Jensen, M.V.; Lu, D.; Ilkayeva, O.; Ronnebaum, S.M.; Becker, T.C.; Newgard, C.B. The Mitochondrial 2-Oxoglutarate Carrier Is Part of a Metabolic Pathway That Mediates Glucose- and Glutamine-Stimulated Insulin Secretion. J. Biol. Chem. 2010, 285, 16530–16537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, M.; Park, D.; Luciani, D.S.; Kida, K.; Palmieri, F.; Blacque, O.E.; Johnson, J.D.; Riddle, D.L. MISC-1/OGC Links Mitochondrial Metabolism, Apoptosis and Insulin Secretion. PLoS ONE 2011, 6, e17827. [Google Scholar] [CrossRef] [PubMed]
- Champattanachai, V.; Marchase, R.B.; Chatham, J.C. Glucosamine Protects Neonatal Cardiomyocytes from Ischemia-Reperfusion Injury via Increased Protein-Associated O-GlcNAc. Am. J. Physiol.-Cell Physiol. 2007, 292, C178–C187. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.P.; Zachara, N.E.; Ngoh, G.A.; Hill, B.G.; Teshima, Y.; Bhatnagar, A.; Hart, G.W.; Marbán, E. Cardioprotection by N-Acetylglucosamine Linkage to Cellular Proteins. Circulation 2008, 117, 1172–1182. [Google Scholar] [CrossRef] [Green Version]
- Verschueren, K.H.G.; Blanchet, C.; Felix, J.; Dansercoer, A.; De Vos, D.; Bloch, Y.; Van Beeumen, J.; Svergun, D.; Gutsche, I.; Savvides, S.N.; et al. Structure of ATP Citrate Lyase and the Origin of Citrate Synthase in the Krebs Cycle. Nature 2019, 568, 571–575. [Google Scholar] [CrossRef]
- Zhao, S.; Torres, A.; Henry, R.A.; Trefely, S.; Wallace, M.; Lee, J.V.; Carrer, A.; Sengupta, A.; Campbell, S.L.; Kuo, Y.-M.; et al. ATP-Citrate Lyase Controls a Glucose-to-Acetate Metabolic Switch. Cell Rep. 2016, 17, 1037–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langston, P.K.; Nambu, A.; Jung, J.; Shibata, M.; Aksoylar, H.I.; Lei, J.; Xu, P.; Doan, M.T.; Jiang, H.; MacArthur, M.R.; et al. Glycerol Phosphate Shuttle Enzyme GPD2 Regulates Macrophage Inflammatory Responses. Nat. Immunol. 2019, 20, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Iacobazzi, V.; Palmieri, F.; Menga, A. ATP-Citrate Lyase Is Essential for Macrophage Inflammatory Response. Biochem. Biophys. Res. Commun. 2013, 440, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhu, X.; Cui, Y.; Miao, H.; Xu, Y.; Xiong, X.; Tang, X.; Shao, L.; Zhang, Y. Serum Proteome-Wide Identified ATP Citrate Lyase as a Novel Informative Diagnostic and Prognostic Biomarker in Pediatric Sepsis: A Pilot Study. Immun. Inflamm. Dis. 2020. [Google Scholar] [CrossRef]
- Macauley, M.S.; Whitworth, G.E.; Debowski, A.W.; Chin, D.; Vocadlo, D.J. O-GlcNAcase Uses Substrate-Assisted Catalysis: Kinetic Analysis and Development of Highly Selective Mechanism-Inspired Inhibitors. J. Biol. Chem. 2005, 280, 25313–25322. [Google Scholar] [CrossRef] [Green Version]
- Merlet, N.; Piriou, N.; Rozec, B.; Grabherr, A.; Lauzier, B.; Trochu, J.-N.; Gauthier, C. Increased Beta2-Adrenoceptors in Doxorubicin-Induced Cardiomyopathy in Rat. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Caloen, G. van; Schmitz, S.; Baroudi, M.E.; Caignet, X.; Ruys, S.P. dit; Roger, P.P.; Vertommen, D.; Machiels, J.-P. Preclinical Activity of Ribociclib in Squamous Cell Carcinoma of the Head and Neck. Mol. Cancer Ther. 2020, 19, 777–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein–Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denis, M.; Dupas, T.; Persello, A.; Dontaine, J.; Bultot, L.; Betus, C.; Pelé, T.; Dhot, J.; Erraud, A.; Maillard, A.; et al. An O-GlcNAcylomic Approach Reveals ACLY as a Potential Target in Sepsis in the Young Rat. Int. J. Mol. Sci. 2021, 22, 9236. https://doi.org/10.3390/ijms22179236
Denis M, Dupas T, Persello A, Dontaine J, Bultot L, Betus C, Pelé T, Dhot J, Erraud A, Maillard A, et al. An O-GlcNAcylomic Approach Reveals ACLY as a Potential Target in Sepsis in the Young Rat. International Journal of Molecular Sciences. 2021; 22(17):9236. https://doi.org/10.3390/ijms22179236
Chicago/Turabian StyleDenis, Manon, Thomas Dupas, Antoine Persello, Justine Dontaine, Laurent Bultot, Charlotte Betus, Thomas Pelé, Justine Dhot, Angélique Erraud, Anaïs Maillard, and et al. 2021. "An O-GlcNAcylomic Approach Reveals ACLY as a Potential Target in Sepsis in the Young Rat" International Journal of Molecular Sciences 22, no. 17: 9236. https://doi.org/10.3390/ijms22179236