
Targeting Mammalian 5-Lipoxygenase by Dietary Phenolics as an Anti-Inflammatory Mechanism: A Systematic Review

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
1.1. Lipoxygenases
1.2. 5-Lipoxygenase and Inflammation
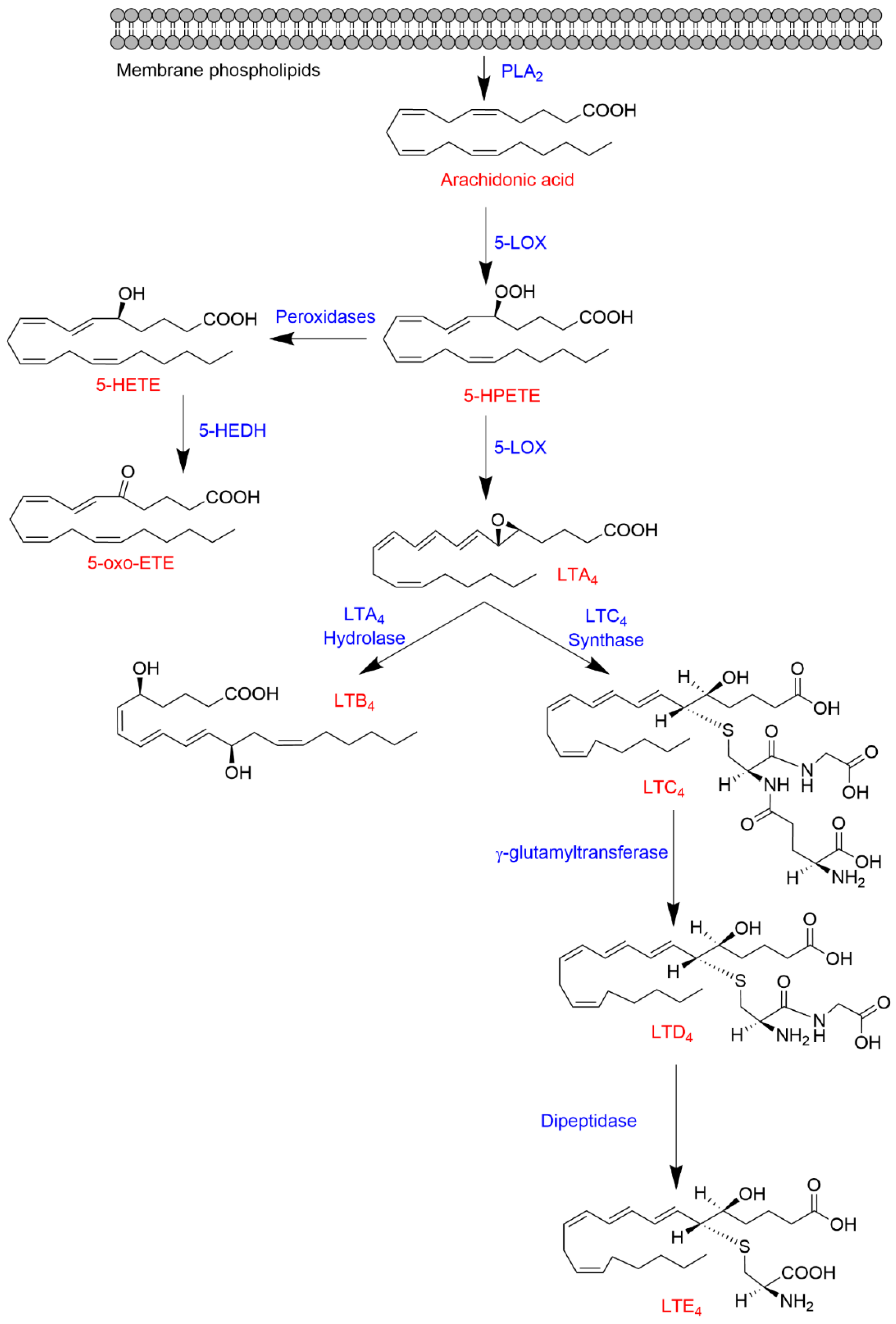
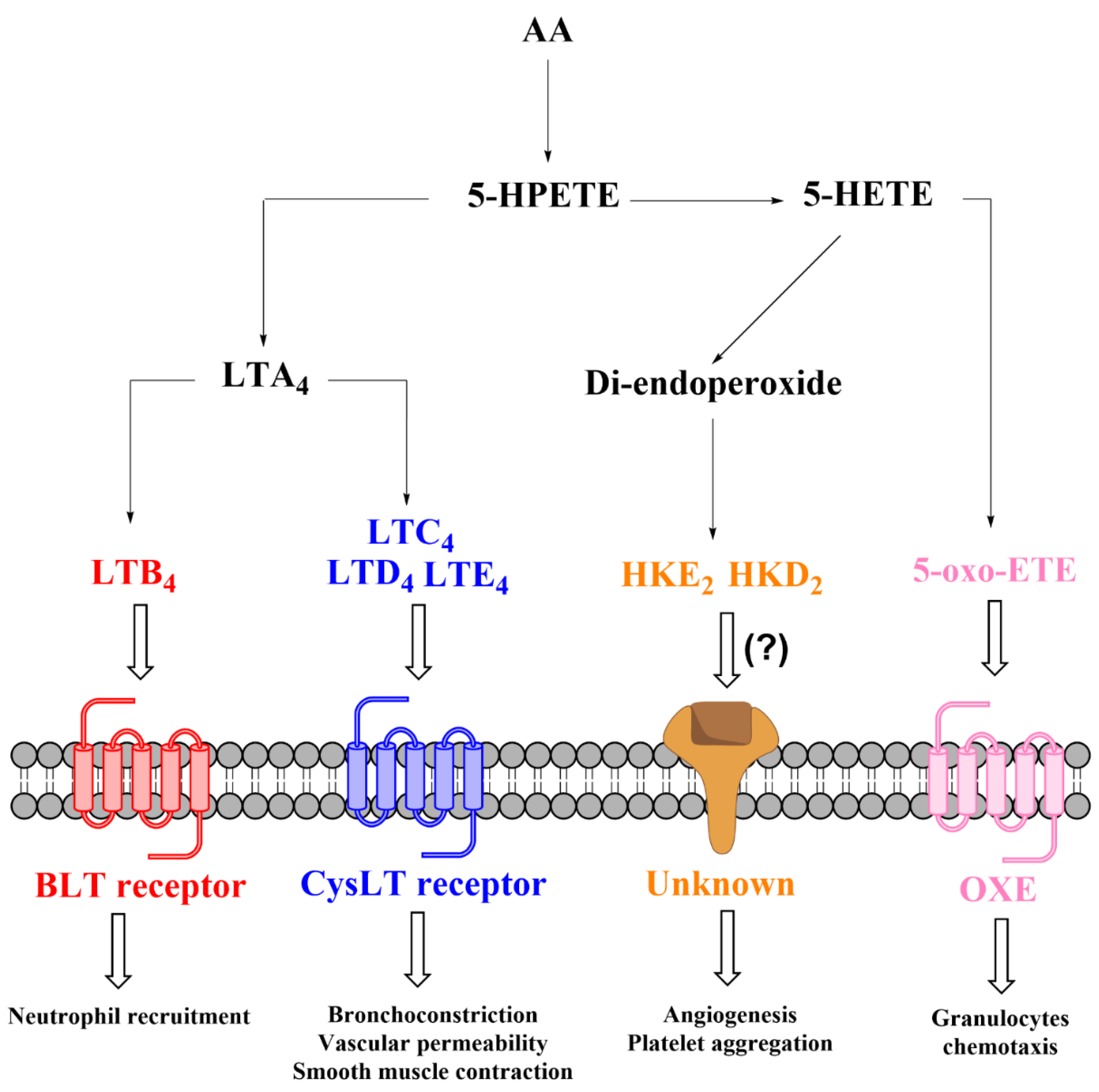
1.3. 5-Lipoxygenase Pathway
1.3.1. Biosynthesis of 5-LOX-Derived Eicosanoids
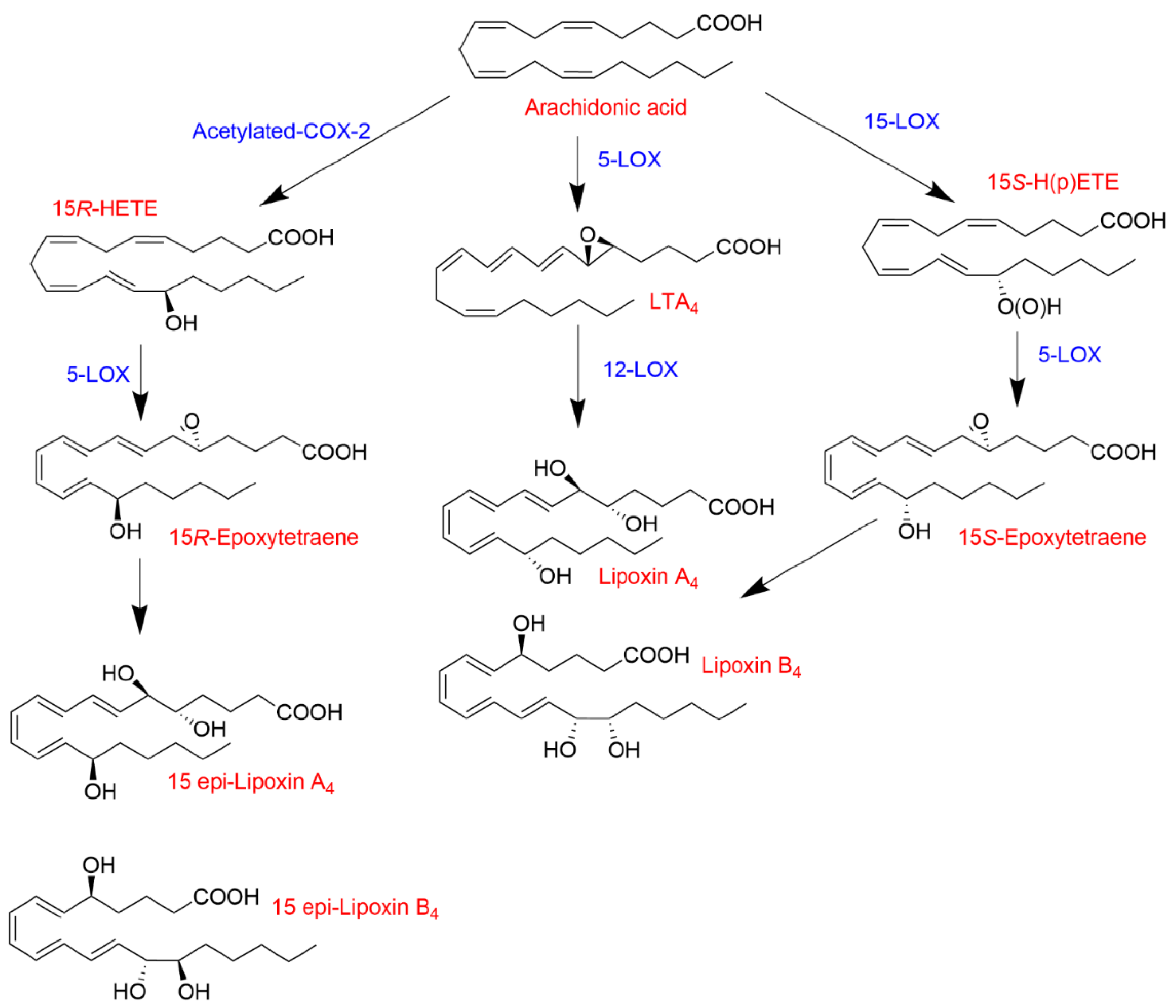
1.3.2. Transcellular Biosynthesis of Leukotrienes and Lipoxins
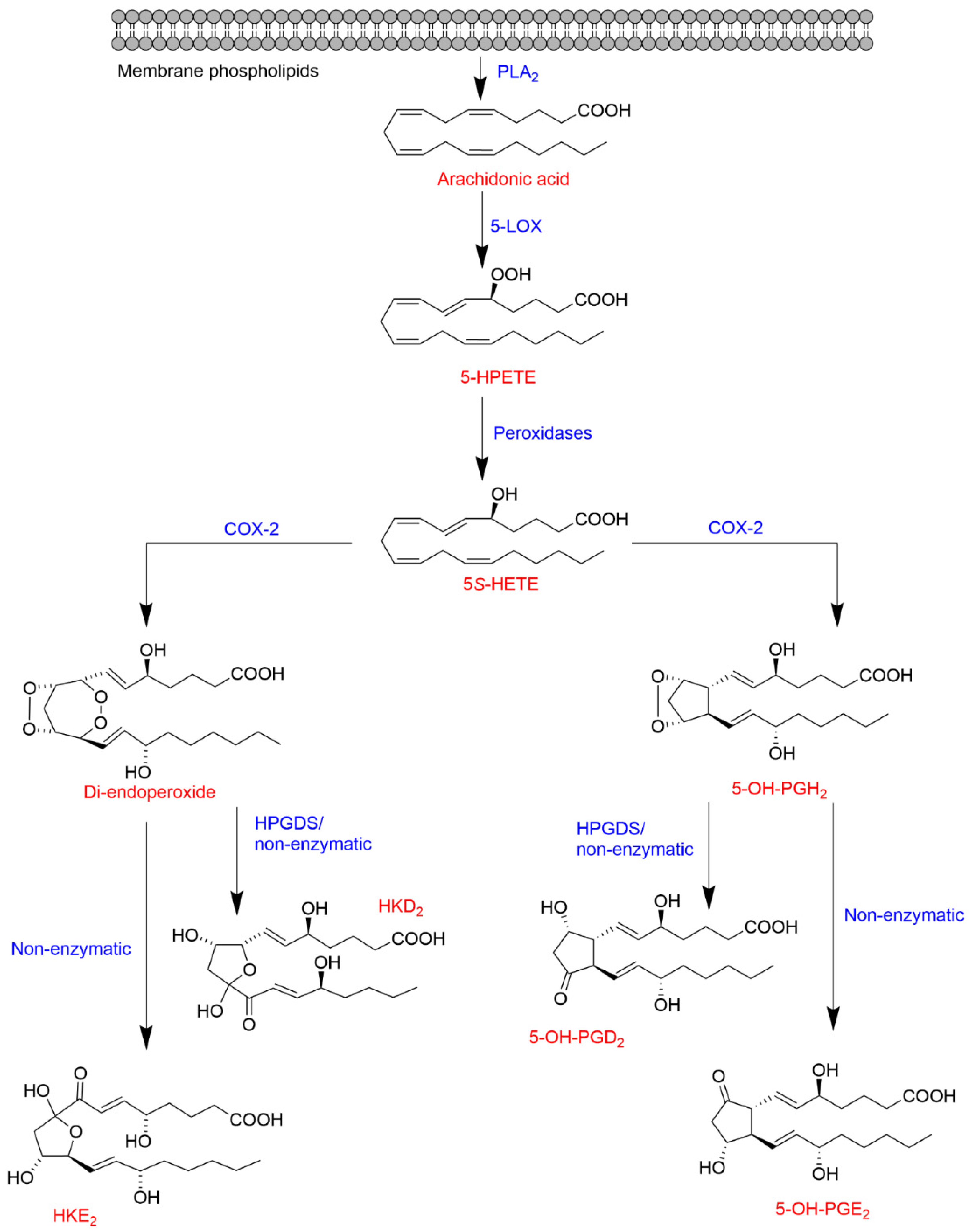
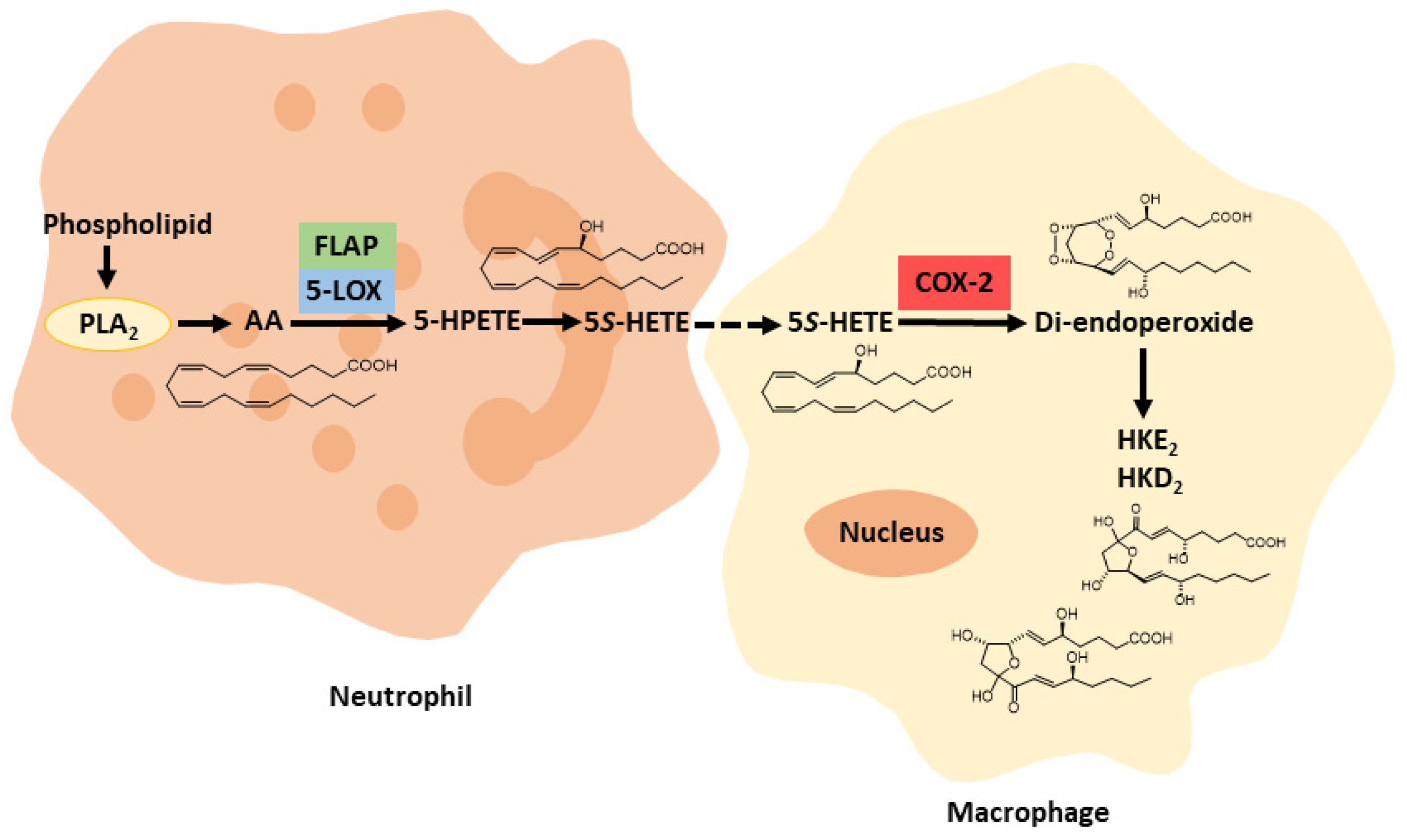
1.3.3. The 5-LOX/COX-2 Crossover Biosynthetic Pathway
1.3.4. Role of 5-Lipoxygenase-Derived Eicosanoids in Inflammation
1.4. Inflammation as a Target of Dietary (Poly)phenols: Role of Their Bioavailability
2. Methods
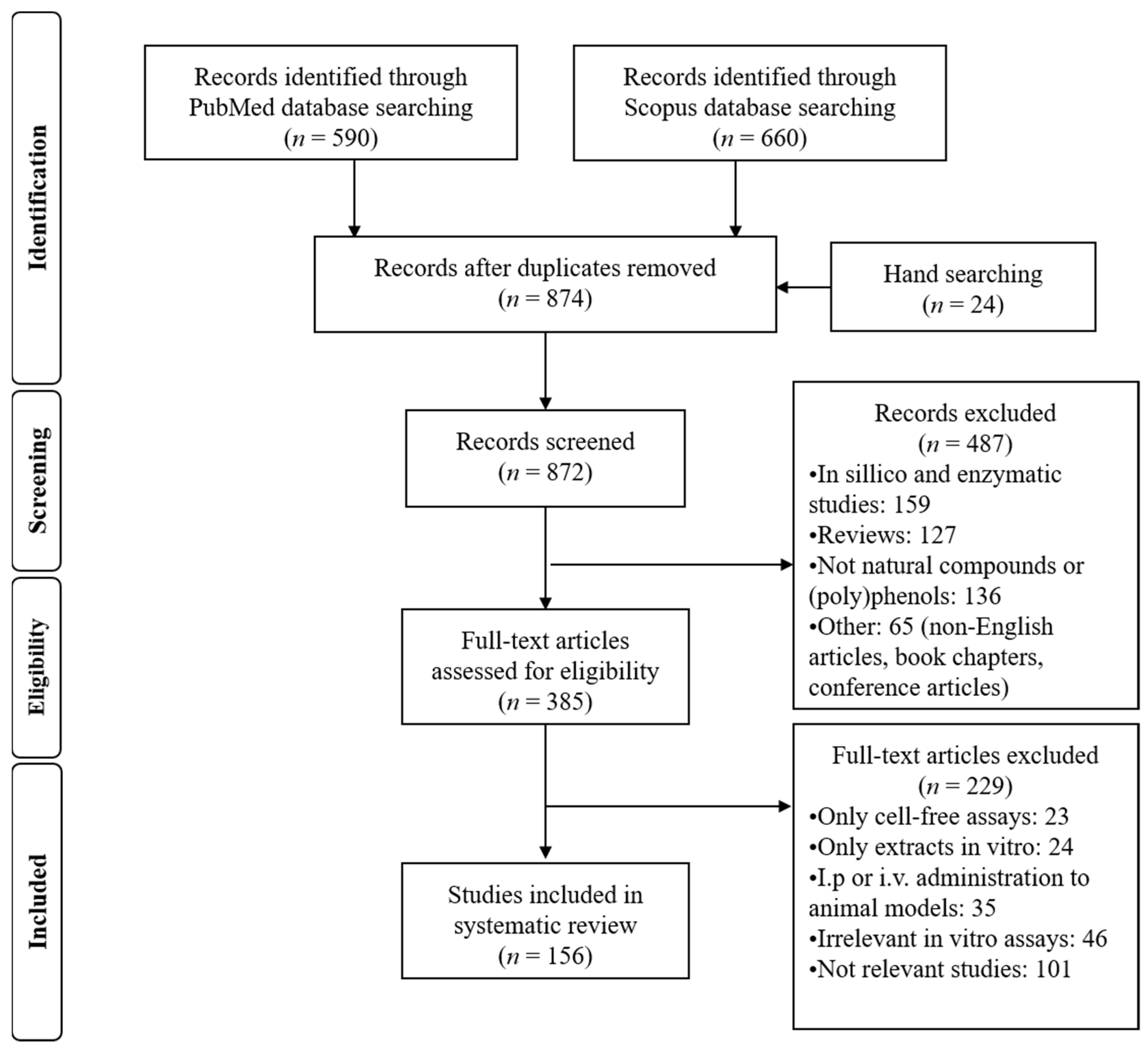
Search Strategy and Study Selection
3. Results and Discussion
3.1. Human Studies
3.2. Animal Studies
3.3. In Vitro Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Brash, A.R. Lipoxygenases: Occurrence, Functions, Catalysis, and Acquisition of Substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Mandal, R.; Singh, A.; Singh, A.P. Legume Lipoxygenase: Strategies for Application in Food Industry. Legum. Sci. 2020, 2, e44. [Google Scholar] [CrossRef]
- Baysal, T.; Demirdöven, A. Lipoxygenase in Fruits and Vegetables: A Review. Enzyme Microb. Technol. 2007, 40, 491–496. [Google Scholar] [CrossRef]
- Ogorodnikova, A.V.; Mukhitova, F.K.; Grechkin, A.N. Oxylipins in the Spikemoss Selaginella Martensii: Detection of Divinyl Ethers, 12-Oxophytodienoic Acid and Related Cyclopentenones. Phytochemistry 2015, 118, 42–50. [Google Scholar] [CrossRef]
- Bennett, M.; Gilroy, D.W. Lipid Mediators in Inflammation. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.R.; O’Neill, G.P.; Liu, Q.; Im, D.S.; Sawyer, N.; Metters, K.M.; Coulombe, N.; Abramovitz, M.; Figueroa, D.J.; Zeng, Z.; et al. Characterization of the Human Cysteinyl Leukotriene CysLT1 Receptor. Nature 1999, 399, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Mashima, R.; Okuyama, T. The Role of Lipoxygenases in Pathophysiology; New Insights and Future Perspectives. Redox Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Ramírez, A.; Mascayano-Collado, C.; Barriga, A.; Echeverría, J.; Urzúa, A. Inhibition of Soybean 15-Lipoxygenase and Human 5-Lipoxygenase by Extracts of Leaves, Stem Bark, Phenols and Catechols Isolated From Lithraea Caustica (Anacardiaceae). Front. Pharmacol. 2020, 11, 594257. [Google Scholar] [CrossRef]
- Kotas, M.E.; Medzhitov, R. Homeostasis, Inflammation, and Disease Susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R. Origin and Physiological Roles of Inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Haeggström, J.Z. Leukotriene Biosynthetic Enzymes as Therapeutic Targets. J. Clin. Investig. 2018, 128, 2680–2690. [Google Scholar] [CrossRef] [PubMed]
- Haeggström, J.Z.; Funk, C.D. Lipoxygenase and Leukotriene Pathways: Biochemistry, Biology, and Roles in Disease. Chem. Rev. 2011, 111, 5866–5898. [Google Scholar] [CrossRef] [PubMed]
- Griesser, M.; Suzuki, T.; Tejera, N.; Mont, S.; Boeglin, W.E.; Pozzi, A.; Schneider, C. Biosynthesis of Hemiketal Eicosanoids by Cross-over of the 5-Lipoxygenase and Cyclooxygenase-2 Pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 6945–6950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. Eicosanoids. Essays Biochem. 2020, 64, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Peters-Golden, M.; Henderson, W.R. Leukotrienes. N. Engl. J. Med. 2007, 357, 1841–1854. [Google Scholar] [CrossRef] [PubMed]
- Bruno, F.; Spaziano, G.; Liparulo, A.; Roviezzo, F.; Nabavi, S.M.; Sureda, A.; Filosa, R.; D’Agostino, B. Recent Advances in the Search for Novel 5-Lipoxygenase Inhibitors for the Treatment of Asthma. Eur. J. Med. Chem. 2018, 153, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Poff, C.D.; Balazy, M. Drugs That Target Lipoxygenases and Leukotrienes as Emerging Therapies for Asthma and Cancer. Curr. Drug Targets Inflamm. Allergy 2004, 3, 19–33. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Jia, Z.; Battino, M.; Miron, A.; Yu, Z.; Cao, H.; Xiao, J. Intracellular Signaling Pathways of Inflammation Modulated by Dietary Flavonoids: The Most Recent Evidence. Crit. Rev. Food Sci. Nutr. 2018, 58, 2908–2924. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Borgeat, P.; Samuelsson, B. Transformation of Arachidonic Acid by Rabbit Polymorphonuclear Leukocytes. Formation of a Novel Dihydroxyeicosatetraenoic Acid. J. Biol. Chem. 1979, 254, 2643–2646. [Google Scholar] [CrossRef]
- Borgeat, P.; Samuelsson, B. Arachidonic Acid Metabolism in Polymorphonuclear Leukocytes: Unstable Intermediate in Formation of Dihydroxy Acids. Proc. Natl. Acad. Sci. USA 1979, 76, 3213–3217. [Google Scholar] [CrossRef] [Green Version]
- Norris, P.C.; Gosselin, D.; Reichart, D.; Glass, C.K.; Dennis, E.A. Phospholipase A2 Regulates Eicosanoid Class Switching during Inflammasome Activation. Proc. Natl. Acad. Sci. USA 2014, 111, 12746–12751. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.-Y.; Shelhamer, J.H. Toll-like Receptor 4 Signaling Regulates Cytosolic Phospholipase A2 Activation and Lipid Generation in Lipopolysaccharide-Stimulated Macrophages. J. Biol. Chem. 2005, 280, 38969–38975. [Google Scholar] [CrossRef] [Green Version]
- Martinez Molina, D.; Wetterholm, A.; Kohl, A.; McCarthy, A.A.; Niegowski, D.; Ohlson, E.; Hammarberg, T.; Eshaghi, S.; Haeggström, J.Z.; Nordlund, P. Structural Basis for Synthesis of Inflammatory Mediators by Human Leukotriene C4 Synthase. Nature 2007, 448, 613–616. [Google Scholar] [CrossRef]
- Murphy, R.C.; Gijón, M.A. Biosynthesis and Metabolism of Leukotrienes. Biochem. J. 2007, 405, 379–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, W.S.; Gravelle, F.; Gravel, S. Metabolism of 5(S)-Hydroxy-6,8,11,14-Eicosatetraenoic Acid and Other 5(S)-Hydroxyeicosanoids by a Specific Dehydrogenase in Human Polymorphonuclear Leukocytes. J. Biol. Chem. 1992, 267, 19233–19241. [Google Scholar] [CrossRef]
- Sala, A.; Folco, G.; Murphy, R.C. Transcellular Biosynthesis of Eicosanoids. Pharmacol. Rep. PR 2010, 62, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Feinmark, S.J.; Cannon, P.J. Endothelial Cell Leukotriene C4 Synthesis Results from Intercellular Transfer of Leukotriene A4 Synthesized by Polymorphonuclear Leukocytes. J. Biol. Chem. 1986, 261, 16466–16472. [Google Scholar] [CrossRef]
- McGee, J.E.; Fitzpatrick, F.A. Erythrocyte-Neutrophil Interactions: Formation of Leukotriene B4 by Transcellular Biosynthesis. Proc. Natl. Acad. Sci. USA 1986, 83, 1349–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabre, J.-E.; Goulet, J.L.; Riche, E.; Nguyen, M.; Coggins, K.; Offenbacher, S.; Koller, B.H. Transcellular Biosynthesis Contributes to the Production of Leukotrienes during Inflammatory Responses in Vivo. J. Clin. Investig. 2002, 109, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Zarini, S.; Gijón, M.A.; Ransome, A.E.; Murphy, R.C.; Sala, A. Transcellular Biosynthesis of Cysteinyl Leukotrienes in Vivo during Mouse Peritoneal Inflammation. Proc. Natl. Acad. Sci. USA 2009, 106, 8296–8301. [Google Scholar] [CrossRef] [Green Version]
- Folco, G.; Murphy, R.C. Eicosanoid Transcellular Biosynthesis: From Cell-Cell Interactions to in Vivo Tissue Responses. Pharmacol. Rev. 2006, 58, 375–388. [Google Scholar] [CrossRef]
- Edenius, C.; Heidvall, K.; Lindgren, J.A. Novel Transcellular Interaction: Conversion of Granulocyte-Derived Leukotriene A4 to Cysteinyl-Containing Leukotrienes by Human Platelets. Eur. J. Biochem. 1988, 178, 81–86. [Google Scholar] [CrossRef]
- Romano, M.; Serhan, C.N. Lipoxin Generation by Permeabilized Human Platelets. Biochemistry 1992, 31, 8269–8277. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hamberg, M.; Samuelsson, B. Lipoxins: Novel Series of Biologically Active Compounds Formed from Arachidonic Acid in Human Leukocytes. Proc. Natl. Acad. Sci. USA 1984, 81, 5335–5339. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Bastida, J.A.; Boeglin, W.E.; Boutaud, O.; Malkowski, M.G.; Schneider, C. Residual Cyclooxygenase Activity of Aspirin-Acetylated COX-2 Forms 15 R-Prostaglandins That Inhibit Platelet Aggregation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 1033–1041. [Google Scholar] [CrossRef]
- Lecomte, M.; Laneuville, O.; Ji, C.; DeWitt, D.L.; Smith, W.L. Acetylation of Human Prostaglandin Endoperoxide Synthase-2 (Cyclooxygenase-2) by Aspirin. J. Biol. Chem. 1994, 269, 13207–13215. [Google Scholar] [CrossRef]
- Clària, J.; Serhan, C.N. Aspirin Triggers Previously Undescribed Bioactive Eicosanoids by Human Endothelial Cell-Leukocyte Interactions. Proc. Natl. Acad. Sci. USA 1995, 92, 9475–9479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N.; Oliw, E. Unorthodox Routes to Prostanoid Formation: New Twists in Cyclooxygenase-Initiated Pathways. J. Clin. Investig. 2001, 107, 1481–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, N.; Arita, M.; Serhan, C.N. Anti-Inflammatory Circuitry: Lipoxin, Aspirin-Triggered Lipoxins and Their Receptor ALX. Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Ringholz, F.C.; Buchanan, P.J.; Clarke, D.T.; Millar, R.G.; McDermott, M.; Linnane, B.; Harvey, B.J.; McNally, P.; Urbach, V. Reduced 15-Lipoxygenase 2 and Lipoxin A4/Leukotriene B4 Ratio in Children with Cystic Fibrosis. Eur. Respir. J. 2014, 44, 394–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snodgrass, R.G.; Brüne, B. Regulation and Functions of 15-Lipoxygenases in Human Macrophages. Front. Pharmacol. 2019, 10, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.; Boeglin, W.E.; Yin, H.; Stec, D.F.; Voehler, M. Convergent Oxygenation of Arachidonic Acid by 5-Lipoxygenase and Cyclooxygenase-2. J. Am. Chem. Soc. 2006, 128, 720–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, F.; Suzuki, T.; Gordon, O.N.; Golding, D.; Okuno, T.; Giménez-Bastida, J.A.; Yokomizo, T.; Schneider, C. Biosynthetic Crossover of 5-Lipoxygenase and Cyclooxygenase-2 Yields 5-Hydroxy-PGE2 and 5-Hydroxy-PGD2. JACS Au 2021. [Google Scholar] [CrossRef]
- Mulugeta, S.; Suzuki, T.; Hernandez, N.T.; Griesser, M.; Boeglin, W.E.; Schneider, C. Identification and Absolute Configuration of Dihydroxy-Arachidonic Acids Formed by Oxygenation of 5S-HETE by Native and Aspirin-Acetylated COX-2. J. Lipid Res. 2010, 51, 575–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejera, N.; Boeglin, W.E.; Suzuki, T.; Schneider, C. COX-2-Dependent and -Independent Biosynthesis of Dihydroxy-Arachidonic Acids in Activated Human Leukocytes. J. Lipid Res. 2012, 53, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Bastida, J.A.; Shibata, T.; Uchida, K.; Schneider, C. Roles of 5-Lipoxygenase and Cyclooxygenase-2 in the Biosynthesis of Hemiketals E2 and D2 by Activated Human Leukocytes. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 1867–1878. [Google Scholar] [CrossRef] [Green Version]
- Ford-Hutchinson, A.W.; Bray, M.A.; Doig, M.V.; Shipley, M.E.; Smith, M.J. Leukotriene B, a Potent Chemokinetic and Aggregating Substance Released from Polymorphonuclear Leukocytes. Nature 1980, 286, 264–265. [Google Scholar] [CrossRef]
- Chen, X.S.; Sheller, J.R.; Johnson, E.N.; Funk, C.D. Role of Leukotrienes Revealed by Targeted Disruption of the 5-Lipoxygenase Gene. Nature 1994, 372, 179–182. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid Storm in Infection and Inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, F.; Yokomizo, T. The Leukotriene Receptors as Therapeutic Targets of Inflammatory Diseases. Int. Immunol. 2019, 31, 607–615. [Google Scholar] [CrossRef]
- Lämmermann, T.; Afonso, P.V.; Angermann, B.R.; Wang, J.M.; Kastenmüller, W.; Parent, C.A.; Germain, R.N. Neutrophil Swarms Require LTB4 and Integrins at Sites of Cell Death in Vivo. Nature 2013, 498, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jeldres, T.; Tyler, C.J.; Boyer, J.D.; Karuppuchamy, T.; Bamias, G.; Dulai, P.S.; Boland, B.S.; Sandborn, W.J.; Patel, D.R.; Rivera-Nieves, J. Cell Trafficking Interference in Inflammatory Bowel Disease: Therapeutic Interventions Based on Basic Pathogenesis Concepts. Inflamm. Bowel Dis. 2019, 25, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Gabrielsen, A.; Agardh, H.E.; Wan, M.; Wetterholm, A.; Wong, C.-H.; Hedin, U.; Swedenborg, J.; Hansson, G.K.; Samuelsson, B.; et al. Expression of 5-Lipoxygenase and Leukotriene A4 Hydrolase in Human Atherosclerotic Lesions Correlates with Symptoms of Plaque Instability. Proc. Natl. Acad. Sci. USA 2006, 103, 8161–8166. [Google Scholar] [CrossRef] [Green Version]
- Spanbroek, R.; Grabner, R.; Lotzer, K.; Hildner, M.; Urbach, A.; Ruhling, K.; Moos, M.P.W.; Kaiser, B.; Cohnert, T.U.; Wahlers, T.; et al. Expanding Expression of the 5-Lipoxygenase Pathway within the Arterial Wall during Human Atherogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 1238–1243. [Google Scholar] [CrossRef] [Green Version]
- Busse, null Leukotrienes and Inflammation. Am. J. Respir. Crit. Care Med. 1998, 157, S210–S213. [CrossRef] [PubMed]
- Hui, Y.; Cheng, Y.; Smalera, I.; Jian, W.; Goldhahn, L.; Fitzgerald, G.A.; Funk, C.D. Directed Vascular Expression of Human Cysteinyl Leukotriene 2 Receptor Modulates Endothelial Permeability and Systemic Blood Pressure. Circulation 2004, 110, 3360–3366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moos, M.P.W.; Mewburn, J.D.; Kan, F.W.K.; Ishii, S.; Abe, M.; Sakimura, K.; Noguchi, K.; Shimizu, T.; Funk, C.D. Cysteinyl Leukotriene 2 Receptor-Mediated Vascular Permeability via Transendothelial Vesicle Transport. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 4352–4362. [Google Scholar] [CrossRef]
- Samuelsson, B. Leukotrienes: Mediators of Immediate Hypersensitivity Reactions and Inflammation. Science 1983, 220, 568–575. [Google Scholar] [CrossRef]
- Erlemann, K.-R.; Rokach, J.; Powell, W.S. Oxidative Stress Stimulates the Synthesis of the Eosinophil Chemoattractant 5-Oxo-6,8,11,14-Eicosatetraenoic Acid by Inflammatory Cells. J. Biol. Chem. 2004, 279, 40376–40384. [Google Scholar] [CrossRef] [Green Version]
- Grant, G.E.; Rokach, J.; Powell, W.S. 5-Oxo-ETE and the OXE Receptor. Prostaglandins Other Lipid Mediat. 2009, 89, 98–104. [Google Scholar] [CrossRef]
- Powell, W.S.; Gravel, S.; MacLeod, R.J.; Mills, E.; Hashefi, M. Stimulation of Human Neutrophils by 5-Oxo-6,8,11,14-Eicosatetraenoic Acid by a Mechanism Independent of the Leukotriene B4 Receptor. J. Biol. Chem. 1993, 268, 9280–9286. [Google Scholar] [CrossRef]
- Boer, R.E.; Giménez-Bastida, J.A.; Boutaud, O.; Jana, S.; Schneider, C.; Sulikowski, G.A. Total Synthesis and Biological Activity of the Arachidonic Acid Metabolite Hemiketal E2. Org. Lett. 2018, 20, 4020–4022. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Suzuki, T.; Sprinkel, K.C.; Boeglin, W.E.; Schneider, C. Biomimetic Synthesis of Hemiketal Eicosanoids for Biological Testing. Prostaglandins Other Lipid Mediat. 2017, 132, 41–46. [Google Scholar] [CrossRef]
- Chicco, F.; Magrì, S.; Cingolani, A.; Paduano, D.; Pesenti, M.; Zara, F.; Tumbarello, F.; Urru, E.; Melis, A.; Casula, L.; et al. Multidimensional Impact of Mediterranean Diet on IBD Patients. Inflamm. Bowel Dis. 2021, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Marcos, L. Mediterranean Diet as a Protection against Asthma: Still Another Brick in Building a Causative Association. Allergol. Immunopathol. (Madr.) 2016, 44, 97–98. [Google Scholar] [CrossRef] [PubMed]
- Temple, N.J.; Guercio, V.; Tavani, A. The Mediterranean Diet and Cardiovascular Disease: Gaps in the Evidence and Research Challenges. Cardiol. Rev. 2019, 27, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Oz, H.S. Chronic Inflammatory Diseases and Green Tea Polyphenols. Nutrients 2017, 9, 660. [Google Scholar] [CrossRef]
- Widmer, R.J.; Freund, M.A.; Flammer, A.J.; Sexton, J.; Lennon, R.; Romani, A.; Mulinacci, N.; Vinceri, F.F.; Lerman, L.O.; Lerman, A. Beneficial Effects of Polyphenol-Rich Olive Oil in Patients with Early Atherosclerosis. Eur. J. Nutr. 2013, 52, 1223–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, I.; Biswas, S.K.; Kirkham, P.A. Regulation of Inflammation and Redox Signaling by Dietary Polyphenols. Biochem. Pharmacol. 2006, 72, 1439–1452. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Martín, A.; Selma, M.V.; Tomás-Barberán, F.A.; González-Sarrías, A.; Espín, J.C. Where to Look into the Puzzle of Polyphenols and Health? The Postbiotics and Gut Microbiota Associated with Human Metabotypes. Mol. Nutr. Food Res. 2020, 64, e1900952. [Google Scholar] [CrossRef]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The Gut Microbiota: A Key Factor in the Therapeutic Effects of (Poly)Phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Frankenfeld, C.L. Cardiometabolic Risk and Gut Microbial Phytoestrogen Metabolite Phenotypes. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Olthof, M.R.; Hollman, P.C.H.; Buijsman, M.N.C.P.; van Amelsvoort, J.M.M.; Katan, M.B. Chlorogenic Acid, Quercetin-3-Rutinoside and Black Tea Phenols Are Extensively Metabolized in Humans. J. Nutr. 2003, 133, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Dei Cas, M.; Ghidoni, R. Dietary Curcumin: Correlation between Bioavailability and Health Potential. Nutrients 2019, 11, 2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giménez-Bastida, J.A.; Martínez-Florensa, M.; Espín, J.-C.; Tomás-Barberán, F.A.; García-Conesa, M.-T. A Citrus Extract Containing Flavanones Represses Plasminogen Activator Inhibitor-1 (PAI-1) Expression and Regulates Multiple Inflammatory, Tissue Repair, and Fibrosis Genes in Human Colon Fibroblasts. J. Agric. Food Chem. 2009, 57, 9305–9315. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.Á.; González-Sarrías, A.; Martínez-Díaz, F.; Abellán, B.; Martínez-Torrano, A.J.; Fernández-López, A.J.; Giménez-Bastida, J.A.; Espín, J.C. Disposition of Dietary Polyphenols in Breast Cancer Patients’ Tumors, and Their Associated Anticancer Activity: The Particular Case of Curcumin. Mol. Nutr. Food Res. 2021, e2100163. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.Á.; Giménez-Bastida, J.A.; González-Sarrías, A.; Espín, J.C. New Insights into the Metabolism of the Flavanones Eriocitrin and Hesperidin: A Comparative Human Pharmacokinetic Study. Antioxid. Basel Switz. 2021, 10, 435. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Espín, J.C.; Tomás-Barberán, F.A. Non-Extractable Polyphenols Produce Gut Microbiota Metabolites That Persist in Circulation and Show Anti-Inflammatory and Free Radical-Scavenging Effects. Trends Food Sci. Technol. 2017, 69, 281–288. [Google Scholar] [CrossRef]
- Erlund, I.; Kosonen, T.; Alfthan, G.; Mäenpää, J.; Perttunen, K.; Kenraali, J.; Parantainen, J.; Aro, A. Pharmacokinetics of Quercetin from Quercetin Aglycone and Rutin in Healthy Volunteers. Eur. J. Clin. Pharmacol. 2000, 56, 545–553. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Giménez-Bastida, J.A.; García-Conesa, M.T.; Gómez-Sánchez, M.B.; García-Talavera, N.V.; Gil-Izquierdo, A.; Sánchez-Alvarez, C.; Fontana-Compiano, L.O.; Morga-Egea, J.P.; Pastor-Quirante, F.A.; et al. Occurrence of Urolithins, Gut Microbiota Ellagic Acid Metabolites and Proliferation Markers Expression Response in the Human Prostate Gland upon Consumption of Walnuts and Pomegranate Juice. Mol. Nutr. Food Res. 2010, 54, 311–322. [Google Scholar] [CrossRef]
- Kunihiro, A.G.; Brickey, J.A.; Frye, J.B.; Luis, P.B.; Schneider, C.; Funk, J.L. Curcumin, but Not Curcumin-Glucuronide, Inhibits Smad Signaling in TGFβ-Dependent Bone Metastatic Breast Cancer Cells and Is Enriched in Bone Compared to Other Tissues. J. Nutr. Biochem. 2019, 63, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Shimoi, K.; Saka, N.; Nozawa, R.; Sato, M.; Amano, I.; Nakayama, T.; Kinae, N. Deglucuronidation of a Flavonoid, Luteolin Monoglucuronide, during Inflammation. Drug Metab. Dispos. Biol. Fate Chem. 2001, 29, 1521–1524. [Google Scholar] [PubMed]
- Shimoi, K.; Saka, N.; Kaji, K.; Nozawa, R.; Kinae, N. Metabolic Fate of Luteolin and Its Functional Activity at Focal Site. BioFactors Oxf. Engl. 2000, 12, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Galindo, P.; Rodriguez-Gómez, I.; González-Manzano, S.; Dueñas, M.; Jiménez, R.; Menéndez, C.; Vargas, F.; Tamargo, J.; Santos-Buelga, C.; Pérez-Vizcaíno, F.; et al. Glucuronidated Quercetin Lowers Blood Pressure in Spontaneously Hypertensive Rats via Deconjugation. PLoS ONE 2012, 7, e32673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishisaka, A.; Kawabata, K.; Miki, S.; Shiba, Y.; Minekawa, S.; Nishikawa, T.; Mukai, R.; Terao, J.; Kawai, Y. Mitochondrial Dysfunction Leads to Deconjugation of Quercetin Glucuronides in Inflammatory Macrophages. PLoS ONE 2013, 8, e80843. [Google Scholar] [CrossRef] [Green Version]
- Kawai, Y.; Nishikawa, T.; Shiba, Y.; Saito, S.; Murota, K.; Shibata, N.; Kobayashi, M.; Kanayama, M.; Uchida, K.; Terao, J. Macrophage as a Target of Quercetin Glucuronides in Human Atherosclerotic Arteries: Implication in the Anti-Atherosclerotic Mechanism of Dietary Flavonoids. J. Biol. Chem. 2008, 283, 9424–9434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menendez, C.; Dueñas, M.; Galindo, P.; González-Manzano, S.; Jimenez, R.; Moreno, L.; Zarzuelo, M.J.; Rodríguez-Gómez, I.; Duarte, J.; Santos-Buelga, C.; et al. Vascular Deconjugation of Quercetin Glucuronide: The Flavonoid Paradox Revealed? Mol. Nutr. Food Res. 2011, 55, 1780–1790. [Google Scholar] [CrossRef]
- Terao, J.; Murota, K.; Kawai, Y. Conjugated Quercetin Glucuronides as Bioactive Metabolites and Precursors of Aglycone in Vivo. Food Funct. 2011, 2, 11–17. [Google Scholar] [CrossRef]
- Fernández-Castillejo, S.; Macià, A.; Motilva, M.-J.; Catalán, Ú.; Solà, R. Endothelial Cells Deconjugate Resveratrol Metabolites to Free Resveratrol: A Possible Role in Tissue Factor Modulation. Mol. Nutr. Food Res. 2019, 63, e1800715. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.A.; Giménez-Bastida, J.A.; González-Sarrías, A.; Espín, J.C. Tissue Deconjugation of Urolithin A Glucuronide to Free Urolithin A in Systemic Inflammation. Food Funct. 2019, 10, 3135–3141. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. PLoS Med. 2021, 18, e1003583. [Google Scholar] [CrossRef] [PubMed]
- Ho, X.L.; Liu, J.J.H.; Loke, W.M. Plant Sterol-Enriched Soy Milk Consumption Modulates 5-Lipoxygenase, 12-Lipoxygenase, and Myeloperoxidase Activities in Healthy Adults – a Randomized-Controlled Trial. Free Radic. Res. 2016, 50, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Schramm, D.D.; Wang, J.F.; Holt, R.R.; Ensunsa, J.L.; Gonsalves, J.L.; Lazarus, S.A.; Schmitz, H.H.; German, J.B.; Keen, C.L. Chocolate Procyanidins Decrease the Leukotriene-Prostacyclin Ratio in Humans and Human Aortic Endothelial Cells. Am. J. Clin. Nutr. 2001, 73, 36–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalhan, R.; Smith, L.J.; Nlend, M.C.; Nair, A.; Hixon, J.L.; Sporn, P.H.S. A Mechanism of Benefit of Soy Genistein in Asthma: Inhibition of Eosinophil P38-Dependent Leukotriene Synthesis. Clin. Exp. Allergy 2007, 38, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidou, V.; Khymenets, O.; Covas, M.-I.; de la Torre, R.; Muñoz-Aguayo, D.; Anglada, R.; Farré, M.; Fito, M. Time Course of Changes in the Expression of Insulin Sensitivity-Related Genes after an Acute Load of Virgin Olive Oil. OMICS J. Integr. Biol. 2009, 13, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Capó, X.; Martorell, M.; Sureda, A.; Riera, J.; Drobnic, F.; Tur, J.A.; Pons, A. Effects of Almond- and Olive Oil-Based Docosahexaenoic- and Vitamin E-Enriched Beverage Dietary Supplementation on Inflammation Associated to Exercise and Age. Nutrients 2016, 8, 619. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zhang, J.; Zhang, Y.; Meng, D.; Yan, D. Anti-Inflammatory Activity Studies on the Stems and Roots of Jasminum Lanceolarium Roxb. J. Ethnopharmacol. 2015, 171, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, R.; Fan, A.; Zhong, G.; He, J. Dendropanax Dentiger (Harms) Merr. Root and Its Major Constituents Exert Therapeutic Effect on Adjuvant-Induced Arthritis in Rats. J. Ethnopharmacol. 2021, 267, 113631. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, R.; Fan, A.; Zhao, J.; Zhang, Y.; He, J. Chemical Composition of Pterospermum Heterophyllum Root and Its Anti-Arthritis Effect on Adjuvant-Induced Arthritis in Rats via Modulation of Inflammatory Responses. Front. Pharmacol. 2020, 11, 584849. [Google Scholar] [CrossRef]
- Zheng, C.-J.; Zhao, X.-X.; Ai, H.-W.; Lin, B.; Han, T.; Jiang, Y.-P.; Xing, X.; Qin, L.-P. Therapeutic Effects of Standardized Vitex Negundo Seeds Extract on Complete Freund’s Adjuvant Induced Arthritis in Rats. Phytomed. Int. J. Phytother. Phytopharm. 2014, 21, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, Y.; Jin, Y.; Son, J.-K.; Lee, S.H.; Chang, H.W. Curcumin Inhibits the Activation of Immunoglobulin E-Mediated Mast Cells and Passive Systemic Anaphylaxis in Mice by Reducing Serum Eicosanoid and Histamine Levels. Biomol. Ther. 2014, 22, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjunatha, H.; Srinivasan, K. Protective Effect of Dietary Curcumin and Capsaicin on Induced Oxidation of Low-Density Lipoprotein, Iron-Induced Hepatotoxicity and Carrageenan-Induced Inflammation in Experimental Rats. FEBS J. 2006, 273, 4528–4537. [Google Scholar] [CrossRef]
- Xia, H.; He, Q.; Wang, H.; Wang, Y.; Yang, Y.; Li, Y.; Zhang, J.; Chen, Z.; Yang, J. Treatment with Either COX-2 Inhibitor or 5-LOX Inhibitor Causes No Compensation between COX-2 Pathway and 5-LOX Pathway in Chronic Aluminum Overload-induced Liver Injury in Rats. Fundam. Clin. Pharmacol. 2019, 33, 535–543. [Google Scholar] [CrossRef]
- Yang, J.-Q.; Zhou, Q.-X.; Liu, B.-Z.; He, B.-C. Protection of Mouse Brain from Aluminum-Induced Damage by Caffeic Acid. CNS Drug Rev. 2008, 14, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Pallio, G.; Bitto, A.; Pizzino, G.; Galfo, F.; Irrera, N.; Minutoli, L.; Arcoraci, V.; Squadrito, G.; Macrì, A.; Squadrito, F.; et al. Use of a Balanced Dual Cyclooxygenase-1/2 and 5-Lypoxygenase Inhibitor in Experimental Colitis. Eur. J. Pharmacol. 2016, 789, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.J.; McCormick, M.E.; Howarth, A.; Aked, D. Leukocyte Recruitment in the Subcutaneous Sponge Implant Model of Acute Inflammation in the Rat Is Not Mediated by Leukotriene B1. Biochem. Pharmacol. 1986, 35, 1709–1717. [Google Scholar] [CrossRef]
- Peskar, B.M.; Lange, K.; Hoppe, U.; Peskar, B.A. Ethanol Stimulates Formation of Leukotriene C4 in Rat Gastric Mucosa. Prostaglandins 1986, 31, 283–293. [Google Scholar] [CrossRef]
- Choi, J.-H.; Chai, Y.-M.; Joo, G.-J.; Rhee, I.-K.; Lee, I.-S.; Kim, K.-R.; Choi, M.-S.; Rhee, S.-J. Effects of Green Tea Catechin on Polymorphonuclear Leukocyte 5′-Lipoxygenase Activity, Leukotriene B4 Synthesis, and Renal Damage in Diabetic Rats. Ann. Nutr. Metab. 2004, 48, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Chang, H.-W.; Rhee, S.-J. Effect of Green Tea Catechin on Arachidonic Acid Cascade in Chronic Cadmium-Poisoned Rats. Asia Pac. J. Clin. Nutr. 2002, 11, 292–297. [Google Scholar] [CrossRef]
- Viji, V.; Helen, A. Inhibition of Lipoxygenases and Cyclooxygenase-2 Enzymes by Extracts Isolated from Bacopa Monniera (L.) Wettst. J. Ethnopharmacol. 2008, 118, 305–311. [Google Scholar] [CrossRef]
- Bhaskar, S.; Kumar, K.S.; Krishnan, K.; Antony, H. Quercetin Alleviates Hypercholesterolemic Diet Induced Inflammation during Progression and Regression of Atherosclerosis in Rabbits. Nutrition 2013, 29, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Nisar, A.; Malik, A.H.; Zargar, M.A. Atropa Acuminata Royle Ex Lindl. Blunts Production of pro-Inflammatory Mediators Eicosanoids., Leukotrienes, Cytokines in Vitro and in Vivo Models of Acute Inflammatory Responses. J. Ethnopharmacol. 2013, 147, 584–594. [Google Scholar] [CrossRef]
- Lin, B.; Zhao, Y.; Han, P.; Yue, W.; Ma, X.-Q.; Rahman, K.; Zheng, C.-J.; Qin, L.-P.; Han, T. Anti-Arthritic Activity of Xanthium Strumarium L. Extract on Complete Freund’s Adjuvant Induced Arthritis in Rats. J. Ethnopharmacol. 2014, 155, 248–255. [Google Scholar] [CrossRef]
- Kavitha, S.; John, F.; Indira, M. Amelioration of Inflammation by Phenolic Rich Methanolic Extract of Ocimum Sanctum Linn. Leaves in Isoproterenol Induced Myocardial Infarction. Indian J. Exp. Biol. 2015, 53, 632–640. [Google Scholar]
- Chang, X.; Luo, F.; Jiang, W.; Zhu, L.; Gao, J.; He, H.; Wei, T.; Gong, S.; Yan, T. Protective Activity of Salidroside against Ethanol-Induced Gastric Ulcer via the MAPK/NF-ΚB Pathway in Vivo and in Vitro. Int. Immunopharmacol. 2015, 28, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, S.; Sudhakaran, P.R.; Helen, A. Quercetin Attenuates Atherosclerotic Inflammation and Adhesion Molecule Expression by Modulating TLR-NF-ΚB Signaling Pathway. Cell. Immunol. 2016, 310, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Yashaswini, P.S.; Sadashivaiah, B.; Ramaprasad, T.R.; Singh, S.A. In Vivo Modulation of LPS Induced Leukotrienes Generation and Oxidative Stress by Sesame Lignans. J. Nutr. Biochem. 2017, 41, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tang, H.; Liu, X.; Chen, H.; Feng, N.; Zhang, J.; Wang, C.; Qiu, M.; Yang, J.; Zhou, X. Frontline Science: Reprogramming COX-2, 5-LOX, and CYP4A-mediated Arachidonic Acid Metabolism in Macrophages by Salidroside Alleviates Gouty Arthritis. J. Leukoc. Biol. 2019, 105, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Sobeh, M.; Mahmoud, M.F.; Rezq, S.; Alsemeh, A.E.; Sabry, O.M.; Mostafa, I.; Abdelfattah, M.A.; Ait El-Allem, K.; El-Shazly, A.M.; Yasri, A.; et al. Salix Tetrasperma Roxb. Extract Alleviates Neuropathic Pain in Rats via Modulation of the NF-ΚB/TNF-α/NOX/INOS Pathway. Antioxidants 2019, 8, 482. [Google Scholar] [CrossRef] [Green Version]
- Yabas, M.; Orhan, C.; Er, B.; Tuzcu, M.; Durmus, A.S.; Ozercan, I.H.; Sahin, N.; Bhanuse, P.; Morde, A.A.; Padigaru, M.; et al. A Next Generation Formulation of Curcumin Ameliorates Experimentally Induced Osteoarthritis in Rats via Regulation of Inflammatory Mediators. Front. Immunol. 2021, 12, 609629. [Google Scholar] [CrossRef]
- Beetens, J.R.; Loots, W.; Somers, Y.; Coene, M.C.; De Clerck, F. Ketoconazole Inhibits the Biosynthesis of Leukotrienes in Vitro and in Vivo. Biochem. Pharmacol. 1986, 35, 883–891. [Google Scholar] [CrossRef]
- Billah, M.M.; Bryant, R.W.; Siegel, M.I. Lipoxygenase Products of Arachidonic Acid Modulate Biosynthesis of Platelet-Activating Factor (1-O-Alkyl-2-Acetyl-Sn-Glycero-3-Phosphocholine) by Human Neutrophils via Phospholipase A2. J. Biol. Chem. 1985, 260, 6899–6906. [Google Scholar] [CrossRef]
- Bossù, E.; Aglianò, A.M.; Desideri, N.; Sestili, I.; Porrà, R.; Grandilone, M.; Quaglia, M.G. LTB4 as Marker of 5-LO Inhibitory Activity of Two New N-Omega-Ethoxycarbonyl-4-Quinolones. J. Pharm. Biomed. Anal. 1999, 19, 539–548. [Google Scholar] [CrossRef]
- Bremm, K.D.; Konig, W.; Alouf, J.E. Effect of Thiol-Activated Toxins (Streptolysin 0, Alveolysin, and Theta Toxin) on the Generation of Leukotrienes and Leukotriene- Inducing and -Metabolizing Enzymes from Human Polymorphonuclear Granulocytes. Infect. Immun. 1985, 50, 8. [Google Scholar] [CrossRef] [Green Version]
- Conti, P.; Panara, M.R.; Barbacane, R.C.; Bongrazio, M.; Dempsey, R.A.; Reale, M. Human Recombinant IL-1 Receptor Antagonist (IL-1Ra) Inhibits Leukotriene B4 Generation from Human Monocyte Suspensions Stimulated by Lipopolysaccharide (LPS). Clin. Exp. Immunol. 1993, 91, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Kubo, K.; Shinozaki, S.; Okada, K.; Matsuzawa, Y.; Kobayashi, T.; Sugane, K. Neutrophil Elastase Inhibitor Reduces Asthmatic Responses in Allergic Sheep. Respir. Physiol. 1995, 100, 91–100. [Google Scholar] [CrossRef]
- Hulkower, K.I.; Pollock, J.S.; Walsh, R.E.; Huang, R.; Otis, E.R.; Brooks, C.D.; Bell, R.L. Leukotrienes Do Not Regulate Nitric Oxide Production in RAW 264.7 Macrophages. Prostaglandins Leukot. Essent. Fat. Acids 1996, 55, 145–149. [Google Scholar] [CrossRef]
- Igarashi, Y.; Lundgren, J.D.; Shelhamer, J.H.; Kaliner, M.A.; White, M.V. Effects of Inhibitors of Arachidonic Acid Metabolism on Serotonin Release from Rat Basophilic Leukemia Cells. Immunopharmacology 1993, 25, 131–144. [Google Scholar] [CrossRef]
- Ishii, K.; Yakuo, I.; Motoyoshi, S.; Nakagawa, H.; Nakamura, H. Inhibition of Leukotriene Production by N-[4-[4-(Diphenylmethyl)-1- Piperazinyl]Butyl]-3-(6-Methyl-3-Pyridyl) Acrylamide (AL-3264), a New Antiallergic Agent. Jpn. J. Pharmacol. 1994, 65, 19–25. [Google Scholar] [CrossRef]
- Johnson-Henry, K.; Wallace, J.L.; Basappa, N.S.; Soni, R.; Wu, G.K.; Sherman, P.M. Inhibition of Attaching and Effacing Lesion Formation Following Enteropathogenic Escherichia Coli and Shiga Toxin-Producing E. Coli Infection. Infect. Immun. 2001, 69, 7152–7158. [Google Scholar] [CrossRef] [Green Version]
- Malleron, J.L.; Roussel, G.; Gueremy, G.; Ponsinet, G.; Robin, J.L.; Terlain, B.; Tissieres, J.M. Penta- and Hexadienoic Acid Derivatives: A Novel Series of 5-Lipoxygenase Inhibitors. J. Med. Chem. 1990, 33, 2744–2749. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Ziereis, K.; Paper, D.H. Ilex Aquifolium: Protection against Enzymatic and Non-Enzymatic Lipid Peroxidation. Planta Med. 1998, 64, 536–540. [Google Scholar] [CrossRef]
- Ohemeng, K.A.; Nguyen, V.N.; Schwender, C.F.; Singer, M.; Steber, M.; Ansell, J.; Hageman, W. Novel Bishydroxamic Acids as 5-Lipoxygenase Inhibitors. Bioorg. Med. Chem. 1994, 2, 187–193. [Google Scholar] [CrossRef]
- Petroni, A.; Blasevich, M.; Papini, N.; Salami, M.; Sala, A.; Galli, C. Inhibition of Leukocyte Leukotriene B4 Production by an Olive Oil-Derived Phenol Identified by Mass-Spectrometry. Thromb. Res. 1997, 87, 315–322. [Google Scholar] [CrossRef]
- Prasad, N.S.; Raghavendra, R.; Lokesh, B.R.; Naidu, K.A. Spice Phenolics Inhibit Human PMNL 5-Lipoxygenase. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 521–528. [Google Scholar] [CrossRef]
- Rotondo, S.; Rajtar, G.; Manarini, S.; Celardo, A.; Rotilio, D.; De Gaetano, G.; Evangelista, V.; Cerletti, C. Effect of Trans -Resveratrol, a Natural Polyphenolic Compound, on Human Polymorphonuclear Leukocyte Function: Trans -Resveratrol and Human PMN Leukocyte Function. Br. J. Pharmacol. 1998, 123, 1691–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tommasini, I.; Guidarelli, A.; Palomba, L.; Cerioni, L.; Cantoni, O. 5-Hydroxyeicosatetraenoic Acid Is a Key Intermediate of the Arachidonate-Dependent Protective Signaling in Monocytes/Macrophages Exposed to Peroxynitrite. J. Leukoc. Biol. 2006, 80, 929–938. [Google Scholar] [CrossRef]
- Van Hilten, J.A.; Elliott, G.R.; Bonta, I.L. Endogenous Lipoxygenase Metabolites Mediate A23187 Induced Macrophage Cytostasis towards P815 Tumor Cells in Vitro. Agents Actions 1989, 26, 170–172. [Google Scholar] [CrossRef]
- Van Hilten, J.A.; Elliott, G.R.; Bonta, I.L. Specific Lipoxygenase Inhibition Reverses Macrophage Cytotasis towards P815 Tumor Cells in Vitro Induced by the Calcium Ionophore A23187. Prostaglandins Leukot. Essent. Fat. Acids 1988, 34, 187–192. [Google Scholar] [CrossRef]
- Kimura, Y.; Okuda, H.; Okuda, T.; Hatano, T.; Arichi, S. Studies on the Activities of Tannins and Related Compounds, X. Effects of Caffeetannins and Related Compounds on Arachidonate Metabolism in Human Polymorphonuclear Leukocytes. J. Nat. Prod. 1987, 50, 392–399. [Google Scholar] [CrossRef]
- Lin, T.-Y.; London, C.A. Characterization and Modulation of Canine Mast Cell Derived Eicosanoids. Vet. Immunol. Immunopathol. 2010, 135, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsen, S.Y.; Tan, X.Y.; Tan, Y.M.; Yan, B.Y.; Loke, W.M. Relative Inhibitions of 5-Lipoxygenase and Myeloperoxidase and Free-Radical Scavenging Activities of Daidzein, Dihydrodaidzein, and Equol. J. Med. Food 2016, 19, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Ammon, H.P.T.; Safayhi, H.; Mack, T.; Sabieraj, J. Mechanism of Antiinflammatory Actions of Curcumine and Boswellic Acids. J. Ethnopharmacol. 1993, 38, 105–112. [Google Scholar] [CrossRef]
- Hong, J. Modulation of Arachidonic Acid Metabolism by Curcumin and Related -Diketone Derivatives: Effects on Cytosolic Phospholipase A2, Cyclooxygenases and 5-Lipoxygenase. Carcinogenesis 2004, 25, 1671–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.; Couto, D.; Alves, A.; Dias, I.; Freitas, M.; Porto, G.; Duarte, J.A.; Fernandes, E. Trihydroxyflavones with Antioxidant and Anti-Inflammatory Efficacy. BioFactors 2012, 38, 378–386. [Google Scholar] [CrossRef]
- Kwon, O.S.; Choi, J.S.; Islam, M.N.; Kim, Y.S.; Kim, H.P. Inhibition of 5-Lipoxygenase and Skin Inflammation by the Aerial Parts of Artemisia Capillaris and Its Constituents. Arch. Pharm. Res. 2011, 34, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Takasugi, M.; Muta, E.; Yamada, K.; Arai, H. A New Method to Evaluate Anti-Allergic Effect of Food Component by Measuring Leukotriene B4 from a Mouse Mast Cell Line. Cytotechnology 2018, 70, 177–184. [Google Scholar] [CrossRef]
- Dehmlow, C.; Murawski, N.; de Groot, H. Scavenging of Reactive Oxygen Species and Inhibition of Arachidonic Acid Metabolism by Silibinin in Human Cells. Life Sci. 1996, 58, 1591–1600. [Google Scholar] [CrossRef]
- Dehmlow, C.; Erhard, J.; de Groot, H. Inhibition of Kupffer Cell Functions as an Explanation for the Hepatoprotective Properties of Silibinin. Hepatology 1996, 23, 749–754. [Google Scholar] [CrossRef]
- Jupp, J.; Hillier, K.; Elliott, D.H.; Fine, D.R.; Bateman, A.C.; Johnson, P.A.; Cazaly, A.M.; Penrose, J.F.; Sampson, A.P. Colonic Expression of Leukotriene-Pathway Enzymes in Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2007, 13, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Han, S.-U.; Lee, K.-M.; Park, K.H.; Cho, S.W.; Hahm, K.-B. 5-LOX Inhibitor Modulates the Inflammatory Responses Provoked by Helicobacter Pylori Infection. Helicobacter 2007, 12. [Google Scholar] [CrossRef]
- Habib, E.S.; El-Bsoumy, E.; Ibrahim, A.K.; Helal, M.A.; El-Magd, M.A.; Ahmed, S.A. Anti-Inflammatory Effect of Methoxyflavonoids from Chiliadenus Montanus (Jasonia Montana) Growing in Egypt. Nat. Prod. Res. 2020, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Loke, W.M.; Proudfoot, J.M.; Stewart, S.; McKinley, A.J.; Needs, P.W.; Kroon, P.A.; Hodgson, J.M.; Croft, K.D. Metabolic Transformation Has a Profound Effect on Anti-Inflammatory Activity of Flavonoids Such as Quercetin: Lack of Association between Antioxidant and Lipoxygenase Inhibitory Activity. Biochem. Pharmacol. 2008, 75, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Bastida, J.A.; González-Sarrías, A.; Espín, J.C.; Schneider, C. Inhibition of 5-Lipoxygenase-Derived Leukotrienes and Hemiketals as a Novel Anti-Inflammatory Mechanism of Urolithins. Mol. Nutr. Food Res. 2020, e2000129. [Google Scholar] [CrossRef]
- Thivierge, M.; Stankova, J.; Rola-Pleszczynski, M. Toll-like Receptor Agonists Differentially Regulate Cysteinyl-Leukotriene Receptor 1 Expression and Function in Human Dendritic Cells. J. Allergy Clin. Immunol. 2006, 117, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Gálvez, M.Á.; González-Sarrías, A.; Espín, J.C. In Vitro Research on Dietary Polyphenols and Health: A Call of Caution and a Guide on How To Proceed. J. Agric. Food Chem. 2018, 66, 7857–7858. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population of Study | Design of the Study | Foodstuff; Intake and Duration | Main Outcomes | Ref. |
|---|---|---|---|---|
| Healthy volunteers (n = 10; 20–55 years) | Randomized, crossover, double-blind, placebo-controlled. | Low- (0.09 mg/g) and high-procyanidin (4.0 mg/g) chocolate bars; 37 g (single dose); Duration: blood collection at 2 and 6 h; wash-out period of 1 week between treatments. | ↑Epicatechin plasma level (especially the high procyanidins consumer group). ↓CysLTs/PGI2 ratio (relative to the effects observed in the low procyanidins consumer group). | [94] |
| Patients suffering mild/moderate persistent asthma (n = 13; 18–65 years) | Quasi-experimental intervention study. No control group. | 1 Soy isoflavones tablets (NOVASOY, ArcherDanielsMidland, Decatur, IL, USA); two 50 mg tablets (once per day); duration: 4 weeks. | ↓LTB4 and FENO in A23187-stimulated eosinophils (ex vivo); no significant changes in pre-bronchodilator FEV1 and Juniper Asthma Control Score. | [95] |
| Healthy volunteers (n = 18; 22–44 years) | Acute consumption, no control group. | Raw virgin olive oil; 50 mL (single dose consumed with 200 g bread); Duration: blood collection at 1 and 6 h; 1 week wash-out period before olive oil consumption. | ↑Hydroxytyrosol in plasma. ↓ALOX5AP gene expression at 1 h (basal values at 6 h). The downregulation was inversely correlated with glucose and insulin levels. | [96] |
| Healthy volunteers of Asian ethnicities (n = 18) | Randomized, crossover, double-blind, placebo-controlled. | 2 Soy milk; 2 daily treatments (20 g); Duration: 3 h after acute consumption followed by 1-week wash-out period and 4-weeks daily exposure. | ↓LTB4 and LXA4 level in plasma after 3 h; ↓LTB4 and F2-isoprostanes in plasma and urine after 4-weeks daily exposure; ↑LXA4 in plasma after 4-weeks daily exposure; ↓MPO activity, serum lipid hydroperoxides and hsCRP in plasma after 4-weeks daily exposure. | [93] |
| Senior and young taekwondo athletes (n = 10; 18–57 years old) | Quasi-experimental intervention study, no placebo. | Isotonic beverage enriched with almond (0.3%), sucrose (0.8%), oils (0.2% 3 DHA-S and 0.6% olive oil), and α-tocopherol acetate (vitamin E); duration: 5 weeks (consumed 5 days a week); blood samples taken at the beginning and at the final of the (1 h before and after each stress test). | Beverage supplementation exerted ↓sL-Selectin, sICAM3 and ↑IL-6 in young athletes (after exercise) as well as ↑TNF-α level in plasma in the young group (in absence of exercise) and in the senior group (after exercise). The beverage consumption exerted no significant effects on lipoxin, PGE2, PGE1, and NF-κB. Modulation of TNF-α, 15-LOX2, COX-2, IL-1β and IL-8 mRNA expression in PBMC; No Effect on TLR4, NF-κB, 5-LOX, IL-10, IL-15, HSP72 expression (mRNA) in PBMC. | [97] |
| Animal Model | Extract/Compound Assayed | Dose/Duration | Main Outcomes | Ref. |
|---|---|---|---|---|
| ♂ Wistar rats; carrageenin-soaked sponges implanted subcutaneously. | Quercetin and NDGA. | 100 mg/kg; administered (16 and) 1 h prior sponge implantation. | No effect on leukocyte infiltration, oedema formation or PGE2 and LTB4 formation in A23187-stimulated leukocytes ex vivo. | [107] |
| ♂ Wistar rats; ethanol-induced gastric mucosal damage. | NDGA. | 100 mg/kg (prepared in 0.25% carboxymethylcellulose); administration of a single dose for 30 min. | ↓Gastric lesions and LTC4 biosynthesis; no changes on PGs and TxB2 production. | [108] |
| ♂ Sprague Dawley rats; cadmium-poisoned rats. | Crude catechin powder. 1 | 0.25 and 0.5 g powder/100 g diet; 4 and 20 weeks. | Assays in platelets (ex vivo): ↓PLA2 and COX-1 activity, ↓TxB2; assays in aortic slices: ↓6-keto-PGF1α and ↓LTB4 in A23187-stimulated leukocytes; ↓5-LOX activity (enzyme fraction level). | [109,110] |
| ♂ Wistar rats; carrageenan-induced paw inflammation. | Curcumin, capsaicin, and a mix of curcumin/capsaicin. | Curcumin: 0.2%, capsaicin: 0.015%, curcumin/capsaicin: 0.2/0.015%; 10 weeks. | ↓Volume of paw edema; ↓5-LOX activity in the enzyme obtained from PMNL isolated from blood of the rats; ↓histamine release. | [103] |
| ♂ KM strain mice; aluminum-induced brain damage. | Caffeic acid. | 10 or 30 mg/kg; days. | ↓5-LOX mRNA expression in the cortical brain (at 10 and 30 mg/kg) and protein expression in hippocampi (only at 30 mg/kg); improvement of memory and learning functions together with ↓MDA, ↓ChAT, and ↓amyloid β and amyloid precursor protein. | [105] |
| ♂ Wistar rats; carrageenan-induced paw edema. | Bacopa monniera extracts. | 20–200 mg/kg; 3 and 5 h. | ↓Volume of paw edema; ↓LPS-induced TNF-α release in whole blood (ex vivo); ↓5-LOX and 15-LOX (IC50 = 100 μg/mL) as well as COX-1 (IC50 = 15.66 μg/mL) and COX-2 (IC50 = 1.22 μg) in A23187-induced rat mononuclear cells (ex vivo). | [111] |
| ♀ New Zealand white rabbits; hypercholesterolemic diet. | Quercetin. | 25 mg/kg; 90 days. | ↓5-LOX, 12-LOX, COX, activity in rabbit mononuclear cells; ↓CRP in plasma, ↓MPO activity in the aorta, and improvement of lipid profile and histopathological aortic features. | [112] |
| ♂ Wistar albino rats; carrageenan-induced paw edema. | Atropa acuminata ethanolic extract. | 62.5–500 mg/kg b.w.; up to 4 h. | ↓LTB4 and PGE2 in carrageenan-treated paws; ↓leukocyte and neutrophil recruitment (no effect on mononuclear cells); ↓vascular permeability; ↓paw edema and exudate volume; modulation of the antioxidant status. | [113] |
| ICR mice; IgE/Ag-mediated passive systemic anaphylaxis. | Curcumin. | 20 and 50 mg/kg; 1 h. | ↓LTC4, PGD2 and histamine. | [102] |
| ♂ Wistar rats; CFA-induced rheumatoid arthritis. | Xanthium strumarium extract. | 75 and 300 mg/kg; administered twice a day after the adjuvant arthritis induction for 28 days. | ↓5-LOX and COX-2 expression in PBMCs; ↓paw swelling and arthritic score; ↓TNF-α and IL-1β together with ↑IL-10 in serum; improvement of histopathological features. | [114] |
| ♂ Wistar albino rats; CFA-induced arthritis. | Vitex negundo seed extract. | 85 and 340 mg/kg/day; 28 days. | ↓Paw swelling (from day 14th) and clinical arthritis score; attenuation of CFA-induced weight loss and index of spleen; ↓synovial lining hyperplasia and massive infiltration of mononuclear cells; ↓TNF-α, IL-1β (at both concentrations) and IL-6 (at 340 mg/kg); ↑IL-10; ↓COX-2 and 5-LOX expression in isolated PBMC. | [101] |
| ♂ Sprague Dawley albino rats; Isoproterenol-induced myocardial infarction. | Ocimum sanctum methanolic extract. | 50–250 mg/kg b.w.; 30 days. | ↓TBARS and NF-κB expression in the heart; ↓FLAP and BLT1 (mRNA) expression in the heart; ↓PLA, PLC and PLD activity, whereas ↑SOD activity and phospholipids in the heart; ↓CK-MB, LDH, hsCRP, LTB4, TxB2 (in serum); ↓COX-2 and 5-LOX activity in monocytes; attenuation of the effects of isoproterenol on cardiomyocytes. | [115] |
| ♂ BALB/c mice; ethanol-induced gastric ulcer. | Salidroside. | 20 and 40 mg/kg; 6 days. | ↓5-LOX and COX-2 protein expression; ↓LTB4 level; modulation of the MAPK and NF-κB pathways; ↓IL-6, IL-1β and TNF-α; improvement of gastric histopathological features. | [116] |
| ♂ Wistar rats; carrageenan-induced rat paw edema. | Jasminum laceolarium. | 100–400 mg/kg; 7 days. | ↓5-LOX (only at 400 mg/kg) and COX-2 expression in serum; ↓paw edema. | [98] |
| ♀ Sprague Dawley albino rats; HCD-fed atherosclerotic rats. | Quercetin. | 25 mg/kg b.w.; 60 days. | ↓5-LOX and COX activity as well as IL-6 expression (mRNA) in mononuclear cells; ↓NOS activity and CRP in serum; ↓MDA in serum and aorta. | [117] |
| ♂ Sprague Dawley rats; DNBS- and DSS-induced colitis. | Flavocoxid. | 20 mg/kg/day (twice a day); 4–5 days. | ↓LTB4, PGE2, 6-keto PGF1α, TxB2, and TNF-α serum level; ↓MPO activity and MDA level in colon tissue; ↓histological damage and apoptosis; ↓CD3 in colon tissue. | [106] |
| ♂ Wistar rats; LPS-induced inflammation. | Sesamol, sesamin, and a mix of sesamol/sesamin. | 10 mg/kg b.w.; 15 days. | ↓LTB4, LTC4, MCP-1, IL-1β, CRP, and TNF-α serum level; ↓5-LOX, cPLA2, and BLT-1 protein expression; ↓LTC4 synthase protein expression (only sesamol and sesamin); ↓MDA (liver tissue and serum); ↓NO serum level (only sesamol); modulation of the antioxidant enzymes. | [118] |
| ♂ Wistar rats and New Zealand rabbits; MSU crystal-induced inflammation. | Salidroside. | 40–80 mg/kg for rats and 20–40 mg/kg for rabbits; 6 days. | ↓LTB4, PGE2, and 20-HETE level in synovial fluid macrophages; ↓COX-2, 5-LOX, and CYP4A1 mRNA expression (only at 80 mg/kg) in synovial fluid macrophages; ↓number of leukocytes and neutrophils; binding to the catalytic side of 5-LOX, COX-2, and CYP4A1 (in silico); ↑macrophages polarization; improvement of ankle swelling and histopathological features. | [119] |
| ♂ Wistar rats and Swiss albino mice; carrageenan-inflammation model and acetic acid-induced writhes. | Salix tetrasperma methanolic extract. | 200–600 mg/kg; single dose. | ↓COX-2, 5-LOX, PGE2, TNF-α, iNOS level, and NF-κB activation in sciatic nerve and brain stem; ↓oxidative stress; ↓p53 positive cells in brain stem tissue; ↓paw edema in rats and leukocyte migration in mice; ↓acetic acid-induced writhes; ↑response latency to heat hyperalgesic stimulus; improvement of histopathological features; antipyretic effect. | [120] |
| ♂ Sprague Dawley rats; aluminum gluconate-induced liver injury. | Caffeic acid. | 30 mg/kg. | ↓5-LOX protein expression in the liver (no effect on COX-2); ↓TNF-α, IL-1β, IL-6, MDA and ↑SOD in the liver; improvement of histopathological features; modulation of the alteration of hepatic enzymes; | [104] |
| ♂ Sprague Dawley rats; adjuvant-induced arthritis. | Pterospermun heterophyllum ethanolic extract. | 160–640 mg/kg/day; 22 days. | ↓5-LOX, COX-2, and MMP-2 expression in rat-isolated PBMCs; ↓TNF-α, IL-1β, IL-6, IL-17, RF, and CRP serum level; ↑IL-4 and IL-10 serum level; improvement of histopathological features of the knee joint and arthritis markers. | [100] |
| ♂ Sprague Dawley rats; adjuvant-induced arthritis. | Dendropanax dentiger ethanolic extract. | 127.5–510 mg/kg/day; 22 days. | ↓5-LOX, COX-2, and MMP-2 expression in rat-isolated PBMCs; ↓TNF-α, IL-1β, IL-6, IL-17, RF, and CRP serum level; ↑IL-4 and IL-10 serum level; improvement of histopathological features of the knee joint and arthritis markers. | [99] |
| ♂ Sprague Dawley rats; pharmaco-kinetic study. ♀ Wistar rats; MIA-induced knee OA. | A curcumin formulation (NGUC) or turmeric extract. | NGUC or 95% turmeric extract as 100% (w/v) aqueous solution to deliver 200 mg/kg b.w. equivalent of curcuminoids. | Enhanced total curcuminoids bioavailability in NGUC-treated animals; reduced swelling; improvement of joint architecture; ↓IL-6, IL-1β, TNF-α, CRP, COMP, NF-κB, COX-2, MMP-3, 5-LOX, COX-2 in synovial fluid; ↓MDA, SOD, CAT, and GPx level. | [121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giménez-Bastida, J.A.; González-Sarrías, A.; Laparra-Llopis, J.M.; Schneider, C.; Espín, J.C. Targeting Mammalian 5-Lipoxygenase by Dietary Phenolics as an Anti-Inflammatory Mechanism: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 7937. https://doi.org/10.3390/ijms22157937
Giménez-Bastida JA, González-Sarrías A, Laparra-Llopis JM, Schneider C, Espín JC. Targeting Mammalian 5-Lipoxygenase by Dietary Phenolics as an Anti-Inflammatory Mechanism: A Systematic Review. International Journal of Molecular Sciences. 2021; 22(15):7937. https://doi.org/10.3390/ijms22157937
Chicago/Turabian StyleGiménez-Bastida, Juan Antonio, Antonio González-Sarrías, José Moisés Laparra-Llopis, Claus Schneider, and Juan Carlos Espín. 2021. "Targeting Mammalian 5-Lipoxygenase by Dietary Phenolics as an Anti-Inflammatory Mechanism: A Systematic Review" International Journal of Molecular Sciences 22, no. 15: 7937. https://doi.org/10.3390/ijms22157937