
Co-Ultra PEALut Enhances Endogenous Repair Response Following Moderate Traumatic Brain Injury

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Pre-Clinical Results
2.1.1. Co-Ultra PEALut Reduced Neuronal Degeneration in the Hippocampus and Neurological Deficit
2.1.2. Co-Ultra PEALut Appreciably Promotes Cell Proliferation
2.1.3. The Effects of Co-Ultra PEALut Treatment to Prevent Neurodegeneration
2.1.4. Co-Ultra PEALut Treatment Counteracts Neurodegeneration, Slowing down Programmed Cell Death
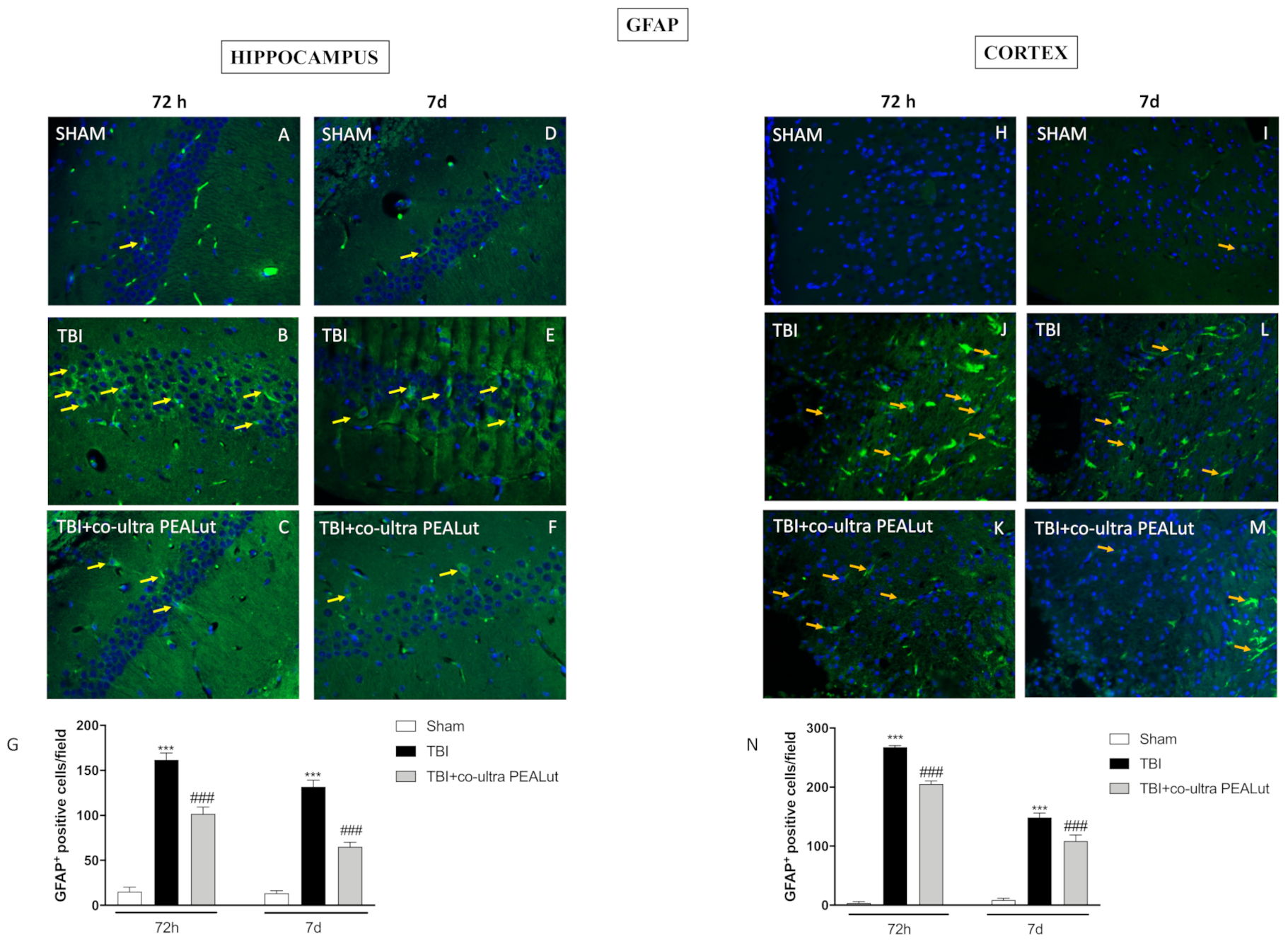
2.1.5. Co-Ultra PEALut Administration Reduced Astrogliosis
2.1.6. Co-Ultra PEALut Administration Reduced Microgliosis
2.1.7. Co-Ultra PEALut Restored BDNF Basal Level
2.1.8. Co-Ultra PEALut Restored NT-3 Basal Level
2.2. Clinical Study Results
2.2.1. Basal Characteristics, Treatment and Neurological Assessment of TBI Patients
2.2.2. Cognitive Outcome
2.2.3. Independency Outcome
2.2.4. Tolerability
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Co-Ultramicronization Process for PEA and Lut
4.3. Pre-Clinical Methods
4.3.1. Animals
4.3.2. Controlled Cortical Impact (CCI) Experimental TBI
4.3.3. Experimental Groups
- Sham group: animals underwent identical surgical procedures except for TBI shock and were maintained under anesthesia for the period of the experiment (n = 10).
- Sham + Co-ultra PEALut group: mice underwent indistinguishable surgical procedures except for TBI shock plus Co-ultra PEALut (1 mg/kg in 1.5% CMC, orally by gavage) administration 1 h after craniotomy and quotidian for 72 h and 7 d (n = 10). (Data not shown)
- TBI + vehicle group: mice underwent CCI (n = 10) plus vehicle, used for co-ultra PEALut, administrated 1 h after craniotomy and quotidian for 72 h and 7 d (n = 10).
- TBI + Co-ultra PEALut group: the mice were subjected to TBI plus administration Co-ultra PEALut (1 mg/kg in 1.5% CMC, orally by gavage) 1 h after craniotomy and quotidian for 72 h and 7 d (n = 10).
4.3.4. Histological Analysis
4.3.5. Immunohistochemistry (IHC)
4.3.6. Behavioral Testing
Morris Water Maze
Rotarod Test
4.3.7. Immunofluorescence of GFAP, Iba1, NeuN, Caspase-3 and Beclin 1
4.3.8. FluoroJade
4.3.9. Western Blot Analysis
4.3.10. Statistical Analysis
4.4. Clinical Methods
4.4.1. Study Setting
4.4.2. Sample Size
4.4.3. Randomization
4.4.4. Primary Endpoint
- (i)
- TBI severity assessment by the Glasgow Coma score and the Marshal Score;
- (ii)
- (iii)
- Depression by the Beck inventory depression scale;
- (iv)
- Independence in activities of daily living and self-care by the Barthel Index.
4.4.5. Blinding
4.4.6. Secondary Endpoints
4.4.7. General Treatment
- Surgical evacuation of hemorrhagic masses and/or “debridement” of outbreaks brain contusion;
- Medical management aimed at the maintenance of euvolemia and adequate brain perfusion. Goals were a systolic blood pressure >90 mmHg and a cerebral perfusion pressure >60 mmHg. Prevention of secondary complications of critical illness included preventive therapy of venous thromboembolism (VTE) using low molecular weight heparin (LMWH) and seizures using levetiracetam prophylactically for the first seven days after injury;
- Patients who underwent surgical treatment also received post-operative intensive care treatments including position of the head high, lower values of end-tidal CO2, sedation with reduced metabolic consumption of O2, increase in plasma osmolarity by administration of mannitol in controlled doses or hypernatremia, therapeutic CSF drainage.
4.4.8. Specific Treatment
4.4.9. Statistical Analysis
5. Patents
- EP 2814489
- EP 2821083
- EP 2985037
- 102015000067344
6. Study Highlights
- The search for molecules that contribute in the neuro-regenerative process has become important, particularly in view of the potential implication to protect the permanent loss of neuron after a head trauma.
- The aim was to appraise the neuro-regenerative properties of a micronized compound comprised of palmitoylethanolamide (PEA) and the vegetable flavonoid luteolin (Lut), which is known as co-ultramicronized PEALut (co-ultra PEALut).
- The co-ultra PEALut treatment helped the neuronal reconstitution process to restore the basal level of both novel neurons and mature neurons in mice
- Co-ultra PEALut could represents a potential therapeutic compound for the improvement of the endogenous reparation response for treating TBI improving the clinical result in traumatic injuries.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethical Approval
Abbreviations
| BDNF | brain-derived neurotrophic factor |
| BNCE | brief neuropsychological cognitive examination |
| BrdU | bromodeoxyuridine |
| CNS | central nervous system |
| CT | computed tomography |
| CCI | controlled cortical impact |
| DG | dentate gyrus |
| DCX | doublecortin |
| ED | emergency department |
| GCS | Glasgow coma scale |
| IRB | institutional review board |
| ICU | intensive care unit |
| ICP | intracranial pressure |
| Lut | luteolin |
| MMSE | mini mental state examination |
| MWM | Morris water maze |
| NPCs | neural precursor cells |
| NSCs | neuronal stem cells |
| NT-3 | neurotrophin-3 |
| NS | not significant |
| PEA | palmitoylethanolamide |
| SVZ | nonventricular zone |
| SGZ | nongranular zone |
| TBI | traumatic brain injury |
| VTE | venous thromboembolism |
References
- Narayan, R.K.; Michel, M.E.; Ansell, B.; Baethmann, A.; Biegon, A.; Bracken, M.B.; Bullock, M.R.; Choi, S.C.; Clifton, G.L.; Contant, C.F.; et al. Clinical trials in head injury. J. Neurotrauma 2002, 19, 503–557. [Google Scholar] [CrossRef]
- Raad, M.; Nohra, E.; Chams, N.; Itani, M.; Talih, F.; Mondello, S.; Kobeissy, F. Autoantibodies in traumatic brain injury and central nervous system trauma. Neuroscience 2014, 281, 16–23. [Google Scholar] [CrossRef]
- Crupi, R.; Cordaro, M.; Cuzzocrea, S.; Impellizzeri, D. Management of Traumatic Brain Injury:From Present to Future. Antioxidants 2020, 9, 297. [Google Scholar] [CrossRef] [Green Version]
- Sophie Su, Y.R.; Veeravagu, A.; Grant, G. Neuroplasticity after Traumatic Brain Injury. In Translational Research in Traumatic Brain Injury; Laskowitz, D., Grant, G., Eds.; CRC Press/Taylor and Francis Group: Boca Raton, FL, USA, 2016. [Google Scholar]
- Kazanis, I. The subependymal zone neurogenic niche: A beating heart in the centre of the brain:how plastic is adult neurogenesis? Opportunities for therapy and questions to be addressed. Brain 2009, 132, 2909–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Cong, D.; Wang, X.; Wang, Y.; Liang, H.; Zhang, X.; Huang, Q. The effect of simvastatin treatment on proliferation and differentiation of neural stem cells after traumatic brain injury. Brain Res. 2015, 1602, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lois, C.; Alvarez-Buylla, A. Proliferating subventricular zone cells in the adult mammalian forebrain can differentiate into neurons and glia. Proc. Natl. Acad. Sci. USA 1993, 90, 2074–2077. [Google Scholar] [CrossRef] [Green Version]
- Denny, C.A.; Kheirbek, M.A.; Alba, E.L.; Tanaka, K.F.; Brachman, R.A.; Laughman, K.B.; Tomm, N.K.; Turi, G.F.; Losonczy, A.; Hen, R. Hippocampal memory traces are differentially modulated by experience, time, and adult neurogenesis. Neuron 2014, 83, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, S.T.; Myczek, K.; Kang, A.P.; Chabrier, M.A.; Baglietto-Vargas, D.; Laferla, F.M. Impact of hippocampal neuronal ablation on neurogenesis and cognition in the aged brain. Neuroscience 2014, 259, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cope, E.C.; Gould, E. Adult Neurogenesis, Glia, and the Extracellular Matrix. Cell Stem Cell 2019, 24, 690–705. [Google Scholar] [CrossRef]
- Manivannan, S.; Marei, O.; Elalfy, O.; Zaben, M. Neurogenesis after traumatic brain injury-The complex role of HMGB1 and neuroinflammation. Neuropharmacology 2021, 183, 108400. [Google Scholar] [CrossRef]
- Redell, J.B.; Maynard, M.E.; Underwood, E.L.; Vita, S.M.; Dash, P.K.; Kobori, N. Traumatic brain injury and hippocampal neurogenesis:Functional implications. Exp. Neurol. 2020, 331, 113372. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Qu, C.; Goussev, A.; Jiang, H.; Lu, C.; Schallert, T.; Mahmood, A.; Chen, J.; Li, Y.; Chopp, M. Statins increase neurogenesis in the dentate gyrus, reduce delayed neuronal death in the hippocampal CA3 region, and improve spatial learning in rat after traumatic brain injury. J. Neurotrauma 2007, 24, 1132–1146. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Ying, Z.; Gomez-Pinilla, F. Omega-3 fatty acids supplementation restores mechanisms that maintain brain homeostasis in traumatic brain injury. J. Neurotrauma 2007, 24, 1587–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muccioli, G.G.; Stella, N. Microglia produce and hydrolyze palmitoylethanolamide. Neuropharmacology 2008, 54, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, L.; Franklin, A.; Witting, A.; Moller, T.; Stella, N. Astrocytes in culture produce anandamide and other acylethanolamides. J. Biol. Chem. 2002, 277, 20869–20876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Crupi, R.; Impellizzeri, D.; Campolo, M.; Marino, A.; Esposito, E.; Cuzzocrea, S. Administration of palmitoylethanolamide (PEA) protects the neurovascular unit and reduces secondary injury after traumatic brain injury in mice. Brain Behav. Immun. 2012, 26, 1310–1321. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Boccella, S.; Iannotta, M.; De Gregorio, D.; Giordano, C.; Belardo, C.; Romano, R.; Palazzo, E.; Scafuro, M.A.; Serra, N.; et al. Palmitoylethanolamide Reduces Neuropsychiatric Behaviors by Restoring Cortical Electrophysiological Activity in a Mouse Model of Mild Traumatic Brain Injury. Front. Pharmacol. 2017, 8, 95. [Google Scholar] [CrossRef] [Green Version]
- Lama, A.; Pirozzi, C.; Annunziata, C.; Morgese, M.G.; Senzacqua, M.; Severi, I.; Calignano, A.; Trabace, L.; Giordano, A.; Meli, R.; et al. Palmitoylethanolamide counteracts brain fog improving depressive-like behaviour in obese mice:Possible role of synaptic plasticity and neurogenesis. Br. J. Pharmacol. 2021, 178, 845–859. [Google Scholar] [CrossRef]
- Fusco, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Gugliandolo, E.; Siracusa, R.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. N-Palmitoylethanolamide-Oxazoline Protects against Middle Cerebral Artery Occlusion Injury in Diabetic Rats by Regulating the SIRT1 Pathway. Int. J. Mol. Sci. 2019, 20, 4845. [Google Scholar] [CrossRef] [Green Version]
- Skaper, S.D.; Buriani, A.; Dal Toso, R.; Petrelli, L.; Romanello, S.; Facci, L.; Leon, A. The ALIAmide palmitoylethanolamide and cannabinoids, but not anandamide, are protective in a delayed postglutamate paradigm of excitotoxic death in cerebellar granule neurons. Proc. Natl. Acad. Sci. USA 1996, 93, 3984–3989. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, C.; Esposito, G.; Blasio, A.; Valenza, M.; Arietti, P.; Steardo, L., Jr.; Carnuccio, R.; De Filippis, D.; Petrosino, S.; Iuvone, T.; et al. Palmitoylethanolamide counteracts reactive astrogliosis induced by beta-amyloid peptide. J. Cell Mol. Med. 2011, 15, 2664–2674. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Chow, M.P.; Huang, W.C.; Lin, Y.C.; Chang, Y.J. Flavonoids inhibit tumor necrosis factor-alpha-induced up-regulation of intercellular adhesion molecule-1 (ICAM-1) in respiratory epithelial cells through activator protein-1 and nuclear factor-kappaB:structure-activity relationships. Mol. Pharmacol. 2004, 66, 683–693. [Google Scholar] [CrossRef]
- Kumazawa, Y.; Kawaguchi, K.; Takimoto, H. Immunomodulating effects of flavonoids on acute and chronic inflammatory responses caused by tumor necrosis factor alpha. Curr. Pharm. Des. 2006, 12, 4271–4279. [Google Scholar] [CrossRef]
- Bhullar, K.S.; Rupasinghe, H.P. Polyphenols:multipotent therapeutic agents in neurodegenerative diseases. Oxid. Med. Cell. Longev. 2013, 2013, 891748. [Google Scholar] [CrossRef] [Green Version]
- Patil, S.P.; Jain, P.D.; Sancheti, J.S.; Ghumatkar, P.J.; Tambe, R.; Sathaye, S. Neuroprotective and neurotrophic effects of Apigenin and Luteolin in MPTP induced parkinsonism in mice. Neuropharmacology 2014, 86, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Crupi, R.; Impellizzeri, D.; Bruschetta, G.; Cordaro, M.; Paterniti, I.; Siracusa, R.; Cuzzocrea, S.; Esposito, E. Co-Ultramicronized Palmitoylethanolamide/Luteolin Promotes Neuronal Regeneration after Spinal Cord Injury. Front. Pharmacol. 2016, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempuraj, D.; Thangavel, R.; Kempuraj, D.D.; Ahmed, M.E.; Selvakumar, G.P.; Raikwar, S.P.; Zaheer, S.A.; Iyer, S.S.; Govindarajan, R.; Chandrasekaran, P.N.; et al. Neuroprotective effects of flavone luteolin in neuroinflammation and neurotrauma. Biofactors 2021, 47, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, H.; Ding, K.; Zhang, L.; Wang, C.; Li, T.; Wei, W.; Lu, X. Luteolin provides neuroprotection in models of traumatic brain injury via the Nrf2-ARE pathway. Free Radic. Biol. Med. 2014, 71, 186–195. [Google Scholar] [CrossRef]
- Yoo, D.Y.; Choi, J.H.; Kim, W.; Nam, S.M.; Jung, H.Y.; Kim, J.H.; Won, M.H.; Yoon, Y.S.; Hwang, I.K. Effects of luteolin on spatial memory, cell proliferation, and neuroblast differentiation in the hippocampal dentate gyrus in a scopolamine-induced amnesia model. Neurol. Res. 2013, 35, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Yao-Yue, C.; Qin, G.W.; Guo, L.H. Luteolin from Purple Perilla mitigates ROS insult particularly in primary neurons. Neurobiol. Aging 2012, 33, 176–186. [Google Scholar] [CrossRef]
- Paterniti, I.; Impellizzeri, D.; Di Paola, R.; Navarra, M.; Cuzzocrea, S.; Esposito, E. A new co-ultramicronized composite including palmitoylethanolamide and luteolin to prevent neuroinflammation in spinal cord injury. J. Neuroinflamm. 2013, 10, 91. [Google Scholar] [CrossRef] [Green Version]
- Paterniti, I.; Cordaro, M.; Campolo, M.; Siracusa, R.; Cornelius, C.; Navarra, M.; Cuzzocrea, S.; Esposito, E. Neuroprotection by association of palmitoylethanolamide with luteolin in experimental Alzheimer’s disease models:the control of neuroinflammation. CNS Neurol. Disord. Drug Targets 2014, 13, 1530–1541. [Google Scholar] [CrossRef]
- Siracusa, R.; Paterniti, I.; Impellizzeri, D.; Cordaro, M.; Crupi, R.; Navarra, M.; Cuzzocrea, S.; Esposito, E. The association of palmitoylethanolamide with luteolin decreases neuroinflammation and stimulates autophagy in Parkinson’s disease model. CNS Neurol. Disord. Drug Targets 2015, 14, 1350–1366. [Google Scholar] [CrossRef]
- Cordaro, M.; Impellizzeri, D.; Paterniti, I.; Bruschetta, G.; Siracusa, R.; De Stefano, D.; Cuzzocrea, S.; Esposito, E. Neuroprotective Effects of Co-UltraPEALut on Secondary Inflammatory Process and Autophagy Involved in Traumatic Brain Injury. J. Neurotrauma 2016, 33, 132–146. [Google Scholar] [CrossRef]
- Siracusa, R.; Paterniti, I.; Bruschetta, G.; Cordaro, M.; Impellizzeri, D.; Crupi, R.; Cuzzocrea, S.; Esposito, E. The Association of Palmitoylethanolamide with Luteolin Decreases Autophagy in Spinal Cord Injury. Mol. Neurobiol. 2016, 53, 3783–3792. [Google Scholar] [CrossRef] [Green Version]
- Girgis, F.; Pace, J.; Sweet, J.; Miller, J.P. Hippocampal Neurophysiologic Changes after Mild Traumatic Brain Injury and Potential Neuromodulation Treatment Approaches. Front. Syst. Neurosci. 2016, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, I.L.; Gonzalez-Prieto, M.; Garcia-Bueno, B.; Caso, J.R.; Leza, J.C.; Madrigal, J.L.M. Alternative Method to Detect Neuronal Degeneration and Amyloid beta Accumulation in Free-Floating Brain Sections with Fluoro-Jade. ASN Neuro 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.J.; Miller, K.M.; Fugaccia, I.; Scheff, S.W. Regional distribution of fluoro-jade B staining in the hippocampus following traumatic brain injury. Exp. Neurol. 2005, 193, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Chang, E.; Igarashi, T.; Noble, L.J. Neuronal injury and loss after traumatic brain injury:time course and regional variability. Brain Res. 2001, 917, 45–54. [Google Scholar] [CrossRef]
- Chirumamilla, S.; Sun, D.; Bullock, M.R.; Colello, R.J. Traumatic brain injury induced cell proliferation in the adult mammalian central nervous system. J. Neurotrauma 2002, 19, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Selimi, F.; Lohof, A.M.; Heitz, S.; Lalouette, A.; Jarvis, C.I.; Bailly, Y.; Mariani, J. Lurcher GRID2-induced death and depolarization can be dissociated in cerebellar Purkinje cells. Neuron 2003, 37, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Erlich, S.; Shohami, E.; Pinkas-Kramarski, R. Neurodegeneration induces upregulation of Beclin 1. Autophagy 2006, 2, 49–51. [Google Scholar] [CrossRef] [Green Version]
- Lossi, L.; Castagna, C.; Merighi, A. Caspase-3 Mediated Cell Death in the Normal Development of the Mammalian Cerebellum. Int. J. Mol. Sci. 2018, 19, 3999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Guise, E.; Gosselin, N.; Leblanc, J.; Champoux, M.C.; Couturier, C.; Lamoureux, J.; Dagher, J.; Marcoux, J.; Maleki, M.; Feyz, M. Clock drawing and mini-mental state examination in patients with traumatic brain injury. Appl. Neuropsychol. 2011, 18, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Kernie, S.G.; Parent, J.M. Forebrain neurogenesis after focal Ischemic and traumatic brain injury. Neurobiol. Dis. 2010, 37, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Rolfe, A.; Sun, D. Stem Cell Therapy in Brain Trauma:Implications for Repair and Regeneration of Injured Brain in Experimental TBI Models. In Brain Neurotrauma: Molecular, Neuropsychological, and Rehabilitation Aspects; Kobeissy, F.H.P., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2015. [Google Scholar]
- Sun, D.; Colello, R.J.; Daugherty, W.P.; Kwon, T.H.; McGinn, M.J.; Harvey, H.B.; Bullock, M.R. Cell proliferation and neuronal differentiation in the dentate gyrus in juvenile and adult rats following traumatic brain injury. J. Neurotrauma 2005, 22, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Rola, R.; Mizumatsu, S.; Otsuka, S.; Morhardt, D.R.; Noble-Haeusslein, L.J.; Fishman, K.; Potts, M.B.; Fike, J.R. Alterations in hippocampal neurogenesis following traumatic brain injury in mice. Exp. Neurol. 2006, 202, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Cuzzocrea, S. Palmitoylethanolamide in homeostatic and traumatic central nervous system injuries. CNS Neurol. Disord. Drug Targets 2013, 12, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Impellizzeri, D.; Mazzon, E.; Paterniti, I.; Cuzzocrea, S. Neuroprotective activities of palmitoylethanolamide in an animal model of Parkinson’s disease. PLoS ONE 2012, 7, e41880. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare Mannelli, L.; Pacini, A.; Corti, F.; Boccella, S.; Luongo, L.; Esposito, E.; Cuzzocrea, S.; Maione, S.; Calignano, A.; Ghelardini, C. Antineuropathic profile of N-palmitoylethanolamine in a rat model of oxaliplatin-induced neurotoxicity. PLoS ONE 2015, 10, e0128080. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ren, T.; Xu, L.; Wang, Y.; Yang, B.; Luo, H.; Zeng, Z.; Zhang, Y.; Du, G.; Zhu, M.; et al. Propane-2-sulfonic acid octadec-9-enyl-amide, a novel peroxisome proliferator-activated receptors alpha and gamma dual agonist, enhances hippocampal neurogenesis and neuroplasticity in rats with cerebral ischaemia. Neuroreport 2019, 30, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Fidaleo, M.; Fanelli, F.; Ceru, M.P.; Moreno, S. Neuroprotective properties of peroxisome proliferator-activated receptor alpha (PPARalpha) and its lipid ligands. Curr. Med. Chem. 2014, 21, 2803–2821. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.F.; Gu, S.; Shan, C.; Marchado, S.; Arias-Carrion, O. Oxidative Stress and Adult Neurogenesis. Stem Cell Rev. Rep. 2015, 11, 706–709. [Google Scholar] [CrossRef]
- Kempermann, G.; Kuhn, H.G.; Gage, F.H. Experience-induced neurogenesis in the senescent dentate gyrus. J. Neurosci. 1998, 18, 3206–3212. [Google Scholar] [CrossRef] [Green Version]
- Neuberger, E.J.; Swietek, B.; Corrubia, L.; Prasanna, A.; Santhakumar, V. Enhanced Dentate Neurogenesis after Brain Injury Undermines Long-Term Neurogenic Potential and Promotes Seizure Susceptibility. Stem Cell Rep. 2017, 9, 972–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; ZhuGe, Q.; Zhong, M.; Chen, G.; Shao, B.; Wang, H.; Mao, X.; Xie, L.; Jin, K. Neurogenesis in adult human brain after traumatic brain injury. J. Neurotrauma 2013, 30, 1872–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Gao, X.; Michalski, S.; Zhao, S.; Chen, J. Traumatic Brain Injury Severity Affects Neurogenesis in Adult Mouse Hippocampus. J. Neurotrauma 2016, 33, 721–733. [Google Scholar] [CrossRef]
- Kernie, S.G.; Erwin, T.M.; Parada, L.F. Brain remodeling due to neuronal and astrocytic proliferation after controlled cortical injury in mice. J. Neurosci. Res. 2001, 66, 317–326. [Google Scholar] [CrossRef]
- Mueller, F.J.; McKercher, S.R.; Imitola, J.; Loring, J.F.; Yip, S.; Khoury, S.J.; Snyder, E.Y. At the interface of the immune system and the nervous system:how neuroinflammation modulates the fate of neural progenitors in vivo. In Opportunities and Challenges of the Therapies Targeting CNS Regeneration; Ernst Schering Research Foundation Workshop, Springer: Berlin, Germany, 2005; Volume 53, pp. 83–114. [Google Scholar] [CrossRef]
- Wilhelmsson, U.; Pozo-Rodrigalvarez, A.; Kalm, M.; de Pablo, Y.; Widestrand, A.; Pekna, M.; Pekny, M. The role of GFAP and vimentin in learning and memory. Biol. Chem. 2019, 400, 1147–1156. [Google Scholar] [CrossRef]
- Skaper, S.D.; Facci, L.; Giusti, P. Mast cells, glia and neuroinflammation:partners in crime? Immunology 2014, 141, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Di Marzo, V. The pharmacology of palmitoylethanolamide and first data on the therapeutic efficacy of some of its new formulations. Br. J. Pharmacol. 2017, 174, 1349–1365. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Fusco, R.; Siracusa, R.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; Di Paola, R. Neuroprotective Effect of Artesunate in Experimental Model of Traumatic Brain Injury. Front. Neurol. 2018, 9, 590. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Miao, Z.; Ye, Y.; Zhao, P.; Fan, L.; Bao, Z.; Tu, Y.; Li, C.; Chao, H.; Xu, X.; et al. Curcumin alleviates neuroinflammation, enhances hippocampal neurogenesis, and improves spatial memory after traumatic brain injury. Brain Res. Bull. 2020, 162, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins:roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazquez, C.; Chiarlone, A.; Bellocchio, L.; Resel, E.; Pruunsild, P.; Garcia-Rincon, D.; Sendtner, M.; Timmusk, T.; Lutz, B.; Galve-Roperh, I.; et al. The CB(1) cannabinoid receptor signals striatal neuroprotection via a PI3K/Akt/mTORC1/BDNF pathway. Cell Death Differ. 2015, 22, 1618–1629. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des. Devel. Ther. 2016, 10, 23–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caltagirone, C.; Cisari, C.; Schievano, C.; Di Paola, R.; Cordaro, M.; Bruschetta, G.; Esposito, E.; Cuzzocrea, S.; Stroke Study, G. Co-ultramicronized Palmitoylethanolamide/Luteolin in the Treatment of Cerebral Ischemia:from Rodent to Man. Transl. Stroke Res. 2016, 7, 54–69. [Google Scholar] [CrossRef] [Green Version]
- van Zomeren, A.H.; van den Burg, W. Residual complaints of patients two years after severe head injury. J. Neurol. Neurosurg. Psychiatry 1985, 48, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Brooks, D.N.; McKinlay, W. Personality and behavioural change after severe blunt head injury—A relative’s view. J. Neurol. Neurosurg. Psychiatry 1983, 46, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Campolo, M.; Esposito, E.; Ahmad, A.; Di Paola, R.; Paterniti, I.; Cordaro, M.; Bruschetta, G.; Wallace, J.L.; Cuzzocrea, S. Hydrogen sulfide-releasing cyclooxygenase inhibitor ATB-346 enhances motor function and reduces cortical lesion volume following traumatic brain injury in mice. J. Neuroinflamm. 2014, 11, 196. [Google Scholar] [CrossRef] [Green Version]
- Campolo, M.; Esposito, E.; Cuzzocrea, S. A Controlled Cortical Impact Preclinical Model of Traumatic Brain Injury. Methods Mol. Biol. 2018, 1727, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Farr, S.A.; Cuzzocrea, S.; Esposito, E.; Campolo, M.; Niehoff, M.L.; Doyle, T.M.; Salvemini, D. Adenosine A3 receptor as a novel therapeutic target to reduce secondary events and improve neurocognitive functions following traumatic brain injury. J. Neuroinflamm. 2020, 17, 339. [Google Scholar] [CrossRef]
- Siracusa, R.; Impellizzeri, D.; Cordaro, M.; Crupi, R.; Esposito, E.; Petrosino, S.; Cuzzocrea, S. Anti-Inflammatory and Neuroprotective Effects of Co-UltraPEALut in a Mouse Model of Vascular Dementia. Front. Neurol. 2017, 8, 233. [Google Scholar] [CrossRef]
- Casili, G.; Campolo, M.; Paterniti, I.; Lanza, M.; Filippone, A.; Cuzzocrea, S.; Esposito, E. Dimethyl Fumarate Attenuates Neuroinflammation and Neurobehavioral Deficits Induced by Experimental Traumatic Brain Injury. J. Neurotrauma 2018, 35, 1437–1451. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Casili, G.; Paterniti, I.; Filippone, A.; Lanza, M.; Ardizzone, A.; Scuderi, S.A.; Cuzzocrea, S.; Esposito, E. Effect of a Product Containing Xyloglucan and Pea Protein on a Murine Model of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 3596. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.; Chimirri, S.; Aiello, R.; De Fazio, S.; Leo, A.; Rispoli, V.; Marra, R.; Labate, A.; De Fazio, P.; Citraro, R.; et al. Lamotrigine positively affects the development of psychiatric comorbidity in epileptic animals, while psychiatric comorbidity aggravates seizures. Epilepsy Behav. 2013, 28, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Vorhees, C.V.; Williams, M.T. Morris water maze:procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Rakoczy, S.; Brown-Borg, H. Assessment of spatial memory in mice. Life Sci. 2010, 87, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Crupi, R.; Campolo, M.; Genovese, T.; Esposito, E.; Cuzzocrea, S. Absence of TLR4 reduces neurovascular unit and secondary inflammatory process after traumatic brain injury in mice. PLoS ONE 2013, 8, e57208. [Google Scholar] [CrossRef] [Green Version]
- Filippone, A.; Consoli, G.M.L.; Granata, G.; Casili, G.; Lanza, M.; Ardizzone, A.; Cuzzocrea, S.; Esposito, E.; Paterniti, I. Topical Delivery of Curcumin by Choline-Calix[4]arene-Based Nanohydrogel Improves Its Therapeutic Effect on a Psoriasis Mouse Model. Int. J. Mol. Sci. 2020, 21, 5053. [Google Scholar] [CrossRef] [PubMed]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Ball, T.D.; Pastore, R.E.; Sollman, M.J.; Burright, R.G.; Donovick, P.J. Evaluation of a neuropsychological screen in an incarcerated population. Clin. Neuropsychol. 2009, 23, 1037–1049. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | ||||||||
|---|---|---|---|---|---|---|---|---|
| Educational Level | 15–20 | 21–30 | 31–40 | 41–50 | 51–60 | 61–70 | 71–80 | >80 |
| Low | 79 | 72 | 73 | 65 | 63 | 59 | 52 | 44 |
| High | 74 | 82 | 77 | 77 | 73 | 61 | 59 | 45 |
| Study Group (Glialia) | Control Group | P | |
|---|---|---|---|
| N. of Patients | 15 | 15 | NS |
| Gender | 13M/2F | 11M/4F | NS |
| Age | 49.7 ± 17.9 | 53.8 ± 17.5 | NS |
| GCS at admission | 11.2 ± 1.3 | 11.2 ± 1.4 | NS |
| MMSE score | 21.1 ± 4.6 | 21.5 ± 3.1 | NS |
| BNCE | 52.3 ± 17.5% | 47.1 ± 19.3% | NS |
| Marshall Scale I II III IV V VI | - 6 3 4 1 1 | - 6 4 3 2 - | NS |
| Surgical Treatment | 4 | 3 | NS |
| OUTCOME | Study Group | Control Group | P |
|---|---|---|---|
| MMSE score | 28.1 ± 2.4 | 26.1 ± 2.8 | 0.04 |
| Δ MMSE | 7.4 ± 1.7 | 4.6 ± 3.9 | 0.02 |
| MMSE score > 26 | 15/15 | 9/15 | 0.02 |
| BNCE | 73.4 ± 11.2% | 63 ± 24.9% | 0.13 |
| BCNE Interference Memory | 85.7 ± 25.2% | 51.9 ± 30.2% | 0.03 |
| Barthel Index | 98.7 ± 3.6 | 97 ± 5.9 | 0.27 |
| Becks Depression Inventory | 5.2 ± 3.7 | 9.9 ± 6.7 | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campolo, M.; Crupi, R.; Cordaro, M.; Cardali, S.M.; Ardizzone, A.; Casili, G.; Scuderi, S.A.; Siracusa, R.; Esposito, E.; Conti, A.; et al. Co-Ultra PEALut Enhances Endogenous Repair Response Following Moderate Traumatic Brain Injury. Int. J. Mol. Sci. 2021, 22, 8717. https://doi.org/10.3390/ijms22168717
Campolo M, Crupi R, Cordaro M, Cardali SM, Ardizzone A, Casili G, Scuderi SA, Siracusa R, Esposito E, Conti A, et al. Co-Ultra PEALut Enhances Endogenous Repair Response Following Moderate Traumatic Brain Injury. International Journal of Molecular Sciences. 2021; 22(16):8717. https://doi.org/10.3390/ijms22168717
Chicago/Turabian StyleCampolo, Michela, Rosalia Crupi, Marika Cordaro, Salvatore Massimo Cardali, Alessio Ardizzone, Giovanna Casili, Sarah Adriana Scuderi, Rosalba Siracusa, Emanuela Esposito, Alfredo Conti, and et al. 2021. "Co-Ultra PEALut Enhances Endogenous Repair Response Following Moderate Traumatic Brain Injury" International Journal of Molecular Sciences 22, no. 16: 8717. https://doi.org/10.3390/ijms22168717